
Formation of the Outer Shell Layer in Pinctada margaritifera: Structural and Biochemical Evidence for a Sequential Development of the Calcite Units

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Major Steps in the Development of the Calcite Prism in the Pinctada Shell
2.1. The Post-Metamorphosis Mineralization
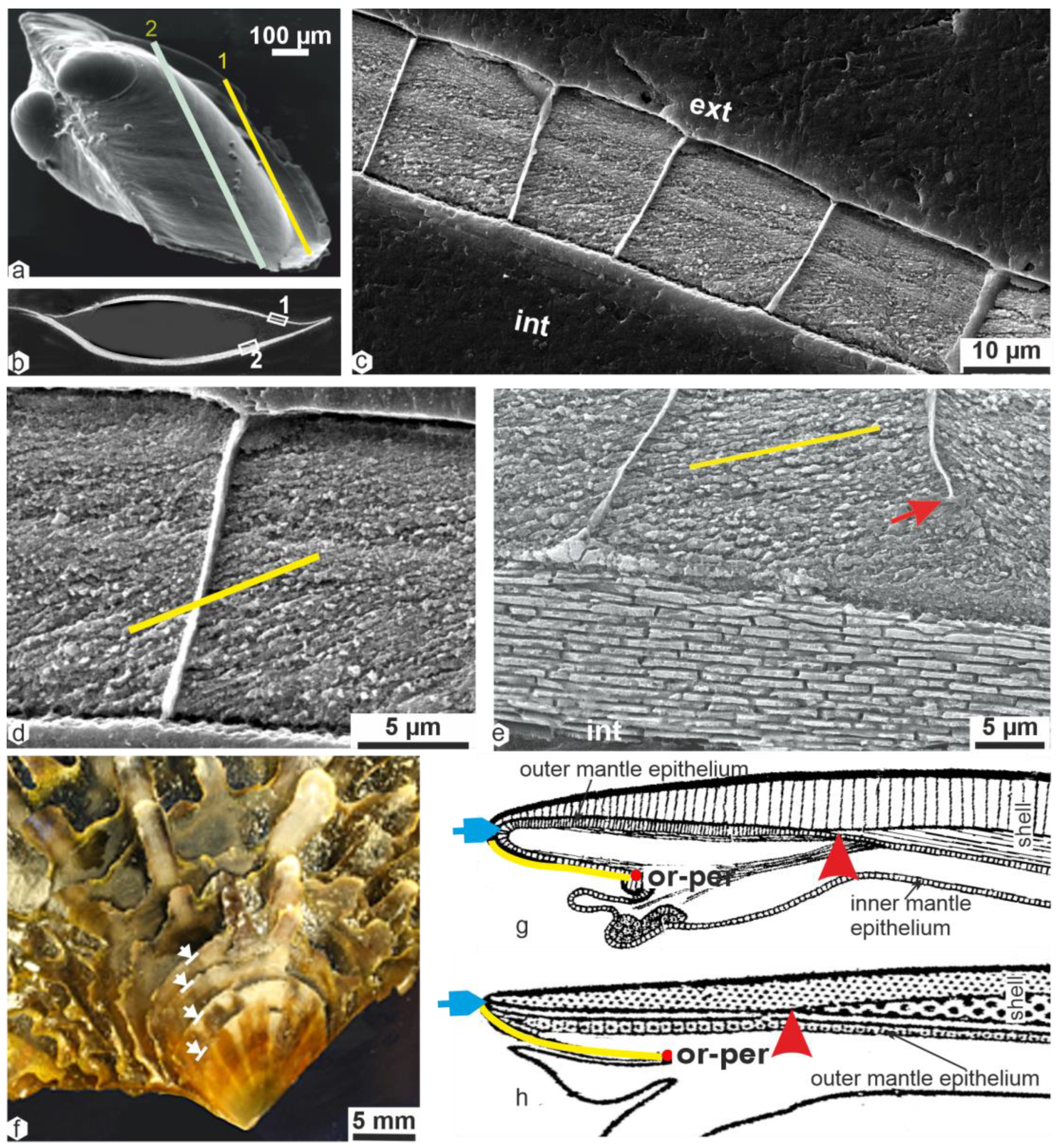
2.2. Microstructural Consequences of the Occurrence of Shell Scales
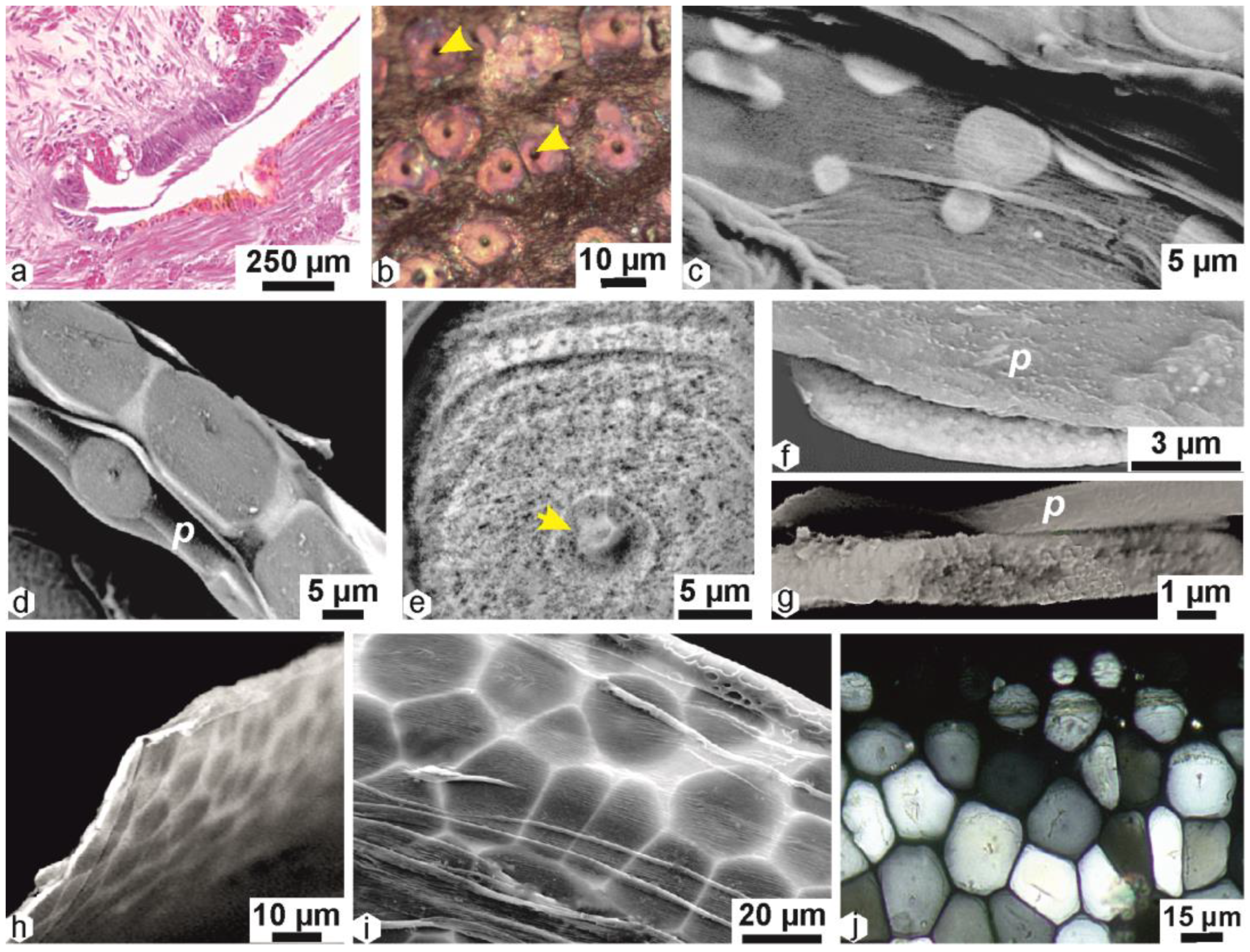
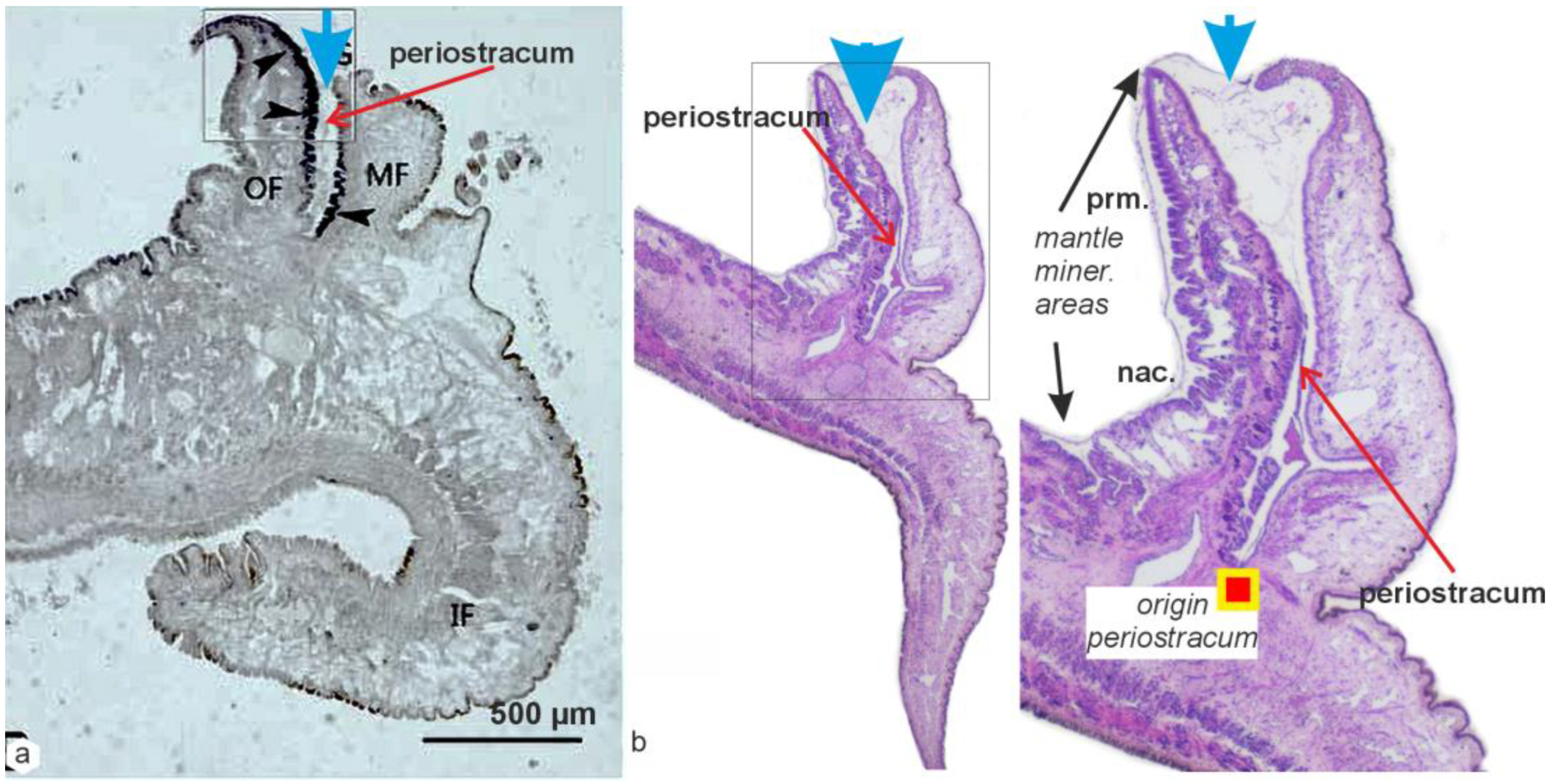
2.3. Calcification Associated with Periostracum Growth: Specific Mineralization Process Predating Prism Formation
2.4. Origin of the Prismatic Units at the Shell Grooving Edge: Transition between the Calcite Disks of the Flexible Shell and the Mantle-Secreted Calcite Prisms
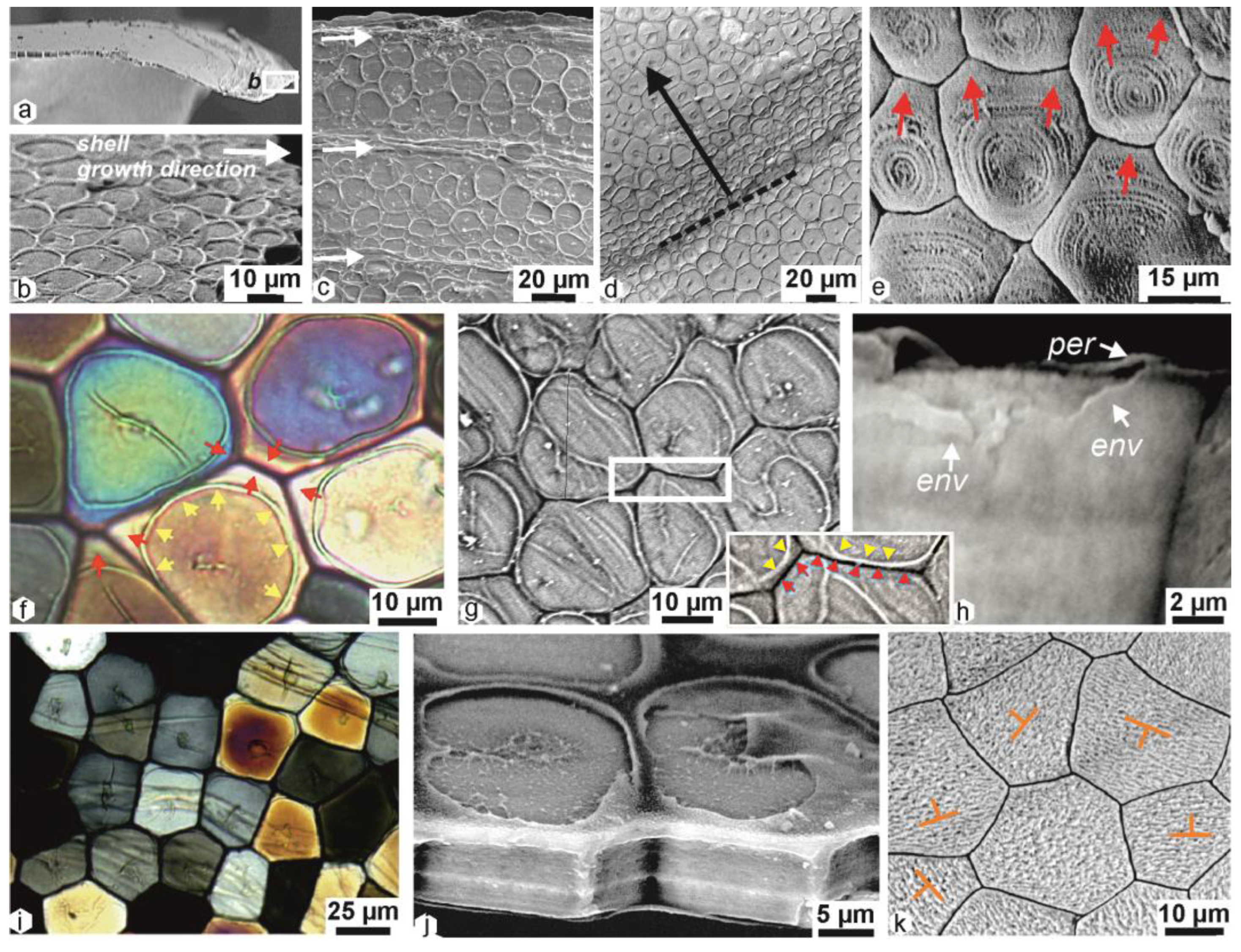
2.5. Structural Differentiation of the Prisms during Growth
3. Discussion: Biomineralization of Pinctada Prisms—Integrating Biological Data for Structural Analysis and Re-Examination of Former Models
3.1. Prisilkin-39 Calcite-Producing Protein Is Limited to Initial Mineralization Area
3.2. Former Interpretations of Prism Formation and Origin of the Prismatic Shell Layer in Pinctada
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A


References
- Mutvei, H.; Dunca, E.; Timm, H.; Slepukhina, T. Structure and growth rates of bivalve shells as indicators of environmental changes and pollution. Bull. Inst. Océanogr. Monaco 1996, 14, 65–72. [Google Scholar]
- Gröcke, D.R.; Gillikin, D.P. Advances in mollusc sclerochronology and sclerochemistry: Tools for understanding climate and environment. Geo-Mar. Lett. 2008, 28, 265–268. [Google Scholar] [CrossRef]
- Taylor, J.D. The structural evolution of the bivalve shell. Palaeontology 1973, 16, 519–534. [Google Scholar]
- Kobayashi, I. Various patterns of biomineralization and its phylogenetic significance in bivalve mollusks. In The Mechanism of Biomineralization in Animals and Plants; Omori, M., Watabe, N., Eds.; Tokai University Press: Tokyo, Japan, 1980; pp. 145–155. [Google Scholar]
- Kobayashi, I. Evolutionary trends of shell microstructure in Bivalve Molluscs. In Mechanisms and Phylogeny of Mineralization in Biological Systems; Suga, S., Nakahara, H., Eds.; Springer: Tokyo, Japan, 1991; pp. 415–419. [Google Scholar] [CrossRef]
- D’Orbigny, A. Paléontologie Française: Description Zoologique Et Géologique de tous les Animaux Mollusques et Rayonnés Fossiles de France: Comprenant leur Application à la Reconnaissance des Couches; Chez L’auteur: Paris, France; pp. 1842–1860, 4000p, 1440 pl.
- Stenzel, H.B. Oysters: Composition of the larval shell. Science 1964, 145, 155–156. [Google Scholar] [CrossRef] [PubMed]
- Carriker, M.R.; Palmer, R.E. Ultrastructural morphogenesis of prodissoconch and early dissoconch valves of the oyster Crassostrea virginica. Proc. Natl. Shellfish. Assoc. 1979, 69, 103–128. [Google Scholar]
- Eyster, L.S. Shell inorganic composition and onset of shell mineralization during Bivalve and Gastropod embryogenesis. Biol. Bull. 1986, 170, 211–231. [Google Scholar] [CrossRef]
- Weiss, I.M.; Tuross, N.; Addadi, L.; Weiner, S. Mollusc larval shell formation: Amorphous calcium carbonate is a precursor phase for aragonite. J. Exper. Zool. 2002, 293, 478–491. [Google Scholar] [CrossRef]
- Gray, J.E. Conchological observations, being an attempt to fix the study of conchology on a firm basis. Zool. J. Lond. 1825, 1, 204–223. [Google Scholar]
- Kobayashi, I. Shell structure of Pinctada fucata martensii (Dunker) in the larval and juvenile stages. Earth Sci. 1981, 35, 245–252, (In Japanese with English Abstract). [Google Scholar]
- Alagarswami, K.; Dharmara, J.A.; Chellam, A.; Velayudhan, T.S. Larval and juvenile rearing of black-lip pearl oyster, Pinctada margaritifera (Linnaeus). Aquaculture 1989, 76, 43–56. [Google Scholar] [CrossRef]
- Mao Che, L.; Golubic, S.; Le Campion-Alsumard, T.; Payri, C. Developmental aspects of biomineralization in the Polynesian pearl oyster Pinctada margaritifera var. cumingii. Oceanol. Acta 2000, 24, S37–S49. [Google Scholar] [CrossRef]
- Doroudi, M.S.; Southgate, P.C. Embryonic and larval development of Pinctada margaritifera (Linnaeus, 1758). Moll. Res. 2003, 23, 101–107. [Google Scholar] [CrossRef]
- Yokoo, N.; Suzuki, M.; Saruwatari, K.; Aoki, H.; Watanabe, K.; Nagasawa, H.; Kogure, K. Microstructures of the larval shell of a pearl oyster, Pinctada fucata, investigated by FIB-TEM technique. Am. Miner. 2011, 96, 1020–1027. [Google Scholar] [CrossRef]
- Wilbur, K.M. Shell formation and regeneration. In Physiology of Mollusca; Wilbur, K.M., Yonge, C.M., Eds.; Academic Press: New York, NY, USA; London, UK, 1964; Volume 1, pp. 243–282. [Google Scholar]
- Wada, K. Biomineralogy and Pearl Culture—3. J. Gemmol. Soc. Jpn. 1975, 2, 99–104. [Google Scholar] [CrossRef]
- Saleuddin, A.S.M. Shell formation in molluscs with special reference to periostracum formation and regeneration. In Pathways in Malacology; Van der Spoel, S., Van Bruggen, A.C., Lever, J., Eds.; Bohn, Scheltema & Holkema: Utrecht, The Netherlands, 1979; pp. 47–81. [Google Scholar]
- Haas, W. Evolution of calcareous hardparts in primitive molluscs. Malacologia 1981, 21, 403–418. [Google Scholar]
- Cuif, J.P.; Burghammer, M.; Chamard, V.; Dauphin, Y.; Godard, P.; Le Moullac, G.; Nehrke, G.; Perez-Huerta, A. Evidence of a biological control over origin, growth and end of the calcite prisms in the shells of Pinctada margaritifera (Pelecypod, Pterioidea). Minerals 2014, 4, 815–834. [Google Scholar] [CrossRef]
- Harper, E.M.; Checa, A.G. Tightly shut: Flexible valve margins and microstructural asymmetry in pterioid bivalves. Mar. Bio. 2020, 167, 78. [Google Scholar] [CrossRef]
- Beniash, E.; Aizenberg, J.; Addadi, L.; Weiner, S. Amorphous calcium carbonate transforms into calcite during sea urchin larval spicule growth. Proc. R. Soc. Lond. 1997, B264, 461–465. [Google Scholar] [CrossRef]
- Watabe, N.; Wada, K. On the shell structures of Japanese pearl oyster, Pinctada martensii (Dunker). (I) Prismatic layer. Rep. Fac. Fish. Prefec. Univ. Mie 1956, 2, 227–231. [Google Scholar]
- Dauphin, Y.; Cuif, J.P.; Doucet, J.; Salomé, M.; Susini, J.; Williams, C.T. In situ chemical speciation of sulfur in calcitic biominerals and the simple prism concept. J. Struct. Biol. 2003, 142, 272–280. [Google Scholar] [CrossRef]
- Dauphin, Y.; Zolotoyabko, E.; Berner, A.; Lakin, E.; Rollion-Bard, C.; Cuif, J.P.; Fratzl, P. Breaking the long-standing morphological paradigm: Individual prisms in the pearl oyster shell grow perpendicular to the c-axis of calcite. J. Struct. Biol. 2019, 205, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Cuif, J.P.; Belhadj, O.; Borensztajn, S.; Geze, M.; Trigos-Santos, S.; Prado, P.; Dauphin, Y. Prism substructures in the shell of Pinna nobilis (Linnaeus, 1758) Mollusca—Evidence for a three-dimensional pulsed-growth model. Heliyon 2020, 6, e04513. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Jing, G.; Yan, Z.; Li, C.; Gong, N.; Zhu, F.; Li, D.; Zhang, Y.; Zheng, G.; Wang, H.; et al. Cloning and characterization of Prisilkin-39, a novel matrix protein serving a dual role in the prismatic layer formation from the oyster Pinctada fucata. J. Biol. Chem. 2009, 284, 10841–10854. [Google Scholar] [CrossRef]
- Carpenter, W.B. On the microscopic structure of shells: Part I. Br. Assoc. Adv. Sci. 1844, 14, 1–24. [Google Scholar]
- Carpenter, W.B. On the microscopic structure of shells: Part II. Br. Assoc. Adv. Sci. 1847, 17, 93–134. [Google Scholar]
- Schmidt, W.J. Die Bausteine des Tierkörpers in Polarisiertem Lichte; Cohen, F.: Bonn, Germany, 1924; 528p. [Google Scholar]
- Boggild, O.B. The shell structure of the molluscs. D. Kgl. Danske Vidensk. Selsk. Skr. Naturvidensk. Ogmathem. 1930, 9, 231–326. [Google Scholar]
- Taylor, J.D.; Kennedy, W.J.; Hall, A. The shell structure and mineralogy of the Bivalvia. I. Introduction. Nuculacae—Trigonacae. Bull. Br. Mus. Nat. Hist. Zool. 1969, 3, 1–125. [Google Scholar]
- Taylor, J.D.; Kennedy, W.J.; Hall, A. The shell structure and mineralogy of the Bivalvia. II. Lucinacea-Clavagellacea. Conclusions. Bull.Br. Mus. Nat. Hist. Zool. 1973, 22, 253–294. [Google Scholar] [CrossRef]
- Nakahara, H.; Bevelander, G. The formation and growth of the prismatic layer of Pinctada radiata. Calcif. Tissue Res. 1971, 7, 31–45. [Google Scholar] [CrossRef]
- Checa, A.G.; Rodriguez-Navarro, A.B.; Esteban-Delgado, F.J. The nature and formation of calcitic columnar prismatic shell layers in pteriomorphian bivalves. Biomaterials 2005, 26, 6404–6414. [Google Scholar] [CrossRef]
- Checa, A.G.; Bonarski, J.T.; Willinger, M.G.; Faryna, M.; Berent, K.; Kania, B.; Gonzalez-Segura, A.; Pina, C.M.; Pospiech, J.; Morawiec, A. Crystallographic orientation in homogeneity and crystal splitting in biogenic calcite. J. Roy. Soc. Interface 2013, 10, 20130425. [Google Scholar] [CrossRef] [PubMed]
- Checa, A.; Macias-Sanchez, E.; Harper, E.M.; Cartwright, J.H.E. Organic membranes determine the pattern of the columnar prismatic layer of mollusc shells. Proc. R. Soc. 2016, B283, 20160032. [Google Scholar] [CrossRef] [PubMed]
- Checa, A.; Salas, C. Periostracum and shell formation in the Bivalvia. In Treatise on Line, 93; part N, revised; University of Kansas, Paleontological Institute: Lawrence, KS, USA, 2017; Volume 1, Chapter 3; pp. 1–51. [Google Scholar]
- Checa, A.G. Physical and biological determinants of the fabrication of molluscan shell microstructures. Front. Mar. Sci. 2018, 5, 353. [Google Scholar]
- Ubukata, T. Architectural constraints of the morphogenesis of prismatic structure in Bivalvia. Palaeontology 1994, 37, 241–261. [Google Scholar]
- Okumura, T.; Suzuki, M.; Nagasawa, H.; Kogure, T. Microstructural variation of biogenic calcite with intracrystalline organic macromolecules. Cryst. Growth Des. 2011, 12, 224–230. [Google Scholar] [CrossRef]
- Okumura, T.; Suzuki, M.; Nagasawa, H.; Kogure, T. Microstructural control of calcite via incorporation of intracrystalline organic molecules in shells. J. Cryst. Growth 2013, 381, 114–120. [Google Scholar] [CrossRef]
- Suzuki, M.; Okumura, T.; Nagasawa, H.; Kogure, T. Localization of intracrystalline organic macromolecules in mollusk shells. J. Crys. Growth 2011, 337, 24–29. [Google Scholar] [CrossRef]
- Dauphin, Y. Soluble organic matrices of the calcitic prismatic shell layers of two pteriomorphid Bivalves: Pinna nobilis and Pinctada margaritifera. J. Biol. Chem. 2003, 278, 15168–15177. [Google Scholar] [CrossRef]
- Duboisset, J.; Ferrand, P.; Baroni, A.; Grünewald, T.; Dicko, H.; Grauby, O.; Vidal-Dupio, J.; Saulnier, D.; Le Moullac, G.; Rosenthal, M.; et al. Amorphous-to-crystal transition in the layer-by-layer growth of bivalve shell prism. Acta Biomater. 2022, 142, 194–207. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuif, J.-P.; Fougerouse, A.; Lo, C.; Dauphin, Y. Formation of the Outer Shell Layer in Pinctada margaritifera: Structural and Biochemical Evidence for a Sequential Development of the Calcite Units. Minerals 2023, 13, 1301. https://doi.org/10.3390/min13101301
Cuif J-P, Fougerouse A, Lo C, Dauphin Y. Formation of the Outer Shell Layer in Pinctada margaritifera: Structural and Biochemical Evidence for a Sequential Development of the Calcite Units. Minerals. 2023; 13(10):1301. https://doi.org/10.3390/min13101301
Chicago/Turabian StyleCuif, Jean-Pierre, Angélique Fougerouse, Cedrik Lo, and Yannicke Dauphin. 2023. "Formation of the Outer Shell Layer in Pinctada margaritifera: Structural and Biochemical Evidence for a Sequential Development of the Calcite Units" Minerals 13, no. 10: 1301. https://doi.org/10.3390/min13101301