
Spatial Study of Enzymatic Activities from Bacterial Isolates in a Mediterranean Urban Park


Abstract
:1. Introduction
2. Materials and Methods
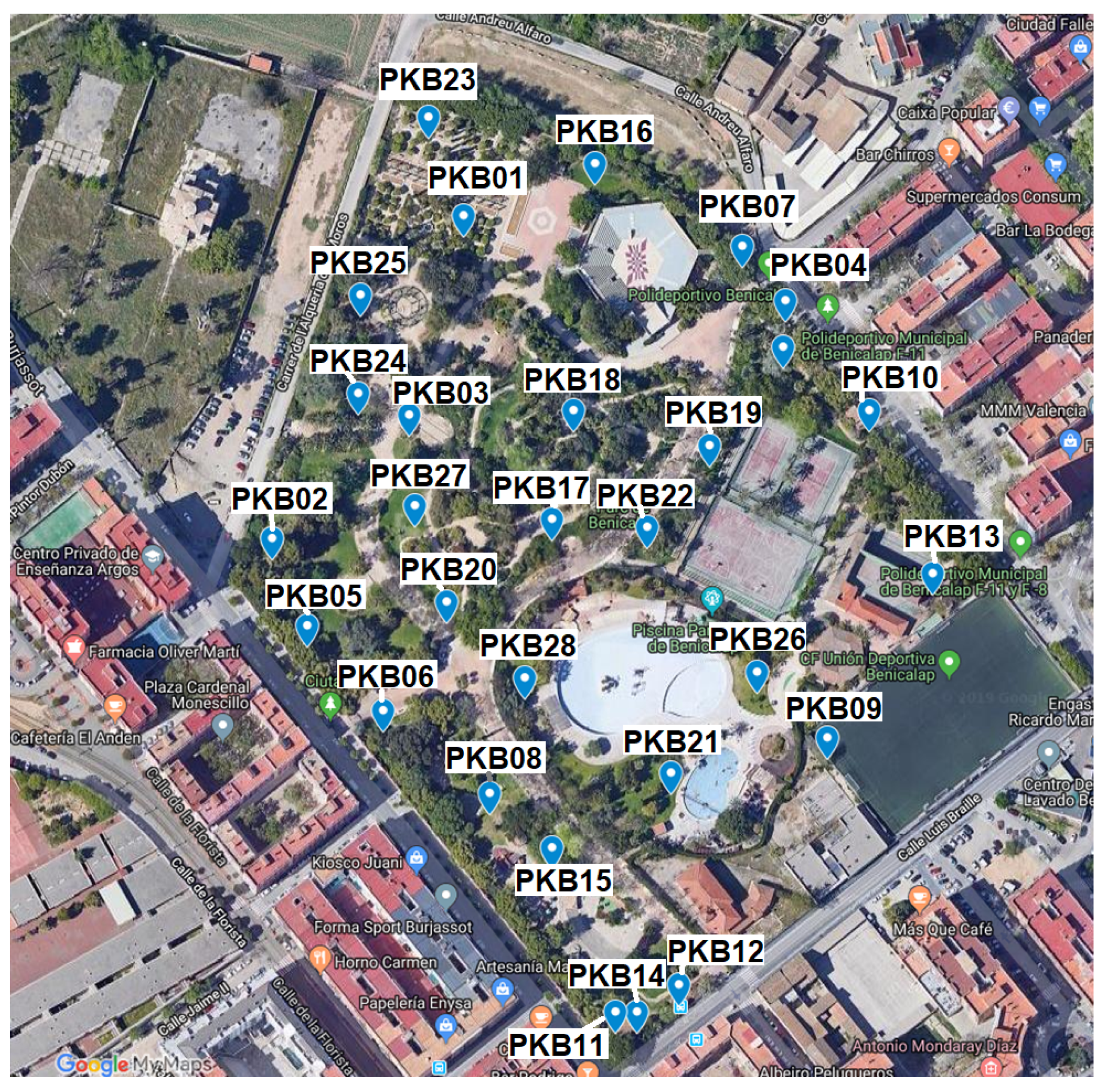
2.1. Sample Collection
2.2. Soil Characterization
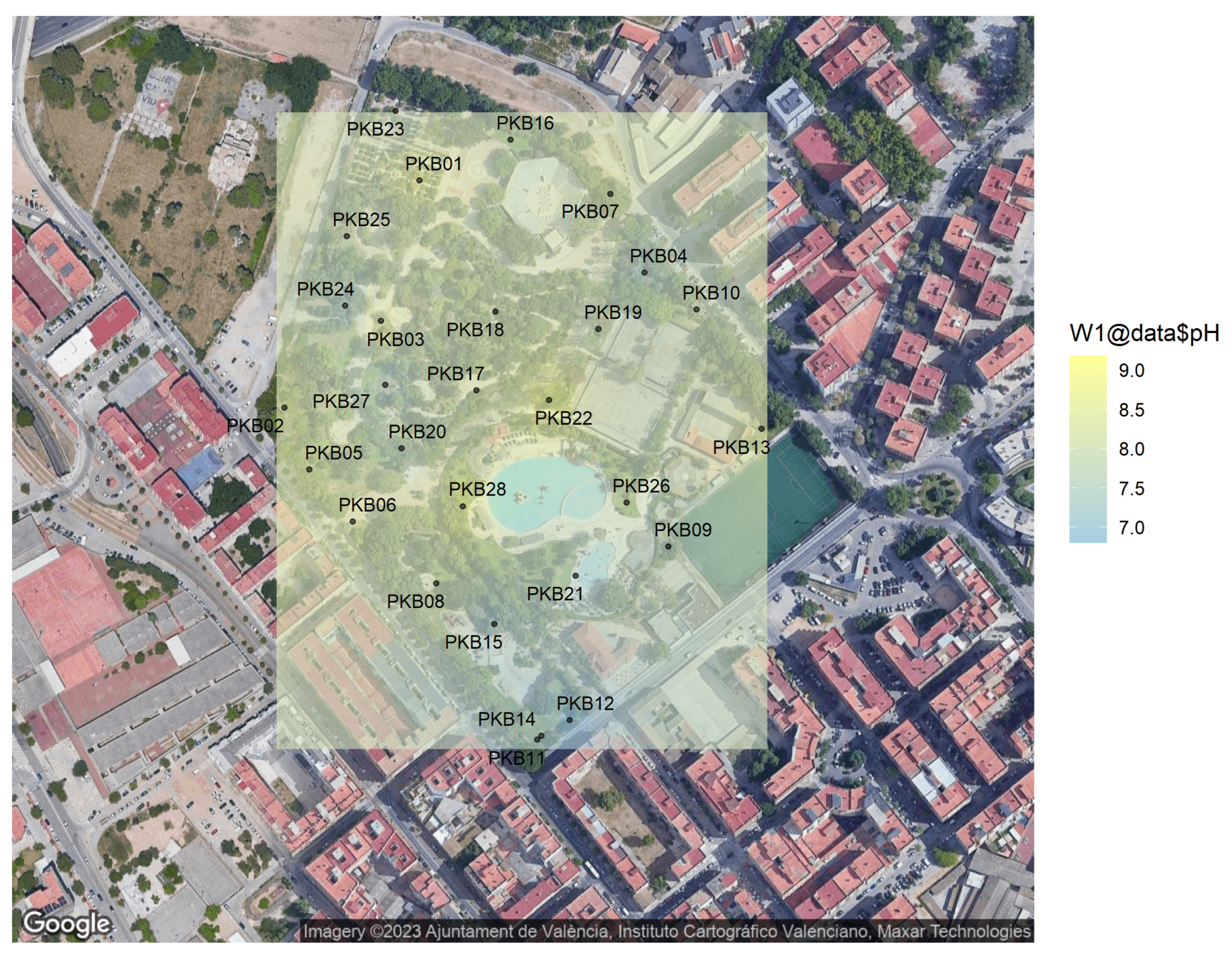
2.2.1. pH Measurement
2.2.2. Color and Texture Determination
2.3. Isolation and Antibiosis Characterization of Soil Bacteria
2.4. Microbial Identification
2.4.1. 16S rDNA Partial Sequence
2.4.2. MALDI-TOF
2.5. Assays for Extracellular Hydrolytic Enzymes Production
- Casein agar: 10 bacteriological peptone, 4 NaCl, 3 meat extract, 15 agar; 10% of skim milk was sterilized in a separate vessel. Once sterile, both parts were poured onto Petri plates. The plates were incubated at 28 °C for 3 days. The presence of casein gives the environment an opaque appearance that disappears when casein is hydrolyzed. A transparent halo must appear in the middle of the proteolytic microorganisms grown on casein. The non-proteolytic ones do not produce a change in the original aspect of the environment.
- TWEEN-80 agar: This medium consists of a synthetic lipid containing ester links between sorbitol and oleic acid (Tween-80) and calcium salts. When microorganisms possessing esterase activity (lipase) are present, they can hydrolyze the ester link, leading to the release of oleic acid from Tween-80. In the presence of an excess amount of Ca, the released oleic acid forms small crystalline oleate crystals that create an opaque halo around the growth area.
- Starch Agar: To reveal the presence of starch, it was necessary to dye it with Lugol. We added 2 milliliters of Lugol to the plate and observed the development of the dark violet color in areas where there was starch. If transparent halos appeared around the growth of a microorganism, this indicated that there was polysaccharide hydrolysis.
- DNase agar: 20 tryptose, 2 deoxyribonucleic acid, 5 sodium chloride, 12 agar. The agar medium was inoculated and incubated with the organisms, then the surface growth was flooded with 1N hydrochloric acid. Polymerized DNA, which is present in the medium, will precipitate in the presence of 1N HCl and cause the medium to become opaque. If the organisms produce enough DNase enzymes to hydrolyze the DNA, clear zones will be observed around the colonies. As a control, Staphylococcus aureus CECT4013 was used.
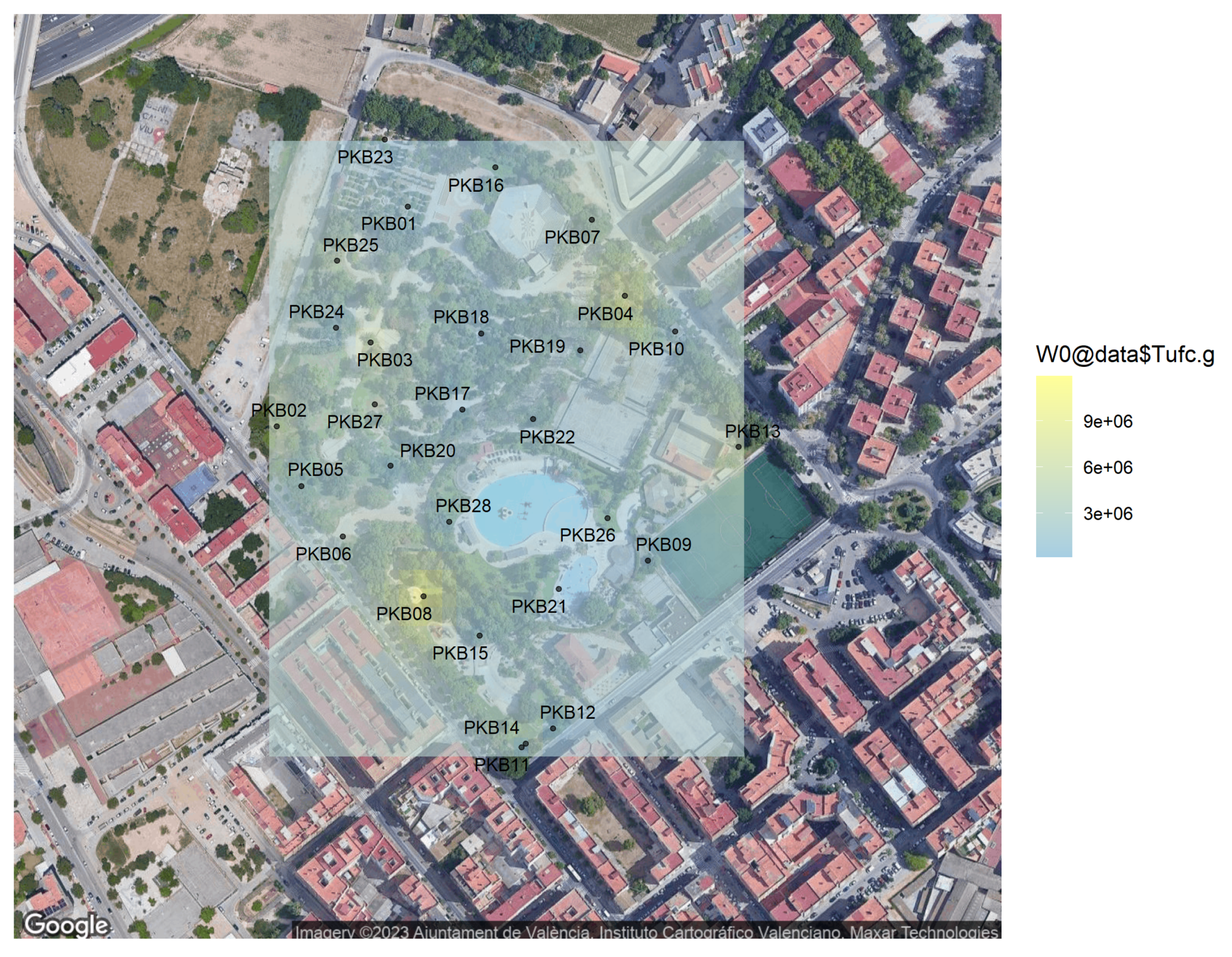
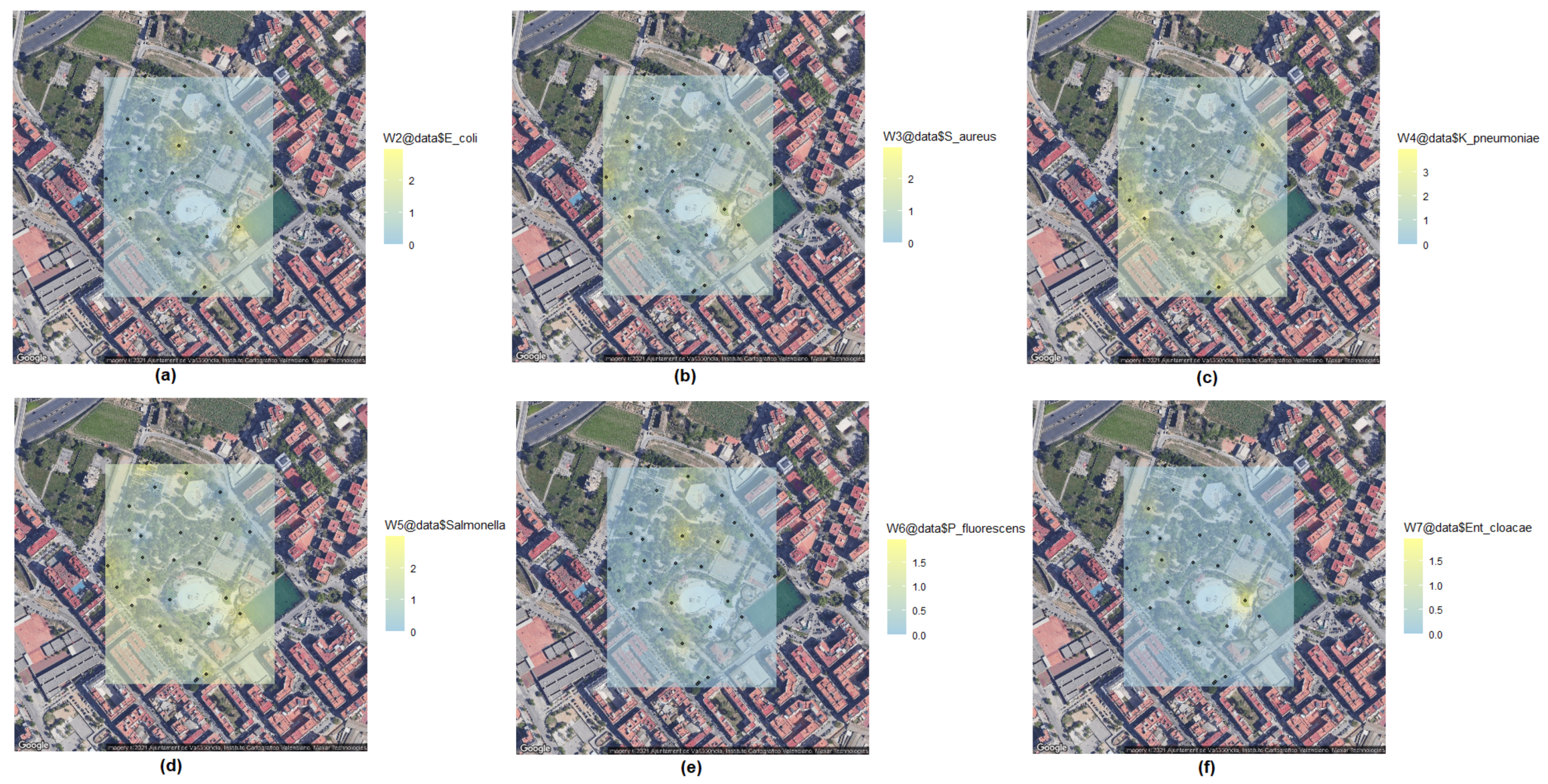
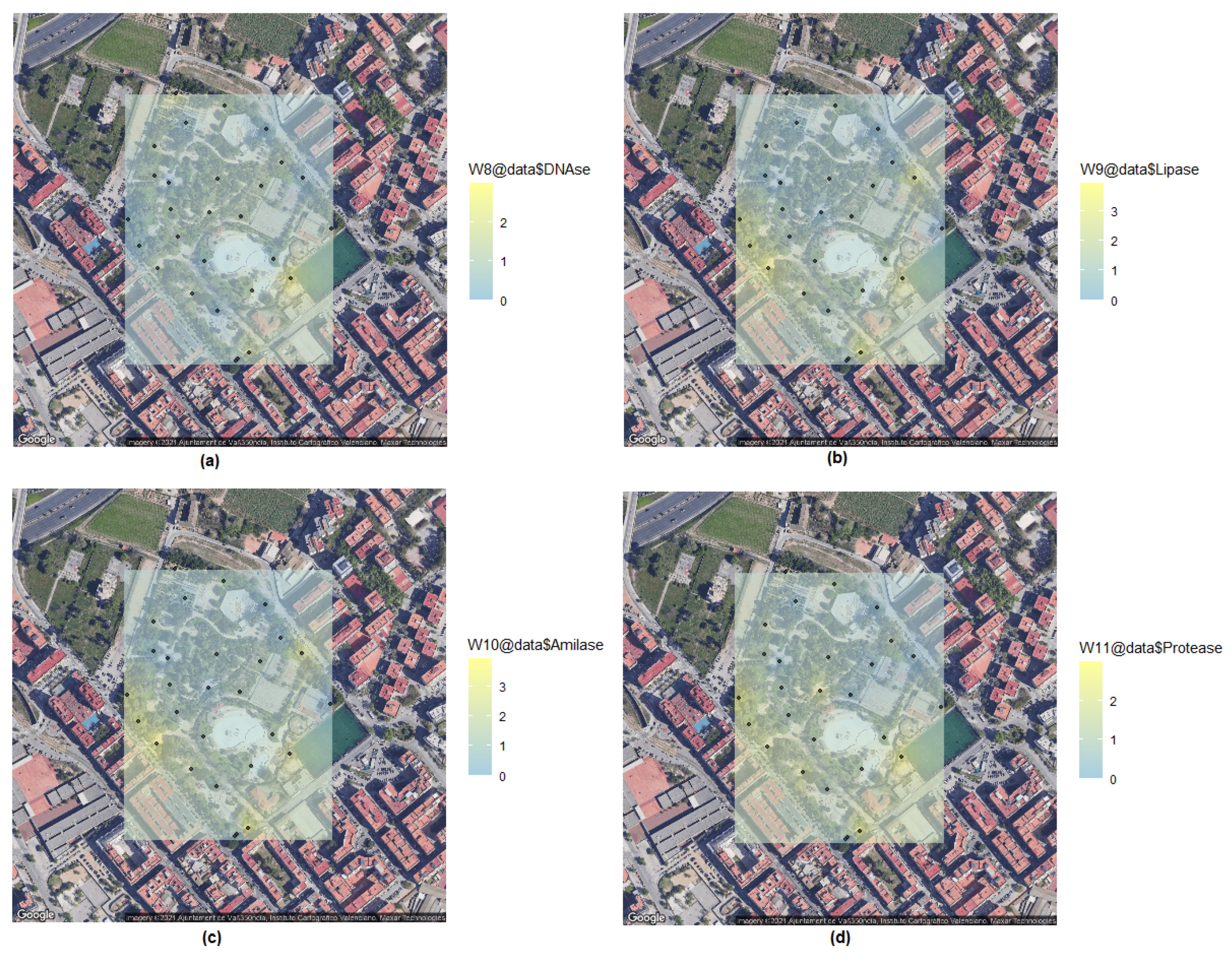
2.6. Kriging Method for Biological Spatial Statistics
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Delaimy, W.; Webb, M. Community Gardens as Environmental Health Interventions: Benefits Versus Potential Risks. Curr. Environ. Health Rep. 2017, 4, 252–265. [Google Scholar] [CrossRef]
- Mafiz, A.I.; Perera, L.N.; He, Y.; Zhang, W.; Xiao, S.; Hao, W.; Sun, S.; Zhou, K.; Zhang, Y. Case study on the soil antibiotic resistome in an urban community garden. Int. J. Antimicrob. Agents 2018, 52, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.; Warren, P.; Thompson, K.; Smith, R. Urban domestic gardens (IV): The extent of the resource and its associated features. Biodivers. Conserv. 2005, 14, 3327–3349. [Google Scholar] [CrossRef]
- Loram, A.; Tratalos, J.; Warren, P.; Gaston, K. Urban domestic gardens (X): The extent & structure of the resource in five major cities. Landsc. Ecol. 2007, 22, 601–615. [Google Scholar] [CrossRef]
- Mathieu, R.; Freeman, C.; Aryal, J. Mapping private gardens in urban areas using object-oriented techniques and very high-resolution satellite imagery. Landsc. Urban Plan. 2007, 81, 179–192. [Google Scholar] [CrossRef]
- Perera, L.N.; Mafiz, A.I.; Amarasekara, N.R.; Chang, E.; Rao, V.B.K.; Zhang, Y. Antimicrobial-resistant E. coli and Enterococcus spp. Recovered from urban community gardens. Food Control 2020, 108, 106857. [Google Scholar] [CrossRef]
- Yuan, W.; Zhang, Y.; Riaz, L.; Yang, Q.; Du, B.; Wang, R. Multiple antibiotic resistance and DNA methylation in Enterobacteriaceae isolates from different environments. J. Hazard. Mater. 2021, 402, 123822. [Google Scholar] [CrossRef]
- Matthiessen, L.; Bergström, R.; Dustdar, S.; Meulien, P.; Draghia-Akli, R. Increased momentum in antimicrobial resistance research. Lancet 2016, 388, 865. [Google Scholar] [CrossRef]
- Mao, D.; Yu, S.; Rysz, M.; Luo, Y.; Yang, F.; Li, F.; Hou, J.; Mu, Q.; Alvarez, P. Prevalence and proliferation of antibiotic resistance genes in two municipal wastewater treatment plants. Water Res. 2015, 85, 458–466. [Google Scholar] [CrossRef]
- Rodriguez-Mozaz, S.; Chamorro, S.; Marti, E.; Huerta, B.; Gros, M.; Sànchez-Melsió, A.; Borrego, C.; Barceló, D.; Balcázar, J. Occurrence of antibiotics and antibiotic resistance genes in hospital and urban wastewaters and their impact on the receiving river. Water Res. 2015, 69, 234–242. [Google Scholar] [CrossRef]
- Yuan, W.; Tian, T.; Yang, Q.; Riaz, L. Transfer potentials of antibiotic resistance genes in Escherichia spp. strains from different sources. Chemosphere 2019, 246, 125736. [Google Scholar] [CrossRef] [PubMed]
- Neiderud, C.J. How urbanization affects the epidemiology of emerging infectious diseases. Afr. J. Disabil. 2015. [Google Scholar] [CrossRef] [PubMed]
- Cooley, J.D.; Wong, W.C.; Jumper, C.A.; Straus, D.C. Correlation between the prevalence of certain fungi and sick building syndrome. Occup. Environ. Med. 1998, 55, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, N.; Siddique, N.; Custovic, A. Allergic disease in urban and rural populations: Increasing prevalence with increasing urbanization. Allergy Eur. J. Allergy Clin. Immunol. 2005, 60, 1357–1360. [Google Scholar] [CrossRef]
- Gilbert, J.A.; Stephens, B. Microbiology of the built environment. Nat. Rev. Microbiol. 2018, 16, 661–670. [Google Scholar] [CrossRef]
- ECDC. ECDC Ears-Net Database. Available online: https://www.ecdc.europa.eu/en/antimicrobial-resistance/surveillance-and-disease-data/data-ecdc (accessed on 24 August 2021).
- Danko, D.; Bezdan, D.; Afshin, E.E.; Ahsanuddin, S.; Bhattacharya, C.; Butler, D.J.; Chng, K.R.; Donnellan, D.; Hecht, J.; Jackson, K.; et al. A global metagenomic map of urban microbiomes and antimicrobial resistance. Cell 2021, 184, 3376–3393.e17. [Google Scholar] [CrossRef]
- Arnal, D.; Moya, C.; Filippelli, L.; Segura-Garcia, J.; Maicas, S. Bacteria spatial tracking in Urban Park soils with MALDI-TOF Mass Spectrometry and Specific PCR. Biodata Min. 2023, 16, 1. [Google Scholar] [CrossRef]
- Thompson, J.; Pollio, A.; Turk, P. Comparison of munsell soil color charts and the globe soil color book. Soil Sci. Soc. Am. J. 2013, 77, 2089–2093. [Google Scholar] [CrossRef]
- Porta-Casanellas, J. Técnicas y Experiments en Edafología; Colegio Oficial de Ingenieros Agrónomos de Cataluña: Barcelona, Spain, 1986. [Google Scholar]
- Alden, A. Sand, Silt, and Clay Soil Classification Diagram; ThoughtCo.: New York, NY, USA, 2020. [Google Scholar]
- Jorge, N.F.; Clark, J.; Cárdenas, M.L.; Geoghegan, H.; Shannon, V. Measuring soil colour to estimate soil organic carbon using a large-scale citizen science-based approach. Sustainability 2021, 13, 11029. [Google Scholar] [CrossRef]
- Arahal, D.R.; Sánchez, E.; Macián, M.C.; Garay, E. Value of recN sequences for species identification and as a phylogenetic marker within the family “Leuconostocaceae”. Int. Microbiol. 2008, 11, 33–39. [Google Scholar] [CrossRef]
- Small World Initiative. Crowdsourcing Antibiotic Discovery. Available online: https://www.smallworldinitiative.org/about (accessed on 27 December 2019).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.; Klepel, S.; Renner, U.; Kostrzewa, M. Fast and reliable MALDI-TOF MS-based microorganism identification. Nat. Methods 2006, 3, i–ii. [Google Scholar] [CrossRef]
- Isaaks, E.-H.; Srivastava, R.M. An Introduction to Applied Geostatistics; Oxford University Press: New York, NY, USA, 1989. [Google Scholar]
- Cressie, N. Statistics for Spatial Data; John Wiley: New York, NY, USA, 1993. [Google Scholar]
- Madigan, M.; Bender, D.; Buckley, D.; Sattley, W.; Stahl, D. Brock Biology of Microorganisms, 15th ed.; Pearson: London, UK, 2018. [Google Scholar]
- Jordán, M.M.; Almendro-Candel, M.B.; Navarro-Pedreño, J.; Pardo, F.; García-Sánchez, E.; Bech, J. Bioavailability, mobility and leaching of phosphorus in a Mediterranean agricultural soil (ne Spain) amended with different doses of biosolids. Environ. Geochem. Health 2022, 44, 7–14. [Google Scholar] [CrossRef]
- Jordán-Vidal, M.; Montero-Martínez, M.A.; García-Sánchez, E.; Martinez-Poveda, A. Metal contamination of agricultural soils amended with biosolids (sewage sludge) at a ceramic production area in NE Spain: A 10-year resampling period. Soil Use Manag. 2021, 37, 307–318. [Google Scholar] [CrossRef]
- Maicas, S.; Fouz, B.; Figàs-Segura, A.; Zueco, J.; Rico, H.; Navarro, A.; Carbó, E.; Segura-Garcia, J.; Biosca, E. Implementation of Antibiotic Discovery by Student Crowdsourcing in the Valèncian Community Through a Service Learning Strategy. Front. Microbiol. 2020, 11, 564030. [Google Scholar] [CrossRef] [PubMed]
- Maicas, S.; Segura-Garcia, J. Spatial distribution of antibiotic-producing bacteria in urban areas. A case study in València (Spain). IEEE/Acm Trans. Comput. Biol. Bioinform. 2020, 18, 2877–2883. [Google Scholar] [CrossRef] [PubMed]
- Kateete, D.; Kimani, C.; Katabazi, F.; Okeng, A.; Okee, M.; Nanteza, A.; Joloba, M.; Najjuka, F. Identification of Staphylococcus aureus: DNase and Mannitol salt agar improve the efficiency of the tube coagulase test. Ann. Clin. Microbiol. Antimicrob. 2010, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Ten, L.; Liu, Q.M.; Im, W.T.; Lee, M.; Yang, D.C.; Lee, S.T. Pedobacter ginsengisoli sp. nov., a DNase-producing bacterium isolated from soil of a ginseng field in South Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 2565–2570. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Lysak, L.V.; Lapygina, E.V. The Diversity of Bacterial Communities in Urban Soils. Eurasian Soil Sci. 2018, 51, 1050–1056. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code | GPS Coord | Depth | pH | Color | Texture | Total ufc·g |
|---|---|---|---|---|---|---|
| PKB01 | 39.4986, −0.3973 | 2–5 | 8.71 | Dark | Sandy-loam | 1.70 × 10 |
| PKB02 | 39.4974, −0.3982 | 0–2 | 8.46 | Brown | Sandy-loam | 4.70 × 10 |
| PKB03 | 39.4979, −0.3976 | 0–2 | 8.63 | Brownish dark | Sandy-loam | 6.00 × 10 |
| PKB04 | 39.4981, −0.3958 | 2–5 | 7.60 | Grey | Sandy-loam | 9.00 × 10 |
| PKB05 | 39.4971, −0.3981 | 0–2 | 8.57 | Brown | Sandy-loam | 9.40 × 10 |
| PKB06 | 39.4968, −0.3978 | 0–2 | 8.53 | Dark | Sandy-loam | 3.20 × 10 |
| PKB07 | 39.4985, −0.3960 | 5–10 | 8.48 | Dark | Sandy-loam | 4.00 × 10 |
| PKB08 | 39.4965, −0.3972 | 2–5 | 8.19 | Dark | Sandy-loam | 1.20 × 10 |
| PKB09 | 39.4967, −0.3957 | 2–5 | 7.80 | Dark | Sandy-loam | 7.40 × 10 |
| PKB10 | 39.4979, −0.3955 | 0–2 | 8.13 | Brown | Sandy-loam | 7.00 × 10 |
| PKB11 | 39.4957, −0.3965 | 2–5 | 7.50 | Dark | Sandy-loam | 6.60 × 10 |
| PKB12 | 39.4958, −0.3963 | 2–5 | 6.79 | Brownish dark | Sandy-loam | 9.70 × 10 |
| PKB13 | 39.4973, −0.3950 | 2–5 | 8.68 | Brownish dark | Sandy-loam | 6.40 × 10 |
| PKB14 | 39.4957, −0.3965 | 0–2 | 7.21 | Brownish dark | Sandy-loam | 5.70 × 10 |
| PKB15 | 39.4963, −0.3968 | 0–2 | 7.33 | Brown | Sandy | 6.60 × 10 |
| PKB16 | 39.4988, −0.3967 | 2–5 | 8.00 | Dark | Sandy | 4.60 × 10 |
| PKB17 | 39.4975, −0.3969 | 0–2 | 8.33 | Dark | Sandy-loam | 3.00 × 10 |
| PKB18 | 39.4979, −0.3968 | 0–2 | 8.28 | Bright | Sandy | 4.90 × 10 |
| PKB19 | 39.4978, −0.3961 | 2–5 | 8.09 | Dark | Sandy-loam | 1.20 × 10 |
| PKB20 | 39.4972, −0.3974 | 0–2 | 7.89 | Dull | Sandy | 5.60 × 10 |
| PKB21 | 39.4966, −0.3963 | 0–2 | 7.63 | Light | Sandy | 1.20 × 10 |
| PKB22 | 39.4975, −0.3965 | 5–10 | 9.24 | Dull | Sandy-loam | 9.30 × 10 |
| PKB23 | 39.4990, −0.3975 | 2–5 | 7.85 | Dark | Sandy | 4.30 × 10 |
| PKB24 | 39.4980, −0.3978 | 0–2 | 7.55 | Brownish dark | Sandy-loam | 6.80 × 10 |
| PKB25 | 39.4983, −0.3978 | 2–5 | 8.11 | Brownish dark | Sandy-loam | 4.10 × 10 |
| PKB26 | 39.4969, −0.3959 | 2–5 | 8.06 | Dull orange | Sandy | 7.20 × 10 |
| PKB27 | 39.4976, −0.3976 | 2–5 | 7.47 | Grey | Sandy-loam | 3.30 × 10 |
| PKB28 | 39.4969, −0.3970 | 0–2 | 9.00 | Dull orange | Sandy-loam | 7.30 × 10 |
| Sample | E. coli | S. aureus | K. pneumoniae | Salmonella sp. | P. fluorescens | Ent. cloacae |
|---|---|---|---|---|---|---|
| PKB01 | 0 | 0 | 0 | 0 | 0 | 0 |
| PKB02 | 0 | 3 | 3 | 3 | 0 | 0 |
| PKB03 | 0 | 0 | 0 | 0 | 0 | 0 |
| PKB04 | 1 | 1 | 0 | 1 | 0 | 0 |
| PKB05 | 0 | 0 | 3 | 2 | 0 | 0 |
| PKB06 | 1 | 2 | 4 | 2 | 0 | 0 |
| PKB07 | 0 | 0 | 0 | 0 | 0 | 0 |
| PKB08 | 1 | 1 | 2 | 2 | 0 | 0 |
| PKB09 | 3 | 2 | 3 | 3 | 0 | 0 |
| PKB10 | 0 | 1 | 3 | 0 | 0 | 0 |
| PKB11 | 0 | 0 | 0 | 0 | 0 | 0 |
| PKB12 | 2 | 1 | 4 | 3 | 0 | 0 |
| PKB13 | 1 | 1 | 0 | 1 | 0 | 0 |
| PKB14 | 1 | 0 | 1 | 1 | 0 | 0 |
| PKB15 | 0 | 0 | 2 | 2 | 1 | 0 |
| PKB16 | 0 | 0 | 1 | 2 | 1 | 0 |
| PKB17 | 0 | 0 | 0 | 1 | 0 | 0 |
| PKB18 | 3 | 2 | 1 | 1 | 2 | 0 |
| PKB19 | 0 | 0 | 0 | 1 | 1 | 0 |
| PKB20 | 0 | 0 | 1 | 2 | 0 | 0 |
| PKB21 | 0 | 0 | 2 | 1 | 0 | 0 |
| PKB22 | 0 | 0 | 1 | 1 | 0 | 0 |
| PKB23 | 1 | 1 | 0 | 3 | 0 | 0 |
| PKB24 | 0 | 2 | 1 | 1 | 0 | 0 |
| PKB25 | 0 | 1 | 2 | 1 | 0 | 1 |
| PKB26 | 0 | 2 | 1 | 2 | 0 | 2 |
| PKB27 | 0 | 1 | 0 | 1 | 0 | 1 |
| PKB28 | 0 | 0 | 0 | 0 | 1 | 0 |
| Total | 14 | 21 | 35 | 37 | 6 | 4 |
| Sample | DNase | Lipase | Amylase | Protease |
|---|---|---|---|---|
| PKB01 | 0 | 0 | 0 | 0 |
| PKB02 | 0 | 3 | 3 | 3 |
| PKB03 | 0 | 0 | 0 | 0 |
| PKB04 | 1 | 1 | 0 | 1 |
| PKB05 | 0 | 0 | 3 | 2 |
| PKB06 | 1 | 2 | 4 | 2 |
| PKB07 | 0 | 0 | 0 | 0 |
| PKB08 | 1 | 1 | 2 | 2 |
| PKB09 | 3 | 2 | 3 | 3 |
| PKB10 | 0 | 1 | 3 | 0 |
| PKB11 | 0 | 0 | 0 | 0 |
| PKB12 | 2 | 1 | 4 | 3 |
| PKB13 | 1 | 2 | 0 | 1 |
| PKB14 | 1 | 0 | 1 | 1 |
| PKB15 | 0 | 1 | 1 | 1 |
| PKB16 | 1 | 0 | 1 | 1 |
| PKB17 | 1 | 0 | 0 | 2 |
| PKB18 | 1 | 0 | 1 | 1 |
| PKB19 | 1 | 0 | 1 | 0 |
| PKB20 | 1 | 0 | 1 | 1 |
| PKB21 | 1 | 0 | 1 | 1 |
| PKB22 | 1 | 0 | 1 | 0 |
| PKB23 | 2 | 0 | 1 | 1 |
| PKB24 | 1 | 0 | 0 | 1 |
| PKB25 | 1 | 2 | 1 | 1 |
| PKB26 | 0 | 1 | 1 | 1 |
| PKB27 | 1 | 1 | 1 | 1 |
| PKB28 | 0 | 0 | 1 | 1 |
| Total | 10 | 12 | 24 | 19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maicas, S.; Segura-Garcia, J. Spatial Study of Enzymatic Activities from Bacterial Isolates in a Mediterranean Urban Park. Land 2023, 12, 655. https://doi.org/10.3390/land12030655
Maicas S, Segura-Garcia J. Spatial Study of Enzymatic Activities from Bacterial Isolates in a Mediterranean Urban Park. Land. 2023; 12(3):655. https://doi.org/10.3390/land12030655
Chicago/Turabian StyleMaicas, Sergi, and Jaume Segura-Garcia. 2023. "Spatial Study of Enzymatic Activities from Bacterial Isolates in a Mediterranean Urban Park" Land 12, no. 3: 655. https://doi.org/10.3390/land12030655