
Local and Landscape Factors Influence Plant-Pollinator Networks and Bee Foraging Behavior across an Urban Corridor
 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Study System
2.2. Local and Landscape Characteristics
2.3. Pollinator Sampling
2.4. Pollen Identification
2.5. Pollen-Based Networks
2.6. Pollen Preference Analysis
3. Results
3.1. Predictors of Pollinator Community and Pollen-Based Networks
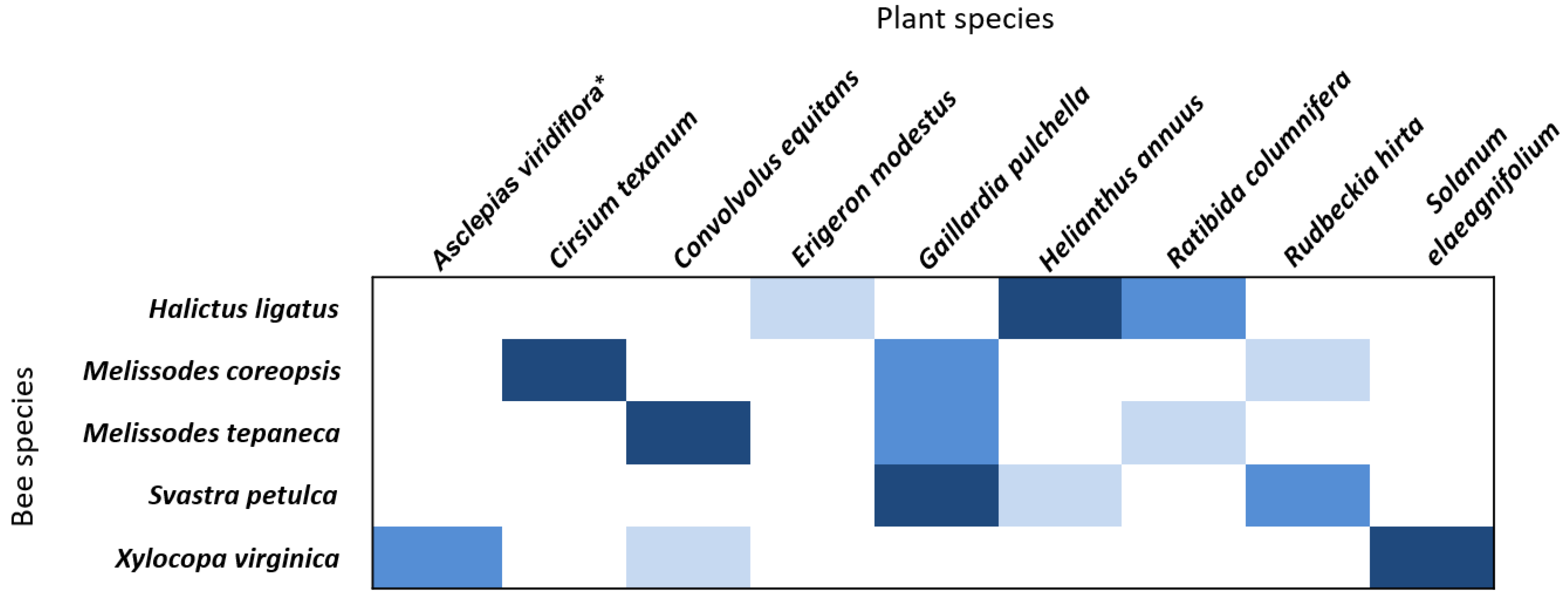
3.2. Pollen Preference Analysis
4. Discussion
4.1. Landscape-Level Habitat Composition Shapes Network Structure
4.2. Local Vegetation Drives Higher Network Connectance
4.3. Pollinators Show Distinct Floral Preferences for Native Flowers in Urban Landscapes
4.4. Conclusions and Conservation Applications
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fehlmann, G.; Oriain, M.J.; Fürtbauer, I.; King, A.J. Behavioral Causes, Ecological Consequences, and Management Challenges Associated with Wildlife Foraging in Human-Modified Landscapes. Bioscience 2021, 71, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Šálek, M.; Drahníková, L.; Tkadlec, E. Changes in Home Range Sizes and Population Densities of Carnivore Species along the Natural to Urban Habitat Gradient. Mamm. Rev. 2015, 45, 1–14. [Google Scholar] [CrossRef]
- Theodorou, P. The Effects of Urbanisation on Ecological Interactions. Curr. Opin. Insect. Sci. 2022, 52, 100922. [Google Scholar] [CrossRef] [PubMed]
- Pellissier, L.; Albouy, C.; Bascompte, J.; Farwig, N.; Graham, C.; Loreau, M.; Maglianesi, M.A.; Melián, C.J.; Pitteloud, C.; Roslin, T.; et al. Comparing Species Interaction Networks along Environmental Gradients. Biol. Rev. 2018, 93, 785–800. [Google Scholar] [CrossRef] [Green Version]
- Baldock, K.C.R.; Goddard, M.A.; Hicks, D.M.; Kunin, W.E.; Mitschunas, N.; Morse, H.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; et al. A Systems Approach Reveals Urban Pollinator Hotspots and Conservation Opportunities. Nat. Ecol. Evol. 2019, 3, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Barr, A.E.; van Dijk, L.J.A.; Hylander, K.; Tack, A.J.M. Local Habitat Factors and Spatial Connectivity Jointly Shape an Urban Insect Community. Landsc. Urban Plan. 2021, 214, 104177. [Google Scholar] [CrossRef]
- Jakobsson, A.; Ågren, J. Distance to Semi-Natural Grassland Influences Seed Production of Insect-Pollinated Herbs. Oecologia 2014, 175, 199–208. [Google Scholar] [CrossRef]
- Corro, E.J.; Ahuatzin, D.A.; Jaimes, A.A.; Favila, M.E.; Ribeiro, M.C.; López-Acosta, J.C.; Dáttilo, W. Forest Cover and Landscape Heterogeneity Shape Ant–Plant Co-Occurrence Networks in Human-Dominated Tropical Rainforests. Landsc. Ecol. 2019, 34, 93–104. [Google Scholar] [CrossRef]
- Seto, K.C.; Fragkias, M.; Güneralp, B.; Reilly, M.K. A Meta-Analysis of Global Urban Land Expansion. PLoS ONE 2011, 6, e23777. [Google Scholar] [CrossRef]
- IPBES. The Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on Pollinators, Pollination, and Food Production; Potts, S.G., Imperatriz-Fonseca, V.L., Ngo, H.T., Eds.; Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2016. [Google Scholar]
- Ollerton, J.; Winfree, R.; Tarrant, S. How Many Flowering Plants Are Pollinated by Animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Joosten, H.; de Klerk, P. Importance of Pollinators in Changing Landscapes for World Crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar]
- Ballare, K.M.; Neff, J.L.; Ruppel, R.; Jha, S. Multi-Scalar Drivers of Biodiversity: Local Management Mediates Wild Bee Community Response to Regional Urbanization. Ecol. Appl. 2019, 29, e01869. [Google Scholar] [CrossRef]
- Ayers, A.C.; Rehan, S.M. Supporting Bees in Cities: How Bees Are Influenced by Local and Landscape Features. Insects 2021, 12, 128. [Google Scholar] [CrossRef] [PubMed]
- Egerer, M.; Cecala, J.M.; Cohen, H. Wild Bee Conservation within Urban Gardens and Nurseries: Effects of Local and Landscape Management. Sustainability 2020, 12, 293. [Google Scholar] [CrossRef] [Green Version]
- Spiesman, B.J.; Inouye, B.D. Habitat Loss Alters the Architecture of Plant-Pollinator Interaction Networks. Ecology 2013, 94, 2688–2696. [Google Scholar] [CrossRef] [Green Version]
- Weiner, C.N.; Werner, M.; Linsenmair, K.E.; Bluthgen, N. Land-Use Impacts on Plant-Pollinator Networks: Interaction Strength and Specialization Predict Pollinator Declines. Ecology 2014, 95, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Theodorou, P.; Albig, K.; Radzevičiūtė, R.; Settele, J.; Schweiger, O.; Murray, T.E.; Paxton, R.J. The Structure of Flower Visitor Networks in Relation to Pollination across an Agricultural to Urban Gradient. Funct. Ecol. 2017, 31, 838–847. [Google Scholar] [CrossRef]
- Jauker, F.; Jauker, B.; Grass, I.; Steffan-Dewenter, I.; Wolters, V. Partitioning Wild Bee and Hoverfly Contributions to Plant–Pollinator Network Structure in Fragmented Habitats. Ecology 2019, 100, e02569. [Google Scholar] [CrossRef]
- Grass, I.; Jauker, B.; Steffan-Dewenter, I.; Tscharntke, T.; Jauker, F. Past and Potential Future Effects of Habitat Fragmentation on Structure and Stability of Plant–Pollinator and Host–Parasitoid Networks. Nat. Ecol. Evol. 2018, 2, 1408–1417. [Google Scholar] [CrossRef]
- Tommasi, N.; Biella, P.; Maggioni, D.; Fallati, L.; Agostinetto, G.; Labra, M.; Galli, P.; Galimberti, A. DNA Metabarcoding Unveils the Effects of Habitat Fragmentation on Pollinator Diversity, Plant-Pollinator Interactions, and Pollination Efficiency in Maldive Islands. Mol. Ecol. 2022, 1–11. [Google Scholar] [CrossRef]
- Moreira, E.F.; Boscolo, D.; Viana, B.F. Spatial Heterogeneity Regulates Plant-Pollinator Networks across Multiple Landscape Scales. PLoS ONE 2015, 10, e0123628. [Google Scholar] [CrossRef] [PubMed]
- Vaudo, A.D.; Patch, H.M.; Mortensen, D.A.; Tooker, J.F.; Grozinger, C.M. Macronutrient Ratios in Pollen Shape Bumble Bee (Bombus Impatiens) Foraging Strategies and Floral Preferences. Proc. Natl. Acad. Sci. USA 2016, 113, E4035–E4042. [Google Scholar] [CrossRef] [Green Version]
- Aebischer, N.J.; Robertson, P.A.; Kenward, R.E. Compositional Analysis of Habitat Use from Animal Radio-Tracking Data. Ecology 1993, 74, 1313–1325. [Google Scholar] [CrossRef]
- Thorp, R.W. The Collection of Pollen by Bees. In Pollen and Pollination; Danfi, A., Hesse, M., Pacini, E., Eds.; Springer Vienna: Vienna, Austria, 2000; pp. 211–223. [Google Scholar]
- Roulston, A.H.; Cane, J.H.; Buchmann, S.L. What Governs Protein Content of Pollen: Pollinator Preferences, Pollen-Pistil Interactions, or Phylogeny? Ecol. Monogr. 2000, 70, 617–643. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Tooker, J.F.; Patch, H.M.; Biddinger, D.J.; Coccia, M.; Crone, M.K.; Fiely, M.; Francis, J.S.; Hines, H.M.; Hodges, M.; et al. Pollen Protein: Lipid Macronutrient Ratios May Guide Broad Patterns of Bee Species Floral Preferences. Insects 2020, 11, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.A.; Arceo-Gomez, G. Pollen Transport Networks Reveal Highly Diverse and Temporally Stable Plant-Pollinator Interactions in an Appalachian Floral Community. AoB Plants 2021, 13, plab062. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless Honey Bee Abundance. Science (1979) 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Winfree, R.; Williams, N.M.; Dushoff, J.; Kremen, C. Native Bees Provide Insurance against Ongoing Honey Bee Losses. Ecol. Lett. 2007, 10, 1105–1113. [Google Scholar] [CrossRef]
- Cusser, S.; Haddad, N.M.; Jha, S. Unexpected Functional Complementarity from Non-Bee Pollinators Enhances Cotton Yield. Agric. Ecosyst. Environ. 2021, 314, 107415. [Google Scholar] [CrossRef]
- Redhead, J.W.; Woodcock, B.A.; Pocock, M.J.O.; Pywell, R.F.; Vanbergen, A.J.; Oliver, T.H. Potential Landscape-Scale Pollinator Networks across Great Britain: Structure, Stability and Influence of Agricultural Land Cover. Ecol. Lett. 2018, 21, 1821–1832. [Google Scholar] [CrossRef] [Green Version]
- Cusser, S.; Goodell, K. Diversity and Distribution of Floral Resources Influence the Restoration of Plant-Pollinator Networks on a Reclaimed Strip Mine. Restor. Ecol. 2013, 21, 713–721. [Google Scholar] [CrossRef]
- Russo, L.; Shea, K. Deliberately Increased Network Connectance in a Plant-Pollinator Community Experiment. J. Complex Netw. 2017, 5, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, A.D.; Ruppel, R.; Jha, S. Generalist Behavior Describes Pollen Foraging for Perceived Oligolectic and Polylectic Bees. Environ. Entomol. 2016, 45, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhang, M. Exploring the Patterns and Drivers of Urban Expansion in the Texas Triangle Megaregion. Land 2021, 10, 1244. [Google Scholar] [CrossRef]
- Day, F.A.; Vaughan, J.W. The Evolving Landscape of the Austin-San Antonio Corridor. In Landscapes, Identities, and Development; Roca, Z., Claval, P., Agnew, J., Eds.; Routledge: New York, NY, USA, 2011; pp. 275–285. [Google Scholar]
- Frey, W.H. Population Growth in Metro America since 1980: Putting the Volatile 2000s in Perspective; Metropolitan Policy Program at Brookings: Washington, DC, USA, 2012. [Google Scholar]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee Foraging Ranges and Their Relationship to Body Size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef]
- Birdshire, K.R.; Carper, A.L.; Briles, C.E. Bee Community Response to Local and Landscape Factors along an Urban-Rural Gradient. Urban. Ecosyst. 2020, 23, 689–702. [Google Scholar] [CrossRef] [Green Version]
- Homer, C.J.; Fry, A.; Barnes, C.A. The National Land Cover Database. US Geol. Surv. Fact Sheet 2012, 3020, 1–4. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System Open Source Geospatial Foundation Project. 2014. Available online: https://qgis.org/en/site/ (accessed on 23 September 2022).
- Quistberg, R.D.; Bichier, P.; Philpott, S.M. Landscape and Local Correlates of Bee Abundance and Species Richness in Urban Gardens. Environ. Entomol. 2016, 45, 592–601. [Google Scholar] [CrossRef]
- Lane, I.G.; Portman, Z.M.; Herron-Sweet, C.H.; Pardee, G.L.; Cariveau, D.P. Differences in Bee Community Composition between Restored and Remnant Prairies Are More Strongly Linked to Forb Community Differences than Landscape Differences. J. Appl. Ecol. 2022, 59, 129–140. [Google Scholar] [CrossRef]
- de Manincor, N.; Hautekèete, N.; Mazoyer, C.; Moreau, P.; Piquot, Y.; Schatz, B.; Schmitt, E.; Zélazny, M.; Massol, F. How Biased Is Our Perception of Plant-Pollinator Networks? A Comparison of Visit- and Pollen-Based Representations of the Same Networks. Acta Oecologica 2020, 105, 103551. [Google Scholar] [CrossRef]
- Tur, C.; Vigalondo, B.; Trøjelsgaard, K.; Olesen, J.M.; Traveset, A. Downscaling Pollen-Transport Networks to the Level of Individuals. J. Anim. Ecol. 2014, 83, 306–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alarcón, R. Congruence between Visitation and Pollen-Transport Networks in a California Plant-Pollinator Community. Oikos 2010, 119, 35–44. [Google Scholar] [CrossRef]
- Urbanowicz, C.; Virginia, R.A.; Irwin, R.E. The Response of Pollen-Transport Networks to Landscape-Scale Climate Variation. Polar. Biol. 2017, 40, 2253–2263. [Google Scholar] [CrossRef]
- O’Connell, M.; Jordan, Z.; McGilvray, E.; Cohen, H.; Liere, H.; Lin, B.B.; Philpott, S.M.; Jha, S. Reap What You Sow: Local Plant Composition Mediates Bumblebee Foraging Patterns within Urban Garden Landscapes. Urban. Ecosyst. 2021, 24, 391–404. [Google Scholar] [CrossRef]
- Williams, N.M.; Kremen, C. Resource Distributions among Habitats Determine Solitary Bee Offspring Production in a Mosaic Landscape. Ecol. Appl. 2007, 17, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Joosten, H.; de Klerk, P. What’s in a Name? Some Thoughts on Pollen Classification, Identification, and Nomenclature in Quaternary Palynology. Rev. Paleobot. Palynol. 2002, 122, 29–45. [Google Scholar] [CrossRef]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring Specialization in Species Interaction Networks. BMC Ecol. 2006, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The Modularity of Pollination Networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef] [Green Version]
- Jordano, P. Patterns of Mutualistic Interactions in Pollination and Seed Dispersal: Connectance, Dependence Assymetries, and Coevolution. Am. Nat. 1987, 129, 657–677. [Google Scholar] [CrossRef]
- van Altena, C.; Hemerik, L.; de Ruiter, P.C. Food Web Stability and Weighted Connectance: The Complexity-Stability Debate Revisited. Theor. Ecol. 2016, 9, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Staniczenko, P.P.A.; Kopp, J.C.; Allesina, S. The Ghost of Nestedness in Ecological Networks. Nat. Commun. 2013, 4, 13191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormann, C.F.; Fründ, J.; Blüthgen, N.; Gruber, B. Indices, Graphs and Null Models: Analyzing Bipartite Ecological Networks. Open Ecol. J. 2009, 2, 7–24. [Google Scholar] [CrossRef]
- Ulrich, W.; Almeida-Neto, M.; Gotelli, N.J. A Consumer’s Guide to Nestedness Analysis. Oikos 2009, 118, 3–17. [Google Scholar] [CrossRef]
- Novella-Fernandez, R.; Rodrigo, A.; Arnan, X.; Bosch, J. Interaction Strength in Plant-Pollinator Networks: Are We Using the Right Measure? PLoS ONE 2019, 14, e0225930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–47. [Google Scholar] [CrossRef]
- Cohen, H.; Philpott, S.M.; Liere, H.; Lin, B.B.; Jha, S. The Relationship between Pollinator Community and Pollination Services Is Mediated by Floral Abundance in Urban Landscapes. Urban. Ecosyst. 2021, 24, 275–290. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S.; Adler, D.; Bates, D.; Baud-Bovy, G.; Ellison, S. Package “Car”. 2016. Available online: https://cran.r-project.org/web/packages/car/index.html (accessed on 23 September 2022).
- Barton, K. Package “MuMin”. 2009. Available online: https://cran.r-project.org/web/packages/MuMIn/index.html (accessed on 23 September 2022).
- Grueber, C.E.; Nakagawa, S.; Laws, R.J.; Jamieson, I.G. Multimodel Inference in Ecology and Evolution: Challenges and Solutions. J. Evol. Biol. 2011, 24, 699–711. [Google Scholar] [CrossRef]
- Cane, J.H.; Sipes, S. Floral Specialization by Bees: Analytical Methods and a Revised Lexicon for Oligolecty. In Plant-Pollinator Interactions: From Specialization to Generalization; Waser, N., Ollerton, J., Eds.; University of Chicago Press: Chicago, IL, USA, 2006; pp. 99–112. [Google Scholar]
- Minckley, R.L.; Roulston, T.H. Incidental Mutualisms and Pollen Specialization among Bees. In Plant-Pollinator Interactions: From Specialization to Generalization; Waser, N.M., Ollerton, J., Eds.; Chicago Press: Chicago, IL, USA, 2006; pp. 69–98. [Google Scholar]
- Davis, E.S.; Reid, N.; Paxton, R.J. Quantifying Forage Specialisation in Polyphagic Insects: The Polylectic and Rare Solitary Bee, Colletes Floralis (Hymenoptera: Colletidae). Insect Conserv. Divers. 2012, 5, 289–297. [Google Scholar] [CrossRef]
- Jha, S.; Stefanovich, L.; Kremen, C. Bumble Bee Pollen Use and Preference across Spatial Scales in Human-Altered Landscapes. Ecol. Entomol. 2013, 38, 570–579. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.H. The Comparison of Usage and Availability Measurements for Evaluating Resource Preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- Calenge, C. The Package “Adehabitat” for the R Software: A Tool for the Analysis of Space and Habitat Use by Animals. Ecol. Modell. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Blüthgen, N. Why Network Analysis Is Often Disconnected from Community Ecology: A Critique and an Ecologist’s Guide. Basic. Appl. Ecol. 2010, 11, 185–195. [Google Scholar] [CrossRef]
- Morozumi, C.; Loy, X.; Reynolds, V.; Schiffer, A.; Morrison, B.; Savage, J.; Brosi, B. Simultaneous Niche Expansion and Contraction in Plant–Pollinator Networks under Drought. Oikos 2022, e09265. [Google Scholar] [CrossRef]
- Gómez-Martínez, C.; González-Estévez, M.A.; Cursach, J.; Lázaro, A. Pollinator Richness, Pollination Networks, and Diet Adjustment along Local and Landscape Gradients of Resource Diversity. Ecol. Appl. 2022, 32, e2634. [Google Scholar] [CrossRef] [PubMed]
- Frund, J.; Dormann, C.F.; Holzschuh, A.; Tscharntke, T. Bee Diversity Effects on Pollination Depend on Functional Complementarity and Niche Shifts. Ecology 2013, 94, 2042–2054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doré, M.; Fontaine, C.; Thébault, E. Relative Effects of Anthropogenic Pressures, Climate, and Sampling Design on the Structure of Pollination Networks at the Global Scale. Glob. Chang. Biol. 2021, 27, 1266–1280. [Google Scholar] [CrossRef] [PubMed]
- Bolin, A.; Smith, H.G.; Lonsdorf, E.V.; Olsson, O. Scale-Dependent Foraging Tradeoff Allows Competitive Coexistence. Oikos 2018, 127, 1575–1585. [Google Scholar] [CrossRef] [Green Version]
- Hopfenmüller, S.; Steffan-Dewenter, I.; Holzschuh, A. Trait-Specific Responses of Wild Bee Communities to Landscape Composition, Configuration and Local Factors. PLoS ONE 2014, 9, e104439. [Google Scholar] [CrossRef] [Green Version]
- Mallinger, R.E.; Gibbs, J.; Gratton, C. Diverse Landscapes Have a Higher Abundance and Species Richness of Spring Wild Bees by Providing Complementary Floral Resources over Bees’ Foraging Periods. Landsc. Ecol. 2016, 31, 1523–1535. [Google Scholar] [CrossRef]
- Rohr, R.P.; Saavedra, S.; Bascompte, J. On the Structural Stability of Mutualistic Systems. Science (1979) 2014, 345, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Thébault, E.; Fontaine, C. Stability of Ecological Communities and the Architecture of Mutualistic and Trophic Networks. Science (1979) 2010, 329, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Senapathi, D.; Goddard, M.A.; Kunin, W.E.; Baldock, K.C.R. Landscape Impacts on Pollinator Communities in Temperate Systems: Evidence and Knowledge Gaps. Funct. Ecol. 2017, 31, 26–37. [Google Scholar] [CrossRef]
- Griffin, S.R.; Bruninga-Socolar, B.; Gibbs, J. Bee Communities in Restored Prairies Are Structured by Landscape and Management, Not Local Floral Resources. Basic. Appl. Ecol. 2021, 50, 144–154. [Google Scholar] [CrossRef]
- Potts, S.G.; Vulliamy, B.; Roberts, S.; O’Toole, C.; Dafni, A.; Ne’eman, G.; Willmer, P. Role of Nesting Resources in Organising Diverse Bee Communities in a Mediterranean Landscape. Ecol. Entomol. 2005, 30, 78–85. [Google Scholar] [CrossRef]
- Baumgartner, M.T. Connectance and Nestedness as Stabilizing Factors in Response to Pulse Disturbances in Adaptive Antagonistic Networks. J. Theor. Biol. 2020, 486, 110073. [Google Scholar] [CrossRef]
- Potter, L.B.; Hoque, N. Texas Population Projections 2010–2050; Office of the State Demographer, 2014. Available online: https://demographics.texas.gov/Resources/publications/2019/20190128_PopProjectionsBrief.pdf (accessed on 23 September 2022).
- Dutta, D.; Rahman, A.; Paul, S.K.; Kundu, A. Impervious Surface Growth and Its Inter-Relationship with Vegetation Cover and Land Surface Temperature in Peri-Urban Areas of Delhi. Urban Clim. 2021, 37, 100799. [Google Scholar] [CrossRef]
- Morandin, L.A.; Kremen, C. Hedgerow Restoration Promotes Pollinator Populations and Exports Native Bees to Adjacent Fields. Ecol. Appl. 2013, 23, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Forup, M.L.; Henson, K.S.E.; Craze, P.G.; Memmott, J. The Restoration of Ecological Interactions: Plant-Pollinator Networks on Ancient and Restored Heathlands. J. Appl. Ecol. 2008, 45, 742–752. [Google Scholar] [CrossRef] [Green Version]
- Layek, U.; Manna, S.S.; Karmakar, P. Pollen Foraging Behaviour of Honey Bee (Apis mellifera L.) in Southern West Bengal, India. Palynology 2020, 44, 114–126. [Google Scholar] [CrossRef]
- Oleques, S.S.; Vizentin-Bugoni, J.; Overbeck, G.E. Influence of Grazing Intensity on Patterns and Structuring Processes in Plant–Pollinator Networks in a Subtropical Grassland. Arthropod Plant Interact. 2019, 13, 757–770. [Google Scholar] [CrossRef]
- Mayer, C.; Soka, G.; Picker, M. The Importance of Monkey Beetle (Scarabaeidae: Hopliini) Pollination for Aizoaceae and Asteraceae in Grazed and Ungrazed Areas at Paulshoek, Succulent Karoo, South Africa. J. Insect. Conserv. 2006, 10, 323–333. [Google Scholar] [CrossRef]
- van Klink, R.; Nolte, S.; Mandema, F.S.; Lagendijk, D.D.G.; WallisDeVries, M.F.; Bakker, J.P.; Esselink, P.; Smit, C. Effects of Grazing Management on Biodiversity across Trophic Levels–The Importance of Livestock Species and Stocking Density in Salt Marshes. Agric. Ecosyst. Environ. 2016, 235, 329–339. [Google Scholar] [CrossRef]
- Torres, C.; Galetto, L. Are Nectar Sugar Composition and Corolla Tube Length Related to the Diversity of Insects That Visit Asteraceae Flowers? Plant Biol. 2002, 4, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Neff, J.L. Components of Nest Provisioning Behavior in Solitary Bees (Hymenoptera: Apoidea). Apidologie 2008, 39, 30–45. [Google Scholar] [CrossRef] [Green Version]
- Ivey, C.T.; Martinez, P.; Wyatt, R. Variation in Pollinator Effectiveness in Swamp Milkweed, Asclepias Incarnata (Apocynaceae). Am. J. Bot. 2003, 90, 214–225. [Google Scholar] [CrossRef] [Green Version]
- Warzecha, D.; Diekötter, T.; Wolters, V.; Jauker, F. Attractiveness of Wildflower Mixtures for Wild Bees and Hoverflies Depends on Some Key Plant Species. Insect Conserv. Divers 2018, 11, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Brosi, B.J.; Briggs, H.M. Single Pollinator Species Losses Reduce Floral Fidelity and Plant Reproductive Function. Proc. Natl. Acad. Sci. USA 2013, 110, 13044–13048. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.M.; Brittain, C.; Hendrix, S.D.; Thorp, R.; Williams, N.; Kremen, C. Wild Pollination Services to California Almond Rely on Semi-Natural Habitat. J. Appl. Ecol. 2012, 49, 723–732. [Google Scholar] [CrossRef]
- Vaca-Uribe, J.L.; Figueroa, L.L.; Santamaría, M.; Poveda, K. Plant Richness and Blooming Cover Affect Abundance of Flower Visitors and Network Structure in Colombian Orchards. Agric. Entomol. 2021, 23, 545–556. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Response | Estimate | Std. Error | Z Value | p Value | R2 Value |
|---|---|---|---|---|---|
| H2’ | |||||
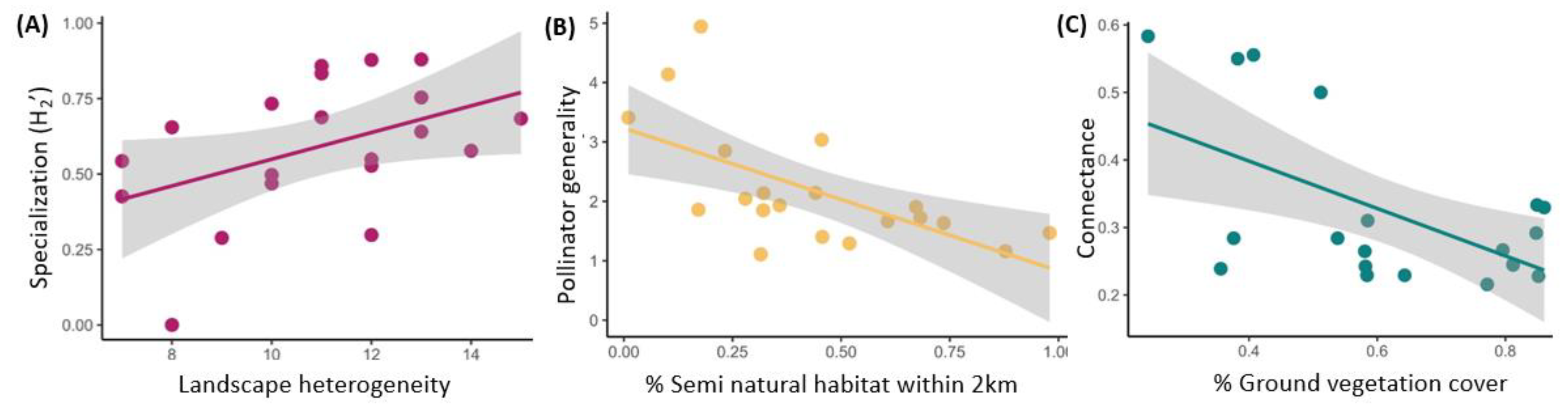
| Landscape heterogeneity | 0.1003 | 0.044 | 2.072 | 0.038 | -- |
| R2GLMM (m) | -- | -- | -- | -- | 0.262 |
| Connectance | |||||
| % Ground vegetation | −0.071 | 0.023 | 2.780 | 0.005 | -- |
| % Bare ground | −0.025 | 0.024 | 0.968 | 0.333 | -- |
| R2GLMM (m) | -- | -- | -- | -- | 0.421 |
| Nestedness | |||||
| % Bare ground | −0.018 | 0.042 | 0.414 | 0.679 | -- |
| Landscape heterogeneity | −0.029 | 0.052 | 0.544 | 0.586 | -- |
| R2GLMM (m) | -- | -- | -- | -- | 0.208 |
| Generality | |||||
| % Semi-natural habitat | −0.607 | 0.171 | 3.270 | 0.001 | -- |
| Landscape heterogeneity | −0.157 | 0.197 | 0.203 | 0.437 | -- |
| R2GLMM (m) | -- | -- | -- | -- | 0.528 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pardee, G.L.; Ballare, K.M.; Neff, J.L.; Do, L.Q.; Ojeda, D.; Bienenstock, E.J.; Brosi, B.J.; Grubesic, T.H.; Miller, J.A.; Tong, D.; et al. Local and Landscape Factors Influence Plant-Pollinator Networks and Bee Foraging Behavior across an Urban Corridor. Land 2023, 12, 362. https://doi.org/10.3390/land12020362
Pardee GL, Ballare KM, Neff JL, Do LQ, Ojeda D, Bienenstock EJ, Brosi BJ, Grubesic TH, Miller JA, Tong D, et al. Local and Landscape Factors Influence Plant-Pollinator Networks and Bee Foraging Behavior across an Urban Corridor. Land. 2023; 12(2):362. https://doi.org/10.3390/land12020362
Chicago/Turabian StylePardee, Gabriella L., Kimberly M. Ballare, John L. Neff, Lauren Q. Do, DianaJoyce Ojeda, Elisa J. Bienenstock, Berry J. Brosi, Tony H. Grubesic, Jennifer A. Miller, Daoqin Tong, and et al. 2023. "Local and Landscape Factors Influence Plant-Pollinator Networks and Bee Foraging Behavior across an Urban Corridor" Land 12, no. 2: 362. https://doi.org/10.3390/land12020362