
Responses of Picea meyeri to Climatic Factors Revealed by Tree Ring Isotopes and Water Use Efficiency on Luya Mountain of North-Central China
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
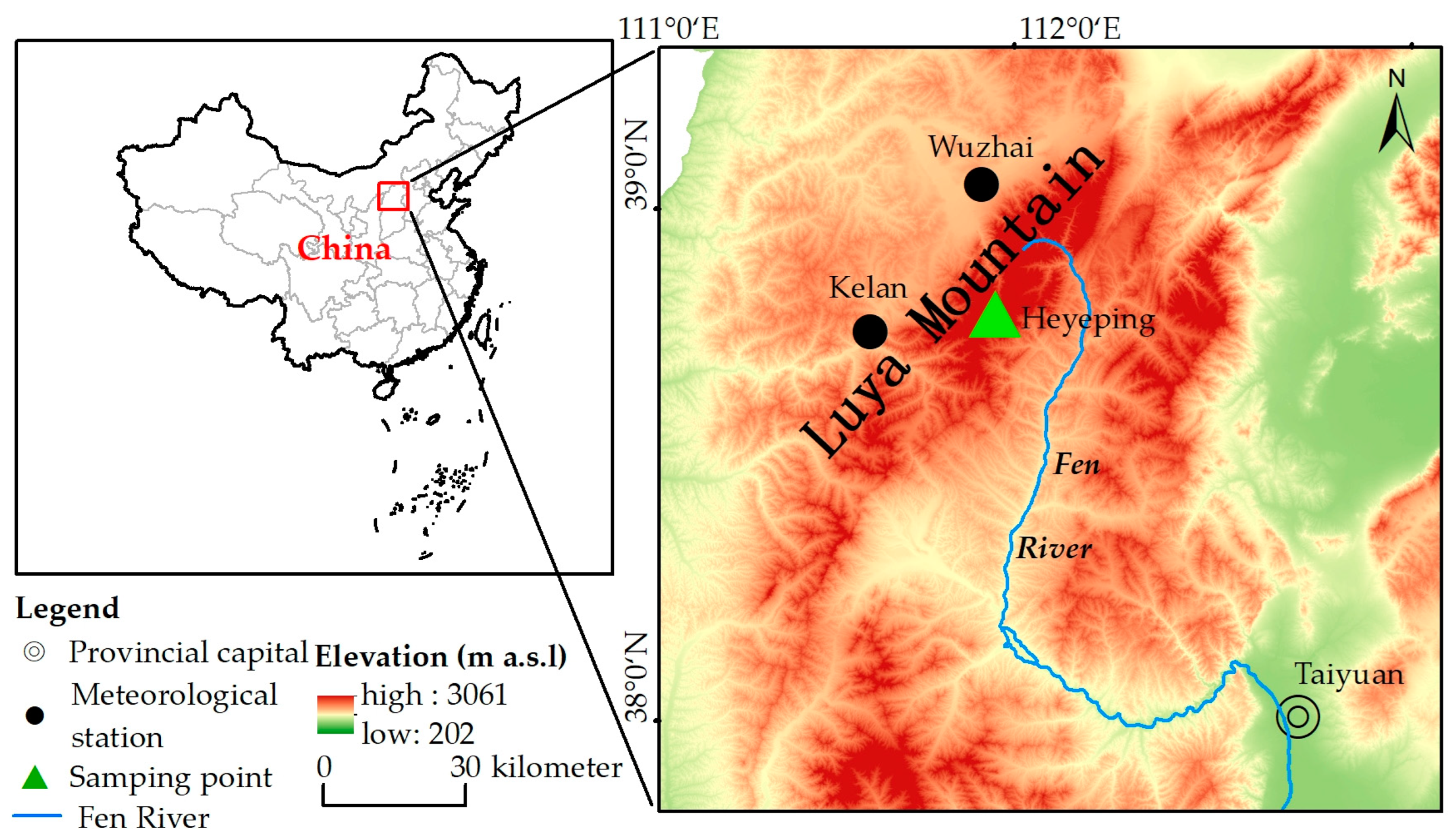
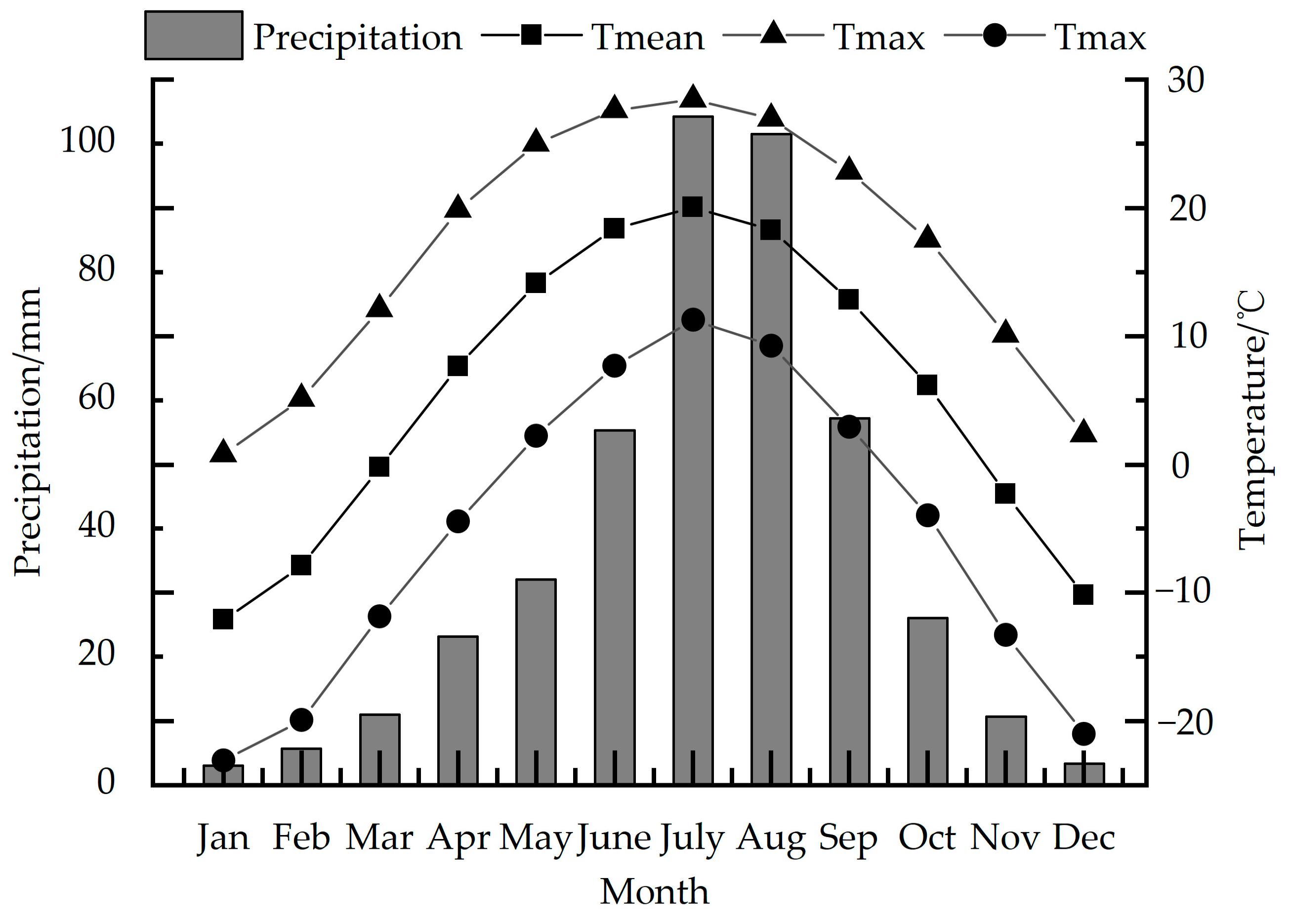
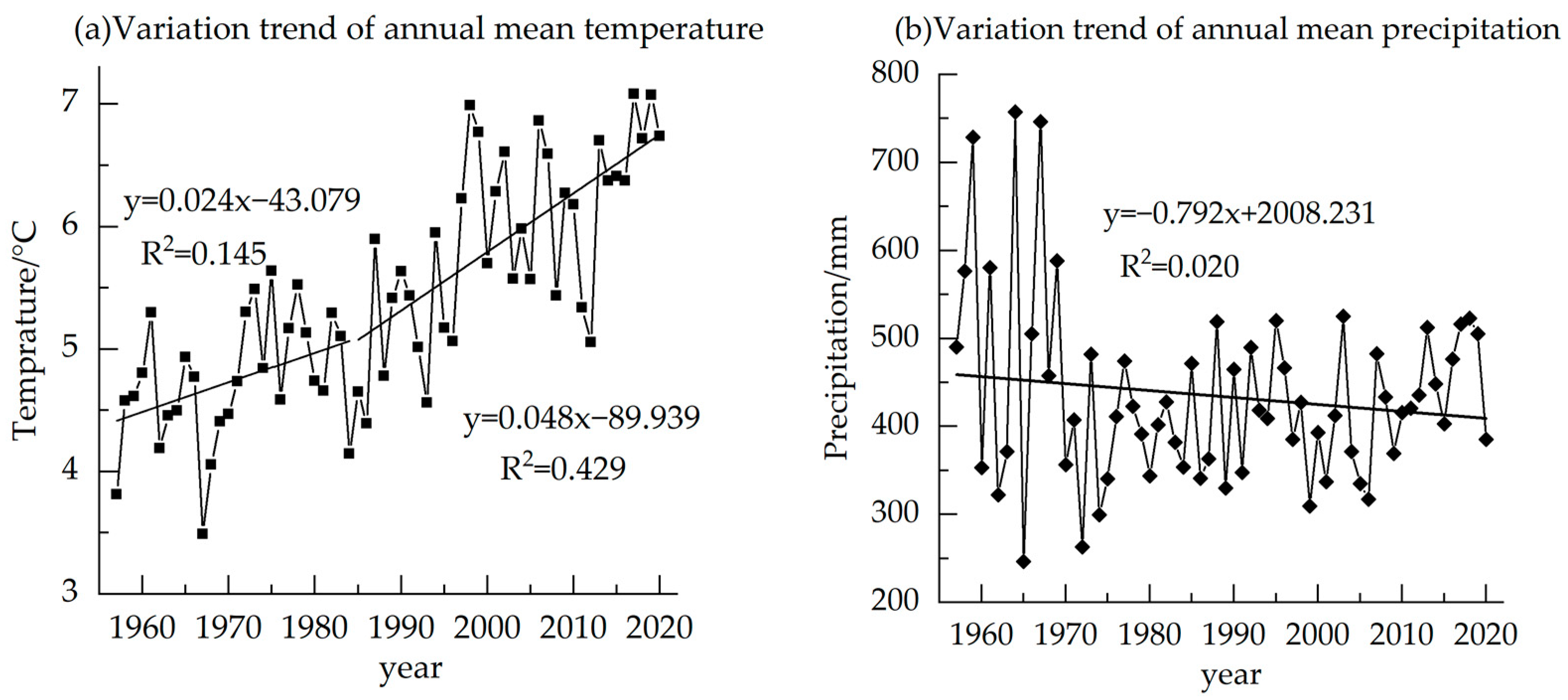
2.1. Overview of the Study Area
2.2. Sample Collection and Treatment
2.3. Calculation of WUE
2.4. Data Collection and Processing
3. Results
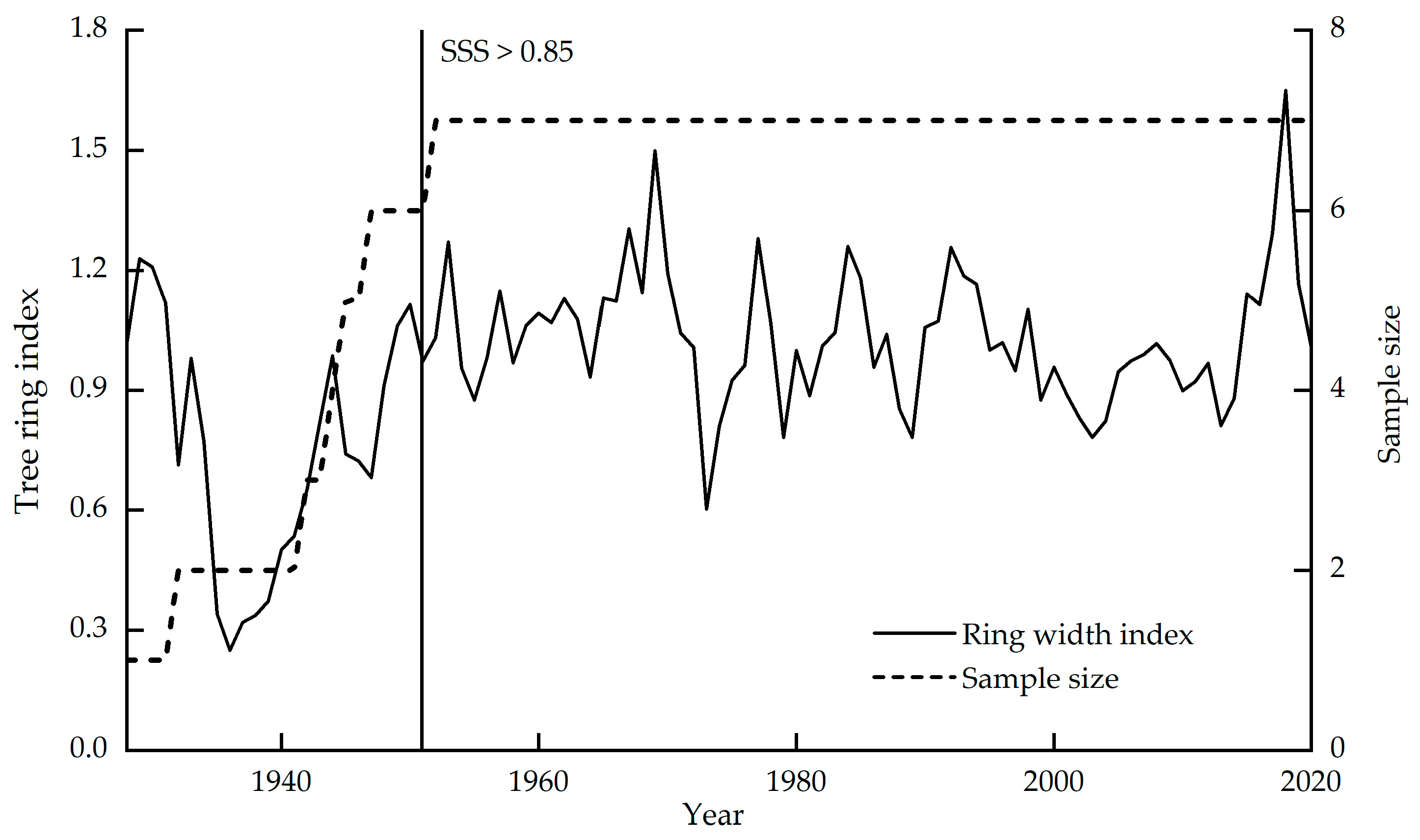
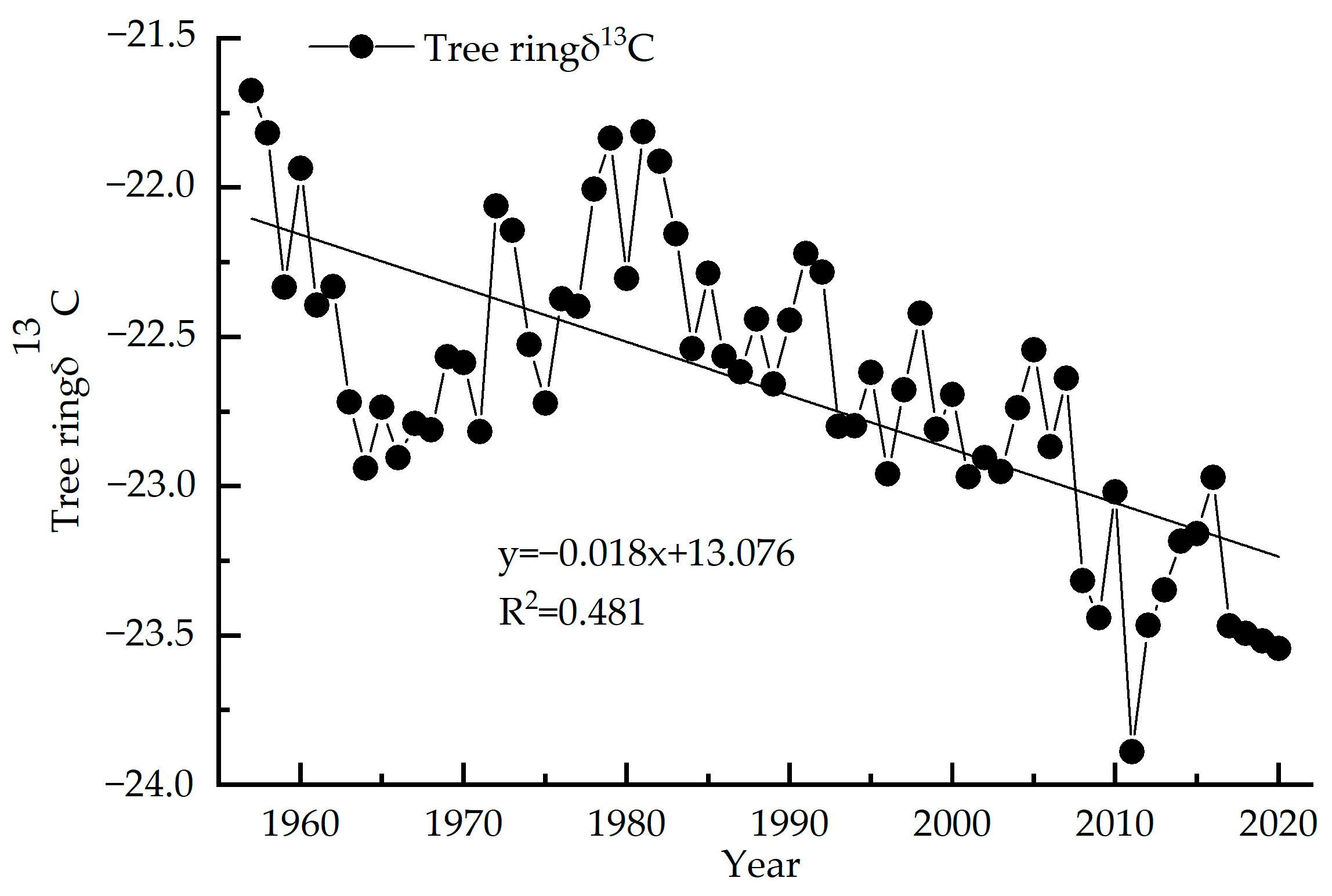
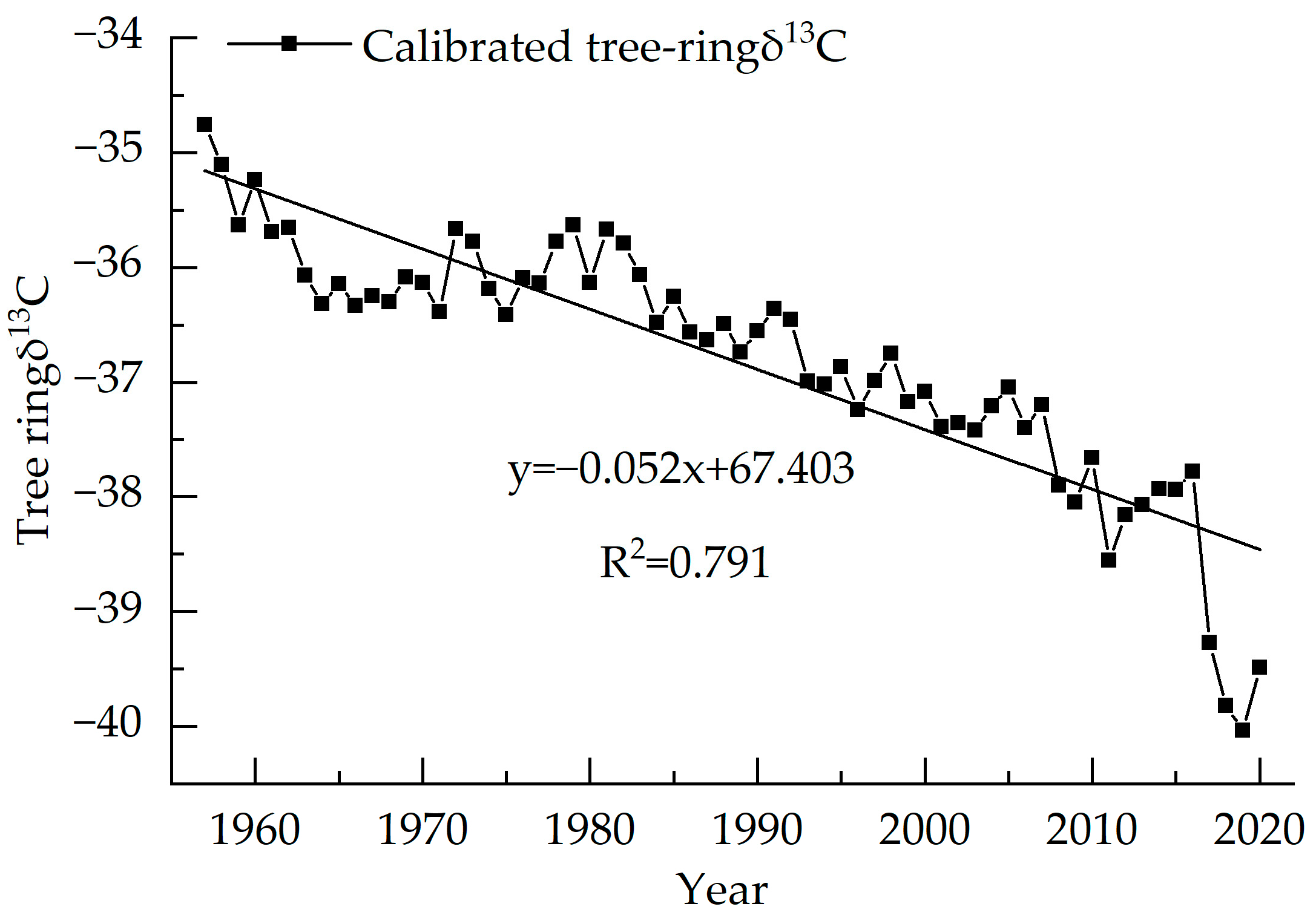
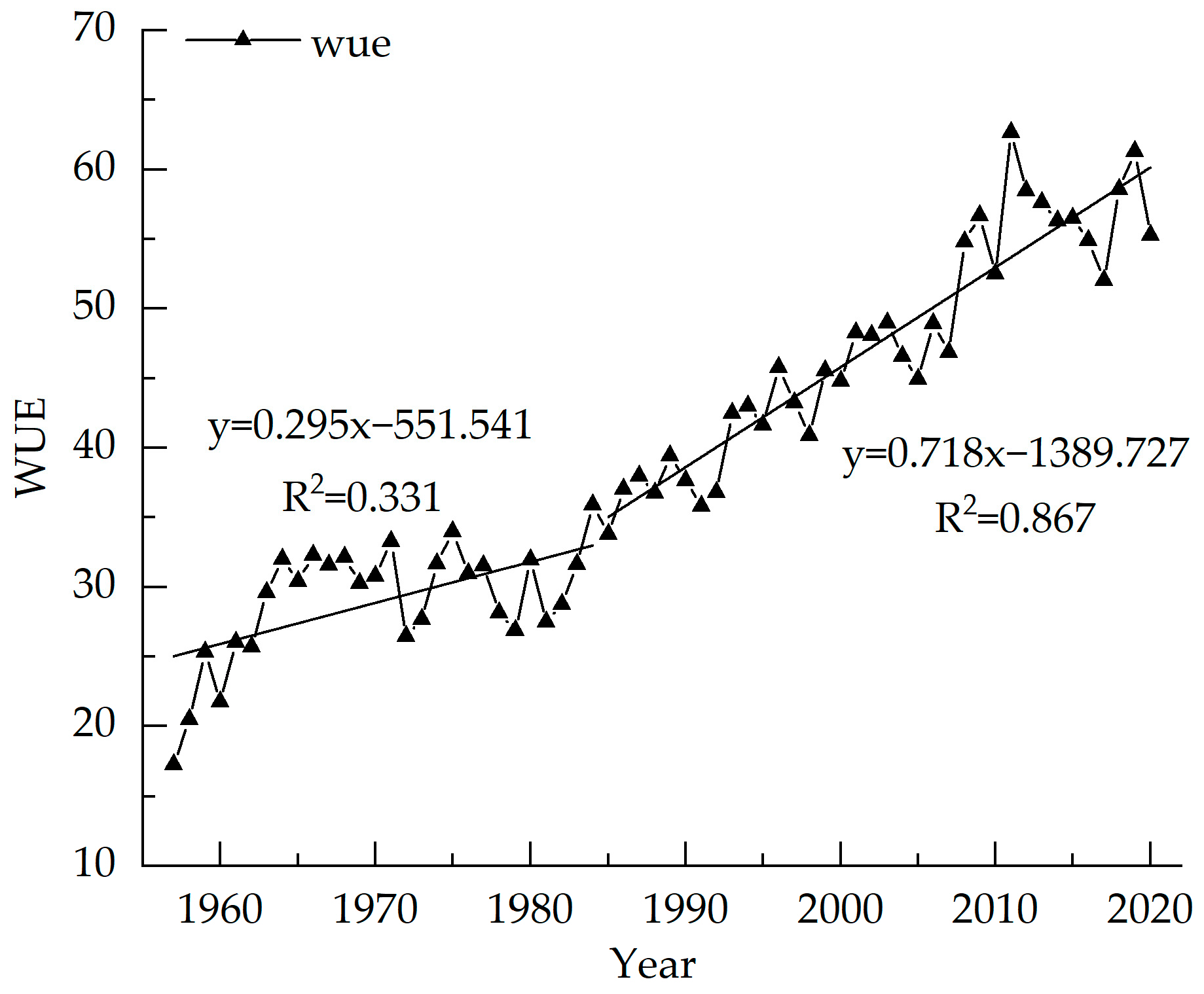
3.1. Picea Meyeri δ13C Value and WUE Sequence Characteristics
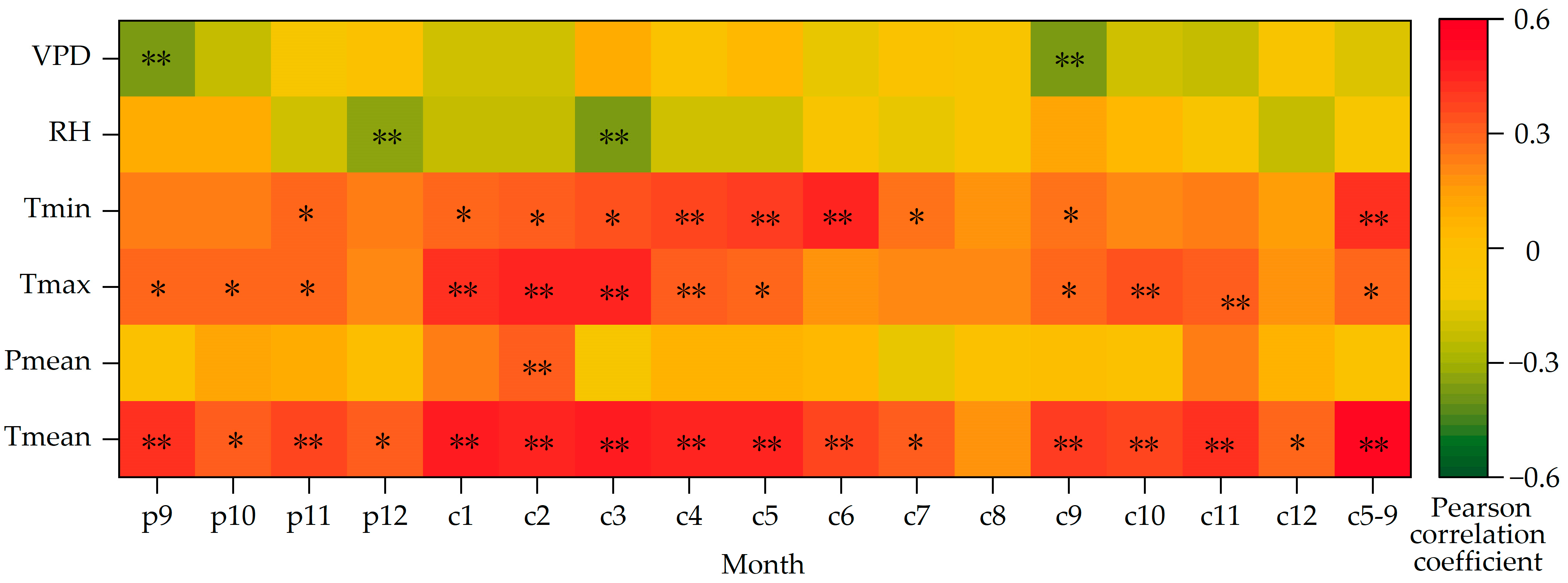
3.2. Response of Stable Carbon Isotope Sequence to Changes in Temperature and Precipitation
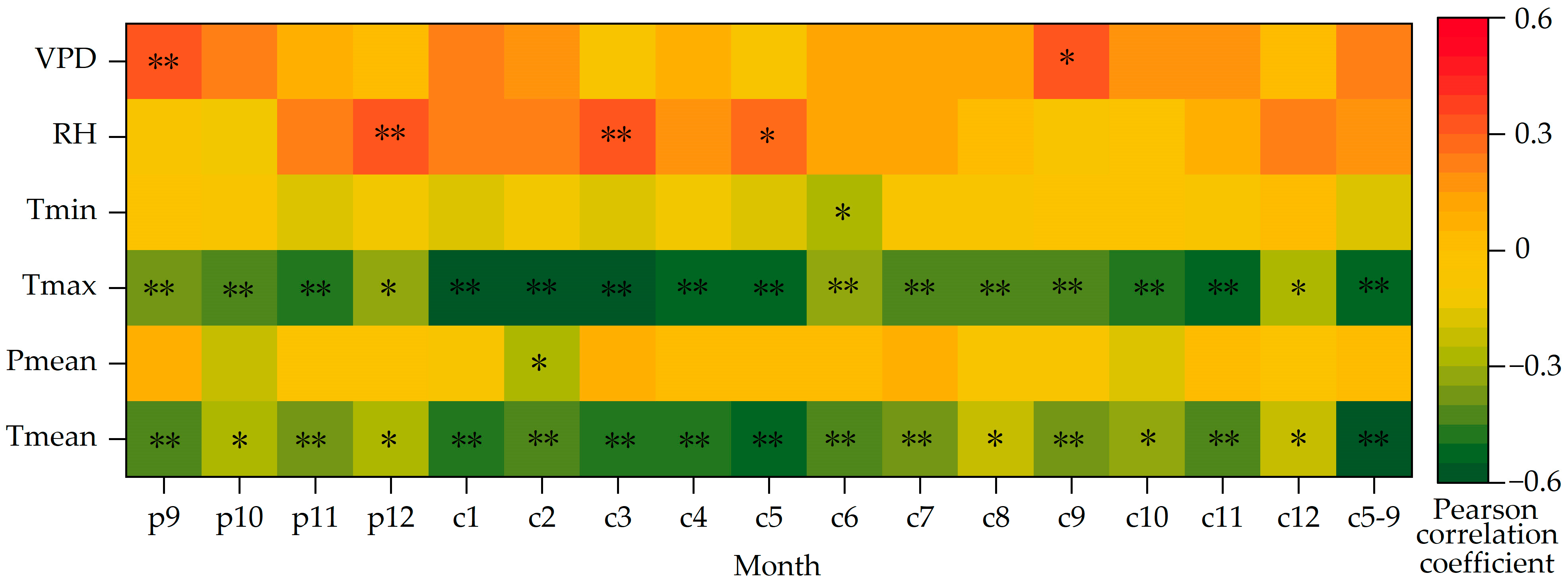
3.3. Relationship between δ13C and Climate Factors
3.4. Response of WUE to Climate Change
4. Discussion
4.1. Analysis of the Main Climate Impact Factors of δ13C of Picea meyeri
4.2. Analysis of the Main Climate Impact Factors of WUE of Picea meyeri
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Yang, Y.; Zhang, W.; Ren, F.; Wang, G.; Dong, M. Stem radius growth features of Picea meyeri and its relationship with environmental factors at the treeline of Luya Mountain. Acta Ecol. Sin. 2009, 29, 6793–6804. (In Chinese) [Google Scholar] [CrossRef]
- Liu, X.; Xu, G.; Wang, W.; Wu, G.; Zeng, X.; An, W.; Zhang, X. Tree-ring stable isotopes proxies: Progress problems and prospects. Quat. Sci. 2015, 35, 1245–1260. (In Chinese) [Google Scholar] [CrossRef]
- Chen, T.; Qin, D.; Li, J.; Ren, J.; Sun, W. Response of CO2 concentration parameters and water-use efficiency derived from tree-ring δ13C series to atmospheric CO2 increase. J. Glaciol. Geocryol. 2001, 23, 41–45. (In Chinese) [Google Scholar] [CrossRef]
- Chen, T.; Qin, D.; He, Y.; Li, J.; Liu, X.; Ren, J. Feasibility study on extracting atmospheric carbon information from tree-ring carbon sequence. Mar. Geol. Quat. Geol. 2002, 22, 79–83. (In Chinese) [Google Scholar] [CrossRef]
- Chen, T.; Chen, F.; An, L.; Liu, X. Variations of tree-ring and foliar δ13C values of Sabina przewalskii with altitude. J. Glaciol. Geocryol. 2004, 26, 767–771. (In Chinese) [Google Scholar] [CrossRef]
- Qin, L.; Shang, H.; Zhang, T.; Liu, W.; Zhang, R. Response comparison of the tree-ring δ13C to climate on the southern and northern slopes of Tianshan Mountains. Acta Ecol. Sin. 2021, 41, 5713–5724. (In Chinese) [Google Scholar] [CrossRef]
- Jiang, Y.; Yang, Y.; Dong, M.; Zhang, W.; Ren, F. Comparison of radial growth characteristics of Picea meyeri and Larix principis-rupprechtii in Luya mountain. Chin. J. Appl. Ecol. 2009, 20, 1271–1277. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, Y.; Wang, M.; Zhang, L.; Dong, M.; Guo, Y. The response of radial growth of Picea meyeri with different altitudes on the sunny slope of Luya Mountain to climate warming. Chin. J. Plant Ecol. 2013, 37, 1142–1152. (In Chinese) [Google Scholar] [CrossRef]
- Wang, J.; Li, S.; Guo, Y.; Yang, Q.; Ren, R.; Han, Y. Responses of Larix principis-rupprechtii Radial Growth to Climatic Factors at Different Elevations on Guancen Mountain, North-Central China. Forests 2022, 13, 99. [Google Scholar] [CrossRef]
- Guo, Y.; Li, S.; Wang, J.; Han, Y. Response divergence of radial growth to climate change in earlywood and latewood of Larix principis-rupprechtii in Luya Mountain. Arid. Zone Res. 2022, 39, 1449–1463. (In Chinese) [Google Scholar] [CrossRef]
- Wilson, A.T.; Grinsted, M.J. 12C/13C in cellulose and lignin as palaeothermometers. Nature 1977, 265, 133–135. [Google Scholar] [CrossRef]
- Mccarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef]
- Borella, S.; Leuenberger, M.; Saurer, M.; Siegwolf, R. Reducing uncertainties in δ13C analysis of tree rings: Pooling, milling, and cellulose extraction. J. Geophys. Res. 1998, 103, 19519–19526. [Google Scholar] [CrossRef]
- Ma, L.; Liu, Y.; Zhao, J. Relationship between stable carbon isotope content and environment in tree rings of Pinus Tabulaeformis in Helan Mountain. Environ. Sci. 2003, 24, 49–53. (In Chinese) [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, J.; Shang, Z. Sensitivity of tree ring stable carbon isotopes to climate change: Research progress and review. Chin. J. Ecol. 2014, 33, 2538–2547. (In Chinese) [Google Scholar] [CrossRef]
- Wu, X. Tree Rings and Climate Change; Beijing Meteorological Press: Beijing, China, 1990. (In Chinese) [Google Scholar]
- Macfarlane, C.; Warren, C.R.; White, D.A.; Adams, M.A. A rapid and simple method for processing wood to crude cellulose for analysis of stable carbon isotopes in tree rings. Tree Physiol. 1999, 19, 831–835. [Google Scholar] [CrossRef]
- Freyer, H.D.; Belacy, N. 13C/12C records in northern hemispheric trees during the past 500 years—Anthropogenic impact and climatic superpositions. J. Geophys. Res. Ocean. 1983, 88, 6844–6852. [Google Scholar] [CrossRef]
- Young, G.H.F.; Bale, R.J.; Loader, N.J.; Mccarroll, D.; Nayling, N.; Vousden, N. Central England temperature since AD 1850: The potential of stable carbon isotopes in British oak trees to reconstruct past summer temperatures. J. Quat. Sci. 2012, 27, 606–614. [Google Scholar] [CrossRef]
- Li, Z.; Liu, R.; An, Z.; Liu, Y.; Wu, X. Tree-ring evidence for stable carbon isotopes from increasing atmospheric CO2 concentration since the Industrial Revolution. Chin. Sci. Bull. 1994, 39, 2172–2174. (In Chinese) [Google Scholar]
- Xu, H.; Hong, Y.; Zhu, Y.; Liu, G. Information of low cloud amount recorded in δ13C series of tree ring cellulose of Pinus Koraiensis in Antu Area. Geochimica 2002, 31, 309–314. (In Chinese) [Google Scholar] [CrossRef]
- Liu, X.; Zhao, L.; Voelker, S.; Xu, G.; Zeng, X.; Zhang, X.; Zhang, L.; Sun, W.; Zhang, Q.; Wu, G.; et al. Warming and CO2 enrichment modified the ecophysiological responses of Dahurian larch and Mongolia pine during the past century in the permafrost of northeastern China. Tree Physiol. 2018, 39, 88–103. [Google Scholar] [CrossRef]
- Xu, G.; Liu, X.; Qin, D.; Chen, T.; An, W.; Wang, W.; Wu, G.; Zeng, X.; Ren, J. Climate warming and increasing atmospheric CO2 have contributed to increased intrinsic water-use efficiency on the northeastern Tibetan Plateau since 1850. Trees 2013, 27, 465–475. [Google Scholar] [CrossRef]
- Zhang, Y.; Pang, R.; Gu, F.; Liu, S. Temporal-spatial variations of WUE and its response to climate change in alpine area of southwestern China. Acta Ecol. Sin. 2016, 36, 1515–1525. (In Chinese) [Google Scholar] [CrossRef]
- Pieters, A.J.; Núñez, M. Photosynthesis, water use efficiency, and the δ13C in two rice genotypes with contrasting response to water deficit. Photosynthetica 2008, 46, 574–580. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Szejner, P. Intra-annual tree-ring isotope variations: Do they occur when environment remains constant? Trees 2022, 36, 865–868. [Google Scholar] [CrossRef]
- Pu, X.; Wang, X.; Lyu, L. Recent warming-induced tree growth enhancement at the Tibetan treeline and the link to improved water-use efficiency. Forests 2021, 12, 1702. [Google Scholar] [CrossRef]
- Quadri, P.; Silva, L.; Zavaleta, E.S. Climate-induced reversal of tree growth patterns at a tropical treeline. Sci. Adv. 2021, 7, eabb7572. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Feng, Q.; Si, J.; Chang, Z.; Cao, G.; Chen, K. Research on characteristics of water use efficiency variations of Populus euphratica during the growing season in extremely arid region of China. J. Desert Res. 2012, 32, 724–729. (In Chinese) [Google Scholar]
- Kannenberg, S.A.; Driscoll, A.W.; Szejner, P.; Anderegg, W.R.L.; Ehleringer, J.R. Rapid increases in shrubland and forest intrinsic water-use efficiency during an ongoing megadrought. Proc. Natl. Acad. Sci. USA 2021, 118, e2118052118. [Google Scholar] [CrossRef]
- Zhang, J. Vertical zones of vegetation in Luya Mountain in Shanxi Province. Sci. Geogr. Sin. 1989, 9, 346–353. (In Chinese) [Google Scholar] [CrossRef]
- Ma, Z. Shanxi Vegetation; Science and Technology Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Ge, Q. Climate Change in Past Dynasties of China; Science Press: Beijing, China, 2011. (In Chinese) [Google Scholar]
- Yi, L.; Liu, Y.; Song, H.; Li, Q.; Cai, Q.; Yang, Y.; Sun, J. Summer temperature variations since 1676 AD in Luya Mountain, Shanxi Province of China, inferred from tree rings. J. Glaciol. Geocryol. 2006, 28, 330–336. (In Chinese) [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–75. [Google Scholar]
- Kagawa, A.; Sano, M.; Nakatsuka, T.; Ikeda, T.; Kubo, S. An optimized method for stable isotope analysis of tree rings by extracting cellulose directly from cross-sectional laths. Chem. Geol. 2015, 393–394, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Green, J.W. Methods of Carbonhy Date Chemistry; Academic Press: New York, NY, USA, 1963. [Google Scholar]
- Liu, X.; Liu, Y.; Xu, G.; Cai, Q.; An, W.; Wang, W. Pretreatment of tree-ring samples for stable isotope analysis. J. Glaciol. Geocryol. 2010, 32, 1242–1250. (In Chinese) [Google Scholar]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and intercellular carbon dioxide concentration in leaves. Aust. J. Plant Physiol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, L.; Cai, Q.; An, Z.; Liu, W.; Gao, L. Reconstruction of summer (June–August) air temperature in Helan Mountain since 1890 using tree ring stable carbon isotopes. Sci. Sin. 2002, 32, 667–674. (In Chinese) [Google Scholar] [CrossRef]
- Hu, Z.; Yu, G.; Wang, Q.; Zhao, F. Ecosystem level water use efficiency: A review. Acta Ecol. Sin. 2009, 29, 1498–1507. (In Chinese) [Google Scholar] [CrossRef]
- Wang, J.; He, Q.; Shang, Z.; Qian, J.; Yang, H.; Zhao, X. Sensitivity of the pine tree δ13C on MT. Zijin at Nanjing to climatic change. Resour. Environ. Yangtze Basin 2012, 21, 1106–1111. (In Chinese) [Google Scholar]
- Sidorova, O.V.C.; Myglan, V.S.; Fonti, M.V.; Saurer, M. Isotopic responses to dry and wet episodes as captured in tree rings of southern Altai relict forests. Eur. J. For. Res. 2021, 140, 527–535. [Google Scholar] [CrossRef]
- Pu, X.; Wang, X.; Lyu, L. Tree-Ring isotopes provide clues for sink limitation on treeline formation on the Tibetan Plateau. Atmosphere 2021, 12, 540. [Google Scholar] [CrossRef]
- Pandey, S.; Cherubini, P.; Saurer, M.; Carrer, M.; Petit, G. Effects of climate change on treeline trees in Sagarmatha (Mt. Everest, Central Himalaya). J. Veg. Sci. 2020, 31, 1144–1153. [Google Scholar] [CrossRef]
- Zheng, Z. The Analysis of the Stable Carbon Isotope Composition of Tree Rings of Pinus Thunbergii and Climate Significance in Tashan Mountain, Shandong Province. Master’s Thesis, Shandong Normal University, Jinan, China, 2014. [Google Scholar]
- Liu, Y.; Wang, Y.; Li, Q.; Song, H.; Linderholm, H.W.; Leavitt, S.W.; Wang, R.; An, Z. Tree-ring stable carbon isotope-based May-July temperature reconstruction over Nanwutai, China, for the past century and its record of 20th century warming. Quat. Sci. Rev. 2014, 93, 67–76. [Google Scholar] [CrossRef]
- Zhou, J.; Meng, P.; Zhang, J.; Guan, C.; Sun, S. Differences in the response of radial growth and intrinsic water use efficiency of Robinia pseudoacacia to climatic factors in Minquan of He’nan province and Baishui of Shaanxi Province. For. Res. 2021, 34, 1–8. (In Chinese) [Google Scholar] [CrossRef]
- Treml, V.; Tumajer, J.; Jandová, K.; Oulehle, F.; Rydval, M.; Čada, V.; Treydte, K.; Mašek, J.; Vondrovicová, L.; Lhotáková, Z.; et al. Increasing water-use efficiency mediates effects of atmospheric carbon, sulfur, and nitrogen on growth variability of central European conifers. Sci. Total Environ. 2022, 838, 156483. [Google Scholar] [CrossRef]
- Manrique-Alba, À.; Beguería, S.; Tomas-Burguera, M.; Camarero, J.J. Increased post-drought growth after thinning in Pinus nigra plantations. Forests 2021, 12, 985. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, Y.; Dong, M.; Kang, M.; Yang, H. Relationship between the radial growth of Picea meyeri and climate along elevations of the Luyashan Mountain in North-Central China. For. Ecol. Manag. 2012, 265, 142–149. [Google Scholar] [CrossRef]
- Liu, N.; Liu, Y.; Zhou, Q.; Bao, G. Droughts and broad-scale climate variability reflected by temperature sensitive tree growth in the Qinling Mountains, central China. Int. J. Biometeorol. 2013, 57, 169–177. [Google Scholar] [CrossRef]
- Liang, E.; Shao, X.; Hu, Y.; Lin, J. Dendroclimatic evaluation of climate-growth relationships of Meyer spruce (Picea meyeri) on a sandy substrate in semi-arid grassland, north China. Trees 2001, 15, 230–235. [Google Scholar] [CrossRef]
- Liang, E.; Shao, X.; Kong, Z.; Lin, J. The extreme drought in the 1920s and its effect on tree growth deduced from tree ring analysis: A case study in North China. Ann. For. Sci. 2003, 60, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Fan, R.; Tanekura, K.; Morozumi, T.; Shingubara, R.; Tei, S.; Nogovitcyn, A.; Starostin, E.; Maximov, T.C.; Sugimoto, A. Adaptation of Willows in River Lowlands to Flooding under Arctic Amplification: Evidence from Nitrogen Content and Stable Isotope Dynamics. Wetlands 2020, 40, 2413–2424. [Google Scholar] [CrossRef]
- Liu, W.; Cao, W. Niche characteristics of main plant species in spruce-fir forests in Changbai Mountains. Chin. J. Ecol. 2011, 30, 1766–1774. (In Chinese) [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tmean | Pmean | Tmax | Tmin | RH | VPD | |
|---|---|---|---|---|---|---|
| δ 13 C | −0.711 ** | −0.005 | −0.676 ** | −0.162 | 0.347 ** | 0.263 * |
| WUE | 0.718 ** | −0.061 | 0.457 ** | 0.424 ** | −0.342 ** | −0.254 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.; Li, S.; Wang, J.; Guo, Y.; Fan, R.; Zhang, H.; Liu, Q.; Zhao, Y. Responses of Picea meyeri to Climatic Factors Revealed by Tree Ring Isotopes and Water Use Efficiency on Luya Mountain of North-Central China. Atmosphere 2023, 14, 615. https://doi.org/10.3390/atmos14040615
Han Y, Li S, Wang J, Guo Y, Fan R, Zhang H, Liu Q, Zhao Y. Responses of Picea meyeri to Climatic Factors Revealed by Tree Ring Isotopes and Water Use Efficiency on Luya Mountain of North-Central China. Atmosphere. 2023; 14(4):615. https://doi.org/10.3390/atmos14040615
Chicago/Turabian StyleHan, Yijie, Shuheng Li, Jiachuan Wang, Yili Guo, Rong Fan, Huan Zhang, Qi Liu, and Yiqi Zhao. 2023. "Responses of Picea meyeri to Climatic Factors Revealed by Tree Ring Isotopes and Water Use Efficiency on Luya Mountain of North-Central China" Atmosphere 14, no. 4: 615. https://doi.org/10.3390/atmos14040615