Arable Weed Patterns According to Temperature and Latitude Gradient in Central and Southern Spain


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
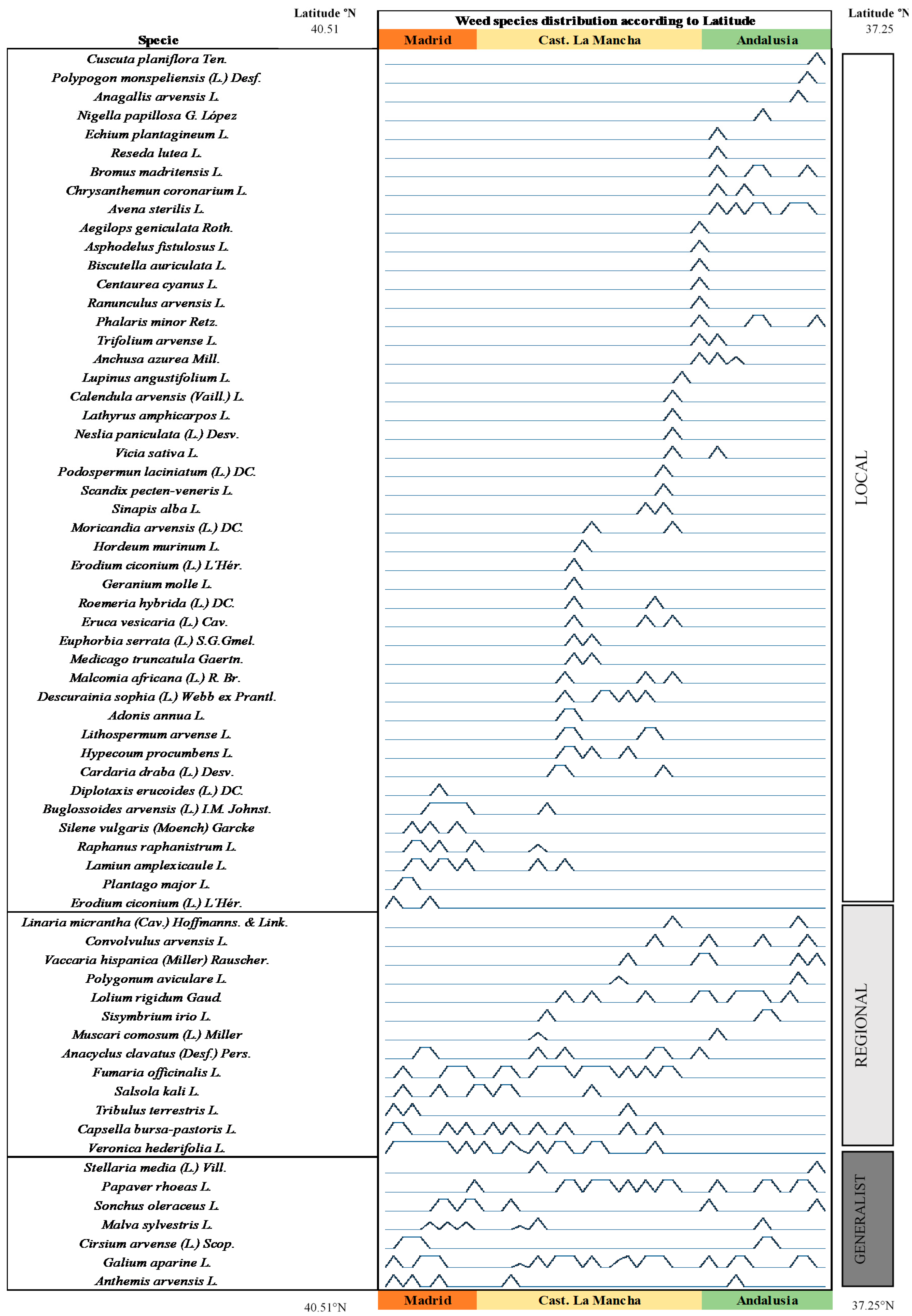
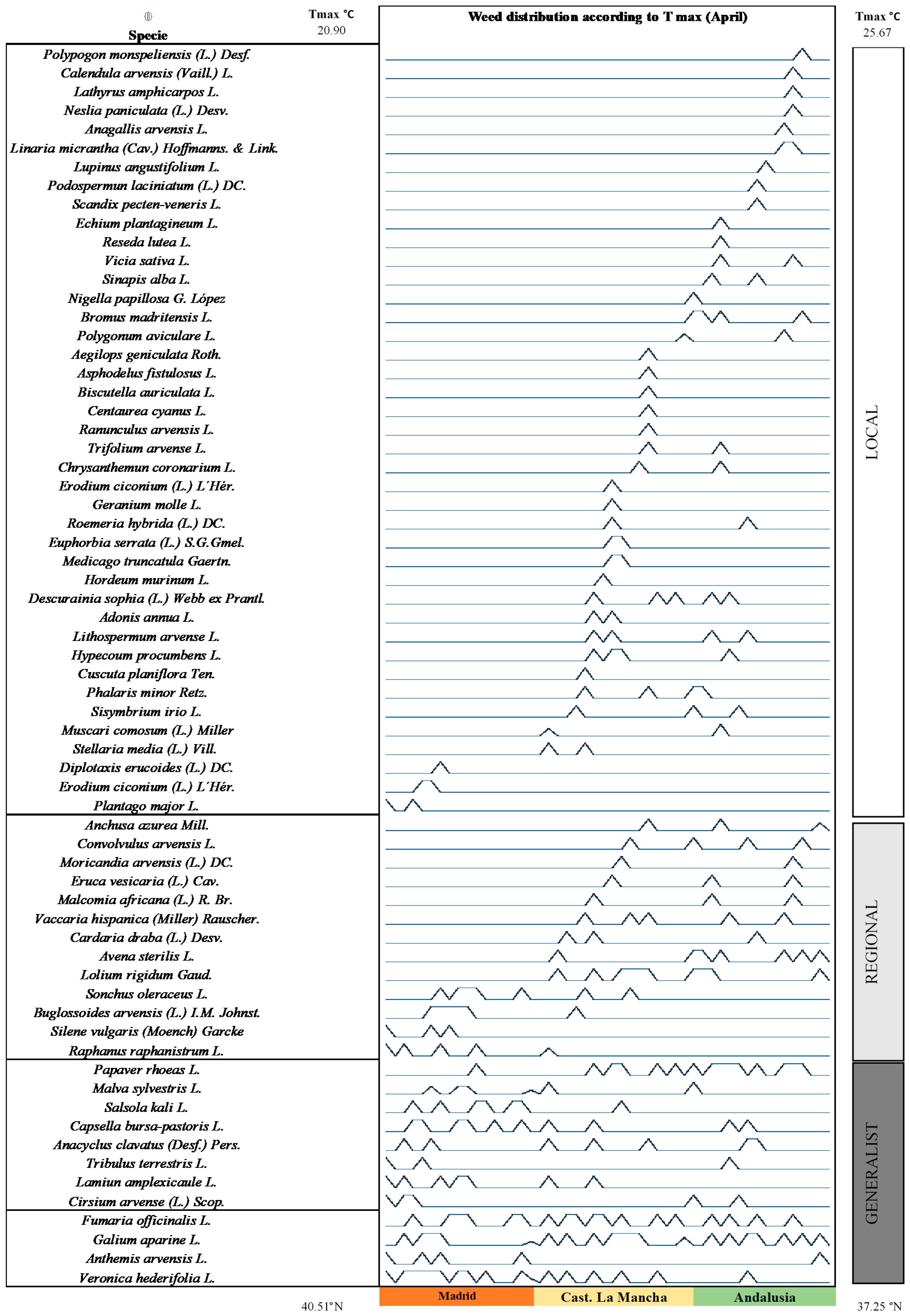
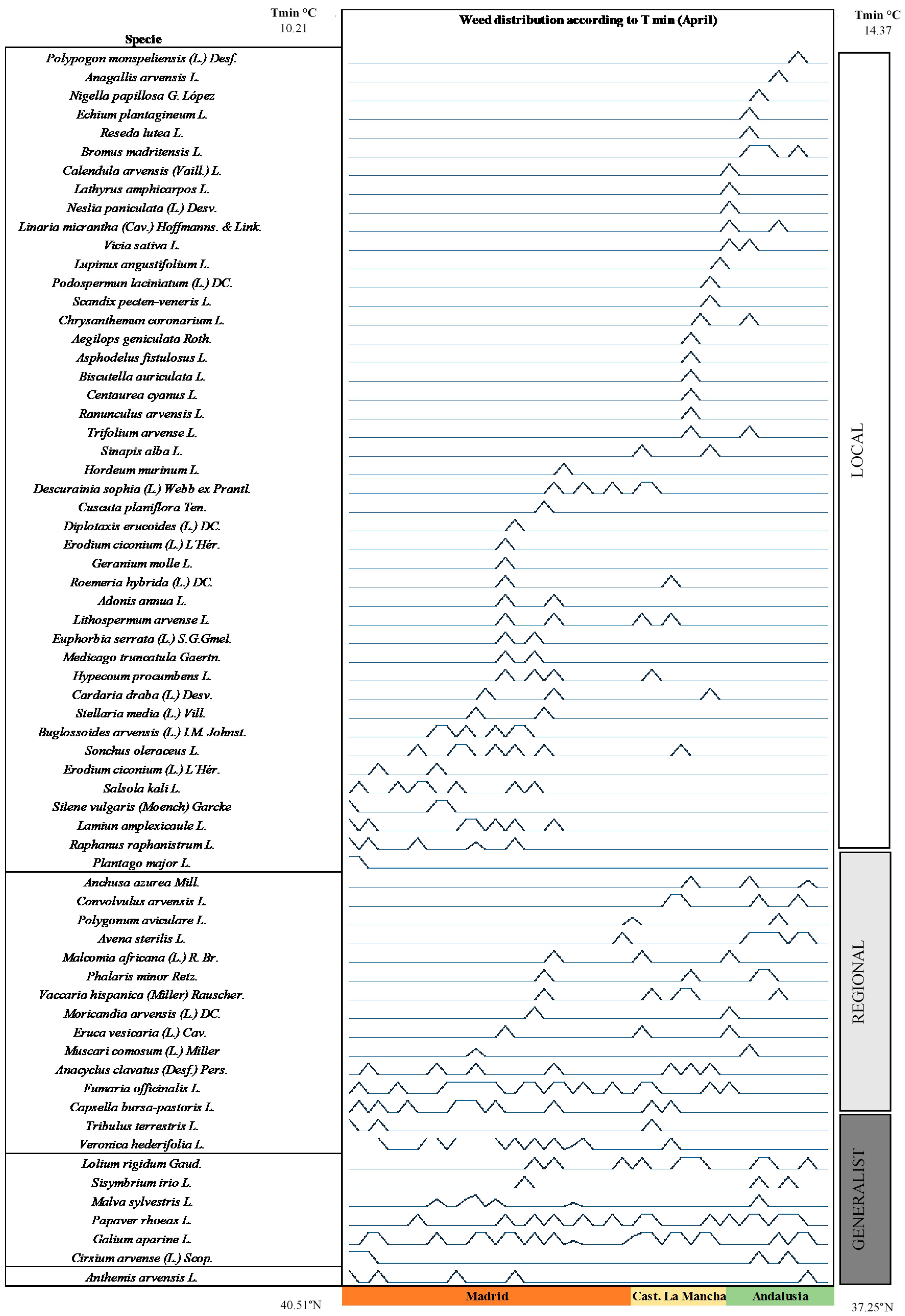
3. Results

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Reidsma, P.; Tekelenbur, T.; van den Berg, M.; Alkemade, R. Impacts of land-use change on biodiversity: An assessment of agricultural biodiversity in the European Union. Agric. Ecosyst. Environ. 2006, 114, 86–102. [Google Scholar] [CrossRef]
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Biodiversity—Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Brock, W.A.; Carpenter, S.R.; Scheffer, M. Regime Shifts, Environmental Signals, Uncertainty, and Policy Choice. In Complexity Theory for a Sustainable Future; Norberg, J., Cumming, G.S., Eds.; Columbia University Press: New York, NY, USA, 2008; pp. 180–206. [Google Scholar]
- Samhouri, J.F.; Levin, P.S.; Ainsworth, C.H. Identifying thresholds for ecosystem-based management. PLoS ONE 2010, 5, e8907. [Google Scholar] [CrossRef]
- Pautasso, M.; Dehnen-Schmutz, K.; Holdenrieder, O.; Pietravalle, S.; Salama, N.; Jeger, M.J.; Lange, E.; Hehl-Lange, S. Plant health and global change—Some implications for landscape management. Biol. Rev. 2010, 85, 729–755. [Google Scholar] [CrossRef]
- Schneider, A.; Havlík, P.; Schmid, E.; Valin, H.; Mosnier, A.; Obersteiner, M.; Böttcher, H.; Skalsky´, R.; Balkovič, J.; Sauer, T.; et al. Impacts of population growth, economic development, and technical change on global food production and consumption. Agric. Syst. 2011, 104, 204–215. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [Green Version]
- Petit, J.R.; Jouzel, J.; Raynaud, D.; Barkov, N.I.; Barnola, J.M.; Basile, I.; Bender, M.; Chappellaz, J.; Davis, M.; Delaygue, G.; et al. Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica. Nature 1999, 399, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Loss, S.R.; Terwilliger, L.A.; Peterson, A.C. Assisted colonization: Integrating conservation strategies in the face of climate change. Biol. Conserv. 2011, 144, 92–100. [Google Scholar] [CrossRef]
- Fried, G.; Norton, L.R.; Reboud, X. Environmental and management factors determining weed species composition and diversity in France. Agric. Ecosyst. Environ. 2008, 128, 68–76. [Google Scholar] [CrossRef]
- Potts, G.R.; Ewald, J.A.; Aebischer, N.J. Long-term changes in the flora of the cereal ecosystem on the Sussex Downs, England, focusing on the years 1968–2005. J. Appl. Ecol. 2010, 47, 215–226. [Google Scholar] [CrossRef]
- Andreasen, C.; Streibig, J.C. Evaluation of changes in weed flora in arable fields of Nordic countries—Based on Danish long-term surveys. Weed Res. 2011, 51, 214–226. [Google Scholar] [CrossRef]
- Salonen, J.; Hyvönen, T.; Kaseva, J.; Jalli, H. Impact of changed cropping practices on weed occurrence in spring cereals in Finland—A comparison of surveys in 1997–1999 and 2007–2009. Weed Res. 2013, 53, 110–120. [Google Scholar] [CrossRef]
- Post, E.; Forchhammer, M.C.; Stenseth, N.C.; Callaghan, T.V. The timing of life-history events in a changing climate. Proc. R. Soc. Lond. Ser. B. 2001, 268, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Nogues-Bravo, D. Predicting the past distribution of species climatic niches. Glob. Ecol. Biogeogr. 2009, 18, 521–531. [Google Scholar] [CrossRef]
- Estrella, N.; Sparks, T.H.; Menzel, A. Effects of temperature, phase type and timing, location, and human density on plant phenological responses in Europe. Clim. Res. 2009, 39, 235–248. [Google Scholar] [CrossRef]
- Kaukoranta, T.; Hakala, K. Impact of spring warming on sowing times of cereal, potato and sugar beet in Finland. Agric. Food Sci. 2008, 17, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Fleming, A.; Vanclay, F. Farmer responses to climate change and sustainable agriculture. A review. Agron. Sustain. Develop. 2010, 30, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Daccache, A.; Keay, C.A.; Jones, R.J.A.; Weatherhead, E.K.; Stalham, M.A.; Knox, J.W. Climate change and land suitability for potato production in England and Wales: Impacts and adaptation. J. Agric. Sci. 2012, 150, 161–177. [Google Scholar] [CrossRef] [Green Version]
- Gerowitt, B.; Bertke, E.; Hespelt, S.-K.; Tute, C. Towards multifunctional agriculture—Weeds as ecological goods? Weed Res. 2003, 43, 227–235. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Fuhrer, J. Agroecosystem responses to combinations of elevated CO2, ozone, and global climate change. Agric. Ecosyst. Environ. 2003, 97, 1–20. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Petit, S.; Boursault, A.; Le Guilloux, M.; Munier-Jolain, N.; Reboud, X. Weeds in agricultural landscapes. A review. Agron. Sustain. Develop. 2011, 31, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Clements, D.R.; Weise, S.F.; Swanton, C.J. Integrated weed management and weed species diversity. Phytoprotection 1994, 75, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Fried, G.; Petit, S.; Reboud, X. A specialist-generalist classification of the arable flora and its response to changes in agricultural practices. BMC Ecol. 2010, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chapin, F.S., III; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek P., M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef]
- Booth, B.D.; Swanton, C.J. Assembly theory applied to weed communities. Weed Sci. 2002, 50, 2–13. [Google Scholar] [CrossRef]
- Eriksson, O. Species pools in cultural landscapes—Niche construction, ecological opportunity and niche shifts. Ecography 2013, 36, 403–413. [Google Scholar] [CrossRef]
- Hawes, C.; Haughton, A.J.; Bohan, D.A.; Squire, G.R. Functional approaches for assessing plant and invertebrate abundance patterns in arable systems. Basic Appl. Ecol. 2009, 10, 34–42. [Google Scholar] [CrossRef]
- Urruty, N.; Deveaud, T.; Guyomard, H.; Boiffin, J. Impacts of agricultural land use changes on pesticide use in French agriculture. Eur. J. Agron. 2016, 80, 113–123. [Google Scholar] [CrossRef]
- Hyvönen, T.; Huusela-Veistola, E. Arable weeds as indicators of agricultural intensity—A case study from Finland. Biol. Conserv. 2008, 141, 2857–2864. [Google Scholar] [CrossRef]
- Dukes, J.S.; Pontius, J.; Orwig, D.; Garnas, J.R.; Rodgers, V.L.; Brazee, N.; Cooke, B.; Theoharides, K.A.; Stange, E.E.; Harrington, R.; et al. Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? Can. J. For. Res. 2009, 39, 231–248. [Google Scholar] [CrossRef]
- Singer, A.; Travism, J.M.J.; Johst, K. Interspecific interactions affect species and community responses to climate shifts. Oikos 2013, 122, 358–366. [Google Scholar] [CrossRef]
- Malavasi, M.; Santoro, R.; Cutini, M.; Acosta, A.T.R.; Carranza, M.L. The impact of human pressure on landscape patterns and plant species richness in Mediterranean coastal dunes. Plant Biosyst. 2016, 150, 73–82. [Google Scholar] [CrossRef]
- Bloomfield, J.P.; Williams, R.J.; Gooddy, D.C.; Cape, J.N.; Guha, P. Impacts of climate change on the fate and behaviour of pesticides in surface and groundwater—A UK perspective. Sci. Total Environ. 2006, 369, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Hanzlik, K.; Gerowitt, B. Occurrence and distribution of important weed species in German winter oilseed rape fields. J. Plant. Dis. Prot. 2012, 119, 107–120. [Google Scholar] [CrossRef]
- Zimdahl, R.L. Weed-Crop Competition: A Review, 2nd ed.; Blackwell: Ames, IA, USA, 2004; p. 220. [Google Scholar]
- Jump, A.S.; Peñuelas, J. Running and stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- Cimalova, S.; Lososova, Z. Arable weed vegetation of the northeastern part of the Czech Republic: Effects of environmental factors on species composition. Plant Ecol. 2009, 203, 45–57. [Google Scholar] [CrossRef]
- Silc, U.; Vrbnicanin, S.; Bozi, C.D.; Carni, A.; Stevanovic, Z.D. Weed vegetation in the northwestern Balkans: Diversity and species composition. Weed Res. 2009, 49, 602–612. [Google Scholar] [CrossRef]
- Hulme, P.E.; Barrett, S.C.H. Integrating trait- and niche-based approaches to assess contemporary evolution in alien plant species. J. Ecol. 2013, 101, 68–77. [Google Scholar] [CrossRef]
- Lososova, Z.; Chytry, M.; Kühn, I.; Hájeka, O.; Horáková, V.; Pysek, P.; Tichy, L. Patterns of plant traits in annual vegetation of man-made habitats in Central Europe. Perspect. Plant. Ecol. Evol. Syst. 2006, 8, 69–81. [Google Scholar] [CrossRef]
- Hyvönen, T.; Luoto, M.; Uotila, P. Assessment of weed establishment risk in a changing European climate. Agric. Food Sci. 2012, 21, 348–360. [Google Scholar] [CrossRef] [Green Version]
- Peters, K.; Breitsameter, L.; Gerowitt, B. Impact of climate change on weeds in agriculture: A review. Agron. Sustain. Develop. 2014, 34, 707–721. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandía, M.L.; Casanova, C.; Sánchez, F.J.; Tenorio, J.L.; Santín-Montanyá, M.I. Arable Weed Patterns According to Temperature and Latitude Gradient in Central and Southern Spain. Atmosphere 2020, 11, 853. https://doi.org/10.3390/atmos11080853
Gandía ML, Casanova C, Sánchez FJ, Tenorio JL, Santín-Montanyá MI. Arable Weed Patterns According to Temperature and Latitude Gradient in Central and Southern Spain. Atmosphere. 2020; 11(8):853. https://doi.org/10.3390/atmos11080853
Chicago/Turabian StyleGandía, María Luisa, Carlos Casanova, Francisco Javier Sánchez, José Luís Tenorio, and María Inés Santín-Montanyá. 2020. "Arable Weed Patterns According to Temperature and Latitude Gradient in Central and Southern Spain" Atmosphere 11, no. 8: 853. https://doi.org/10.3390/atmos11080853