
Heterochromatin Is Not the Only Place for satDNAs: The High Diversity of satDNAs in the Euchromatin of the Beetle Chrysolina americana (Coleoptera, Chrysomelidae)

 , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Insects, Preparation of Chromosome Spreads, and C-Banding
2.2. DNA Extraction, Genome Sequencing, and Bioinformatic Analysis
2.3. Fluorescence In Situ Hybridization (FISH) and Chromosome In Silico Mapping (CHRISMAPP)
3. Results and Discussion
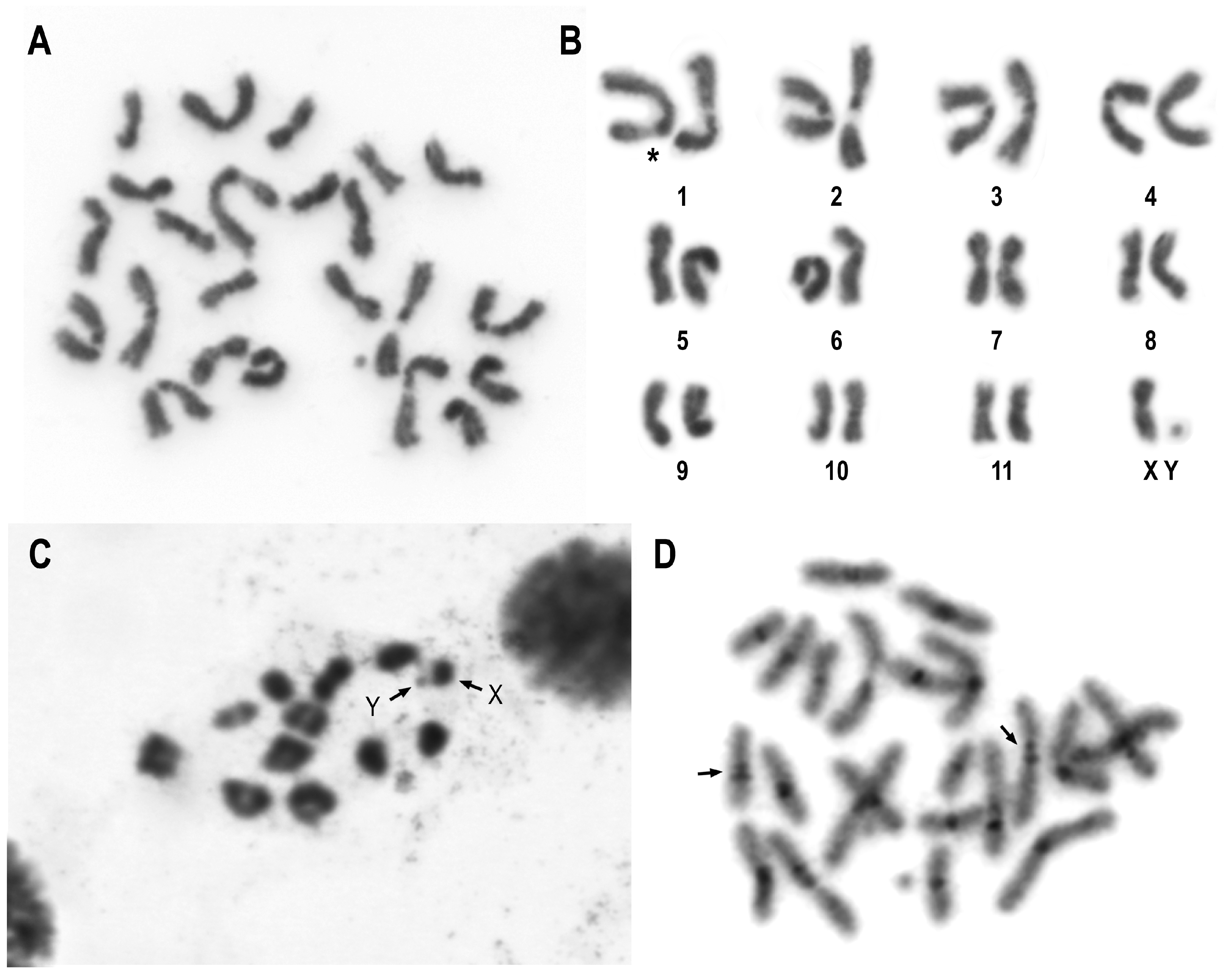
3.1. General Features of Chromosomes of C. americana
3.2. Satellitome Analysis of C. americana Genome Reveals Extensive Diversity of satDNAs with Amplification of Few Families
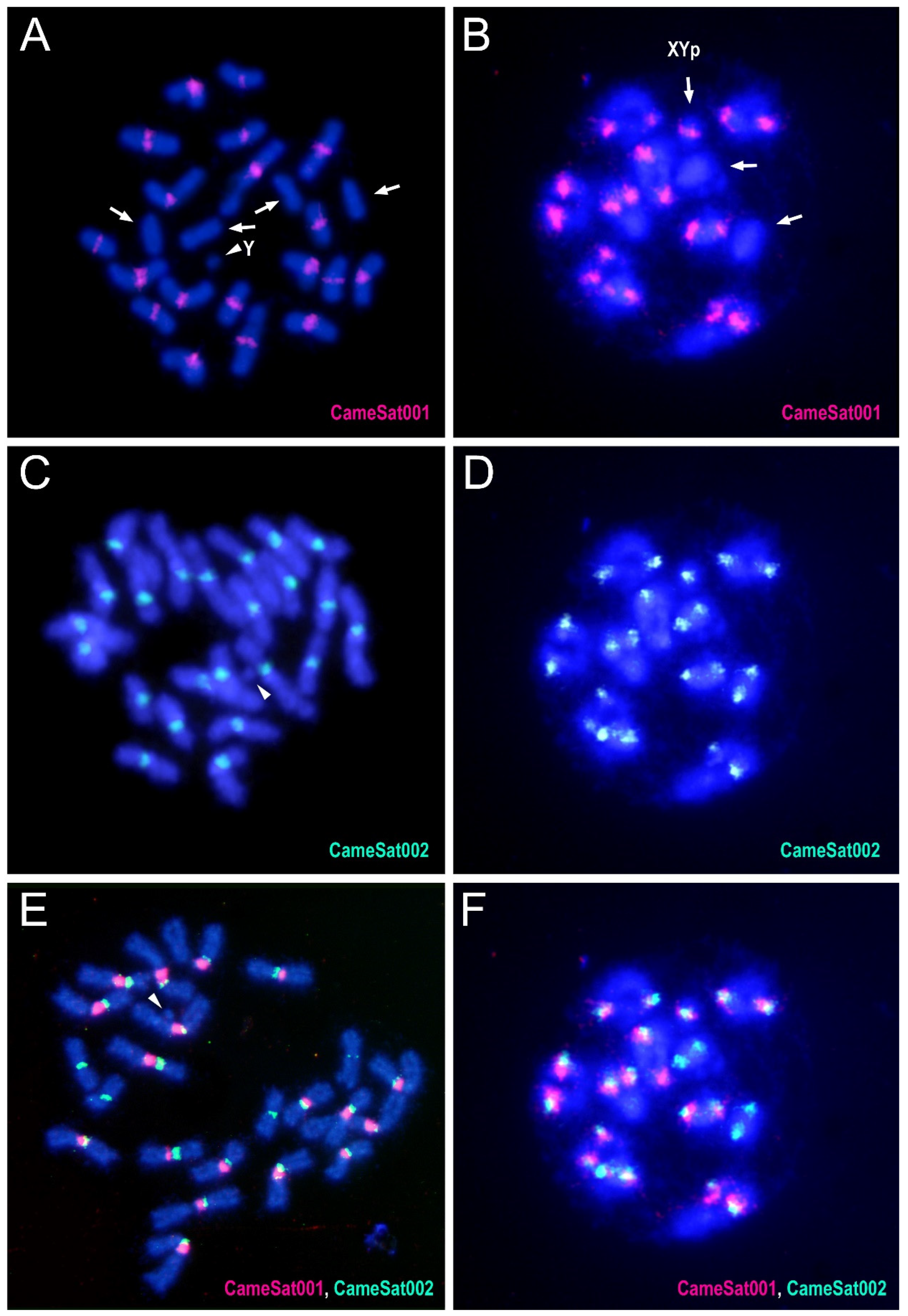
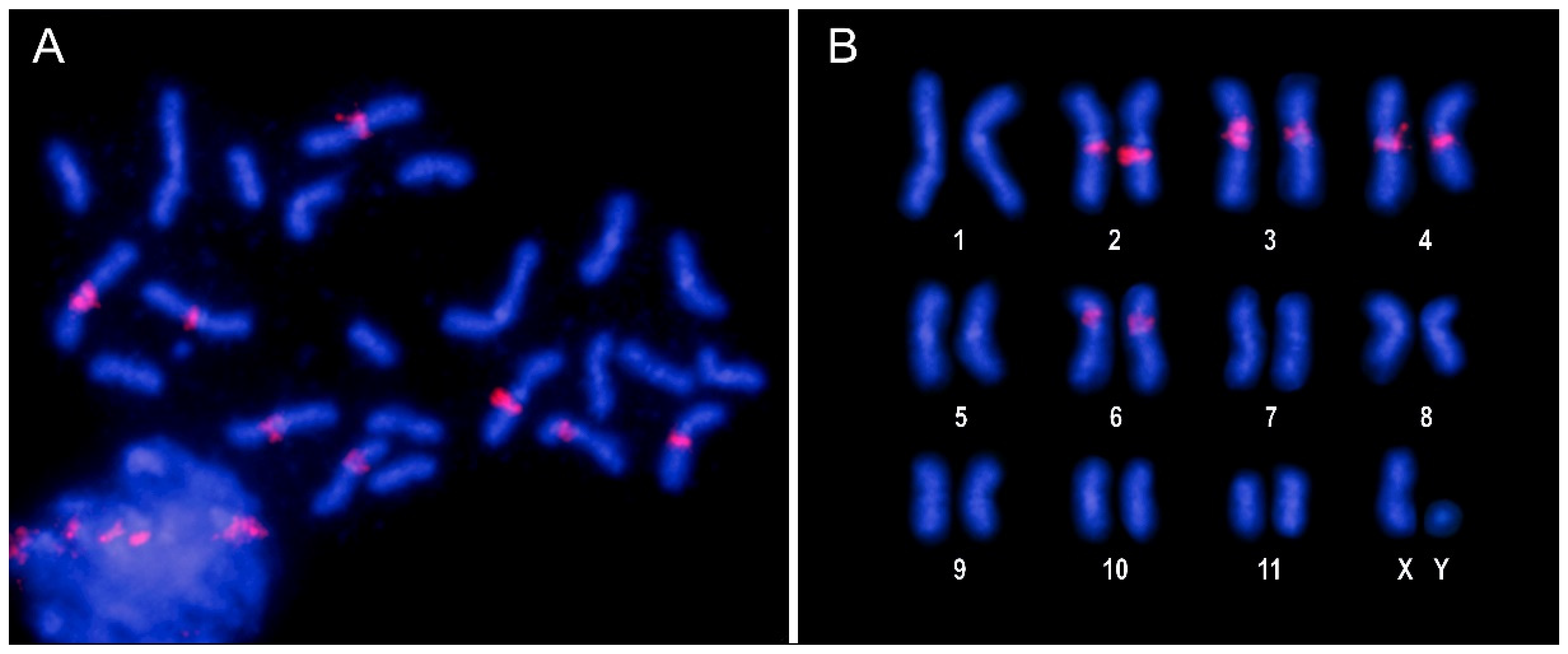
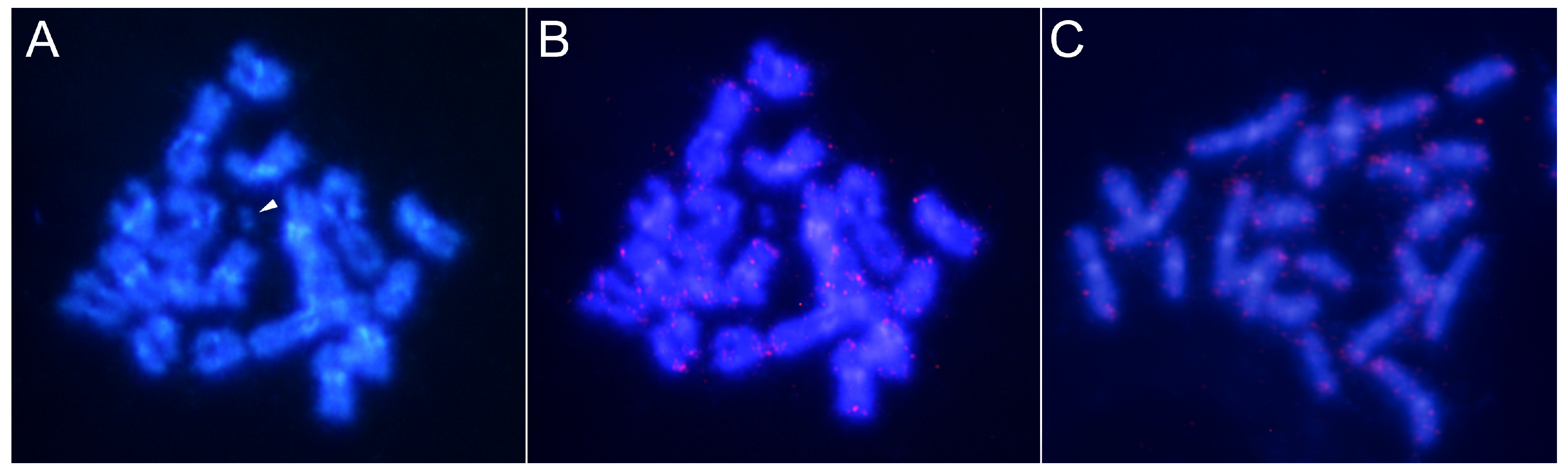
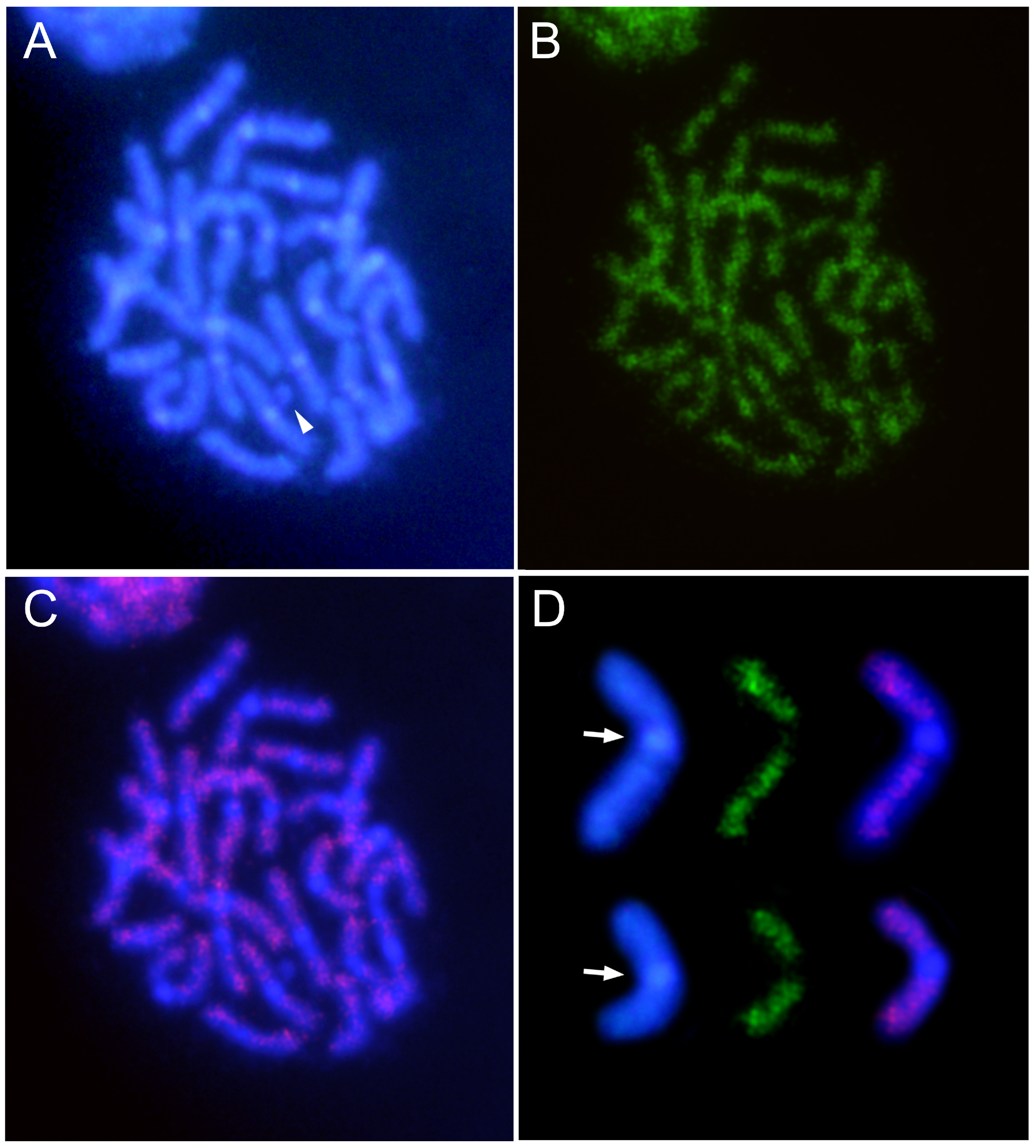
3.3. Characterization and Chromosomal Localization of the Main Families of Satellite DNA Shows an Unpreceded High Number of satDNAs in Euchromatin
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bouchard, P.; Bousquet, Y.; Davies, A.E.; Alonso–Zarazaga, M.A.; Lawrence, J.F.; Lyal, C.H.C.; Newton, A.F.; Reid, C.A.M.; Schmitt, M.; Ślipiński, S.A.; et al. Family–group names in Coleoptera (Insecta). Zookeys 2011, 88, 1–972. [Google Scholar] [CrossRef] [PubMed]
- Daccordi, M. Notes on the distribution of the Chrysomelinae and their possible origin. In Chrysomelidae Biology Vol. 1: The Classification. Phylogeny and Genetics; Jolivet, P., Cox, M.L., Eds.; SPB Academy Publishing: Amsterdam, The Netherlands, 1996; pp. 399–412. [Google Scholar]
- Beenen, R.; Roques, A. Leaf and seed beetles (Coleoptera, Chrysomelidae). Chapter 8.3. BioRisk 2010, 4, 267–292. [Google Scholar] [CrossRef]
- Kippenberg, H. Chrysomelinae. In Chrysomeloidea; Löbl, I., Smetana, A., Eds.; Catalogue of Palaearctic Coleoptera (vol. 6); Brill: Stenstrup, Denmark, 2010; pp. 390–443. [Google Scholar]
- Bieńkowski, A.O.; Orlova–Bienkowskaja, M.J. Alien leaf beetles (Coleoptera, Chrysomelidae) of European Russia and some general tendencies of leaf beetle invasions. PLoS ONE 2018, 13, e0203561. [Google Scholar] [CrossRef] [PubMed]
- Hadjiconstantis, M.; Zoumides, C. First records of the pest leaf beetle Chrysolina (Chrysolinopsis) americana (Linnaeus, 1758) (Coleoptera, Chrysomelidae) in Cyprus—A study initiated from social media. Biodivers. Data J. 2021, 9, e61349. [Google Scholar] [CrossRef] [PubMed]
- John, B.; Miklos, G.L. Functional aspects of satellite DNA and heterochromatin. Int. Rev. Cytol. 1979, 58, 1–114. [Google Scholar] [CrossRef] [PubMed]
- López–Flores, I.; Garrido–Ramos, M.A. The repetitive DNA content of eukaryotic genomes. Genome Dyn. 2012, 7, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S. So much “junk” DNA in our genome. Brookhaven Sysp. Biol. 1972, 23, 366–370. [Google Scholar]
- Louzada, S.; Lopes, M.; Ferreira, D.; Adega, F.; Escudeiro, A.; Gama–Carvalho, M.; Chaves, R. Decoding the role of Satellite DNA in genome architecture and plasticity–An evolutionary and clinical affair. Genes 2020, 11, 72. [Google Scholar] [CrossRef]
- Thakur, J.; Packiaraj, J.; Henikoff, S. Sequence, chromatin and evolution of satellite DNA. Int. J. Mol. Sci. 2021, 22, 4309. [Google Scholar] [CrossRef]
- Adega, F.; Guedes-Pinto, H.; Chaves, R. Satellite DNA in the karyotype evolution of domestic animals—Clinical considerations. Cytogenet. Genome Res. 2009, 126, 12–20. [Google Scholar] [CrossRef]
- Shatskikh, A.S.; Kotov, A.A.; Adashev, V.E.; Bazylev, S.S.; Olenina, L.V. Functional significance of satellite DNAs: Insights from Drosophila. Front. Cell Dev. Biol. 2020, 8, 312. [Google Scholar] [CrossRef]
- Talbert, P.B.; Henikoff, S. The genetics and epigenetics of satellite centromeres. Genome Res. 2022, 32, 608–615. [Google Scholar] [CrossRef]
- Plohl, M.; Meštrović, N.; Mravinac, B. Centromere identity from the DNA point of view. Chromosoma 2014, 123, 313–325. [Google Scholar] [CrossRef]
- Henikoff, S.; Ahmad, K.; Malik, H.S. The centromere paradox: Stable inheritance with rapidly evolving DNA. Science 2001, 293, 1098–1102. [Google Scholar] [CrossRef]
- Larracuente, A.M. The organization and evolution of the Responder satellite in species of the Drosophila melanogaster group: Dynamic evolution of a target of meiotic drive. BMC Evol. Biol. 2014, 14, 233. [Google Scholar] [CrossRef]
- Ferree, P.M.; Barbash, D.A. Species–specific heterochromatin prevents mitotic chromosome segregation to cause hybrid lethality in Drosophila. PLoS Biol. 2009, 7, e1000234. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kinney, N.A.; Timoshevskiy, V.A.; Sharakhova, M.V.; Sharakhov, I.V. Structural variation of the X chromosome heterochromatin in the Anopheles gambiae complex. Genes 2020, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Podgornaya, O.I. Nuclear organization by satellite DNA, SAF–A/hnRNPU and matrix attachment regions. Semin. Cell Dev. Biol. 2022, 128, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Singchat, W.; Jehangir, M.; Suntronpong, A.; Panthum, T.; Malaivijitnond, S.; Srikulnath, K. Dark matter of primate genomes: Satellite DNA repeats and their evolutionary dynamics. Cells 2020, 9, 2714. [Google Scholar] [CrossRef] [PubMed]
- Biscotti, M.A.; Canapa, A.; Forconi, M.; Olmo, E.; Barucca, M. Transcription of tandemly repetitive DNA: Functional roles. Chromosome Res. 2015, 23, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Feliciello, I.; Akrap, I.; Ugarković, Đ. Satellite DNA modulates gene expression in the beetle Tribolium castaneum after heat stress. PLoS Genet. 2015, 11, e1005466. [Google Scholar] [CrossRef]
- Ferreira, D.; Meles, S.; Escudeiro, A.; Mendes–da–Silva, A.; Adega, F.; Chaves, R. Satellite non–coding RNAs: The emerging players in cells, cellular pathways and cancer. Chromosome Res. 2015, 23, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Porokhovnik, L.N.; Veiko, N.N.; Ershova, E.S.; Kostyuk, S.V. The role of human satellite III (1q12) copy number variation in the adaptive response during aging, stress, and pathology: A pendulum model. Genes 2021, 12, 1524. [Google Scholar] [CrossRef] [PubMed]
- Ugarković, Đ.; Sermek, A.; Ljubić, S.; Feliciello, I. Satellite DNAs in health and disease. Genes 2022, 13, 1154. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.; Louzada, S.; Ferreira, D.; Veríssimo, G.; Eleutério, D.; Gama–Carvalho, M.; Chaves, R. Human satellite 1A analysis provides evidence of pericentromeric transcription. BMC Biol. 2023, 21, 28. [Google Scholar] [CrossRef] [PubMed]
- Montiel, E.E.; Mora, P.; Rico–Porras, J.M.; Palomeque, T.; Lorite, P. Satellitome of the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), the most diverse among insects. Front. Ecol. Evol. 2022, 10, 826808. [Google Scholar] [CrossRef]
- Lower, S.S.; McGurk, M.P.; Clark, A.G.; Barbash, D.A. Satellite DNA evolution: Old ideas, new approaches. Curr. Opin. Genet. Dev. 2018, 48, 70–78. [Google Scholar] [CrossRef]
- Šatović, E. Tools and databases for solving problems in detection and identification of repetitive DNA sequences. Period. Biol. 2020, 121, 7–14. [Google Scholar] [CrossRef]
- Novák, P.; Neumann, P.; Macas, J. Graph–based clustering and characterization of repetitive sequences in next–generation sequencing data. BMC Bioinform. 2010, 11, 378. [Google Scholar] [CrossRef]
- Novák, P.; Neumann, P.; Pech, J.; Steinhaisl, J.; Macas, J. RepeatExplorer: A Galaxy–based web server for genome–wide characterization of eukaryotic repetitive elements from next–generation sequence reads. Bioinformatics 2013, 29, 792–793. [Google Scholar] [CrossRef]
- Novák, P.; Ávila Robledillo, L.; Koblížková, A.; Vrbová, I.; Neumann, P.; Macas, J. TAREAN: A computational tool for identification and characterization of satellite DNA from unassembled short reads. Nucleic Acids Res. 2017, 45, e111. [Google Scholar] [CrossRef] [PubMed]
- Ruiz–Ruano, F.J.; López–León, M.D.; Cabrero, J.; Camacho, J.P.M. High–throughput analysis of the satellitome illuminates satellite DNA evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef]
- Mora, P.; Vela, J.; Ruiz–Ruano, F.J.; Ruiz–Mena, A.; Montiel, E.E.; Palomeque, T.; Lorite, P. Satellitome analysis in the ladybird beetle Hippodamia variegata (Coleoptera, Coccinellidae). Genes 2020, 11, 783. [Google Scholar] [CrossRef]
- Oppert, B.; Muszewska, A.; Steczkiewicz, K.; Šatović-Vukšić, E.; Plohl, M.; Fabrick, J.A.; Vinokurov, K.S.; Koloniuk, I.; Johnston, J.S.; Smith, T.P.L.; et al. The genome of Rhyzopertha dominica (Fab.) (Coleoptera: Bostrichidae): Adaptation for success. Genes 2022, 13, 446. [Google Scholar] [CrossRef] [PubMed]
- Gržan, T.; Dombi, M.; Despot-Slade, E.; Veseljak, D.; Volarić, M.; Meštrović, N.; Plohl, M.; Mravinac, B. The low-copy-number satellite DNAs of the model beetle Tribolium castaneum. Genes 2023, 14, 999. [Google Scholar] [CrossRef]
- Oppert, B.; Dossey, A.T.; Chu, F.-C.; Šatović-Vukšić, E.; Plohl, M.; Smith, T.P.L.; Koren, S.; Olmstead, M.L.; Leierer, D.; Ragan, G.; et al. The genome of the yellow mealworm, Tenebrio molitor: It’s bigger than you think. Genes 2023, 14, 2209. [Google Scholar] [CrossRef]
- Petitpierre, E. Cytogenetics, cytotaxonomy and genetics of Chrysomelidae. In Biology of Chrysomelidae; Jolivet, P., Petitpierre, E., Hsiao, T.H., Eds.; Series Entomologica (Vol. 42); Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988; pp. 131–159. [Google Scholar]
- Petitpierre, E. Karyological analysis of the subfamily Chrysomelinae (Coleoptera: Chrysomelidae). Cut. Edge Res. Biol. 2023, 5, 104–121. [Google Scholar] [CrossRef]
- Blackmon, H.; Jonika, M.M.; Alfieri, J.M.; Demuth, J.P. Drift drives the evolution of chromosome number I: The impact of trait transitions on genome evolution in Coleoptera. J. Hered. 2024, 115, esae001. [Google Scholar] [CrossRef]
- Lorite, P.; Palomeque, T.; Garnería, I.; Petitpierre, E. Characterization and chromosome location of satellite DNA in the leaf beetle Chrysolina americana (Coleoptera, Chrysomelidae). Genetica 2001, 110, 143–150. [Google Scholar] [CrossRef]
- Lorite, P.; Carrillo, J.A.; Garnería, I.; Petitpierre, E.; Palomeque, T. Satellite DNA in the elm leaf beetle, Xanthogaleruca luteola (Coleoptera, Chrysomelidae): Characterization, interpopulation analysis, and chromosome location. Cytogenet. Genome Res. 2002, 98, 302–307. [Google Scholar] [CrossRef]
- Lorite, P.; Torres, M.I.; Palomeque, T. Characterization of two unrelated satellite DNA families in the Colorado potato beetle Leptinotarsa decemlineata (Coleoptera, Chrysomelidae). Bull. Entomol. Res. 2013, 103, 538–546. [Google Scholar] [CrossRef]
- Palomeque, T.; Muñoz–López, M.; Carrillo, J.A.; Lorite, P. Characterization and evolutionary dynamics of a complex family of satellite DNA in the leaf beetle Chrysolina carnifex (Coleoptera, Chrysomelidae). Chromosome Res. 2005, 13, 795–807. [Google Scholar] [CrossRef]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High throughput Sequence Data [Online]. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 December 2023).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones–Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved telomere detection using a telomere repeat probe (TTAGGG)n generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef]
- Cabral-de-Mello, D.C.; Marec, F. Universal fluorescence in situ hybridization (FISH) protocol for mapping repetitive DNAs in insects and other arthropods. Mol. Genet. Genom. 2021, 296, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Pinkel, D.; Straume, T.; Gray, J.M. Cytogenetic analysis using quantitative, high sensitivity fluorescence hybridization. Proc. Natl. Acad. Sci. USA 1986, 83, 2934–2938. [Google Scholar] [CrossRef] [PubMed]
- Tunjić-Cvitanić, M.; Pasantes, J.J.; García-Souto, D.; Cvitanić, T.; Plohl, M.; Šatović-Vukšić, E. Satellitome analysis of the Pacific oyster Crassostrea gigas reveals new pattern of satellite DNA organization, highly scattered across the genome. Int. J. Mol. Sci. 2021, 22, 6798. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org (accessed on 10 February 2024).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer International Publishing: Cham, Switzerland, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 5 February 2024).
- Petitpierre, E. Karyometric differences among nine species of the genus Chrysolina Mots. (Coleoptera, Chrysomelidae). Can. J. Genet. Cytol. 1983, 25, 33–39. [Google Scholar] [CrossRef]
- Blackmon, H.; Demuth, J.P. Coleoptera Karyotype Database. Coleopterists Bull. 2015, 69, 174–175. [Google Scholar] [CrossRef]
- Smith, S.G.; Virkki, N. Coleoptera. In Animal Cytogenetics; John, B., Ed.; Insecta 5 (Vol. 3); Gebrüder Borntraeger: Berlin, Germany, 1978; p. 366. [Google Scholar]
- Palomeque, T.; Lorite, P. Satellite DNA in insects: A review. Heredity 2008, 100, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Rozek, M.; Lachowska, D.; Petitpierre, E.; Holecova, M. C–bands on chromosomes of 32 beetle species (Coleoptera: Elateridae, Cantharidae, Oedemeridae, Cerambycidae, Anthicidae, Chrysomelidae, Attelabidae, and Curculionidae). Hereditas 2004, 140, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, R.G.; Mulhair, P.O.; Holland, P.W.H. GC content across insect genomes: Phylogenetic patterns, causes and consequences. J. Mol. Evol. 2024. (advance online publications). [Google Scholar] [CrossRef]
- Kuznetsova, V.; Grozeva, S.; Gokhman, V. Telomere structure in insects: A review. J. Zool. Syst. Evol. Res. 2020, 58, 127–158. [Google Scholar] [CrossRef]
- Vítková, M.; Král, J.; Traut, W.; Zrzavý, J.; Marec, F. The evolutionary origin of insect telomeric repeats, (TTAGG)n. Chromosome Res. 2005, 13, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Mravinac, B.; Meštrović, N.; Cavrak, V.V.; Plohl, M. TCAGG, an alternative telomeric sequence in insects. Chromosoma 2011, 120, 367–376. [Google Scholar] [CrossRef]
- Mora, P.; Pita, S.; Montiel, E.E.; Rico–Porras, J.M.; Palomeque, T.; Panzera, F.; Lorite, P. Making the genome huge: The case of Triatoma delpontei, a Triatominae species with more than 50% of its genome full of satellite DNA. Genes 2023, 14, 371. [Google Scholar] [CrossRef]
- Anjos, A.; Milani, D.; Bardella, V.B.; Paladini, A.; Cabral-de-Mello, D.C. Evolution of satDNAs on holocentric chromosomes: Insights from hemipteran insects of the genus Mahanarva. Chromosome Res. 2023, 31, 5. [Google Scholar] [CrossRef]
- Ruiz–Ruano, F.J.; Castillo–Martínez, J.; Cabrero, J.; Gómez, R.; Camacho, J.P.M.; López–León, M.D. High–throughput analysis of satellite DNA in the grasshopper Pyrgomorpha conica reveals abundance of homologous and heterologous higher–order repeats. Chromosoma 2018, 127, 323–340. [Google Scholar] [CrossRef]
- Šatović-Vukšić, E.; Plohl, M. Satellite DNAs—From localized to highly dispersed genome components. Genes 2023, 14, 742. [Google Scholar] [CrossRef] [PubMed]
- Cabrero, J.; Perfectti, F.; Gómez, R.; Camacho, J.P.; López-León, M.D. Population variation in the A chromosome distribution of satellite DNA and ribosomal DNA in the grasshopper Eyprepocnemis plorans. Chromosome Res. 2003, 11, 375–381. [Google Scholar] [CrossRef]
- Milani, D.; Ramos, É.; Loreto, V.; Martí, D.A.; Cardoso, A.L.; de Moraes, K.C.M.; Martins, C.; Cabral-de-Mello, D.C. The satellite DNA AflaSAT-1 in the A and B chromosomes of the grasshopper Abracris flavolineata. BMC Genet. 2017, 18, 81. [Google Scholar] [CrossRef] [PubMed]
- Elder, J.F., Jr.; Turner, B.J. Concerted evolution at the population level: Pupfish HindIII satellite DNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 994–998. [Google Scholar] [CrossRef]
- Feliciello, I.; Akrap, I.; Brajković, J.; Zlatar, I.; Ugarković, Đ. Satellite DNA as a driver of population divergence in the red flour beetle Tribolium castaneum. Genome Biol. Evol. 2014, 7, 228–239. [Google Scholar] [CrossRef]
- Rovatsos, M.; Marchal, J.A.; Giagia-Athanasopoulou, E.; Sánchez, A. Molecular composition of heterochromatin and its contribution to chromosome variation in the Microtus thomasi/Microtus atticus species complex. Genes 2021, 12, 807. [Google Scholar] [CrossRef]
- Hall, A.B.; Papathanos, P.A.; Sharma, A.; Cheng, C.; Akbari, O.S.; Assour, L.; Bergman, N.H.; Cagnetti, A.; Crisanti, A.; Dottorini, T.; et al. Radical remodeling of the Y chromosome in a recent radiation of malaria mosquitoes. Proc. Natl. Acad. Sci. USA 2016, 113, E2114–E2123. [Google Scholar] [CrossRef] [PubMed]
- Cabral-de-Mello, D.C.; Mora, P.; Rico-Porras, J.M.; Ferretti, A.B.S.M.; Palomeque, T.; Lorite, P. The spread of satellite DNAs in euchromatin and insights into the multiple sex chromosome evolution in Hemiptera revealed by repeatome analysis of the bug Oxycarenus hyalinipennis. Insect Mol. Biol. 2023, 32, 725–737. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SatDNA Family | Oligonucleotide | Sequence |
|---|---|---|
| CameSat001-141 | Came_CL2-F Came_CL2-R | ACCATGATGCGTGCCAAGTC |
| GTCAACAGAAATCATTGTCGGGTC | ||
| CameSat002-187 | Came_CL26-F Came_CL26-R | ACAAGTCGAGACATACGAAGCAC |
| CCAAAATACAGAACAAGTCCAGCTG | ||
| CameSat003-10 | Came_CL1 | GACTTGTCCCGACTTGTCCC |
| CameSat004-322 | Came_CL4-F Came_CL4-R | CGAACGCCAATCGATTCAGAATG |
| CAGAATTGCTCTATCTTCAACCGTTC | ||
| CameSat005-499 | Came_CL11-F Came_CL11-R | ATGTCCGTCGTGGTATTAGCCAG |
| ACTCCCAAGCAGCACAGTCTC |
| satDNA Family | % Genome | Divergence (K2P) | RUL (bp) | A+T (%) | satDNA Family | % Genome | Divergence (K2P) | RUL (bp) | A+T (%) |
|---|---|---|---|---|---|---|---|---|---|
| CameSat001-141 | 8.9588 | 7.68 | 141 | 61.0 | CameSat084-141 | 0.0088 | 11.67 | 141 | 67.4 |
| CameSat002-187 | 2.4143 | 7.18 | 187 | 58.8 | CameSat085-625 | 0.0087 | 2.28 | 625 | 61.3 |
| CameSat003-10 | 1.2658 | 9.60 | 10 | 40.0 | CameSat086-159 | 0.0087 | 12.12 | 159 | 66.0 |
| CameSat004-322 | 0.6512 | 17.83 | 322 | 67.4 | CameSat087-171 | 0.0085 | 9.58 | 171 | 66.7 |
| CameSat005-499 | 0.3837 | 11.62 | 499 | 57.5 | CameSat088-132 | 0.0081 | 12.79 | 132 | 56.1 |
| CameSat006-2670 | 0.2748 | 2.59 | 2670 | 56.1 | CameSat089-151 | 0.0080 | 14.26 | 151 | 68.9 |
| CameSat007-313 | 0.2600 | 11.97 | 313 | 62.6 | CameSat090-288 | 0.0079 | 6.61 | 288 | 63.2 |
| CameSat008-454 | 0.2310 | 8.36 | 454 | 60.4 | CameSat091-143 | 0.0079 | 9.77 | 143 | 44.1 |
| CameSat009-206 | 0.1666 | 3.63 | 206 | 61.2 | CameSat092-160 | 0.0078 | 13.12 | 160 | 68.8 |
| CameSat010-3664 | 0.1401 | 3.63 | 3664 | 65.9 | CameSat093-182 | 0.0073 | 16.57 | 182 | 63.7 |
| CameSat011-2186 | 0.1381 | 8.18 | 2186 | 66.3 | CameSat094-142 | 0.0071 | 7.69 | 142 | 62.7 |
| CameSat012-346 | 0.1372 | 10.03 | 346 | 54.0 | CameSat095-231 | 0.0067 | 13.07 | 231 | 61.5 |
| CameSat013-1223 | 0.1284 | 11.99 | 1223 | 63.2 | CameSat096-135 | 0.0066 | 17.45 | 135 | 63.0 |
| CameSat014-163 | 0.1227 | 16.32 | 163 | 58.3 | CameSat097-76 | 0.0066 | 11.95 | 76 | 51.9 |
| CameSat015-288 | 0.1225 | 14.10 | 288 | 58.8 | CameSat098-1148 | 0.0065 | 2.57 | 1148 | 61.1 |
| CameSat016-449 | 0.1132 | 6.45 | 449 | 57.5 | CameSat099-146 | 0.0062 | 10.12 | 146 | 61.0 |
| CameSat017-479 | 0.1115 | 9.50 | 479 | 60.1 | CameSat100-146 | 0.0062 | 10.34 | 146 | 71.2 |
| CameSat018-280 | 0.0978 | 10.52 | 280 | 67.1 | CameSat101-649 | 0.0061 | 2.99 | 649 | 62.4 |
| CameSat019-228 | 0.0967 | 19.91 | 228 | 66.2 | CameSat102-141 | 0.0059 | 11.29 | 141 | 66.0 |
| CameSat020-336 | 0.0665 | 6.27 | 336 | 59.2 | CameSat103-172 | 0.0059 | 14.49 | 172 | 63.4 |
| CameSat021-312 | 0.0663 | 13.13 | 312 | 62.5 | CameSat104-165 | 0.0058 | 16.43 | 165 | 64.8 |
| CameSat022-324 | 0.0612 | 17.54 | 324 | 66.7 | CameSat105-952 | 0.0055 | 2.13 | 952 | 63.8 |
| CameSat023-146 | 0.0571 | 7.12 | 146 | 63.0 | CameSat106-309 | 0.0055 | 7.04 | 309 | 58.3 |
| CameSat024-316 | 0.0544 | 4.70 | 316 | 54.4 | CameSat107-212 | 0.0053 | 4.39 | 212 | 64.6 |
| CameSat025-150 | 0.0517 | 3.75 | 150 | 64.7 | CameSat108-410 | 0.0052 | 18.03 | 410 | 65.4 |
| CameSat026-65 | 0.0514 | 6.85 | 65 | 80.0 | CameSat109-148 | 0.0052 | 11.11 | 148 | 65.5 |
| CameSat027-469 | 0.0496 | 5.69 | 469 | 62.3 | CameSat110-145 | 0.0052 | 13.30 | 145 | 71.0 |
| CameSat028-154 | 0.0493 | 7.78 | 154 | 66.2 | CameSat111-160 | 0.0049 | 10.38 | 160 | 64.4 |
| CameSat029-325 | 0.0461 | 10.47 | 325 | 68.1 | CameSat112-147 | 0.0049 | 2.93 | 147 | 63.9 |
| CameSat030-132 | 0.0452 | 7.28 | 132 | 63.6 | CameSat113-76 | 0.0048 | 11.48 | 76 | 69.7 |
| CameSat031-281 | 0.0440 | 26.18 | 281 | 66.9 | CameSat114-144 | 0.0045 | 15.56 | 144 | 62.5 |
| CameSat032-166 | 0.0434 | 16.03 | 166 | 35.5 | CameSat115-14 | 0.0045 | 6.70 | 14 | 50.0 |
| CameSat033-351 | 0.0429 | 16.55 | 351 | 60.4 | CameSat116-29 | 0.0044 | 8.17 | 29 | 51.7 |
| CameSat034-191 | 0.0416 | 4.64 | 191 | 60.7 | CameSat117-149 | 0.0044 | 8.64 | 149 | 67.1 |
| CameSat035-5-tel | 0.0414 | 0.83 | 5 | 40.0 | CameSat118-144 | 0.0042 | 7.07 | 144 | 66.0 |
| CameSat036-144 | 0.0409 | 4.64 | 144 | 66.0 | CameSat119-140 | 0.0040 | 7.42 | 140 | 60.7 |
| CameSat037-1278 | 0.0405 | 2.77 | 1278 | 64.1 | CameSat120-286 | 0.0040 | 5.94 | 286 | 60.1 |
| CameSat038-50 | 0.0404 | 13.65 | 50 | 66.0 | CameSat121-162 | 0.0039 | 18.95 | 162 | 71.0 |
| CameSat039-400 | 0.0389 | 2.39 | 400 | 66.4 | CameSat122-132 | 0.0038 | 14.77 | 132 | 58.3 |
| CameSat040-67 | 0.0377 | 13.60 | 67 | 64.2 | CameSat123-161 | 0.0038 | 14.31 | 161 | 65.8 |
| CameSat041-142 | 0.0353 | 4.81 | 142 | 62.7 | CameSat124-120 | 0.0037 | 15.51 | 120 | 59.2 |
| CameSat042-250 | 0.0342 | 18.25 | 250 | 66.8 | CameSat125-309 | 0.0036 | 15.66 | 309 | 64.4 |
| CameSat043-151 | 0.0333 | 5.93 | 151 | 65.6 | CameSat126-509 | 0.0036 | 17.22 | 509 | 69.7 |
| CameSat044-2844 | 0.0322 | 11.06 | 2844 | 73.6 | CameSat127-410 | 0.0034 | 10.08 | 410 | 66.1 |
| CameSat045-143 | 0.0319 | 9.01 | 143 | 61.5 | CameSat128-148 | 0.0034 | 6.09 | 148 | 57.4 |
| CameSat046-23 | 0.0301 | 6.47 | 23 | 56.5 | CameSat129-91 | 0.0034 | 19.47 | 91 | 62.6 |
| CameSat047-389 | 0.0298 | 13.10 | 389 | 64.0 | CameSat130-147 | 0.0033 | 11.08 | 147 | 61.2 |
| CameSat048-293 | 0.0286 | 17.01 | 293 | 58.4 | CameSat131-149 | 0.0033 | 12.11 | 149 | 70.5 |
| CameSat049-20 | 0.0265 | 13.40 | 20 | 60.0 | CameSat132-429 | 0.0030 | 3.43 | 429 | 63.4 |
| CameSat050-140 | 0.0258 | 9.00 | 140 | 67.9 | CameSat133-142 | 0.0028 | 6.41 | 142 | 62.7 |
| CameSat051-3051 | 0.0255 | 8.78 | 3051 | 67.5 | CameSat134-166 | 0.0028 | 5.63 | 166 | 69.3 |
| CameSat052-143 | 0.0238 | 6.67 | 143 | 66.4 | CameSat135-148 | 0.0027 | 7.01 | 148 | 66.2 |
| CameSat053-153 | 0.0235 | 9.79 | 153 | 68.6 | CameSat136-255 | 0.0027 | 12.48 | 255 | 65.9 |
| CameSat054-164 | 0.0211 | 8.20 | 164 | 67.1 | CameSat137-360 | 0.0025 | 3.35 | 360 | 59.4 |
| CameSat055-30 | 0.0201 | 13.58 | 30 | 63.3 | CameSat138-156 | 0.0025 | 7.18 | 156 | 57.7 |
| CameSat056-42 | 0.0198 | 19.82 | 42 | 69.0 | CameSat139-369 | 0.0025 | 13.38 | 369 | 65.0 |
| CameSat057-147 | 0.0195 | 9.61 | 147 | 61.9 | CameSat140-84 | 0.0025 | 10.28 | 84 | 57.1 |
| CameSat058-280 | 0.0189 | 6.36 | 280 | 57.1 | CameSat141-163 | 0.0019 | 13.47 | 163 | 65.0 |
| CameSat059-128 | 0.0188 | 19.29 | 128 | 68.8 | CameSat142-408 | 0.0017 | 10.77 | 408 | 67.9 |
| CameSat060-141 | 0.0188 | 14.68 | 141 | 59.6 | CameSat143-184 | 0.0016 | 8.55 | 184 | 65.8 |
| CameSat061-230 | 0.0184 | 7.56 | 230 | 68.3 | CameSat144-265 | 0.0015 | 13.25 | 265 | 63.8 |
| CameSat062-142 | 0.0183 | 12.19 | 142 | 60.6 | CameSat145-168 | 0.0015 | 7.16 | 168 | 64.3 |
| CameSat063-2048 | 0.0182 | 5.04 | 2048 | 64.7 | CameSat146-139 | 0.0015 | 8.36 | 139 | 74.1 |
| CameSat064-104 | 0.0182 | 2.78 | 104 | 51.0 | CameSat147-154 | 0.0014 | 13.08 | 154 | 73.4 |
| CameSat065-148 | 0.0175 | 15.05 | 148 | 67.6 | CameSat148-468 | 0.0014 | 4.06 | 468 | 59.2 |
| CameSat066-149 | 0.0175 | 6.94 | 149 | 66.4 | CameSat149-141 | 0.0014 | 10.48 | 141 | 51.8 |
| CameSat067-168 | 0.0155 | 23.71 | 168 | 69.0 | CameSat150-147 | 0.0013 | 4.43 | 147 | 70.1 |
| CameSat068-153 | 0.0147 | 8.35 | 153 | 58.8 | CameSat151-300 | 0.0013 | 7.26 | 300 | 63.0 |
| CameSat069-142 | 0.0144 | 13.63 | 142 | 55.6 | CameSat152-228 | 0.0012 | 5.41 | 228 | 59.6 |
| CameSat070-13 | 0.0142 | 14.35 | 13 | 53.8 | CameSat153-498 | 0.0012 | 4.75 | 498 | 60.8 |
| CameSat071-20 | 0.0137 | 17.80 | 20 | 70.0 | CameSat154-59 | 0.0012 | 9.39 | 59 | 72.9 |
| CameSat072-193 | 0.0132 | 8.71 | 193 | 65.3 | CameSat155-496 | 0.0011 | 2.09 | 496 | 66.7 |
| CameSat073-20 | 0.0130 | 22.31 | 20 | 45.0 | CameSat156-216 | 0.0011 | 2.84 | 216 | 60.2 |
| CameSat074-1625 | 0.0116 | 1.77 | 1625 | 63.1 | CameSat157-476 | 0.0011 | 2.76 | 476 | 62.8 |
| CameSat075-275 | 0.0116 | 7.13 | 275 | 62.9 | CameSat158-143 | 0.0011 | 9.02 | 143 | 65.0 |
| CameSat076-243 | 0.0114 | 12.69 | 243 | 61.3 | CameSat159-272 | 0.0010 | 2.51 | 272 | 63.6 |
| CameSat077-1817 | 0.0103 | 1.95 | 1817 | 63.1 | CameSat160-36 | 0.0009 | 12.37 | 36 | 63.9 |
| CameSat078-138 | 0.0102 | 14.13 | 138 | 67.4 | CameSat161-127 | 0.0009 | 2.42 | 127 | 65.4 |
| CameSat079-143 | 0.0102 | 6.60 | 143 | 69.2 | CameSat162-133 | 0.0008 | 4.60 | 133 | 70.7 |
| CameSat080-162 | 0.0098 | 16.90 | 162 | 67.9 | CameSat163-416 | 0.0008 | 3.37 | 416 | 60.6 |
| CameSat081-125 | 0.0093 | 16.48 | 125 | 58.4 | CameSat164-112 | 0.0006 | 8.95 | 112 | 67.9 |
| CameSat082-204 | 0.0091 | 4.98 | 204 | 57.8 | CameSat165-105 | 0.0002 | 8.18 | 105 | 60.0 |
| CameSat083-100 | 0.0091 | 15.98 | 100 | 56.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rico-Porras, J.M.; Mora, P.; Palomeque, T.; Montiel, E.E.; Cabral-de-Mello, D.C.; Lorite, P. Heterochromatin Is Not the Only Place for satDNAs: The High Diversity of satDNAs in the Euchromatin of the Beetle Chrysolina americana (Coleoptera, Chrysomelidae). Genes 2024, 15, 395. https://doi.org/10.3390/genes15040395
Rico-Porras JM, Mora P, Palomeque T, Montiel EE, Cabral-de-Mello DC, Lorite P. Heterochromatin Is Not the Only Place for satDNAs: The High Diversity of satDNAs in the Euchromatin of the Beetle Chrysolina americana (Coleoptera, Chrysomelidae). Genes. 2024; 15(4):395. https://doi.org/10.3390/genes15040395
Chicago/Turabian StyleRico-Porras, José M., Pablo Mora, Teresa Palomeque, Eugenia E. Montiel, Diogo C. Cabral-de-Mello, and Pedro Lorite. 2024. "Heterochromatin Is Not the Only Place for satDNAs: The High Diversity of satDNAs in the Euchromatin of the Beetle Chrysolina americana (Coleoptera, Chrysomelidae)" Genes 15, no. 4: 395. https://doi.org/10.3390/genes15040395