
Integrating Genomic and Chromosomal Data: A Cytogenetic Study of Transancistrus santarosensis (Loricariidae: Hypostominae) with Characterization of a ZZ/ZW Sex Chromosome System
 , ,
, ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Molecular Identification of Samples and Phylogenetic Reconstruction
2.3. Cytogenetic Procedures
3. Results
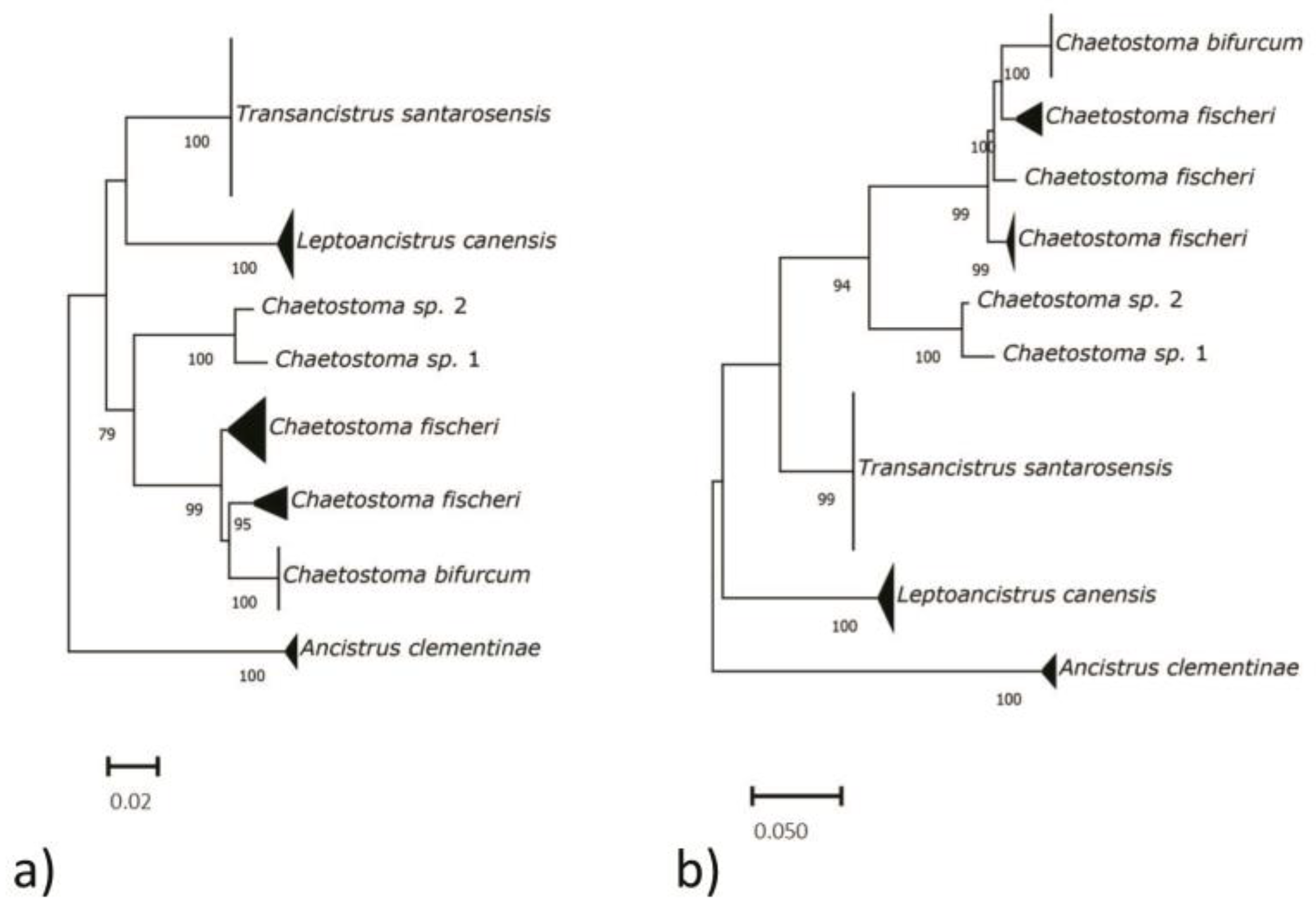
3.1. COI Identification and Phylogenetic Reconstruction
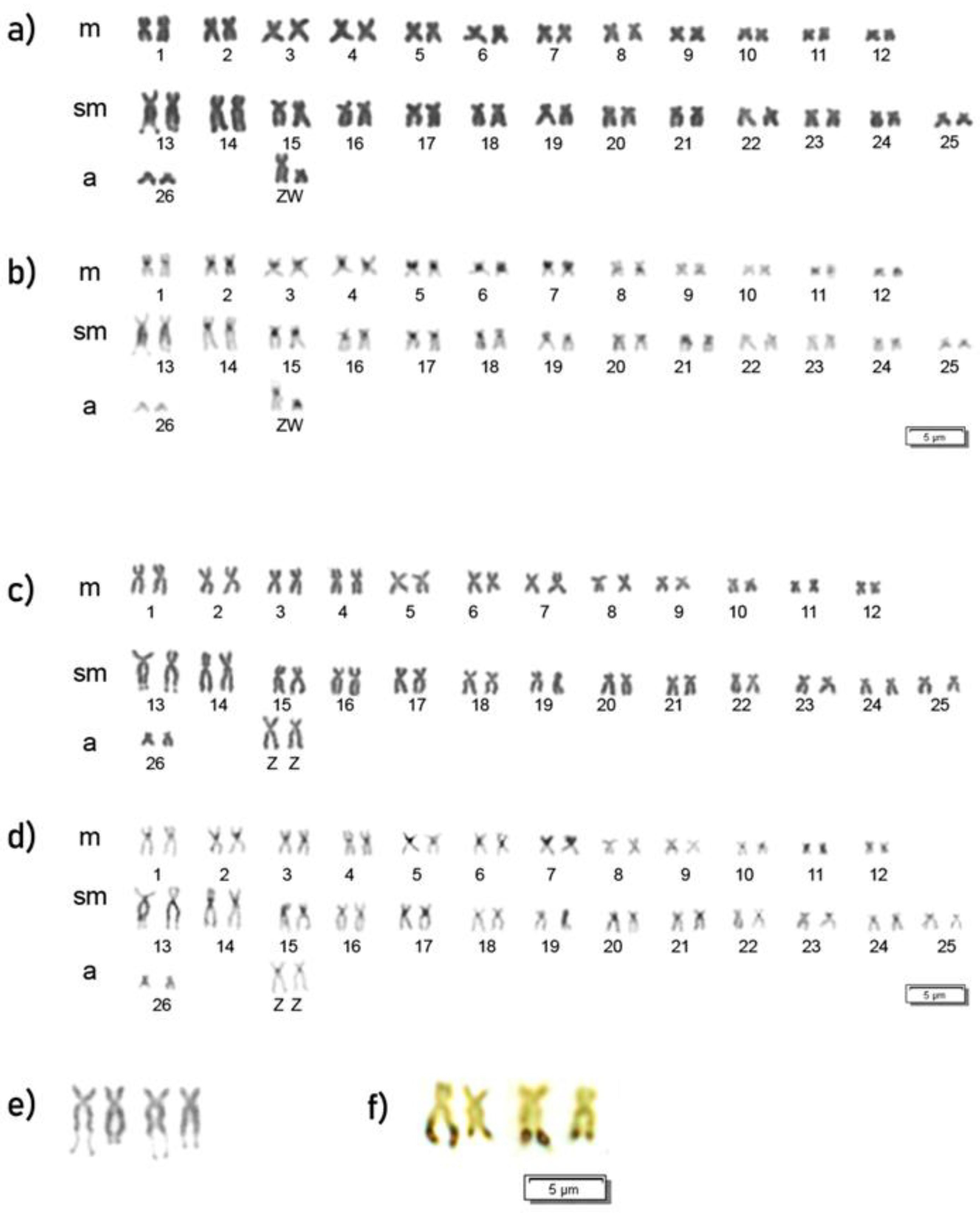
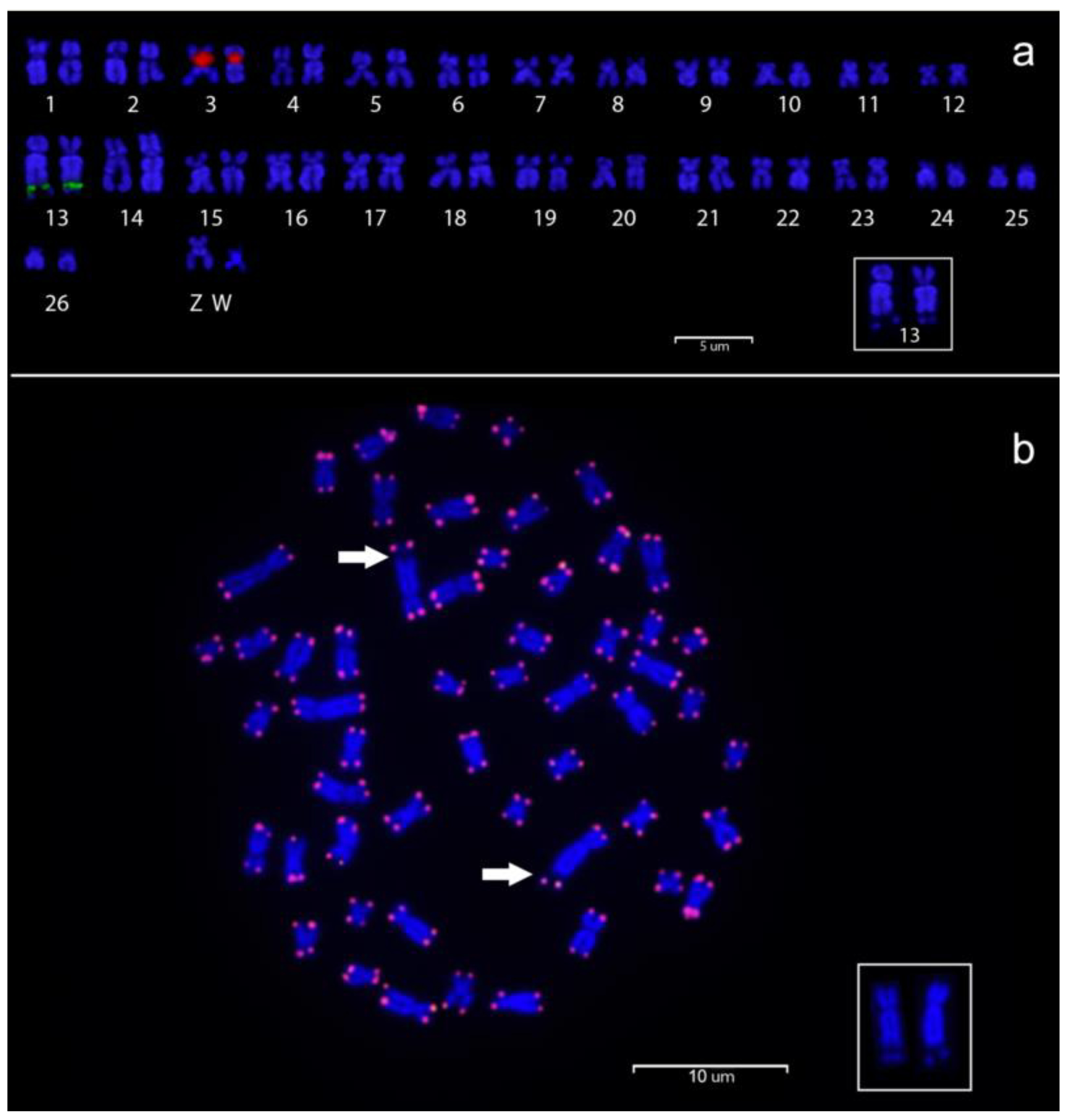
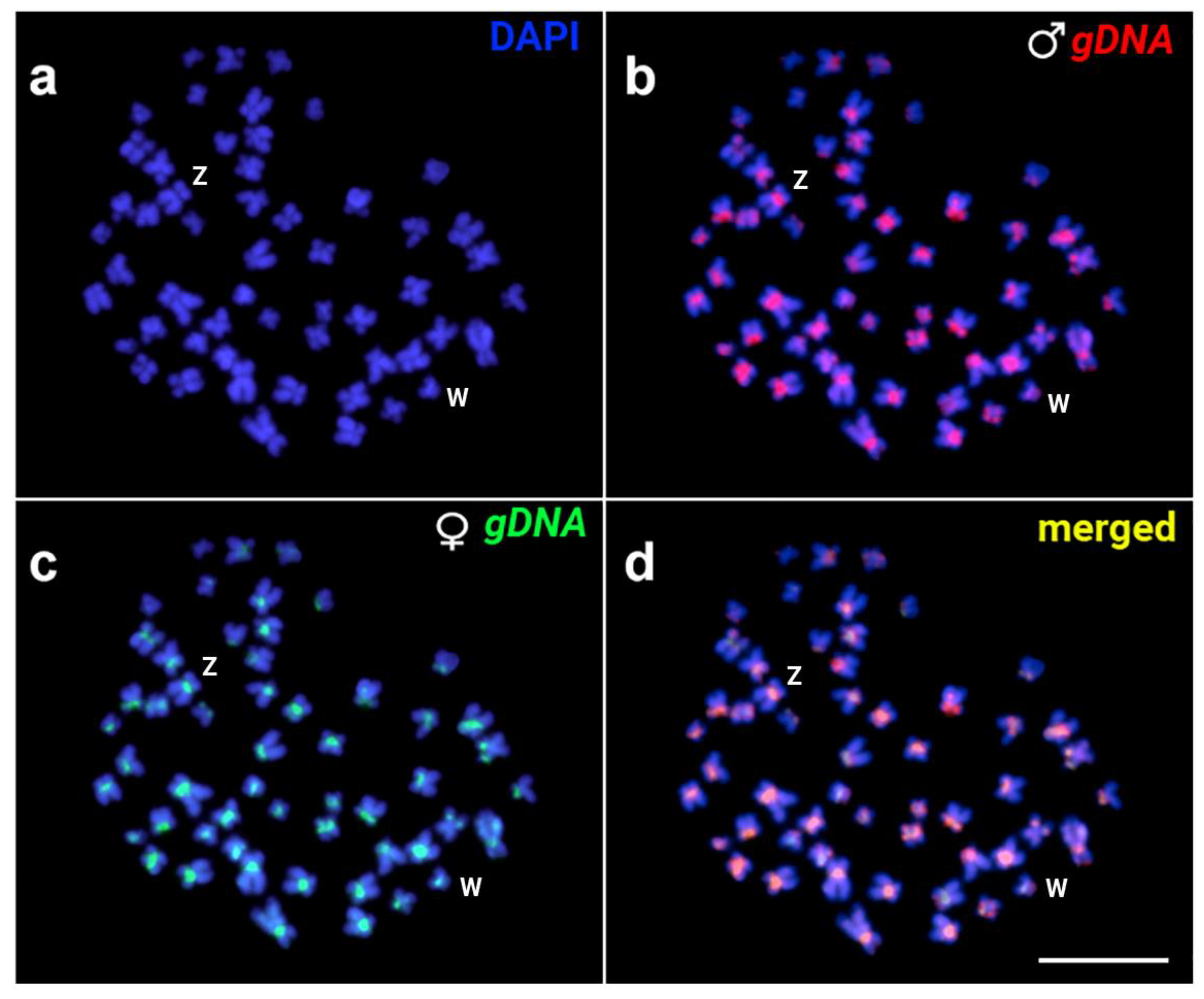
3.2. Cytogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WWF. Living Planet Report—Construire Una Societá Naturepositive; Almond, R.E.A., Grooten, M., Juffe Bignoli, D., Petersen, T., Eds.; WWF: Gland, Switzerland, 2022. [Google Scholar]
- Tisseuil, C.; Cornu, J.F.; Beauchard, O.; Brosse, S.; Darwall, W.; Holland, R.; Hugueny, B.; Tedesco, P.A.; Oberdorff, T. Global Diversity Patterns and Cross-Taxa Convergence in Freshwater Systems. J. Anim. Ecol. 2013, 82, 365–376. [Google Scholar] [CrossRef]
- Reis, R.E.; Albert, J.S.; Di Dario, F.; Mincarone, M.M.; Petry, P.; Rocha, L.A. Fish Biodiversity and Conservation in South America. J. Fish Biol. 2016, 89, 12–47. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological Identifications through DNA Barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Bertollo, L.A.C.; Moreira-Filho, O.; Galetti, P.M. Cytogenetics and Taxonomy: Considerations Based on Chromosome Studies of Freshwater Fish. J. Fish Biol. 1986, 28, 153–159. [Google Scholar] [CrossRef]
- Rossi, A.R. Fish Cytogenetics: Present and Future. Genes 2021, 12, 983. [Google Scholar] [CrossRef]
- Pereira, L.H.G.; Castro, J.R.C.; Vargas, P.M.H.; Gomez, J.A.M.; Oliveira, C. The Use of an Integrative Approach to Improve Accuracy of Species Identification and Detection of New Species in Studies of Stream Fish Diversity. Genetica 2021, 149, 103–116. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Yano, C.F.; Sember, A.; Bertollo, L.A.C. Chromosomal Evolution in Lower Vertebrates: Sex Chromosomes in Neotropical Fishes. Genes 2017, 8, 258. [Google Scholar] [CrossRef]
- Schemberger, M.O.; Nascimento, V.D.; Coan, R.; Ramos, É.; Nogaroto, V.; Ziemniczak, K.; Valente, G.T.; Moreira-Filho, O.; Martins, C.; Vicari, M.R. DNA Transposon Invasion and Microsatellite Accumulation Guide W Chromosome Differentiation in a Neotropical Fish Genome. Chromosoma 2019, 128, 547–560. [Google Scholar] [CrossRef]
- Symonová, R.; Flajšhans, M.; Sember, A.; Havelka, M.; Gela, D.; Kořínková, T.; Rodina, M.; Rábová, M.; Ráb, P. Molecular Cytogenetics in Artificial Hybrid and Highly Polyploid Sturgeons: An Evolutionary Story Narrated by Repetitive Sequences. Cytogenet. Genome Res. 2013, 141, 153–162. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Wagner, F.M.; Ferreira, A.R.; Bertollo, L.A.C. Chromosomes as Tools for Discovering Biodiversity—The Case of Erythrinidae Fish Family. In Recent Trends in Cytogenetic Studies; Tirunilai, P., Ed.; InTech Publisher: Rijeka, Croatia, 2012; pp. 125–146. [Google Scholar] [CrossRef]
- Arai, R. Fish Karyotypes: A Check List; Springer Science & Business Media: Tokio, Japan, 2011; ISBN 9784431538776. [Google Scholar]
- Formenti, G.; Theissinger, K.; Fernandes, C.; Bista, I.; Bombarely, A.; Bleidorn, C.; Ciofi, C.; Crottini, A.; Godoy, J.A.; Höglund, J.; et al. The Era of Reference Genomes in Conservation Genomics. Trends Ecol. Evol. 2022, 37, 197–202. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Moreira-Filho, O.; Almeida-Toledo, L.F.; Bertollo, L.A.C. The Contrasting Role of Heterochromatin in the Differentiation of Sex Chromosomes: An Overview from Neotropical Fishes. J. Fish Biol. 2012, 80, 2125–2139. [Google Scholar] [CrossRef]
- Pertoldi, C.; Randi, E.; Ruiz-González, A.; Vergeer, P.; Ouborg, J. How Can Genomic Tools Contribute to the Conservation of Endangered Organisms. Int. J. Genom. Proteom. 2016, 2016, 4712487. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Eschemeyer’s Catalog of Fishes: Genera, Species. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 25 May 2023).
- Roxo, F.F.; Ochoa, L.E.; Sabaj, M.H.; Lujan, N.K.; Covain, R.; Silva, G.S.C.; Melo, B.F.; Albert, J.S.; Chang, J.; Foresti, F.; et al. Phylogenomic Reappraisal of the Neotropical Catfish Family Loricariidae (Teleostei: Siluriformes) Using Ultraconserved Elements. Mol. Phylogenet. Evol. 2019, 135, 148–165. [Google Scholar] [CrossRef]
- Malabarba, L.R.; Malabarba, M.C. Phylogeny and Classification of Neotropical Fish. In Biology and Physiology of Freshwater Neotropical Fish; Baldisserotto, B., Urbinati, E.C., Cyrino, J.E.P., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 1–19. ISBN 9780128158722. [Google Scholar]
- Lujan, N.K.; Armbruster, J.W.; Lovejoy, N.R.; López-Fernández, H. Multilocus Molecular Phylogeny of the Suckermouth Armored Catfishes (Siluriformes: Loricariidae) with a Focus on Subfamily Hypostominae. Mol. Phylogenet. Evol. 2015, 82 Pt A, 269–288. [Google Scholar] [CrossRef]
- Lujan, N.K.; Meza-Vargas, V.; Barriga-Salazar, R. Two New Chaetostoma Group (Loricariidae: Hypostominae) Sister Genera from Opposite Sides of the Andes Mountains in Ecuador, with the Description of One New Species. Copeia 2015, 103, 651–663. [Google Scholar] [CrossRef]
- Lujan, N.K.; Meza-Vargas, V.; Astudillo-Clavijo, V.; Barriga-Salazar, R.; López-Fernández, H. A Multilocus Molecular Phylogeny for Chaetostoma Clade Genera and Species with a Review of Chaetostoma (Siluriformes: Loricariidae) from the Central Andes. Copeia 2015, 103, 664–701. [Google Scholar] [CrossRef]
- Glugoski, L.; Deon, G.; Schott, S.; Vicari, M.R.; Nogaroto, V.; Moreira-Filho, O. Comparative Cytogenetic Analyses in Ancistrus Species (Siluriformes: Loricariidae). Neotrop. Ichthyol. 2020, 18, e200013. [Google Scholar] [CrossRef]
- Deon, G.A.; Glugoski, L.; Vicari, M.R.; Nogaroto, V.; Sassi, F.M.C.; Cioffi, M.B.; Liehr, T.; Bertollo, L.A.C.; Moreira-Filho, O. Highly Rearranged Karyotypes and Multiple Sex Chromosome Systems in Armored Catfishes from the Genus Harttia (Teleostei, Siluriformes). Genes 2020, 11, 1366. [Google Scholar] [CrossRef]
- Santos da Silva, K.; Glugoski, L.; Vicari, M.R.; de Souza, A.C.P.; Noronha, R.C.R.; Pieczarka, J.C.; Nagamachi, C.Y. Chromosomal Diversification in Ancistrus Species (Siluriformes: Loricariidae) Inferred from Repetitive Sequence Analysis. Front. Genet. 2022, 13, 838462. [Google Scholar] [CrossRef]
- Nirchio, M.; Oliveira, C.; Cioffi, M.B.; Sassi, F.M.C.; Valdiviezo, J.; Paim, F.G.; Soares, L.B.; Rossi, A.R. Occurrence of Sex Chromosomes in Fish of the Genus Ancistrus with a New Description of Multiple Sex Chromosomes in the Ecuadorian Endemic Ancistrus clementinae (Loricariidae). Genes 2023, 14, 306. [Google Scholar] [CrossRef]
- Sassi, F.M.C.; Deon, G.A.; Moreira-Filho, O.; Vicari, M.R.; Bertollo, L.A.C.; Liehr, T.; Oliveira, E.A.; Cioffi, M.B. Multiple Sex Chromosomes and Evolutionary Relationships in Amazonian Catfishes: The Outstanding Model of the Genus Harttia (Siluriformes: Loricariidae). Genes 2020, 11, 1179. [Google Scholar] [CrossRef] [PubMed]
- Sassi, F.M.C.; Moreira-Filho, O.; Deon, G.A.; Sember, A.; Bertollo, L.A.C.; Liehr, T.; Oliveira, V.C.S.; Viana, P.F.; Feldberg, E.; Vicari, M.R.; et al. Adding New Pieces to the Puzzle of Karyotype Evolution in Harttia (Siluriformes, Loricariidae): Investigation of Amazonian Species. Biology 2021, 10, 922. [Google Scholar] [CrossRef]
- Santos da Silva, K.; de Souza, A.C.P.; Pety, A.M.; Noronha, R.C.R.; Vicari, M.R.; Pieczarka, J.C.; Nagamachi, C.Y. Comparative Cytogenetics Analysis Among Peckoltia Species (Siluriformes, Loricariidae): Insights on Karyotype Evolution and Biogeography in the Amazon Region. Front. Genet. 2021, 12, 779464. [Google Scholar] [CrossRef]
- Rubert, M.; da Rosa, R.; Jerep, F.C.; Bertollo, L.A.C.; Giuliano-Caetano, L. Cytogenetic Characterization of Four Species of the Genus Hypostomus Lacépède, 1803 (Siluriformes, Loricariidae) with Comments on Its Chromosomal Diversity. Comp. Cytogenet. 2011, 5, 397–410. [Google Scholar] [CrossRef]
- Marajó, L.; Viana, P.F.; Ferreira, M.; Py-Daniel, L.H.R.; Feldberg, E. Cytogenetics of Two Farlowella Species (Loricariidae: Loricariinae): Implications on the Taxonomic Status of the Species. Neotrop. Ichthyol. 2018, 16. [Google Scholar] [CrossRef]
- Schott, S.C.Q.; Glugoski, L.; Azambuja, M.; Moreira-Filho, O.; Vicari, M.R.; Nogaroto, V. Comparative Cytogenetic and Sequence Analysis of U Small Nuclear RNA Genes in Three Ancistrus Species (Siluriformes: Loricariidae). Zebrafish 2022, 19, 200–209. [Google Scholar] [CrossRef]
- Takagui, F.H.; Baumgärtner, L.; Baldissera, J.N.; Laridondo Lui, R.; Margarido, V.P.; Fonteles, S.B.A.; Garcia, C.; Birindelli, J.O.; Moreira-Filho, O.; Almeida, F.S.; et al. Chromosomal Diversity of Thorny Catfishes (Siluriformes-Doradidae): A Case of Allopatric Speciation Among Wertheimerinae Species of São Francisco and Brazilian Eastern Coastal Drainages. Zebrafish 2019, 16, 477–485. [Google Scholar] [CrossRef]
- Tan, M.; Armbruster, J.W. Cordylancistrus santarosensis (Siluriformes: Loricariidae), a New Species with Unique Snout Deplatation from the Río Santa Rosa, Ecuador. Zootaxa 2012, 3243, 52–58. [Google Scholar] [CrossRef]
- IUCN Red List Committee. Available online: https://www.iucnredlist.org (accessed on 7 June 2023).
- Lyons, T.J. Transancistrus santarosensis. Available online: https://www.iucnredlist.org/es/species/66601846/176676466 (accessed on 7 June 2023).
- Artoni, R.F.; Bertollo, L.A. Trends in the Karyotype Evolution of Loricariidae Fish (Siluriformes). Hereditas 2001, 134, 201–210. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, R.R.; Feldberg, E.; Dos Anjos, M.B.; Zuanon, J. Mechanisms of Chromosomal Evolution and Its Possible Relation to Natural History Characteristics in Ancistrus Catfishes (Siluriformes: Loricariidae). J. Fish Biol. 2009, 75, 2209–2225. [Google Scholar] [CrossRef]
- Mariotto, S.; Miyazawa, C.S. Ancistrus cf. dubius (Siluriformes, Ancistrinae), a Complex of Species. 1. Chromosomic Characterization of Four Populations and Occurrence of Sexual Chromosomes of Type XX/XY, in the Pantanal Basin of Mato Grosso, Brazil. Caryologia 2006, 59, 299–304. [Google Scholar] [CrossRef]
- Sandra, M.; Artoni, R.F.; Miyazawa, C.S. Occurrence of Sexual Chromosome, of the Type ZZ/ZW, in Ancistrus cf. dubius (Loricariidae, Ancistrinae) of the Paraguay River Basin, Mato Grosso, Brazil. Caryologia 2004, 57, 327–331. [Google Scholar] [CrossRef]
- Mariotto, S.; Centofante, L.; Miyazawa, C.S.; Bertollo, L.A.C.; Moreira Filho, O. Chromosome Polymorphism in Ancistrus cuiabae Knaack, 1999 (Siluriformes: Loricariidae: Ancistrini). Neotrop. Ichthyol. 2009, 7, 595–600. [Google Scholar] [CrossRef]
- de Oliveira, R.R.; Feldberg, E.; dos Anjos, M.B.; Zuanon, J. Occurrence of Multiple Sexual Chromosomes (XX/XY1Y2 and Z1Z1Z2Z2/Z1Z2W1W2) in Catfishes of the Genus Ancistrus (Siluriformes: Loricariidae) from the Amazon Basin. Genetica 2008, 134, 243–249. [Google Scholar] [CrossRef]
- Mariotto, S.; Centofante, L.; Moreira-Filho, O. Diversity and Chromosomal Evolution in the genus Ancistrus Kner, 1854 (Loricariidae: Ancistrini) from Three Hydrographic Basins of Mato Grosso State, Brazil. Neotrop. Ichthyol. 2013, 11, 125–131. [Google Scholar] [CrossRef]
- de Moraes, R.L.R.; Sember, A.; Bertollo, L.A.C.; de Oliveira, E.A.; Ráb, P.; Hatanaka, T.; Marinho, M.M.F.; Liehr, T.; Al-Rikabi, A.B.H.; Feldberg, E.; et al. Comparative Cytogenetics and Neo-Y Formation in Small-Sized Fish Species of the Genus Pyrrhulina (Characiformes, Lebiasinidae). Front. Genet. 2019, 10, 678. [Google Scholar] [CrossRef]
- Sember, A.; Bertollo, L.A.C.; Ráb, P.; Yano, C.F.; Hatanaka, T.; de Oliveira, E.A.; Cioffi, M.B. Sex Chromosome Evolution and Genomic Divergence in the Fish Hoplias malabaricus (Characiformes, Erythrinidae). Front. Genet. 2018, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Yano, C.F.; Bertollo, L.A.C.; Cioffi, M.B. Fish-FISH: Molecular Cytogenetics in Fish Species. In Fluorescence In Situ Hybridization (FISH): Application Guide; Liehr, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 429–443. ISBN 9783662529591. [Google Scholar]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA Barcoding Australia’s Fish Species. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Milana, V.; Fusari, A.; Rossi, A.; Sola, L. Molecular and Morphological Identification of an Uncommon Centrolophid Fish. Cent. Eur. J. Biol. 2011, 6, 440–445. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 1, 2.3.1–2.3.22. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Foresti, F.; Almeida Toledo, L.F.; Toledo, S.A. Polymorphic Nature of Nucleolus Organizer Regions in Fishes. Cytogenet. Cell Genet. 1981, 31, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Sumner, A.T. A Simple Technique for Demonstrating Centromeric Heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef] [PubMed]
- Howell, W.M.; Black, D.A. Controlled Silver-Staining of Nucleolus Organizer Regions with a Protective Colloidal Developer: A 1-Step Method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for Centromeric Position on Chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Pinkel, D.; Straume, T.; Gray, J.W. Cytogenetic Analysis Using Quantitative, High-Sensitivity, Fluorescence Hybridization. Proc. Natl. Acad. Sci. USA 1986, 83, 2934–2938. [Google Scholar] [CrossRef]
- Sassi, F.M.C.; Toma, G.A.; Cioffi, M.B. FISH—In Fish Chromosomes. In Cytogenetics and Molecular Cytogenetics; Liehr, T., Ed.; CRC Press: Boca Raton, FL, USA, 2022; pp. 281–296. ISBN 9781003223658. [Google Scholar]
- Utsunomia, R.; de Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Araya-Jaime, C.; Pansonato-Alves, J.C.; Scacchetti, P.C.; Hashimoto, D.T.; Oliveira, C.; Trifonov, V.A.; Porto-Foresti, F.; et al. Uncovering the Ancestry of B Chromosomes in Moenkhausia sanctaefilomenae (Teleostei, Characidae). PLoS ONE 2016, 11, e0150573. [Google Scholar] [CrossRef]
- Pendas, A.M.; Moran, P.; Martinez, J.L.; Garcia-Vazquez, E. Applications of 5S rDNA in Atlantic Salmon, Brown Trout, and in Atlantic Salmon Brown Trout Hybrid Identification. Mol. Ecol. 1995, 4, 275–276. [Google Scholar] [CrossRef] [PubMed]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved Telomere Detection Using a Telomere Repeat Probe (TTAGGG)n Generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; ISBN 9780879695767. [Google Scholar]
- Yang, F.; Trifonov, V.; Ng, B.L.; Kosyakova, N.; Carter, N.P. Generation of Paint Probes by Flow-Sorted and Microdissected Chromosomes. In Fluorescence In Situ Hybridization (FISH)—Application Guide; Liehr, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 35–52. ISBN 9783540705819. [Google Scholar]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Gregory, T.R. Animal Genome Size Database. Available online: http://www.genomesize.com (accessed on 17 January 2023).
- Yoshida, K.; Kitano, J. Tempo and Mode in Karyotype Evolution Revealed by a Probabilistic Model Incorporating Both Chromosome Number and Morphology. PLoS Genet. 2021, 17, e1009502. [Google Scholar] [CrossRef]
- Devlin, R.H.; Nagahama, Y. Sex Determination and Sex Differentiation in Fish: An Overview of Genetic, Physiological, and Environmental Influences. Aquaculture 2002, 208, 191–364. [Google Scholar] [CrossRef]
- Guiguen, Y.; Fostier, A.; Herpin, A. Sex Determination and Differentiation in Fish. In Sex Control in Aquaculture; Wang, H.-P., Piferrer, F., Chen, S.-L., Shen, Z.-G., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2018; pp. 35–63. ISBN 9781119127291. [Google Scholar]
- Kuwamura, T.; Sunobe, T.; Sakai, Y.; Kadota, T.; Sawada, K. Hermaphroditism in Fishes: An Annotated List of Species, Phylogeny, and Mating System. Ichthyol. Res. 2020, 67, 341–360. [Google Scholar] [CrossRef]
- Schartl, M.; Schmid, M.; Nanda, I. Dynamics of Vertebrate Sex Chromosome Evolution: From Equal Size to Giants and Dwarfs. Chromosoma 2016, 125, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.-G.; Wang, H.-P. Environmental Sex Determination and Sex Differentiation in Teleosts—How Sex Is Established. In Sex Control in Aquaculture; Wang, H.-P., Piferrer, F., Chen, S.-L., Shen, Z.-G., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2018; pp. 85–115. ISBN 9781119127291. [Google Scholar]
- Sember, A.; Nguyen, P.; Perez, M.F.; Altmanová, M.; Ráb, P.; Cioffi, M.B. Multiple Sex Chromosomes in Teleost Fishes from a Cytogenetic Perspective: State of the Art and Future Challenges. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20200098. [Google Scholar] [CrossRef]
- Cereali, S.S.; Pomini, E.; Rosa, R.; Zawadzki, C.H.; Froehlich, O.; Giuliano-Caetano, L. Karyotype Description of Two Species of Hypostomus (Siluriformes, Loricariidae) of the Planalto Da Bodoquena, Brazil. Genet. Mol. Res. 2008, 7, 583–591. [Google Scholar] [CrossRef]
- Artoni, R.F.; Venere, P.C.; Bertollo, L.A.C. A Heteromorphic ZZ/ZW Sex Chromosome System in Fish, Genus Hypostomus (Loricariidae). Cytologia 1998, 63, 421–425. [Google Scholar] [CrossRef]
- Rocha-Reis, D.A.; de Oliveira Brandão, K.; de Almeida-Toledo, L.F.; Pazza, R.; Kavalco, K.F. The Persevering Cytotaxonomy: Discovery of a Unique XX/XY Sex Chromosome System in Catfishes Suggests the Existence of a New, Endemic and Rare Species. Cytogenet. Genome Res. 2018, 156, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Marajó, L.; Viana, P.F.; Ferreira, A.M.V.; Py-Daniel, L.H.R.; Cioffi, M.B.; Sember, A.; Feldberg, E. Chromosomal Rearrangements and the First Indication of an ♀X1 X1 X2 X2/♂X1 X2 Y Sex Chromosome System in Rineloricaria Fishes (Teleostei: Siluriformes). J. Fish Biol. 2023, 102, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Ziemniczak, K.; Barros, A.V.; Rosa, K.O.; Nogaroto, V.; Almeida, M.C.; Cestari, M.M.; Moreira-Filho, O.; Artoni, R.F.; Vicari, M.R. Comparative Cytogenetics of Loricariidae (Actinopterygii: Siluriformes): Emphasis in Neoplecostominae and Hypoptopomatinae. Ital. J. Zool. 2012, 79, 492–501. [Google Scholar] [CrossRef]
- Favarato, R.M.; da Silva, M.; de Oliveira, R.R.; Artoni, R.F.; Feldberg, E.; Matoso, D.A. Cytogenetic Diversity and the Evolutionary Dynamics of rDNA Genes and Telomeric Sequences in the Ancistrus Genus (Loricariidae: Ancistrini). Zebrafish 2016, 13, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Prizon, A.C.; Borin-Carvalho, L.A.; Bruschi, D.P.; Otávio Ribeiro, M.; Magrinelli Barbosa, L.; de Brito Ferreira, G.E.; Cius, A.; Zawadzki, C.H.; de Brito Portela-Castro, A.L. Cytogenetic Data on Ancistrus sp. (Siluriformes, Loricariidae) of the Paraguay River Basin (MS) Sheds Light on Intrageneric Karyotype Diversification. Comp. Cytogenet. 2016, 10, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Pety, A.M.; Cardoso, A.L.; Nagamachi, C.Y.; Pieczarka, J.C.; de Sousa, L.M.; Noronha, R.C.R. In Situ Localization of Ribosomal Sites in Peckoltia and Ancistomus (Loricariidae: Hypostominae) from the Amazon Basin. Zebrafish 2018, 15, 263–269. [Google Scholar] [CrossRef]
- Stornioli, J.H.F.; Goes, C.A.G.; Calegari, R.M.; Dos Santos, R.Z.; Giglio, L.M.; Foresti, F.; Oliveira, C.; Penitente, M.; Porto-Foresti, F.; Utsunomia, R. The B Chromosomes of Prochilodus lineatus (Teleostei, Characiformes) Are Highly Enriched in Satellite DNAs. Cells 2021, 10, 1527. [Google Scholar] [CrossRef]
- Kretschmer, R.; Goes, C.A.G.; Bertollo, L.A.C.; Ezaz, T.; Porto-Foresti, F.; Toma, G.A.; Utsunomia, R.; Cioffi, M.B. Satellitome Analysis Illuminates the Evolution of ZW Sex Chromosomes of Triportheidae Fishes (Teleostei: Characiformes). Chromosoma 2022, 131, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.S.; Benjamin, C.; Rodriguez, J.; Dosik, H. Population Heteromorphisms of Ag-Stained Nucleolus Organizer Regions (NORs) in the Acrocentric Chromosomes of East Indians. Hum. Genet. 1981, 59, 412–415. [Google Scholar] [CrossRef]
- Galetti, P.M.; Foresti, F.; Bertollo, L.A.C.; Moreira, O. Characterization of Eight Species of Anostomidae (Cypriniformes) Fish on the Basis of the Nucleolar Organizing Region. Caryologia 1984, 37, 401–406. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, Y.; Wang, Q.; Li, L.; Hu, Y.; Wu, Y.; Gautam, M.; Li, L. R-Loops Mediate Transcription-Associated Formation of Human rDNA Secondary Constrictions. J. Cell. Biochem. 2021, 122, 1517–1533. [Google Scholar] [CrossRef]
- Symonová, R.; Majtánová, Z.; Arias-Rodriguez, L.; Mořkovský, L.; Kořínková, T.; Cavin, L.; Pokorná, M.J.; Doležálková, M.; Flajšhans, M.; Normandeau, E.; et al. Genome Compositional Organization in Gars Shows More Similarities to Mammals than to Other Ray-Finned Fish. J. Exp. Zool. B Mol. Dev. Evol. 2017, 328, 607–619. [Google Scholar] [CrossRef]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary Trends in Animal Ribosomal DNA Loci: Introduction to a New Online Database. Chromosoma 2018, 127, 141–150. [Google Scholar] [CrossRef]
- Janssen, A.; Colmenares, S.U.; Karpen, G.H. Heterochromatin: Guardian of the Genome. Annu. Rev. Cell Dev. Biol. 2018, 34, 265–288. [Google Scholar] [CrossRef]
- Allshire, R.C.; Madhani, H.D. Ten Principles of Heterochromatin Formation and Function. Nat. Rev. Mol. Cell Biol. 2018, 19, 229–244. [Google Scholar] [CrossRef]
- John, B.; Miklos, G.L. Functional Aspects of Satellite DNA and Heterochromatin. Int. Rev. Cytol. 1979, 58, 1–114. [Google Scholar] [CrossRef] [PubMed]
- Henning, F.; Moysés, C.B.; Calcagnotto, D.; Meyer, A.; de Almeida-Toledo, L.F. Independent Fusions and Recent Origins of Sex Chromosomes in the Evolution and Diversification of Glass Knife Fishes (Eigenmannia). Heredity 2011, 106, 391–400. [Google Scholar] [CrossRef]
- Yano, C.F.; Bertollo, L.A.C.; Molina, W.F.; Liehr, T.; Cioffi, M.B. Genomic Organization of Repetitive DNAs and Its Implications for Male Karyotype and the Neo-Y Chromosome Differentiation in Erythrinus erythrinus (Characiformes, Erythrinidae). Comp. Cytogenet. 2014, 8, 139–151. [Google Scholar] [CrossRef]
- Ziemniczak, K.; Traldi, J.B.; Nogaroto, V.; Almeida, M.C.; Artoni, R.F.; Moreira-Filho, O.; Vicari, M.R. In Situ Localization of (GATA)n and (TTAGGG)n Repeated DNAs and W Sex Chromosome Differentiation in Parodontidae (Actinopterygii: Characiformes). Cytogenet. Genome Res. 2014, 144, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Schemberger, M.O.; Bellafronte, E.; Nogaroto, V.; Almeida, M.C.; Schühli, G.S.; Artoni, R.F.; Moreira-Filho, O.; Vicari, M.R. Differentiation of Repetitive DNA Sites and Sex Chromosome Systems Reveal Closely Related Group in Parodontidae (Actinopterygii: Characiformes). Genetica 2011, 139, 1499–1508. [Google Scholar] [CrossRef]
- Paim, F.; Nirchio, M.; Oliveira, C.; Rossi, A.R. Sex Chromosomes and Internal Telomeric Sequences in Dormitator latifrons (Richardson 1844) (Eleotridae: Eleotrinae): An Insight into Their Origin in the Genus. Genes 2020, 11, 659. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.B.; Barbosa, P.; Nogaroto, V.; Almeida, M.C.; Artoni, R.F.; Scacchetti, P.C.; Pansonato-Alves, J.C.; Foresti, F.; Moreira-Filho, O.; Vicari, M.R. Chromosomal Spreading of Microsatellites and (TTAGGG)n Sequences in the Characidium zebra and C. gomesi Genomes (Characiformes: Crenuchidae). Cytogenet. Genome Res. 2016, 149, 182–190. [Google Scholar] [CrossRef]
- De Oliveira, R.R.; Feldberg, E.; dos Anjos, M.B.; Zuanon, J. Karyotype Characterization and ZZ/ZW Sex Chromosome Heteromorphism in Two Species of the Catfish Genus Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from the Amazon Basin. Neotrop. Ichthyol. 2007, 5, 301–306. [Google Scholar] [CrossRef]
- Konerat, J.T.; Bueno, V.; Margarido, V.P.; Portela-Castro, A.L.B.; Martins-Santos, I.C. Diversity of Sex Chromosome Systems in Ancistrini (Loricariidae, Hypostominae): ZZ/ZW in Ancistrus taunayi Miranda Ribeiro, 1918. Cytogenet. Genome Res. 2015, 146, 306–310. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, R.R.; Souza, I.L.; Venere, P.C. Karyotype Description of Three Species of Loricariidae (Siluriformes) and Occurrence of the ZZ/ZW Sexual System in Hemiancistrus spilomma Cardoso & Lucinda, 2003. Neotrop. Ichthyol. 2006, 4, 93–97. [Google Scholar] [CrossRef]
- Andreata, A.A.; do Almeida-Toledo, L.; Oliveira, C.; de Almeida Toledo Filho, S. Chromosome Studies in Hypoptopomatinae (Pisces, Siluriformes, Loricariidae). II. ZZ/ZW Sex-Chromosome System, B Chromosomes, and Constitutive Heterochromatin Differentiation in Microlepidogaster leucofrenatus. Cytogenet. Cell Genet. 1993, 63, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.C.; Ribeiro, M.O.; Dutra, E.S.; Zawadzki, C.H.; Portela-Castro, A.L.B.; Martins-Santos, I.C. Karyotype Structure of Hypostomus cf. plecostomus (Linnaeus, 1758) from Tapajós River Basin, Southern Amazon: Occurrence of Sex Chromosomes (ZZ/ZW) and Their Evolutionary Implications. Genet. Mol. Res. 2015, 14, 6625–6634. [Google Scholar] [CrossRef]
- de Souza, L.K.M.C.; Baumgärtner, L.; Paiva, S.; Zawadzki, C.H.; Martins-Santos, I.C.; de Brito Portela-Castro, A.L. Chromosomal Diversity of Three Species of Hypostomus Lacépède, 1803 (Siluriformes, Loricariidae), from the Paraná River Basin, Brazil: A Species Complex in Hypostomus ancistroides Reinforced by a ZZ/ZW Sex Chromosome System. Zebrafish 2017, 14, 357–363. [Google Scholar] [CrossRef]
- Yano, C.F.; Sember, A.; Kretschmer, R.; Bertollo, L.A.C. Against the mainstream: Exceptional evolutionary stability of ZW sex chromosomes across the fish families Triportheidae and Gasteropelecidae (Teleostei: Characiformes). Chromosome Res. 2021, 29, 391–416. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the Evolution of Heteromorphic Sex Chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef]
- Utsunomia, R.; de Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Goes, C.A.G.; Melo, S.; Ramos, L.P.; Oliveira, C.; Porto-Foresti, F.; Foresti, F.; Hashimoto, D.T. Satellitome Landscape Analysis of Megaleporinus macrocephalus (Teleostei, Anostomidae) Reveals Intense Accumulation of Satellite Sequences on the Heteromorphic Sex Chromosome. Sci. Rep. 2019, 9, 5856. [Google Scholar] [CrossRef] [PubMed]
- Bolzán, A.D. Interstitial Telomeric Sequences in Vertebrate Chromosomes: Origin, Function, Instability and Evolution. Mutat. Res. Rev. Mut. Res. 2017, 773, 51–65. [Google Scholar] [CrossRef]
- Ocalewicz, K.; Furgala-Selezniow, G.; Szmyt, M.; Lisboa, R.; Kucinski, M.; Lejk, A.M.; Jankun, M. Pericentromeric Location of the Telomeric DNA Sequences on the European Grayling Chromosomes. Genetica 2013, 141, 409–416. [Google Scholar] [CrossRef]
- Scacchetti, P.C.; Utsunomia, R.; Pansonato-Alves, J.C.; Vicari, M.R.; Artoni, R.F.; Oliveira, C.; Foresti, F. Chromosomal Mapping of Repetitive DNAs in Characidium (Teleostei, Characiformes): Genomic Organization and Diversification of ZW Sex Chromosomes. Cytogenet. Genome Res. 2015, 146, 136–143. [Google Scholar] [CrossRef]
- Nirchio, M.; Gaviria, J.I.; Siccha-Ramirez, Z.R.; Oliveira, C.; Foresti, F.; Milana, V.; Rossi, A.R. Chromosomal Polymorphism and Molecular Variability in the Pearly Razorfish Xyrichtys novacula (Labriformes, Labridae): Taxonomic and Biogeographic Implications. Genetica 2019, 147, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, A.; Nergadze, S.G.; Santagostino, M.; Giulotto, E. Telomeric Repeats far from the Ends: Mechanisms of Origin and Role in Evolution. Cytogenet. Genome Res. 2008, 122, 219–228. [Google Scholar] [CrossRef]
- Salvadori, S.; Deiana, A.; Elisabetta, C.; Floridia, G.; Rossi, E.; Zuffardi, O. Colocalization of (TTAGGG)n Telomeric Sequences and Ribosomal Genes in Atlantic Eels. Chromosome Res. 1995, 3, 54–58. [Google Scholar] [CrossRef]
- Gornung, E.; Mannarelli, M.E.; Rossi, A.R.; Sola, L. Chromosomal Evolution in Mugilidae (Pisces, Mugiliformes): FISH Mapping of the (TTAGGG) Telomeric Repeat in the Six Mediterranean Mullets. Hereditas 2004, 140, 158–159. [Google Scholar] [CrossRef]
- Rossi, A.R.; Gornung, E.; Sola, L.; Nirchio, M. Comparative Molecular Cytogenetic Analysis of Two Congeneric Species, Mugil curema and M. liza (Pisces, Mugiliformes), Characterized by Significant Karyotype Diversity. Genetica 2005, 125, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Libertini, A.; Vitturi, R.; Lannino, A.; Maone, M.C.; Franzoi, P.; Riccato, F.; Colomba, S. FISH Mapping of 18S rDNA and (TTAGGG)n Sequences in Two Pipefish Species (Gasteroisteiformes: Syngnathidae). J. Genet. 2006, 85, 153–156. [Google Scholar] [CrossRef]
- Slijepcevic, P. Telomeres and Mechanisms of Robertsonian Fusion. Chromosoma 1998, 107, 136–140. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nirchio Tursellino, M.; de Bello Cioffi, M.; de Menezes Cavalcante Sassi, F.; Deon, G.A.; Oliveira, C.; Kuranaka, M.; Valdiviezo-Rivera, J.; Gonzalez, V.H.; Rossi, A.R. Integrating Genomic and Chromosomal Data: A Cytogenetic Study of Transancistrus santarosensis (Loricariidae: Hypostominae) with Characterization of a ZZ/ZW Sex Chromosome System. Genes 2023, 14, 1662. https://doi.org/10.3390/genes14091662
Nirchio Tursellino M, de Bello Cioffi M, de Menezes Cavalcante Sassi F, Deon GA, Oliveira C, Kuranaka M, Valdiviezo-Rivera J, Gonzalez VH, Rossi AR. Integrating Genomic and Chromosomal Data: A Cytogenetic Study of Transancistrus santarosensis (Loricariidae: Hypostominae) with Characterization of a ZZ/ZW Sex Chromosome System. Genes. 2023; 14(9):1662. https://doi.org/10.3390/genes14091662
Chicago/Turabian StyleNirchio Tursellino, Mauro, Marcelo de Bello Cioffi, Francisco de Menezes Cavalcante Sassi, Geize Aparecida Deon, Claudio Oliveira, Mariana Kuranaka, Jonathan Valdiviezo-Rivera, Víctor Hugo Gonzalez, and Anna Rita Rossi. 2023. "Integrating Genomic and Chromosomal Data: A Cytogenetic Study of Transancistrus santarosensis (Loricariidae: Hypostominae) with Characterization of a ZZ/ZW Sex Chromosome System" Genes 14, no. 9: 1662. https://doi.org/10.3390/genes14091662