
Identification of ROH Islands Conserved through Generations in Pigs Belonging to the Nero Lucano Breed

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. DNA Analyses
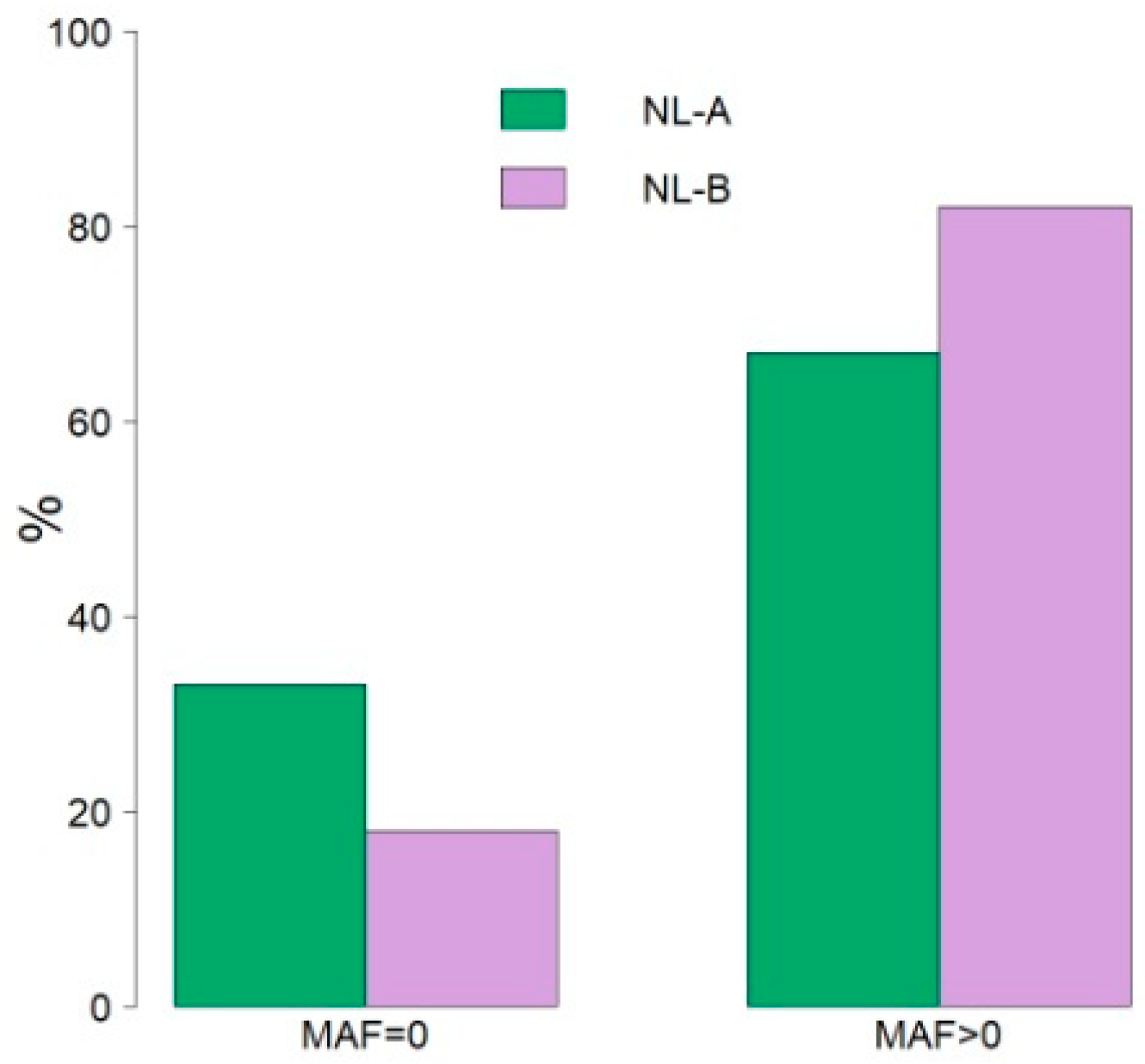
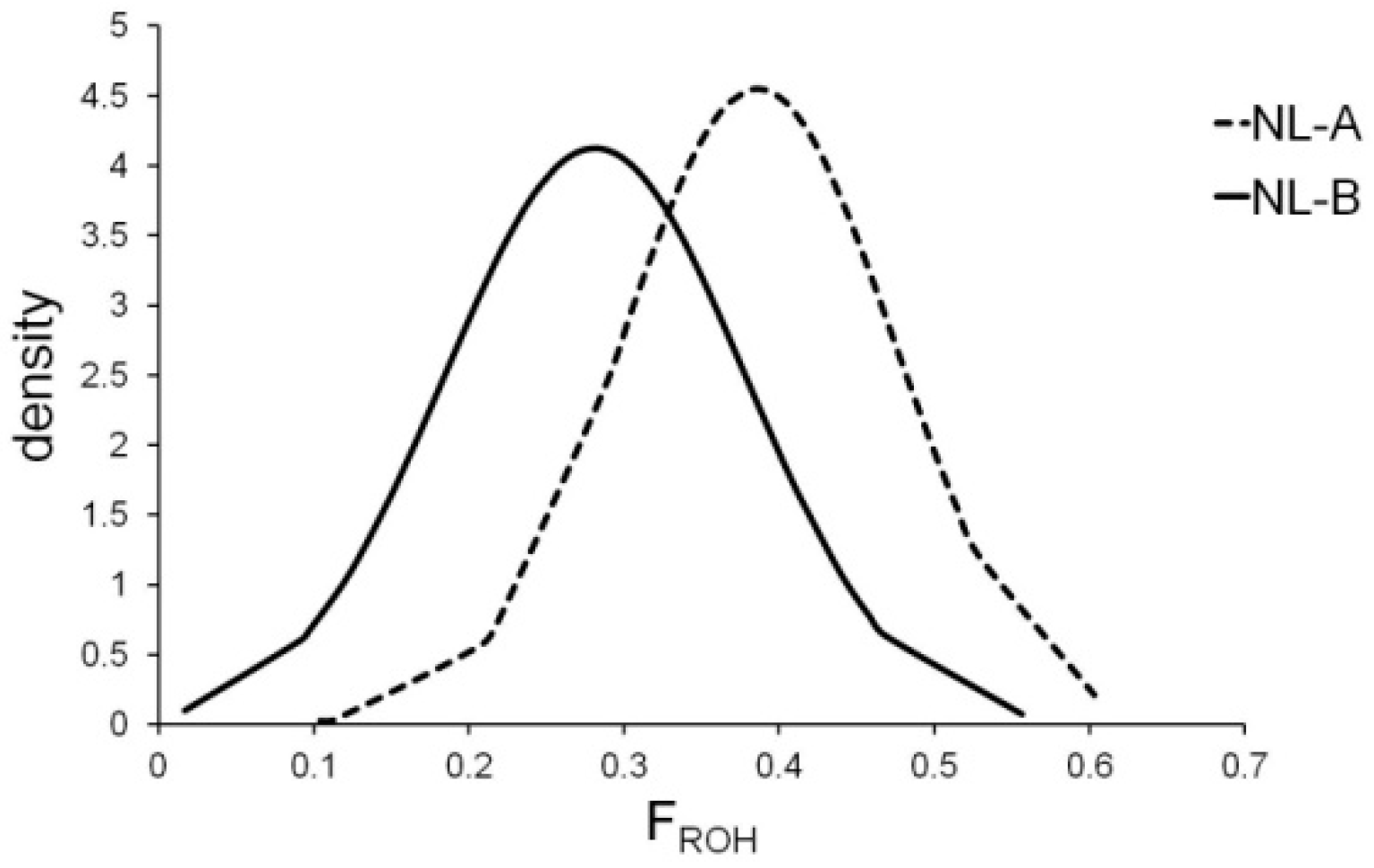
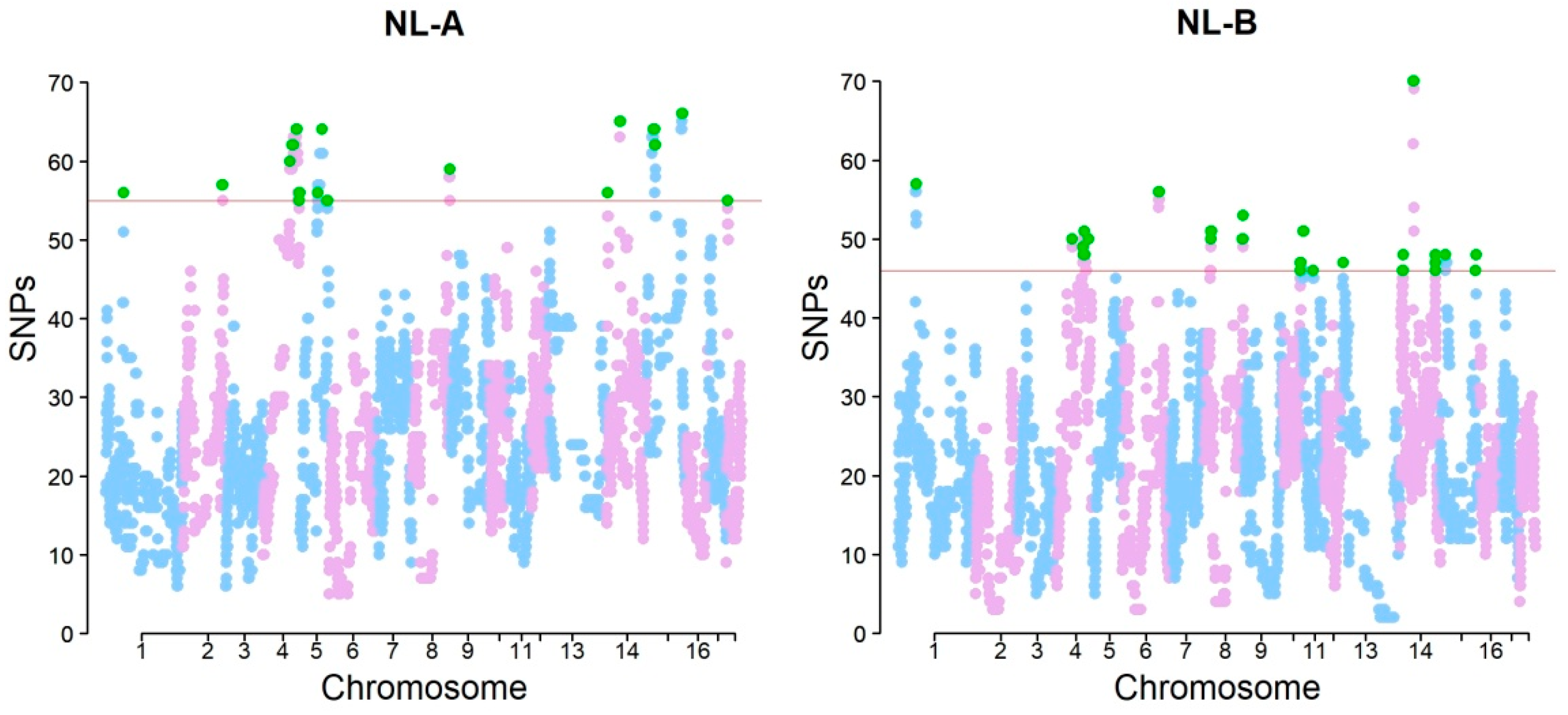
3. Results
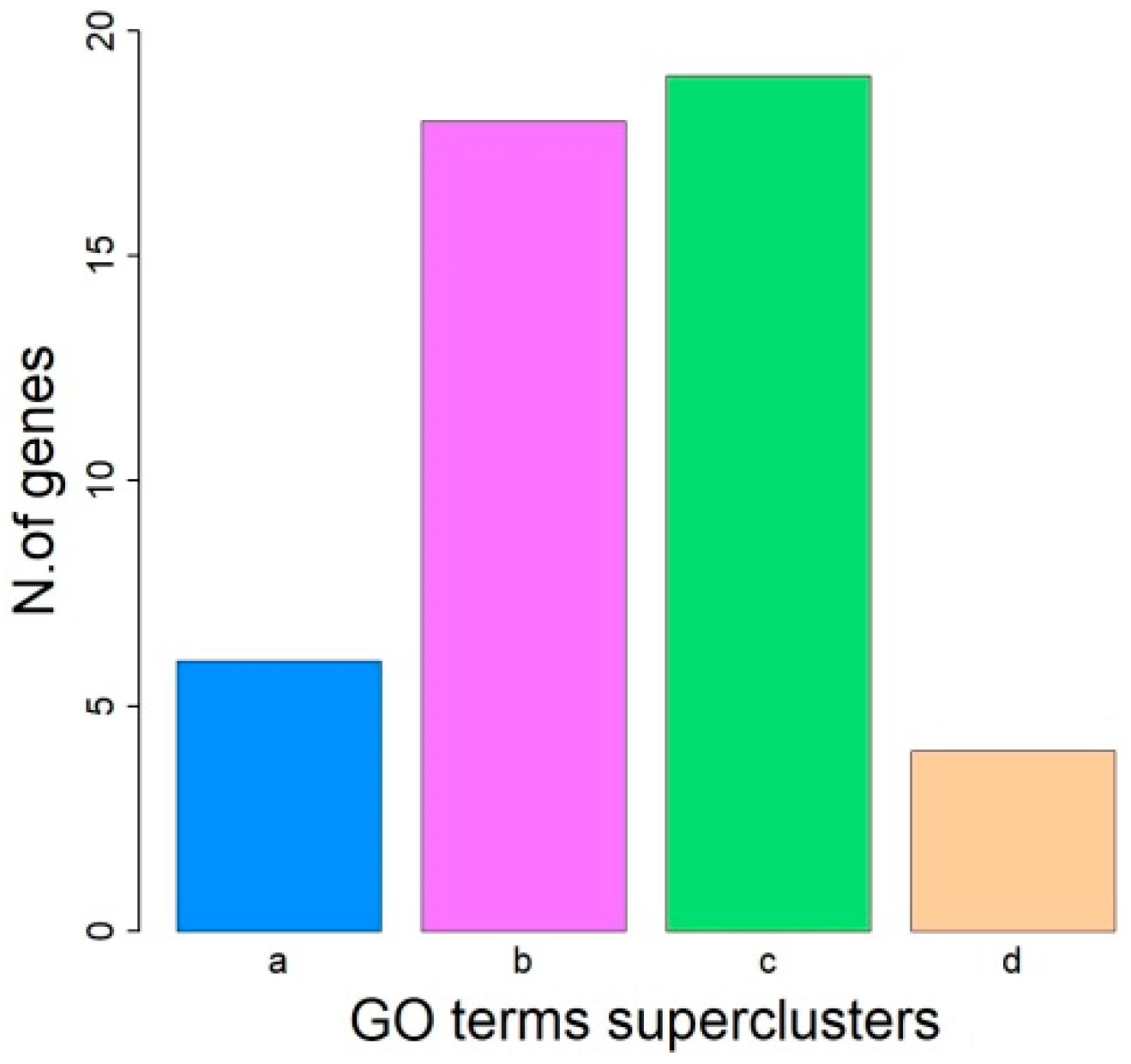
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hilborn, R.; Banobi, J.; Hall, S.J.; Pucylowski, T.; Walsworth, T.E. The environmental cost of animal source foods. Front. Ecol. Environ. 2018, 16, 329–335. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Farm to Fork Strategy: For a Fair, Healthy and Environmentally-Friendly Food System. COM/2020/381 Final. Date of Document: 20/05/2020. Available online: https://ec.europa.eu/commission/presscorner/detail/en/fs_20_908 (accessed on 15 February 2023).
- Farkas, J.; Curik, I.; Csato, L.; Csornyei, Z.; Baumung, R.; Nagy, I. Bayesian inference of inbreeding effects on litter size and gestation length in Hungarian Landrace and Hungarian Large White pigs. Livest. Sci. 2007, 112, 109–114. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhuo, Y.; Ning, C.; Zhou, L.; Liu, J.F. Estimate of inbreeding depression on growth and reproductive traits in a Large White pig population. G3 Genes Genomes Genet. 2022, 12, jkac118. [Google Scholar] [CrossRef]
- Valluzzi, C.; Rando, A.; Di Gregorio, P. Genetic variability of Nero Lucano pig breed at IGF2, LEP, MC4R, PIK3C3, RYR1 and VRTN loci. Ital. J. Anim. Sci. 2019, 18, 1321–1326. [Google Scholar] [CrossRef]
- Valluzzi, C.; Rando, A.; Macciotta, N.P.P.; Gaspa, G.; Di Gregorio, P. The Nero Lucano Pig Breed: Recovery and Variability. Animals 2021, 11, 1331. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A toolset for whole-genome association and population-based linkage analysis. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Supek, F.; Bosnjak, M.; Skunca, N.; Smuc, T. REVIGO summarizes and visualizes long lists of Gene Ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 15 February 2023).
- Szmatoła, T.; Gurgul, A.; Ropka-Molik, K.; Jasielczuk, I.; Ząbek, T.; Bugno-Poniewierska, M. Characteristics of runs of homozygosity in selected cattle breeds maintained in Poland. Livest. Sci. 2016, 188, 72–80. [Google Scholar] [CrossRef]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 family of cytokines in health and disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef] [PubMed]
- Rohrer, G.A.; Nonneman, D.J.; Wiedmann, R.T.; Schneider, J.F. A study of vertebra number in pigs confirms the association of vertnin and reveals additional QTL. BMC Genet. 2015, 16, 129. [Google Scholar] [CrossRef] [Green Version]
- Rohrer, G.A.; Nonneman, D.J. Genetic analysis of teat number in pigs reveals some developmental pathways independent of vertebra number and several loci which only affect a specific side. Genet. Sel. Evol. 2017, 49, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- See, G.M.; Trenhaile-Grannemann, M.D.; Spangler, M.L.; Ciobanu, D.C.; Mote, B.E. A genome-wide association study for gestation length in swine. Anim. Genet. 2019, 50, 539–542. [Google Scholar] [CrossRef]
- Wang, J.Y.; Luo, Y.R.; Fu, W.X.; Lu, X.; Zhou, J.P.; Ding, X.D.; Liu, J.F.; Zhang, Q. Genome-wide association studies for hematological traits in swine. Anim. Genet. 2013, 44, 34–43. [Google Scholar] [CrossRef]
- Nonneman, D.; Lents, C.; Rohrer, G.; Rempel, L.; Vallet, J. Genome-wide association with delayed puberty in swine. Anim. Genet. 2014, 45, 130–132. [Google Scholar] [CrossRef]
- Curik, I.; Ferencakovic, M.; Sölkner, J. Inbreeding and runs of homozygosity: A possible solution to an old problem. Livest. Sci. 2014, 166, 26–34. [Google Scholar] [CrossRef]
- Toro-Ospina, A.M.; Herrera Rios, A.C.; Pimenta Schettini, G.; Vallejo Aristizabal, V.H.; Bizarria dos Santos, W.; Zapata, C.A.; Ortiz Morea, E.G. Identification of Runs of Homozygosity Islands and Genomic Estimated Inbreeding Values in Caqueteño Creole Cattle (Colombia). Genes 2022, 13, 1232. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, P.; Michal, J.J.; Wang, Y.; Zhao, J.; Jiang, Z.; Liu, B. Molecular characterization of the porcine S100A6 gene and analysis of its expression in pigs infected with highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV). J. Appl. Genet. 2015, 56, 355–363. [Google Scholar] [CrossRef]
- Raymond, P.; Bellehumeur, C.; Nagarajan, M.; Longtin, D.; Ferland, A.; Muller, P.; Bissonnette, R.; Simard, C. Porcine reproductive and respiratory syndrome virus (PRRSV) in pig meat. Can. J. Vet. Res. 2017, 81, 162–170. [Google Scholar] [PubMed]
- Ueda, W.; Tohno, M.; Shimazu, T.; Fujie, H.; Aso, H.; Kawai, Y.; Numasaki, M.; Saito, T.; Kitazawa, H. Molecular cloning, tissue expression, and subcellular localization of porcine peptidoglycan recognition proteins 3 and 4. Vet. Immunol. Immunopathol. 2011, 143, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Han, C.; Xie, B.; Wu, Y.; Liu, S.; Chen, K.; Xia, M.; Zhang, Y.; Song, L.; Li, Z.; et al. Rhbdd3 controls autoimmunity by suppressing the production of IL-6 by dendritic cells via K27-linked ubiquitination of the regulator NEMO. Nat. Immunol. 2014, 15, 612–622. [Google Scholar] [CrossRef]
- Starkl, P.; Marichal, T.; Galli, S.J. PLA2G3 promotes mast cell maturation and function. Nat. Immunol. 2013, 14, 527–529. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Qi, N.; Xing, W.; Li, M.; Qian, Y.; Luo, G.; Yu, S. The TGF-β/SMAD signaling pathway prevents follicular atresia by upregulating MORC2. Int. J. Mol. Sci. 2022, 23, 10657. [Google Scholar] [CrossRef]
- Bonnet, A.; Le Cao, K.A.; Sancristobal, M.; Benne, F.; Robert-Granie, C.; Law-So, G.; Fabre, S.; Besse, P.; De Billy, E.; Quesnel, H.; et al. In vivo gene expression in granulosa cells during pig terminal follicular development. Reproduction 2008, 136, 211–224. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yang, M.; Zhou, M.; Wang, Y.; Wu, X.; Zhang, X.; Ding, Y.; Zhao, G.; Yin, Z.; Wang, C. Identification of signatures of selection by whole-genome resequencing of a Chinese native pig. Front. Genet. 2020, 11, 566255. [Google Scholar] [CrossRef]
- Shi, B.; Xue, J.; Zhou, J.; Kasowitz, S.D.; Zhang, Y.; Liang, G.; Guan, Y.; Shi, Q.; Liu, M.; Sha, J.; et al. MORC2B is essential for meiotic progression and fertility. PLoS Genet. 2018, 14, e1007175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udagawa, O.; Ito, C.; Ogonuki, N.; Sato, H.; Lee, S.; Tripvanuntakul, P.; Ichi, I.; Uchida, Y.; Nishimura, T.; Murakami, M.; et al. Oligo-astheno-teratozoospermia in mice lacking ORP4, a sterol-binding protein in the OSBP-related protein family. Genes Cells 2014, 19, 13–27. [Google Scholar] [CrossRef]
- Takahashi, H.; Koshimizu, U.; Miyazaki, J.; Nakamura, T. Impaired spermatogenic ability of testicular germ cells in mice deficient in the LIM-kinase 2 gene. Dev. Biol. 2002, 241, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Kuzmin, A.; Jarvi, K.; Lo, K.; Spencer, L.; Chow, G.Y.; Macleod, G.; Wang, Q.; Varmuza, S. Identification of potentially damaging amino acid substitutions leading to human male infertility. Biol. Reprod. 2009, 81, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonova, M.A.; Getmantseva, L.V.; Vasilenko, V.N.; Klimenko, A.I.; Usatov, A.V.; Yu, B.S.; Yu, K.A.; Shirockova, N.V. Leukemia Inhibitory Factor (LIF) Gene Polymorphism and its Impact on Reproductive Traits of Pigs. Am. J. Anim. Vet. Sci. 2015, 10, 212–216. [Google Scholar] [CrossRef]
- Pitts, M.W.; Reeves, M.A.; Hashimoto, A.C.; Ogawa, A.; Kremer, P.; Seale, L.A.; Berry, M.J. Deletion of selenoprotein M leads to obesity without cognitive deficits. J. Biol. Chem. 2013, 288, 26121–26134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Xu, C.; Meng, F.; Yao, Z.; Fan, Z.; Yang, Y.; Meng, X.; Zhan, Y.; Sun, Y.; Ma, F.; et al. Genome-Wide Association studies for flesh color and intramuscular fat in (Duroc Landrace Large White) crossbred commercial pigs. Genes 2022, 13, 2131. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kim, S.W.; Do, K.T.; Ha, Y.K.; Lee, Y.M.; Yoon, S.H.; Kim, H.B.; Kim, J.J.; Choi, B.H.; Kim, K.S. Analyses of porcine public SNPs in coding-gene regions by re-sequencing and phenotypic association studies. Mol. Biol. Rep. 2011, 38, 3805–3820. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wu, Y.; Qin, C.; He, W.; Wei, X. Phylogenetic and structural analysis of the phospholipase A2 gene family in vertebrates. Int. J. Mol. Med. 2015, 35, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Heid, I.M.; Jackson, A.U.; Randall, J.C.; Winkler, T.W.; Qi, L.; Steinthorsdottir, V.; Thorleifsson, G.; Zillikens, M.C.; Speliotes, E.K.; Mägi, R.; et al. Meta-analysis identifies 13 new loci associated with waist-hip ratio and reveals sexual dimorphism in the genetic basis of fat distribution. Nat. Genet. 2010, 42, 949–960, Erratum in Nat Genet. 2011, 43, 1164. [Google Scholar] [CrossRef] [Green Version]
- Bakoev, S.; Getmantseva, L.; Kostyunina, O.; Bakoev, N.; Prytkov, Y.; Usatov, A.; Tatarinova, T.V. Genome-wide analysis of genetic diversity and artificial selection in Large White pigs in Russia. PeerJ 2021, 9, e11595. [Google Scholar] [CrossRef]
- Li, X.; Yang, S.; Dong, K.; Tang, Z.; Li, K.; Fan, B.; Wang, Z.; Liu, B. Identification of positive selection signatures in pigs by comparing linkage disequilibrium variances. Anim. Genet. 2017, 48, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Uemoto, Y.; Nakano, H.; Kikuchi, T.; Sato, S.; Ishida, M.; Shibata, T.; Kadowaki, H.; Kobayashi, E.; Suzuki, K. Fine mapping of porcine SSC14 QTL and SCD gene effects on fatty acid composition and melting point of fat in a Duroc purebred population. Anim. Genet. 2012, 43, 225–228. [Google Scholar] [CrossRef]
- Munoz, M.; Rodríguez, M.C.; Alves, E.; Folch, J.M.; Ibanez-Escriche, N.; Silio, L.; Fernández, A.I. Genome-wide analysis of porcine backfat and intramuscular fat fatty acid composition using high-density genotyping and expression data. BMC Genom. 2013, 14, 845. [Google Scholar] [CrossRef] [Green Version]
- Cameron, N.D.; Enser, M.; Nute, G.R.; Whittington, F.M.; Penman, J.C.; Fisken, A.C.; Perry, A.M.; Wood, J.D. Genotype with nutrition interaction on fatty acid composition of intramuscular fat and the relationship with flavour of pig meat. Meat Sci. 2000, 55, 187–195. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ROH Class | NL-A | NL-B | ||||
|---|---|---|---|---|---|---|
| ROH Number | SNPs/ROH (Mean ± SD) | % Genome Coverage | ROH Number | SNPs/ROH (Mean ± SD) | % Genome Coverage | |
| <2 Mb | 260 | 57.30 ± 7.67 | 0.29 | 236 | 59.82 ± 8.71 | 0.25 |
| 2–4 Mb | 897 | 85.67 ± 22.93 | 1.73 | 1052 | 84.14 ± 20.18 | 1.85 |
| 4–8 Mb | 859 | 169.26 ± 46.37 | 3.24 | 913 | 161.81 ± 41.76 | 3.16 |
| 8–16 Mb | 707 | 329.64 ± 76.12 | 5.40 | 741 | 321.53 ± 72.55 | 5.07 |
| >16 Mb | 903 | 1180.92 ± 783.82 | 27.97 | 748 | 1002.98 ± 636.18 | 17.78 |
| Total | 3626 | - | 38.63 | 3690 | - | 28.11 |
| SSC | NL-A | NL-B | N. Genes | ||
|---|---|---|---|---|---|
| from-to | N. SNPs | from-to | N. SNPs | ||
| 1 | 61,286,411-63,123,772 | 52 | 61,286,411-63,012,075 | 47 | 1 |
| 4 | 96,094,167-96,682,751 | 19 | 96,094,167-96,243,742 | 8 | 7 |
| 8 | 135,852,700-136,018,155 | 4 | 135,852,700-136,018,155 | 4 | 2 |
| 14 | 46,176,964-48,062,822 | 60 | 46,176,964-47,999,414 | 58 | 39 |
| 15 | 126,097,384-126,642,052 | 17 | 126,097,384-126,642,052 | 17 | 3 |
| GO | Biological Process | Genes |
|---|---|---|
| GO:0006954 | inflammatory response | S100A12, S100A8, S100A9, OSM, PLA2G3, RHBDD3, |
| GO:0048731 | system development | LIF, LIMK2, NF2, S100A8, S100A9, THOC5, AP1B1, CUL3, GAL3ST1, GAS2L1, INPP5J, KREMEN1, NEFH, PLA2G3, RHBDD3, SELENOM, ZNRF3 |
| GO:0051128 | regulation of cellular component organization | LIF, LIMK2, MORC2, NF2, S100A9, CUL3, GAS2L1, HNRNPD, INPP5J, KREMEN1, MTMR3, PLA2G3, |
| GO:0033043 | regulation of organelle organization | LIF, LIMK2, MORC2, NF2, CUL3, GAS2L1, HNRNPD, MTMR3, |
| GO:0009914 | hormone transport | LIF, OSM, PLA2G3, SELENOM, |
| GO:0051046 | regulation of secretion | LIF, S100A8, OSM, PLA2G3, RHBDD3, |
| GO:0046903 | secretion | LIF, S100A8, OSM, PLA2G3, RHBDD3, SELENOM, |
| GO:0050729 | positive regulation of inflammatory response | S100A8, OSM, PLA2G3 |
| GO:0031345 | negative regulation of cell projection organization | LIMK2, INPP5J, KREMEN1, |
| GO:0023061 | signal release | LIF, OSM, PLA2G3, SELENOM, |
| GO:0051247 | positive regulation of protein metabolic process | LIF, LIMK2, S100A8, TBC1D10A, CUL3, HNRNPD, OSM, RHBDD3, |
| GO:0010648 | negative regulation of cell communication | LIF, NF2, CUL3, CASTOR1, KREMEN1, OSM, ZNRF3, |
| GO:0023057 | negative regulation of signaling | LIF, NF2, CUL3, CASTOR1, KREMEN1, OSM, ZNRF3, |
| GO:0048232 | male gamete generation | LIMK2, GAL3ST1, OSBP2, PLA2G3, |
| GO:0046486 | glycerolipid metabolic process | GAL3ST1, INPP5J, MTMR3, PLA2G3, |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Gregorio, P.; Perna, A.; Di Trana, A.; Rando, A. Identification of ROH Islands Conserved through Generations in Pigs Belonging to the Nero Lucano Breed. Genes 2023, 14, 1503. https://doi.org/10.3390/genes14071503
Di Gregorio P, Perna A, Di Trana A, Rando A. Identification of ROH Islands Conserved through Generations in Pigs Belonging to the Nero Lucano Breed. Genes. 2023; 14(7):1503. https://doi.org/10.3390/genes14071503
Chicago/Turabian StyleDi Gregorio, Paola, Annamaria Perna, Adriana Di Trana, and Andrea Rando. 2023. "Identification of ROH Islands Conserved through Generations in Pigs Belonging to the Nero Lucano Breed" Genes 14, no. 7: 1503. https://doi.org/10.3390/genes14071503