
Altered Transcript Levels of MMP13 and VIT Genes in the Muscle and Connective Tissue of Pigs with Umbilical Hernia
 , , and
, , and 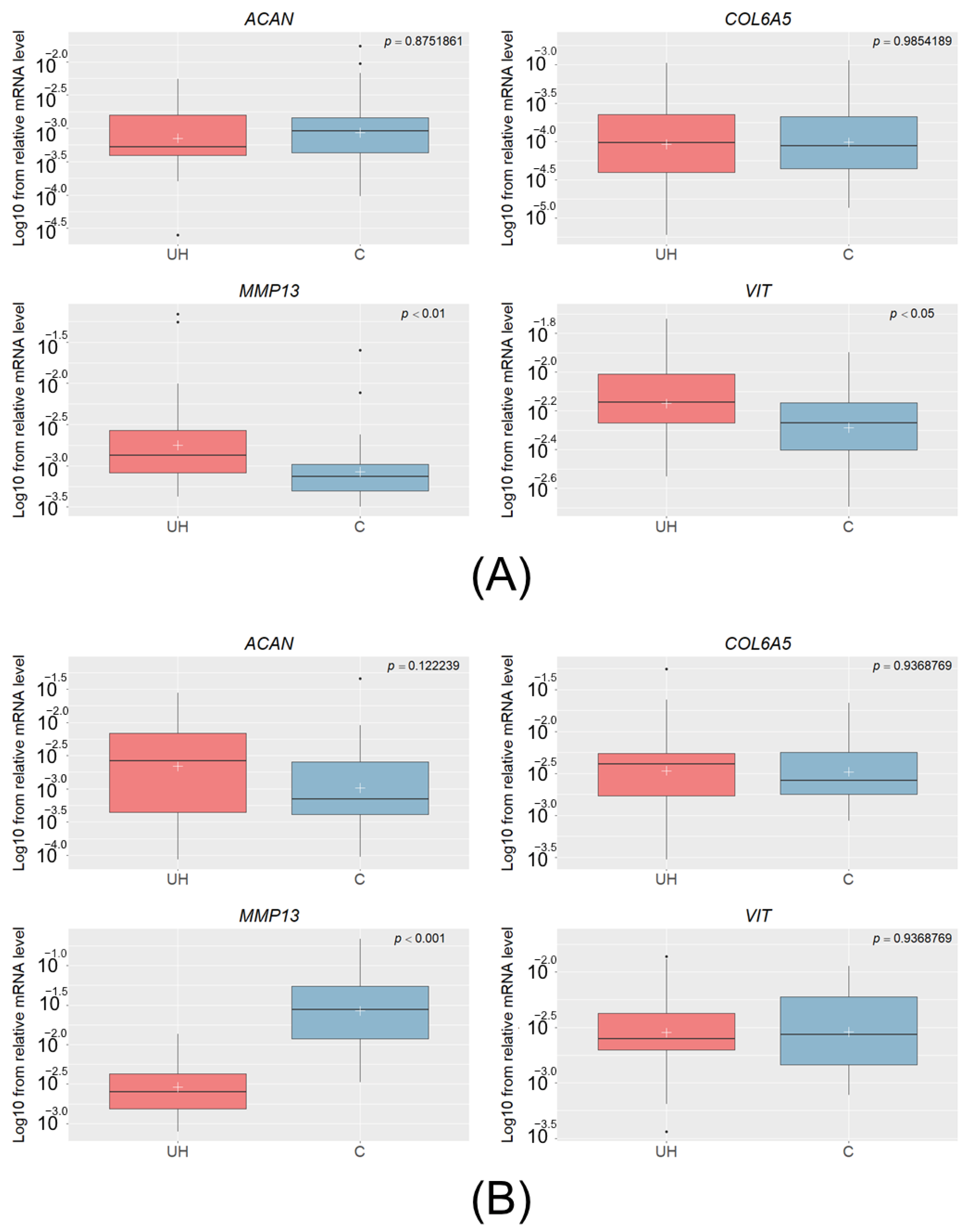{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Groups, Phenotypes, and Sample Collection
2.2.1. Pigs with Umbilical Hernia
2.2.2. Pigs with Inguinal Hernia
2.3. RNA Extraction and cDNA Synthesis for qPCR Assay
2.4. Quantitative Real-Time PCR
2.5. Methylation Analysis
2.6. Statistical Analysis
3. Results
3.1. Relative mRNA Levels of the Studied Genes in the UH Pigs
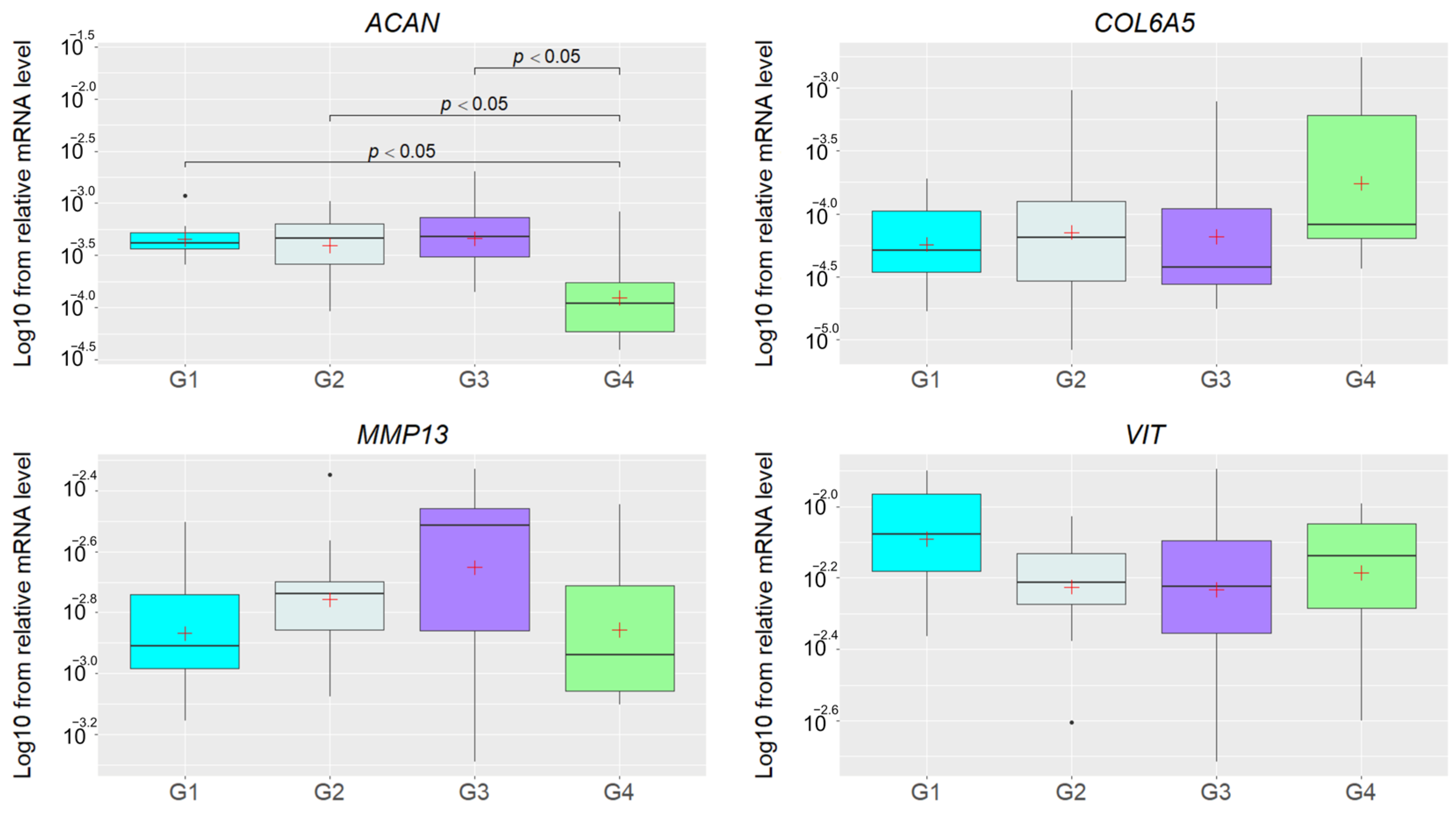
3.2. Relative mRNA Levels of Studied Genes in IH Pigs
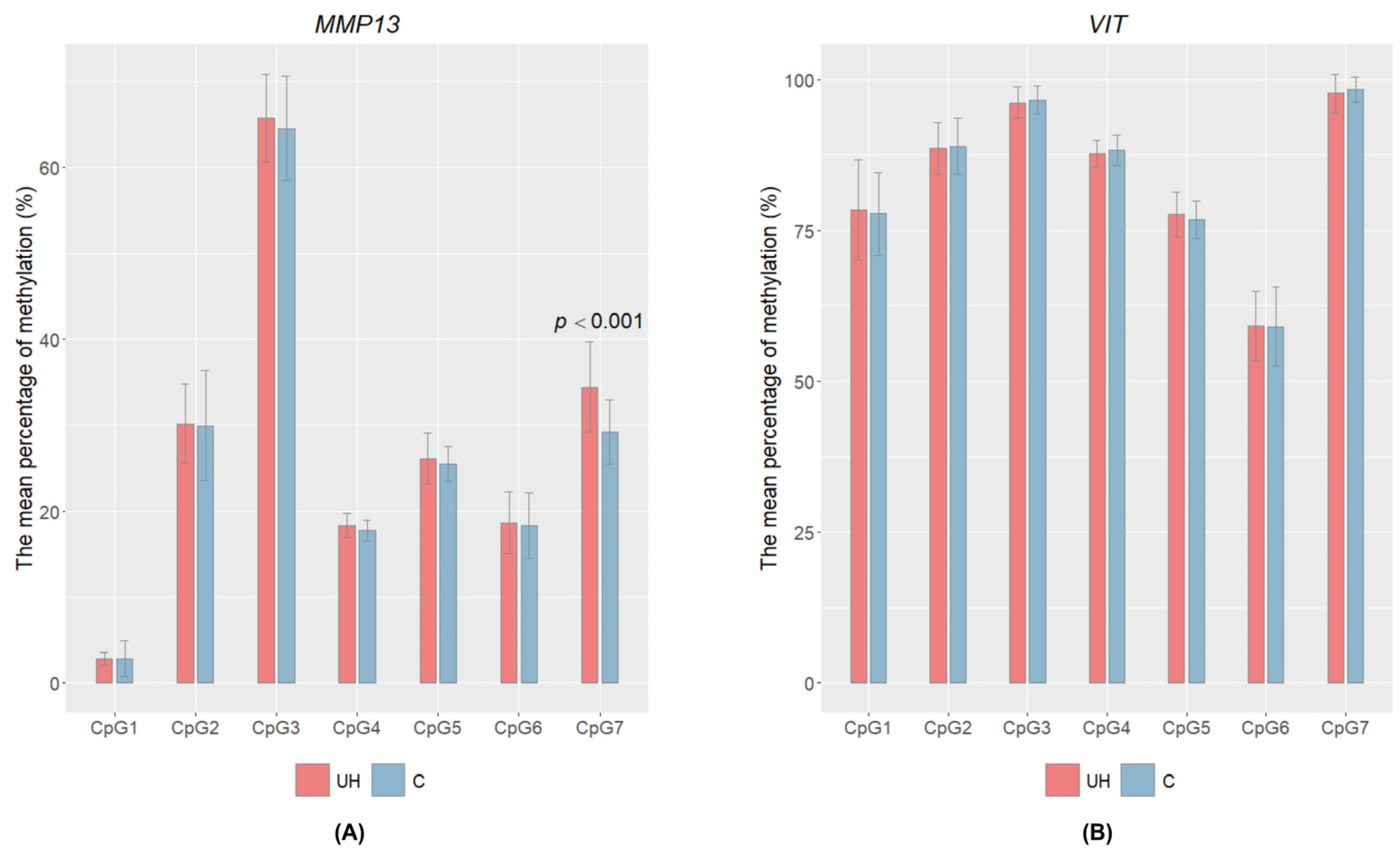
3.3. CpG Methylation Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rauw, W.M.; Rydhmer, L.; Kyriazakis, I.; Øverland, M.; Gilbert, H.; Dekkers, J.C.; Hermesch, S.; Bouquet, A.; Gómez Izquierdo, E.; Louveau, I.; et al. Prospects for Sustainability of Pig Production in Relation to Climate Change and Novel Feed Resources. J. Sci. Food Agric. 2020, 100, 3575–3586. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.J.; Vellinga, T.V.; Steinfeld, H. Issues and Options in Addressing the Environmental Consequences of Livestock Sector’s Growth. Meat Sci. 2010, 84, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Davoli, R.; Braglia, S. Molecular Approaches in Pig Breeding to Improve Meat Quality. Brief. Funct. Genom. Proteom. 2008, 6, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Thorslund, C.A.H.; Aaslyng, M.D.; Lassen, J. Perceived Importance and Responsibility for Market-Driven Pig Welfare: Literature Review. Meat Sci. 2017, 125, 37–45. [Google Scholar] [CrossRef]
- Lago, L.; Nery Da Silva, A.; Zanella, E.; Groke Marques, M.; Peixoto, J.; Da Silva, M.; Ledur, M.; Zanella, R. Identification of Genetic Regions Associated with Scrotal Hernias in a Commercial Swine Herd. Vet. Sci. 2018, 5, 15. [Google Scholar] [CrossRef]
- Souza, M.R.; Ibelli, A.M.G.; Savoldi, I.R.; Cantão, M.E.; Peixoto, J.D.O.; Mores, M.A.Z.; Lopes, J.S.; Coutinho, L.L.; Ledur, M.C. Transcriptome Analysis Identifies Genes Involved with the Development of Umbilical Hernias in Pigs. PLoS ONE 2020, 15, e0232542. [Google Scholar] [CrossRef]
- Rodrigues, A.F.G.; Ibelli, A.M.G.; Peixoto, J.D.O.; Cantão, M.E.; Oliveira, H.C.D.; Savoldi, I.R.; Souza, M.R.; Mores, M.A.Z.; Carreño, L.O.D.; Ledur, M.C. Genes and SNPs Involved with Scrotal and Umbilical Hernia in Pigs. Genes 2021, 12, 166. [Google Scholar] [CrossRef]
- Grindflek, E.; Hansen, M.H.S.; Lien, S.; Van Son, M. Genome-Wide Association Study Reveals a QTL and Strong Candidate Genes for Umbilical Hernia in Pigs on SSC14. BMC Genom. 2018, 19, 412. [Google Scholar] [CrossRef]
- Hammoud, M.; Gerken, J. Inguinal Hernia. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Liao, Y.; Smyth, G.K.; Shi, W. The R Package Rsubread Is Easier, Faster, Cheaper and Better for Alignment and Quantification of RNA Sequencing Reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef]
- Mikami, H.; Fredeen, H.T. A GENETIC STUDY OF CRYPTORCHIDISM AND SCROTAL HERNIA IN PIGS. Can. J. Genet. Cytol. 1979, 21, 9–19. [Google Scholar] [CrossRef]
- Searcy-Bernal, R.; Gardner, I.A.; Hird, D.W. Effects of and Factors Associated with Umbilical Hernias in a Swine Herd. J. Am. Vet. Med. Assoc. 1994, 204, 1660–1664. [Google Scholar] [PubMed]
- Thaller, G.; Dempfle, L.; Hoeschele, I. Investigation of the Inheritance of Birth Defects in Swine by Complex Segregation Analysis. J. Anim. Breed. Genet. 1996, 113, 77–92. [Google Scholar] [CrossRef]
- Sevillano, C.A.; Lopes, M.S.; Harlizius, B.; Hanenberg, E.; Knol, E.F.; Bastiaansen, J. Genome-Wide Association Study Using Deregressed Breeding Values for Cryptorchidism and Scrotal/Inguinal Hernia in Two Pig Lines. Genet. Sel. Evol. 2015, 47, 18. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.S.; Mao, H.R.; Guo, Y.M.; Ren, J.; Xiao, S.J.; Wu, G.Z.; Shen, H.Q.; Wu, L.H.; Ruan, G.F.; Brenig, B.; et al. A Genome-Wide Scan Reveals Candidate Susceptibility Loci for Pig Hernias in an Intercross between White Duroc and Erhualian1. J. Anim. Sci. 2009, 87, 2469–2474. [Google Scholar] [CrossRef]
- Nowacka-Woszuk, J. The Genetic Background of Hernia in Pigs: A Review. Livest. Sci. 2021, 244, 104317. [Google Scholar] [CrossRef]
- Krupova, Z. Candidate Genes for Congenital Malformations in Pigs. Acta Fytotechn Zootech. 2021, 24, 309–314. [Google Scholar] [CrossRef]
- Long, Y.; Su, Y.; Ai, H.; Zhang, Z.; Yang, B.; Ruan, G.; Xiao, S.; Liao, X.; Ren, J.; Huang, L.; et al. A Genome-Wide Association Study of Copy Number Variations with Umbilical Hernia in Swine. Anim. Genet. 2016, 47, 298–305. [Google Scholar] [CrossRef]
- Li, X.; Xu, P.; Zhang, C.; Sun, C.; Li, X.; Han, X.; Li, M.; Qiao, R. Genome-wide Association Study Identifies Variants in the CAPN 9 Gene Associated with Umbilical Hernia in Pigs. Anim. Genet. 2019, 50, 162–165. [Google Scholar] [CrossRef]
- Romano, G.D.S.; Ibelli, A.M.G.; Lorenzetti, W.R.; Weber, T.; Peixoto, J.D.O.; Cantão, M.E.; Mores, M.A.Z.; Morés, N.; Pedrosa, V.B.; Coutinho, L.L.; et al. Inguinal Ring RNA Sequencing Reveals Downregulation of Muscular Genes Related to Scrotal Hernia in Pigs. Genes 2020, 11, 117. [Google Scholar] [CrossRef]
- Greenberg, M.V.C.; Bourc’his, D. The Diverse Roles of DNA Methylation in Mammalian Development and Disease. Nat. Rev. Mol. Cell Biol. 2019, 20, 590–607. [Google Scholar] [CrossRef]
- Lorenzetti, W.R.; Ibelli, A.M.G.; Peixoto, J.D.O.; Mores, M.A.Z.; Savoldi, I.R.; Carmo, K.B.D.; Oliveira, H.C.D.; Ledur, M.C. Identification of Endogenous Normalizing Genes for Expression Studies in Inguinal Ring Tissue for Scrotal Hernias in Pigs. PLoS ONE 2018, 13, e0204348. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-Project.Org (accessed on 10 February 2022).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Wozniak, J.; Loba, W.; Iskrzak, P.; Pszczola, M.; Wojtczak, J.; Switonski, M.; Nowacka-Woszuk, J. A Confirmed Association between DNA Variants in CAPN9, OSM, and ITGAM Candidate Genes and the Risk of Umbilical Hernia in Pigs. Anim. Genet. 2023, 54, 307–314. [Google Scholar] [CrossRef]
- Zheng, Z.S.; Reiner Kasperk, H. Recurrent Inguinal Hernia: Disease of the Collagen Matrix? World J. Surg. 2002, 26, 401–408. [Google Scholar] [CrossRef]
- Bracale, U.; Peltrini, R.; Iacone, B.; Martirani, M.; Sannino, D.; Gargiulo, A.; Corcione, F.; Serra, R.; Bracale, U.M. A Systematic Review on the Role of Matrix Metalloproteinases in the Pathogenesis of Inguinal Hernias. Biomolecules 2023, 13, 1123. [Google Scholar] [CrossRef]
- Amar, S.; Smith, L.; Fields, G.B. Matrix Metalloproteinase Collagenolysis in Health and Disease. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2017, 1864, 1940–1951. [Google Scholar] [CrossRef]
- Vincenti, M.P.; Brinckerhoff, C.E. Transcriptional Regulation of Collagenase (MMP-1, MMP-13) Genes in Arthritis: Integration of Complex Signaling Pathways for the Recruitment of Gene-Specific Transcription Factors. Arthritis Res. Ther. 2002, 4, 157. [Google Scholar] [CrossRef]
- Toriseva, M.; Laato, M.; Carpén, O.; Ruohonen, S.T.; Savontaus, E.; Inada, M.; Krane, S.M.; Kähäri, V.-M. MMP-13 Regulates Growth of Wound Granulation Tissue and Modulates Gene Expression Signatures Involved in Inflammation, Proteolysis, and Cell Viability. PLoS ONE 2012, 7, e42596. [Google Scholar] [CrossRef]
- Lei, H.; Leong, D.; Smith, L.R.; Barton, E.R. Matrix Metalloproteinase 13 Is a New Contributor to Skeletal Muscle Regeneration and Critical for Myoblast Migration. Am. J. Physiol. Cell Physiol. 2013, 305, C529–C538. [Google Scholar] [CrossRef]
- Barrett, L.W.; Fletcher, S.; Wilton, S.D. Regulation of Eukaryotic Gene Expression by the Untranslated Gene Regions and Other Non-Coding Elements. Cell Mol. Life Sci. 2012, 69, 3613–3634. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Bai, C.; Yang, L.; Zheng, Z.; Su, G.; Gao, G.; Wei, Z.; Zuo, Y.; Li, G. DNA Methylation Subpatterns at Distinct Regulatory Regions in Human Early Embryos. Open Biol. 2018, 8, 180131. [Google Scholar] [CrossRef]
- Colombatti, A.; Bonaldo, P.; Doliana, R. Type A Modules: Interacting Domains Found in Several Non-Fibrillar Collagens and in Other Extracellular Matrix Proteins. Matrix 1993, 13, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, C.A.; Hynes, R.O. Distribution and Evolution of von Willebrand/Integrin A Domains: Widely Dispersed Domains with Roles in Cell Adhesion and Elsewhere. MBoC 2002, 13, 3369–3387. [Google Scholar] [CrossRef]
- Roughley, P.J.; Mort, J.S. The Role of Aggrecan in Normal and Osteoarthritic Cartilage. J. Exp. Ortop. 2014, 1, 8. [Google Scholar] [CrossRef] [PubMed]
- Aspberg, A. The Different Roles of Aggrecan Interaction Domains. J. Histochem. Cytochem. 2012, 60, 987–996. [Google Scholar] [CrossRef]
- Miranda-Duarte, A. DNA Methylation in Osteoarthritis: Current Status and Therapeutic Implications. TORJ 2018, 12, 37–49. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wozniak, J.; Loba, W.; Wysocka, A.; Dzimira, S.; Przadka, P.; Switonski, M.; Nowacka-Woszuk, J. Altered Transcript Levels of MMP13 and VIT Genes in the Muscle and Connective Tissue of Pigs with Umbilical Hernia. Genes 2023, 14, 1903. https://doi.org/10.3390/genes14101903
Wozniak J, Loba W, Wysocka A, Dzimira S, Przadka P, Switonski M, Nowacka-Woszuk J. Altered Transcript Levels of MMP13 and VIT Genes in the Muscle and Connective Tissue of Pigs with Umbilical Hernia. Genes. 2023; 14(10):1903. https://doi.org/10.3390/genes14101903
Chicago/Turabian StyleWozniak, Jakub, Weronika Loba, Alicja Wysocka, Stanislaw Dzimira, Przemyslaw Przadka, Marek Switonski, and Joanna Nowacka-Woszuk. 2023. "Altered Transcript Levels of MMP13 and VIT Genes in the Muscle and Connective Tissue of Pigs with Umbilical Hernia" Genes 14, no. 10: 1903. https://doi.org/10.3390/genes14101903