
Transcriptomic Characterization of Genes Regulating the Stemness in Porcine Atrial Cardiomyocytes during Primary In Vitro Culture
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cell Isolation
2.3. Long-Term Primary Cell Culture
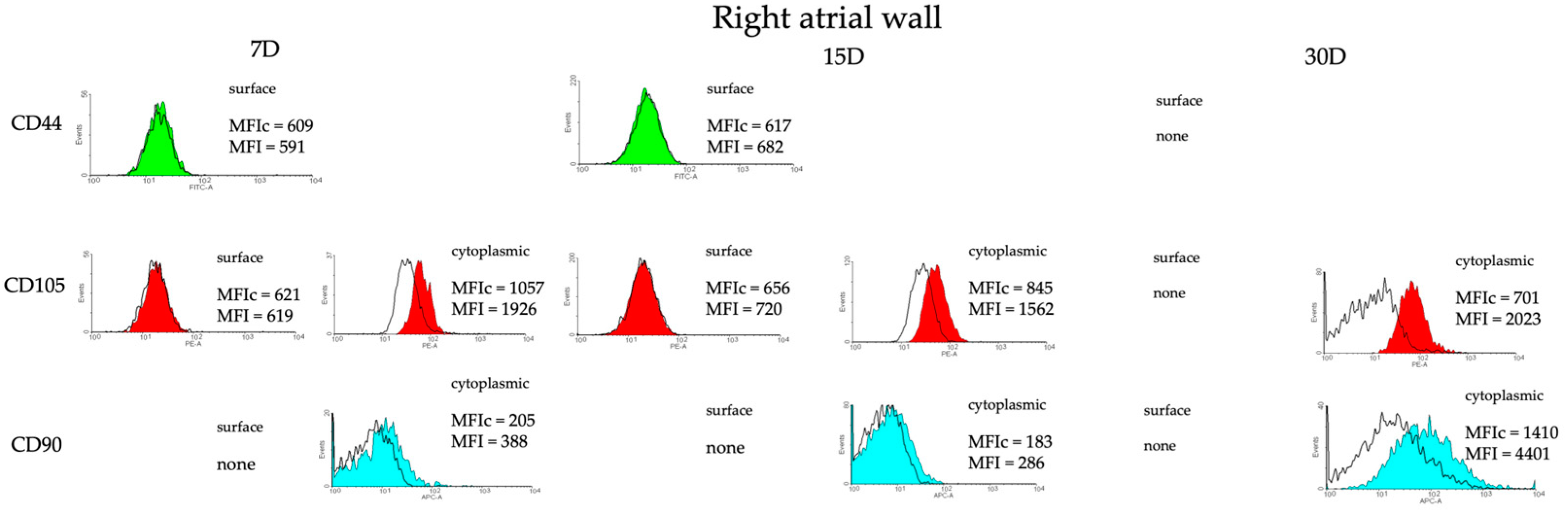
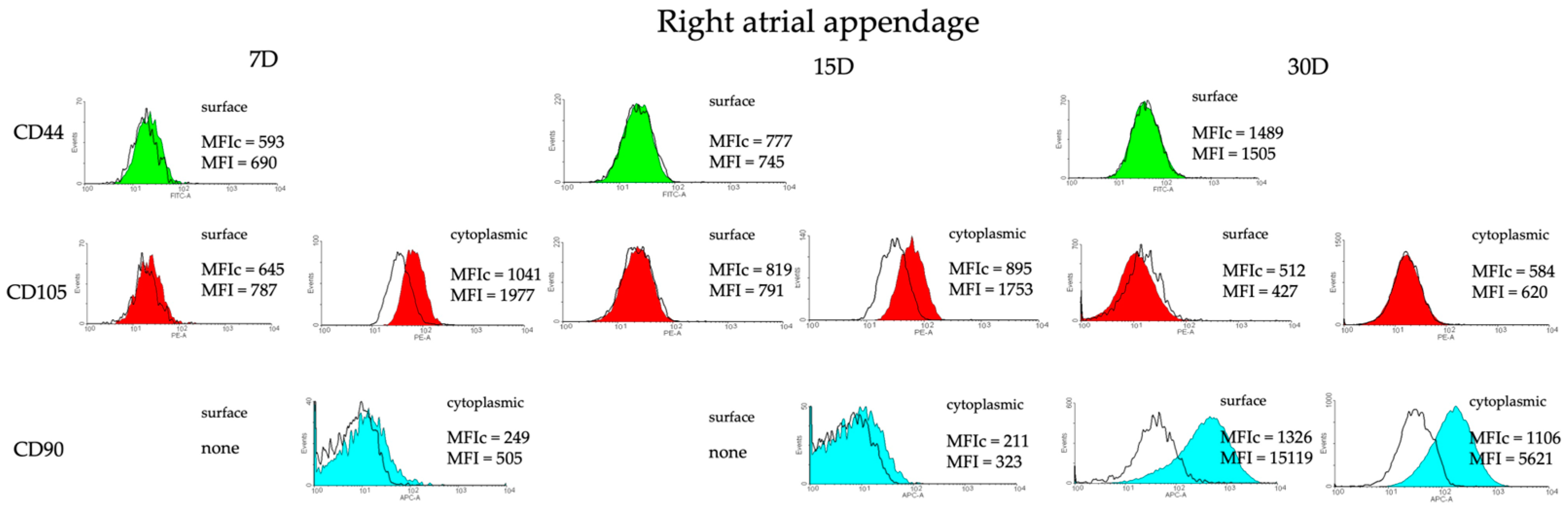
2.4. Detection of Chosen Markers Using Flow Cytometry (FC)
2.5. RNA Extraction and Isolation
2.6. Microarray-Based Transcriptomic Profiling
2.7. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.8. Bioinformatic Data Analysis and Visualization
3. Results
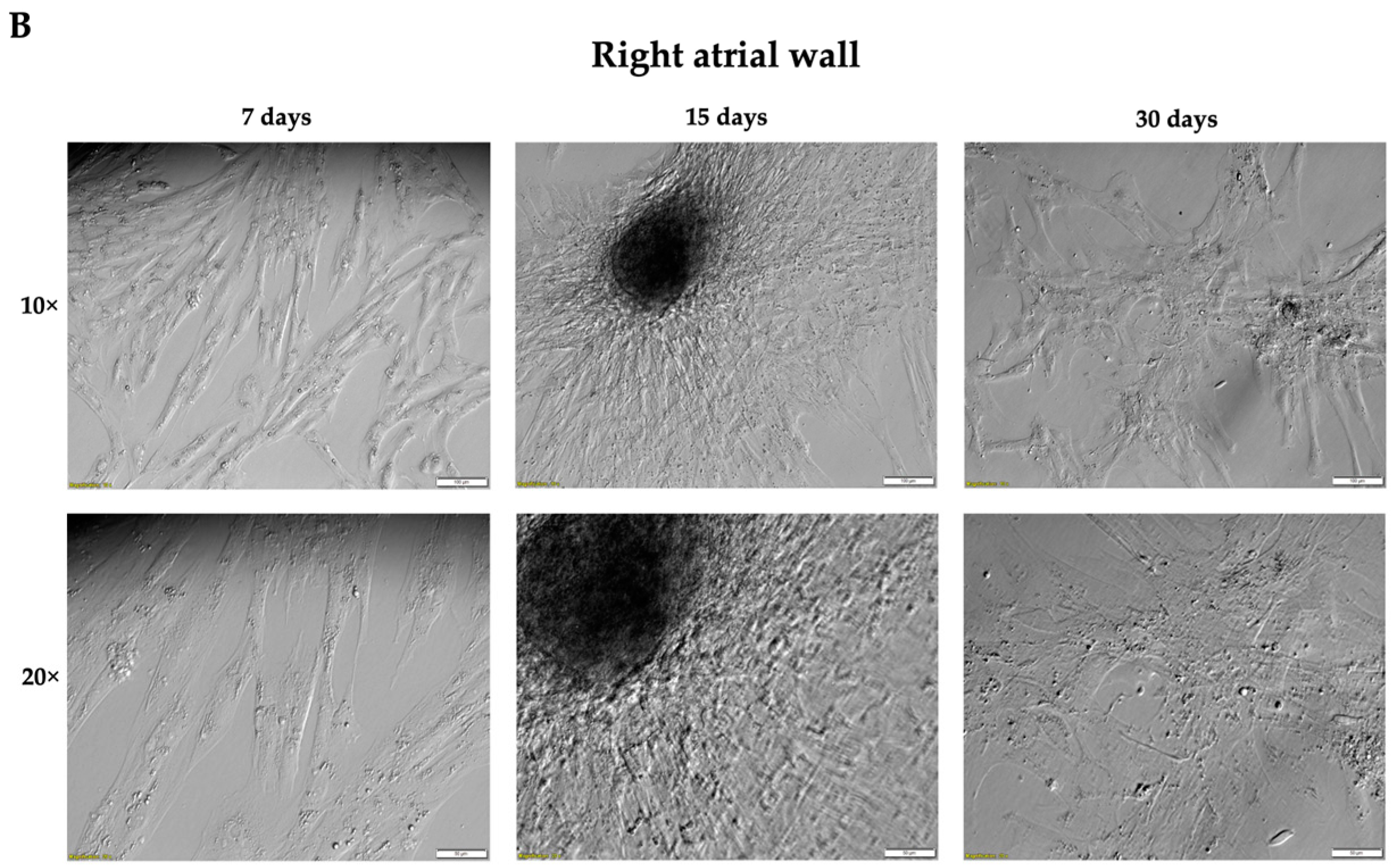
3.1. Isolation of Cells from Two Fragments of Porcine Cardiac Muscle and Long-Term In Vitro Primary Culture
3.2. Identification of Possible Stemness Phenotype of the Cells Isolated from the Heart Tissue
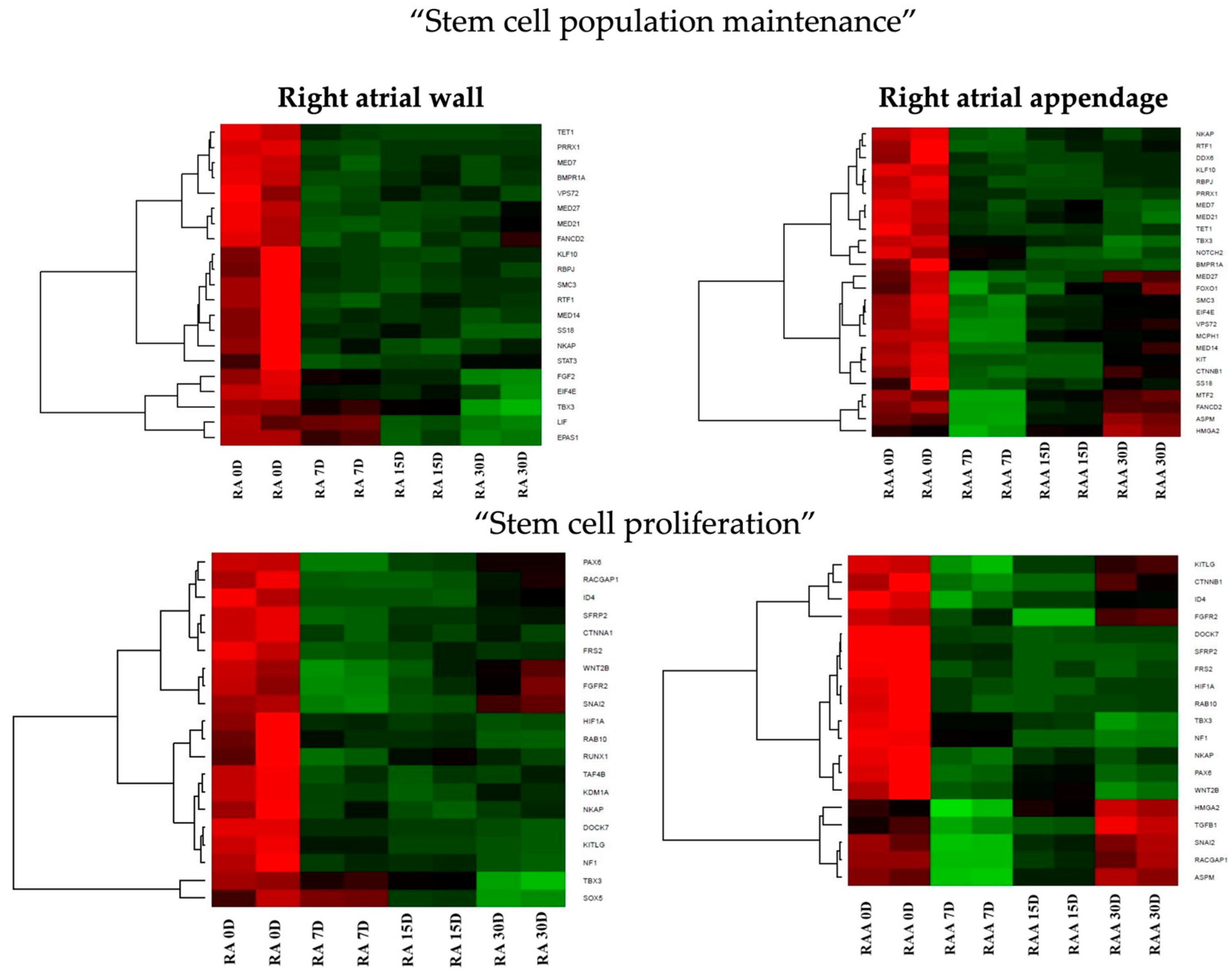
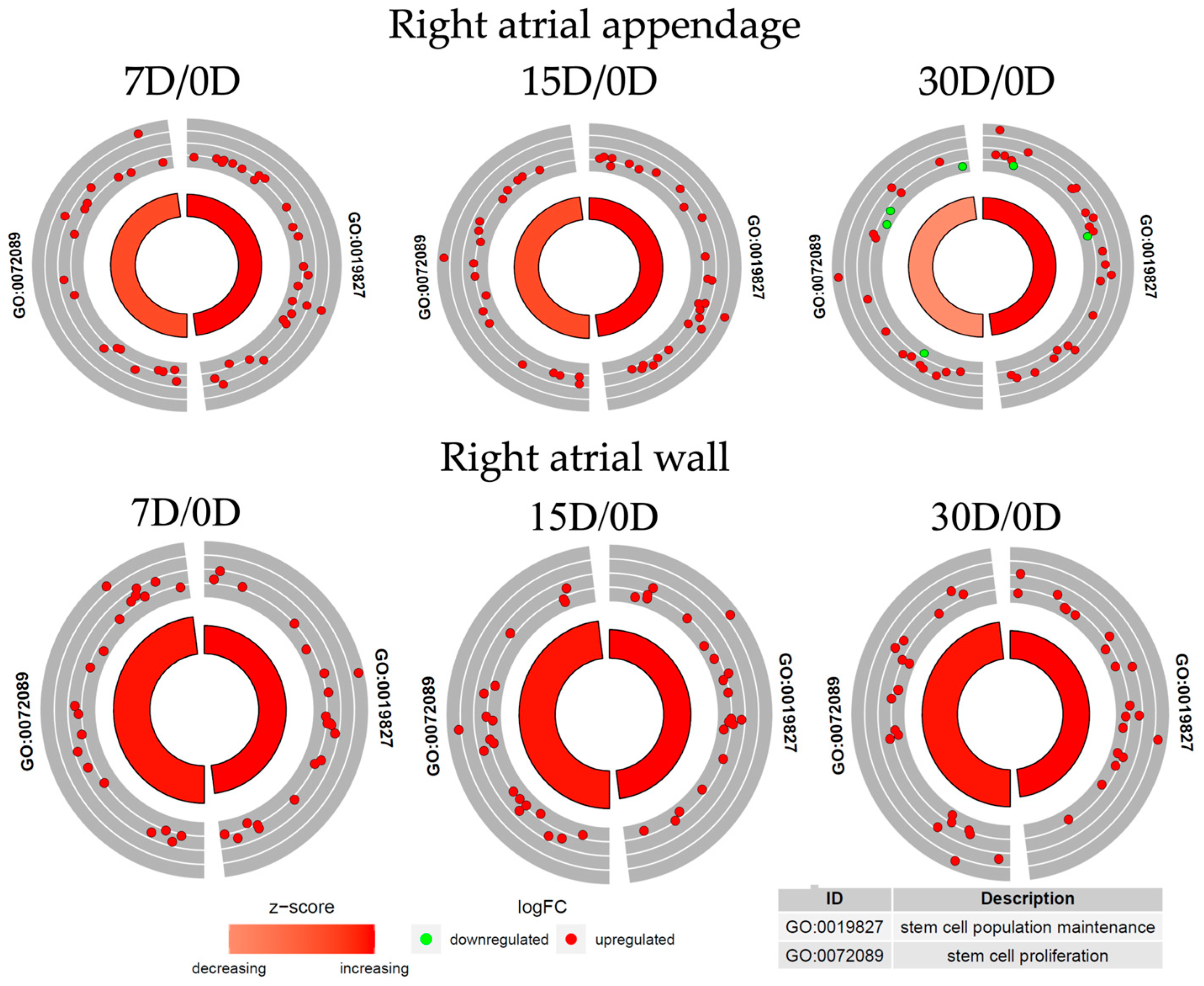
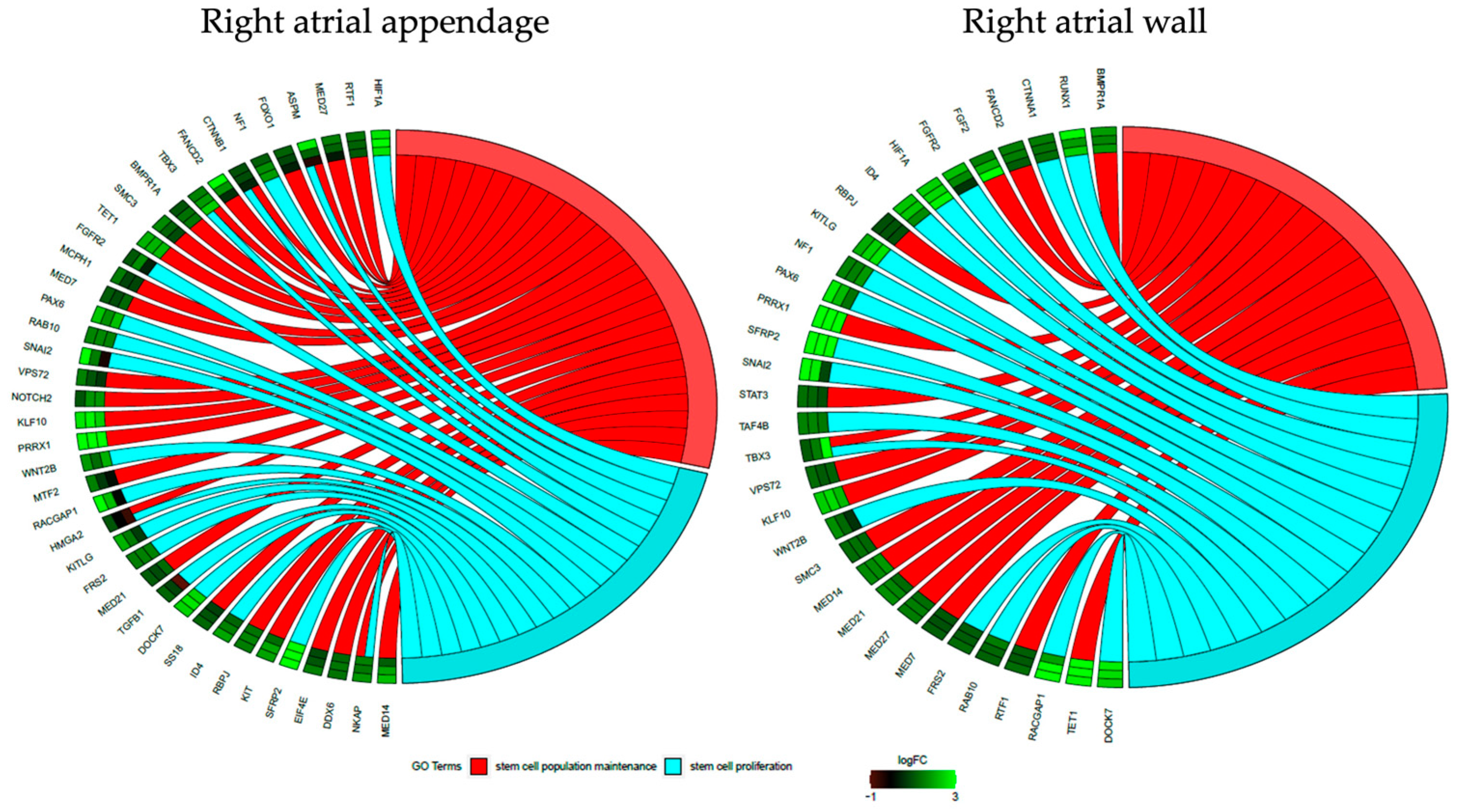
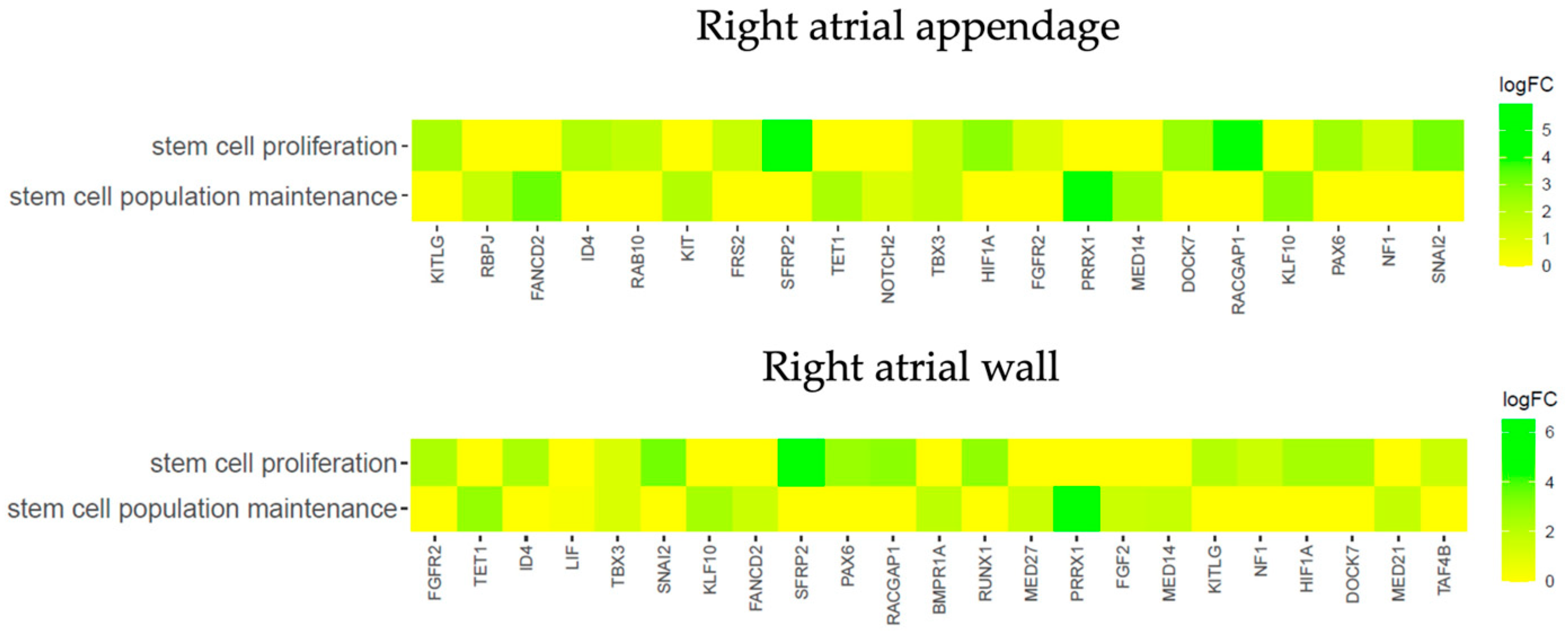
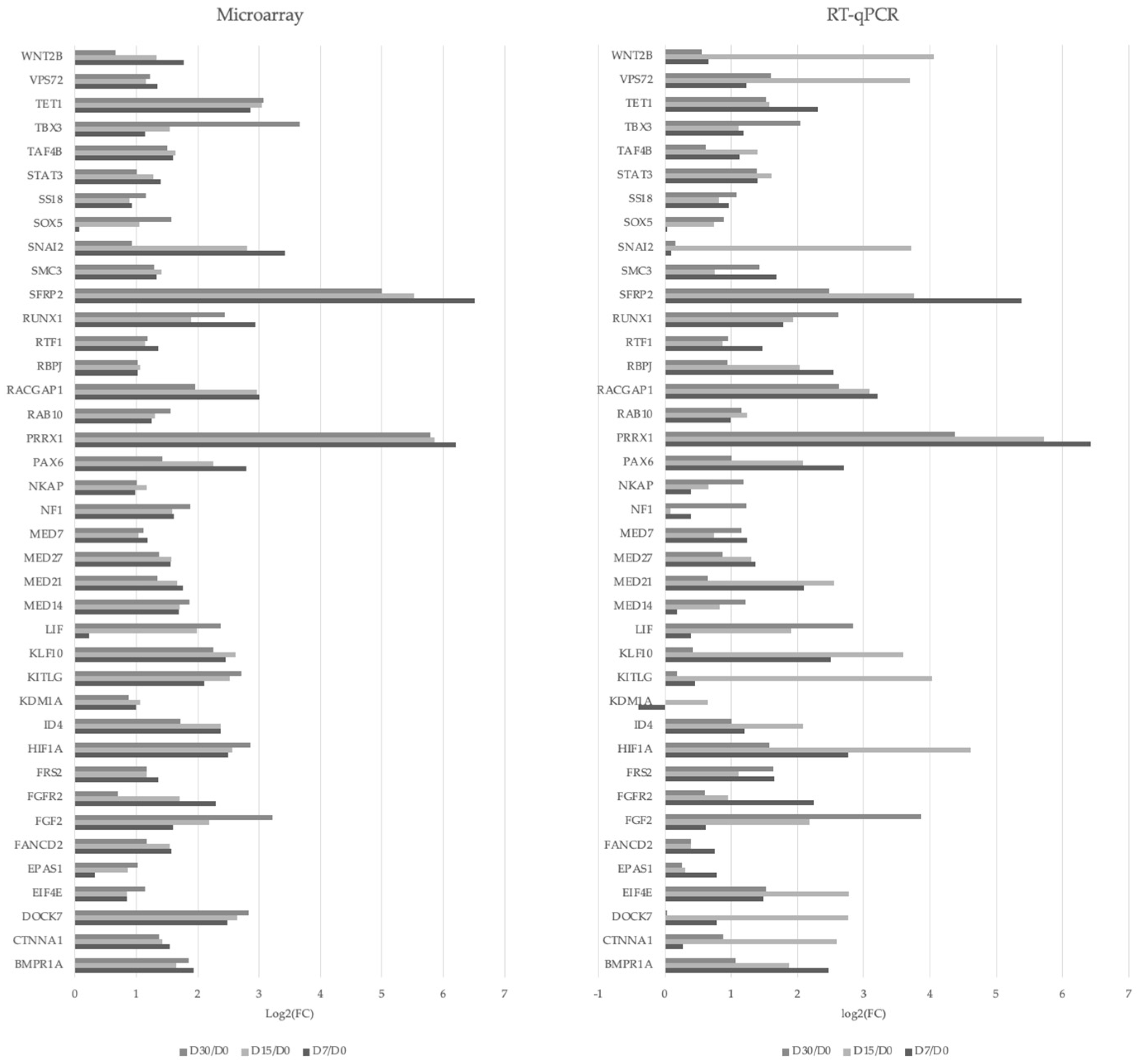
3.3. Transcriptomic Characterization of the Cardiomyocytes during Long-Term In Vitro Culture
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Braunwald, E. The War against Heart Failure: The Lancet Lecture. Lancet 2015, 385, 812–824. [Google Scholar] [CrossRef]
- Yancy, C.W.; Jessup, M.; Bozkurt, B.; Butler, J.; Casey, D.E.; Colvin, M.M.; Drazner, M.H.; Filippatos, G.S.; Fonarow, G.C.; Givertz, M.M.; et al. 2017 ACC/AHA/HFSA Focused Update of the 2013 ACCF/AHA Guideline for the Management of Heart Failure: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines and the Heart Failure Society of America. J. Am. Coll. Cardiol. 2017, 70, e137–e161. [Google Scholar]
- Ziaeian, B.; Fonarow, G.C. Epidemiology and Aetiology of Heart Failure. Nat. Rev. Cardiol. 2016, 13, 368–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yancy, C.W.; Jessup, M.; Bozkurt, B.; Butler, J.; Casey, D.E.; Drazner, M.H.; Fonarow, G.C.; Geraci, S.A.; Horwich, T.; Januzzi, J.L.; et al. 2013 ACCF/AHA Guideline for the Management of Heart Failure: A Report of the American College of Cardiology Foundation/American Heart Association Task Force on Practice Guidelines. Circulation 2013, 128, e147–e239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, O.; Bhardwaj, R.D.; Bernard, S.; Zdunek, S.; Barnabé-Heide, F.; Walsh, S.; Zupicich, J.; Alkass, K.; Buchholz, B.A.; Druid, H.; et al. Evidence for Cardiomyocyte Renewal in Humans. Science 2009, 324, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, O.; Zdunek, S.; Felker, A.; Salehpour, M.; Alkass, K.; Bernard, S.; Sjostrom, S.L.; Szewczykowska, M.; Jackowska, T.; Dos Remedios, C.; et al. Dynamics of Cell Generation and Turnover in the Human Heart. Cell 2015, 161, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.C.H.; Segers, V.F.M.; Davis, M.E.; MacGillivray, C.; Gannon, J.; Molkentin, J.D.; Robbins, J.; Lee, R.T. Evidence from a Genetic Fate-Mapping Study That Stem Cells Refresh Adult Mammalian Cardiomyocytes after Injury. Nat. Med. 2007, 13, 970–974. [Google Scholar] [CrossRef]
- Hsueh, Y.C.; Wu, J.M.F.; Yu, C.K.; Wu, K.K.; Hsieh, P.C.H. Prostaglandin E2 Promotes Post-Infarction Cardiomyocyte Replenishment by Endogenous Stem Cells. EMBO Mol. Med. 2014, 6, 496–503. [Google Scholar] [CrossRef]
- Walsh, S.; Pontén, A.; Fleischmann, B.K.; Jovinge, S. Cardiomyocyte Cell Cycle Control and Growth Estimation in Vivo-An Analysis Based on Cardiomyocyte Nuclei. Cardiovasc. Res. 2010, 86, 365–373. [Google Scholar] [CrossRef]
- Malliaras, K.; Zhang, Y.; Seinfeld, J.; Galang, G.; Tseliou, E.; Cheng, K.; Sun, B.; Aminzadeh, M.; Marbán, E. Cardiomyocyte Proliferation and Progenitor Cell Recruitment Underlie Therapeutic Regeneration after Myocardial Infarction in the Adult Mouse Heart. EMBO Mol. Med. 2013, 5, 191–209. [Google Scholar] [CrossRef]
- Le, T.; Chong, J. Cardiac Progenitor Cells for Heart Repair. Cell Death Discov. 2016, 2, 16052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Z.; Lal, S.; Le, T.Y.L.; dos Remedios, C.; Chong, J.J.H. Cardiac Stem Cells: Translation to Human Studies. Biophys. Rev. 2015, 7, 127–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalise, M.; Marino, F.; Cianflone, E.; Mancuso, T.; Marotta, P.; Aquila, I.; Torella, M.; Nadal-Ginard, B.; Torella, D. Heterogeneity of Adult Cardiac Stem Cells. Adv. Exp. Med. Biol. 2019, 1169, 141–178. [Google Scholar] [PubMed]
- Ellison, G.M.; Vicinanza, C.; Smith, A.J.; Aquila, I.; Leone, A.; Waring, C.D.; Henning, B.J.; Stirparo, G.G.; Papait, R.; Scarfò, M.; et al. Adult C-Kitpos Cardiac Stem Cells Are Necessary and Sufficient for Functional Cardiac Regeneration and Repair. Cell 2013, 154, 827–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison, G.M.; Torella, D.; Dellegrottaglie, S.; Perez-Martinez, C.; Perez De Prado, A.; Vicinanza, C.; Purushothaman, S.; Galuppo, V.; Iaconetti, C.; Waring, C.D.; et al. Endogenous Cardiac Stem Cell Activation by Insulin-like Growth Factor-1/Hepatocyte Growth Factor Intracoronary Injection Fosters Survival and Regeneration of the Infarcted Pig Heart. J. Am. Coll. Cardiol. 2011, 58, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Fransioli, J.; Bailey, B.; Gude, N.A.; Cottage, C.T.; Muraski, J.A.; Emmanuel, G.; Wu, W.; Alvarez, R.; Rubio, M.; Ottolenghi, S.; et al. Evolution of the C-kit-Positive Cell Response to Pathological Challenge in the Myocardium. Stem Cells 2008, 26, 1315–1324. [Google Scholar] [CrossRef] [Green Version]
- Hensley, M.T.; de Andrade, J.; Keene, B.; Meurs, K.; Tang, J.; Wang, Z.; Caranasos, T.G.; Piedrahita, J.; Li, T.S.; Cheng, K. Cardiac Regenerative Potential of Cardiosphere-Derived Cells from Adult Dog Hearts. J. Cell Mol. Med. 2015, 19, 1805–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.T.; White, A.J.; Matsushita, S.; Malliaras, K.; Steenbergen, C.; Zhang, Y.; Li, T.S.; Terrovitis, J.; Yee, K.; Simsir, S.; et al. Intramyocardial Injection of Autologous Cardiospheres or Cardiosphere-Derived Cells Preserves Function and Minimizes Adverse Ventricular Remodeling in Pigs with Heart Failure Post-Myocardial Infarction. J. Am. Coll. Cardiol. 2011, 57, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Malliaras, K.; Li, T.S.; Luthringer, D.; Terrovitis, J.; Cheng, K.; Chakravarty, T.; Galang, G.; Zhang, Y.; Schoenhoff, F.; Van Eyk, J.; et al. Safety and Efficacy of Allogeneic Cell Therapy in Infarcted Rats Transplanted with Mismatched Cardiosphere-Derived Cells. Circulation 2012, 125, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Messina, E.; De Angelis, L.; Frati, G.; Morrone, S.; Chimenti, S.; Fiordaliso, F.; Salio, M.; Battaglia, M.; Latronico, M.V.G.; Coletta, M.; et al. Isolation and Expansion of Adult Cardiac Stem Cells from Human and Murine Heart. Circ. Res. 2004, 95, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Hierlihy, A.M.; Seale, P.; Lobe, C.G.; Rudnicki, M.A.; Megeney, L.A. The Post-Natal Heart Contains a Myocardial Stem Cell Population. FEBS Lett. 2002, 530, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Pfister, O.; Mouquet, F.; Jain, M.; Summer, R.; Helmes, M.; Fine, A.; Colucci, W.S.; Liao, R. CD31- but Not CD31+ Cardiac Side Population Cells Exhibit Functional Cardiomyogenic Differentiation. Circ. Res. 2005, 97, 52–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamahara, K.; Fukushima, S.; Coppen, S.R.; Felkin, L.E.; Varela-Carver, A.; Barton, P.J.R.; Yacoub, M.H.; Suzuki, K. Heterogeneic Nature of Adult Cardiac Side Population Cells. Biochem. Biophys. Res. Commun. 2008, 371, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K.; Nagai, T.; Nishigaki, N.; Oyama, T.; Nishi, J.; Wada, H.; Sano, M.; Toko, H.; Akazawa, H.; Sato, T.; et al. Adult Cardiac Sca-1-Positive Cells Differentiate into Beating Cardiomyocytes. J. Biol. Chem. 2004, 279, 11384–11391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Vliet, P.; Roccio, M.; Smits, A.M.; Van Oorschot, A.A.M.; Metz, C.H.G.; Van Veen, T.A.B.; Sluijter, J.P.G.; Doevendans, P.A.; Goumans, M.J. Progenitor Cells Isolated from the Human Heart: A Potential Cell Source for Regenerative Therapy. Neth. Heart J. 2008, 16, 163–169. [Google Scholar] [CrossRef]
- Winter, E.M.; Grauss, R.W.; Hogers, B.; Van Tuyn, J.; Van Der Geest, R.; Lie-Venema, H.; Steijn, R.V.; Maas, S.; Deruiter, M.C.; Devries, A.A.F.; et al. Preservation of Left Ventricular Function and Attenuation of Remodeling after Transplantation of Human Epicardium-Derived Cells into the Infarcted Mouse Heart. Circulation 2007, 116, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Brown, J.; Kanarek, A.; Rajagopal, J.; Melton, D.A. In Vivo Reprogramming of Adult Pancreatic Exocrine Cells to β-Cells. Nature 2008, 455, 627–632. [Google Scholar] [CrossRef]
- Chong, J.J.H.; Chandrakanthan, V.; Xaymardan, M.; Asli, N.S.; Li, J.; Ahmed, I.; Heffernan, C.; Menon, M.K.; Scarlett, C.J.; Rashidianfar, A.; et al. Adult Cardiac-Resident MSC-like Stem Cells with a Proepicardial Origin. Cell Stem Cell 2011, 9, 527–540. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.J.H.; Reinecke, H.; Iwata, M.; Torok-Storb, B.; Stempien-Otero, A.; Murry, C.E. Progenitor Cells Identified by PDGFR-α Expression in the Developing and Diseased Human Heart. Stem Cells Dev. 2013, 22, 1932–1943. [Google Scholar] [CrossRef] [Green Version]
- Engleka, K.A.; Manderfield, L.J.; Brust, R.D.; Li, L.; Cohen, A.; Dymecki, S.M.; Epstein, J.A. Islet1 Derivatives in the Heart Are of Both Neural Crest and Second Heart Field Origin. Circ. Res. 2012, 110, 922–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laugwitz, K.L.; Moretti, A.; Lam, J.; Gruber, P.; Chen, Y.; Woodard, S.; Lin, L.Z.; Cai, C.L.; Lu, M.M.; Reth, M.; et al. Postnatal Isl1+ Cardioblasts Enter Fully Differentiated Cardiomyocyte Lineages. Nature 2005, 433, 647–653. [Google Scholar] [CrossRef] [Green Version]
- Moretti, A.; Caron, L.; Nakano, A.; Lam, J.T.; Bernshausen, A.; Chen, Y.; Qyang, Y.; Bu, L.; Sasaki, M.; Martin-Puig, S.; et al. Multipotent Embryonic Isl1+ Progenitor Cells Lead to Cardiac, Smooth Muscle, and Endothelial Cell Diversification. Cell 2006, 127, 1151–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolli, R.; Chugh, A.R.; D’Amario, D.; Loughran, J.H.; Stoddard, M.F.; Ikram, S.; Beache, G.M.; Wagner, S.G.; Leri, A.; Hosoda, T.; et al. Cardiac Stem Cells in Patients with Ischaemic Cardiomyopathy (SCIPIO): Initial Results of a Randomised Phase 1 Trial. Lancet 2011, 378, 1847–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makkar, R.R.; Smith, R.R.; Cheng, K.; Malliaras, K.; Thomson, L.E.J.; Berman, D.; Czer, L.S.C.; Marbán, L.; Mendizabal, A.; Johnston, P.V.; et al. Intracoronary Cardiosphere-Derived Cells for Heart Regeneration after Myocardial Infarction (CADUCEUS): A Prospective, Randomised Phase 1 Trial. Lancet 2012, 379, 895–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomczynski, P.; Sacchi, N. Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. The Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction: Twenty-Something Years On. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Koressaar, T.; Remm, M. Enhancements and Modifications of Primer Design Program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [Green Version]
- Kõressaar, T.; Lepamets, M.; Kaplinski, L.; Raime, K.; Andreson, R.; Remm, M. Primer3-Masker: Integrating Masking of Template Sequence with Primer Design Software. Bioinformatics 2018, 34, 1937–1938. [Google Scholar] [CrossRef] [Green Version]
- Yates, A.D.; Achuthan, P.; Akanni, W.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; et al. Ensembl 2020. Nucleic Acids Res. 2020, 48, D682–D688. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, D.T.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating High-Throughput Genomic Analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics Enrichment Tools: Paths toward the Comprehensive Functional Analysis of Large Gene Lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A Web Server for Functional Enrichment Analysis and Functional Annotation of Gene Lists (2021 Update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- The Gene Ontology Consortium; Acencio, M.; Lægreid, A.; Kuiper, M. The Gene Ontology Resource: 20 Years and Still GOing Strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar]
- Walter, W.; Sánchez-Cabo, F.; Ricote, M. GOplot: An R Package for Visually Combining Expression Data with Functional Analysis. Bioinformatics 2015, 31, 2912–2914. [Google Scholar] [CrossRef] [Green Version]
- Beltrami, A.P.; Barlucchi, L.; Torella, D.; Baker, M.; Limana, F.; Chimenti, S.; Kasahara, H.; Rota, M.; Musso, E.; Urbanek, K.; et al. Adult Cardiac Stem Cells Are Multipotent and Support Myocardial Regeneration. Cell 2003, 114, 763–776. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.; Bradfute, S.B.; Gallardo, T.D.; Nakamura, T.; Gaussin, V.; Mishina, Y.; Pocius, J.; Michael, L.H.; Behringer, R.R.; Garry, D.J.; et al. Cardiac Progenitor Cells from Adult Myocardium: Homing, Differentiation, and Fusion after Infarction. Proc. Natl. Acad. Sci. USA 2003, 100, 12313–12318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.R.; Barile, L.; Cho, H.C.; Leppo, M.K.; Hare, J.M.; Messina, E.; Giacomello, A.; Abraham, M.R.; Marbán, E. Regenerative Potential of Cardiosphere-Derived Cells Expanded from Percutaneous Endomyocardial Biopsy Specimens. Circulation 2007, 115, 896–908. [Google Scholar] [CrossRef] [Green Version]
- Nawrocki, M.J.; Jopek, K.; Kaczmarek, M.; Zdun, M.; Mozdziak, P.; Jemielity, M.; Perek, B.; Bukowska, D.; Kempisty, B. Transcriptomic Profile of Genes Regulating the Structural Organization of Porcine Atrial Cardiomyocytes during Primary In Vitro Culture. Genes 2022, 13, 1205. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Weiss, T.S.; Dayoub, R. Thy-1 (CD90)-Positive Hepatic Progenitor Cells, Hepatoctyes, and Non-Parenchymal Liver Cells Isolated from Human Livers. Methods Mol. Biol. 2017, 1506, 75–89. [Google Scholar]
- Nakamura, Y.; Muguruma, Y.; Yahata, T.; Miyatake, H.; Sakai, D.; Mochida, J.; Hotta, T.; Ando, K. Expression of CD90 on Keratinocyte Stem/Progenitor Cells. Br. J. Dermatl. 2006, 154, 1062–1070. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhan, X.Z.; Malola, J.; Li, Z.Y.; Pawar, J.S.; Zhang, H.T.; Zha, Z.G. The Multiple Roles of Thy-1 in Cell Differentiation and Regeneration. Differentiation 2020, 113, 38–48. [Google Scholar] [CrossRef]
- Pinto, A.R.; Ilinykh, A.; Ivey, M.J.; Kuwabara, J.T.; D’antoni, M.L.; Debuque, R.; Chandran, A.; Wang, L.; Arora, K.; Rosenthal, N.A.; et al. Revisiting Cardiac Cellular Composition. Circ. Res. 2016, 118, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Hudon-David, F.; Bouzeghrane, F.; Couture, P.; Thibault, G. Thy-1 Expression by Cardiac Fibroblasts: Lack of Association with Myofibroblast Contractile Markers. J. Mol. Cell Cardiol. 2007, 42, 991–1000. [Google Scholar] [CrossRef]
- Gambini, E.; Pompilio, G.; Biondi, A.; Alamanni, F.; Capogrossi, M.C.; Agrifoglio, M.; Pesce, M. C-Kit+ Cardiac Progenitors Exhibit Mesenchymal Markers and Preferential Cardiovascular Commitment. Cardiovasc. Res. 2011, 89, 362–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, D.R.; Zhang, Y.; Smith, R.R.; Cheng, K.; Terrovitis, J.; Malliaras, K.; Li, T.S.; White, A.; Makkar, R.; Marbán, E. Validation of the Cardiosphere Method to Culture Cardiac Progenitor Cells from Myocardial Tissue. PLoS ONE 2009, 4, e7195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, H.H.L.; Meher Homji, Z.; Gomes, R.S.M.; Sweeney, D.; Thomas, G.N.; Tan, J.J.; Zhang, H.; Perbellini, F.; Stuckey, D.J.; Watt, S.M.; et al. Human Cardiosphere-Derived Cells from Patients with Chronic Ischaemic Heart Disease Can Be Routinely Expanded from Atrial but Not Epicardial Ventricular Biopsies. J. Cardiovasc. Transl. Res. 2012, 5, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Tuyn, J.; Atsma, D.E.; Winter, E.M.; van der Velde-van Dijke, I.; Pijnappels, D.A.; Bax, N.A.M.; Knaän-Shanzer, S.; Gittenberger-de Groot, A.C.; Poelmann, R.E.; van der Laarse, A.; et al. Epicardial Cells of Human Adults Can Undergo an Epithelial-to-Mesenchymal Transition and Obtain Characteristics of Smooth Muscle Cells In Vitro. Stem Cells 2007, 25, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Ibrahim, A.; Hensley, M.T.; Shen, D.; Sun, B.; Middleton, R.; Liu, W.; Smith, R.R.; Marbán, E. Relative Roles of CD90 and C-Kit to the Regenerative Efficacy of Cardiosphere-Derived Cells in Humans and in a Mouse Model of Myocardial Infarction. J. Am. Heart Assoc. 2014, 3, e001260. [Google Scholar] [CrossRef] [Green Version]
- Senbanjo, L.T.; Chellaiah, M.A. CD44: A Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Thorne, R.F.; Wang, Y.; Zhang, Y.; Jing, X.; Zhang, X.D.; de Bock, C.E.; Oliveira, C.S. Evaluating Nuclear Translocation of Surface Receptors: Recommendations Arising from Analysis of CD44. Histochem. Cell Biol. 2020, 153, 77–87. [Google Scholar] [CrossRef]
- Bollini, S.; Vieira, J.M.N.; Howard, S.; Dubè, K.N.; Balmer, G.M.; Smart, N.; Riley, P.R. Re-Activated Adult Epicardial Progenitor Cells Are a Heterogeneous Population Molecularly Distinct from Their Embryonic Counterparts. Stem Cells Dev. 2014, 23, 1719–1730. [Google Scholar] [CrossRef]
- Lebrin, F.; Goumans, M.J.; Jonker, L.; Carvalho, R.L.C.; Valdimarsdottir, G.; Thorikay, M.; Mummery, C.; Arthur, H.M.; Ten Dijke, P. Endoglin Promotes Endothelial Cell Proliferation and TGF-β/ALK1 Signal Transduction. EMBO J. 2004, 23, 4018–4028. [Google Scholar] [CrossRef] [Green Version]
- Singh, E.; Phillips, H.M.; Arthur, H.M. Dynamic Changes in Endoglin Expression in the Developing Mouse Heart. Gene Expr. Patterns 2021, 39, 119165. [Google Scholar] [CrossRef]
- Arthur, H.M.; Ure, J.; Smith, A.J.H.; Renforth, G.; Wilson, D.I.; Torsney, E.; Charlton, R.; Parums, D.V.; Jowett, T.; Marchuk, D.A.; et al. Endoglin, an Ancillary TGFβ Receptor, Is Required for Extraembryonic Angiogenesis and Plays a Key Role in Heart Development. Dev. Biol. 2000, 217, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Sunagawa, Y.; Morimoto, T.; Takaya, T.; Kaichi, S.; Wada, H.; Kawamura, T.; Fujita, M.; Shimatsu, A.; Kita, T.; Hasegawa, K. Cyclin-Dependent Kinase-9 Is a Component of the P300/GATA4 Complex Required for Phenylephrine-Induced Hypertrophy in Cardiomyocytes. J. Biol. Chem. 2010, 285, 9556–9568. [Google Scholar] [CrossRef] [Green Version]
- Bisping, E.; Ikeda, S.; Kong, S.W.; Tarnavski, O.; Bodyak, N.; McMullen, J.R.; Rajagopal, S.; Son, J.K.; Ma, Q.; Springer, Z.; et al. Gata4 Is Required for Maintenance of Postnatal Cardiac Function and Protection from Pressure Overload-Induced Heart Failure. Proc. Natl. Acad. Sci. USA 2006, 103, 14471–14476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamura, S.; Izumiya, Y.; Araki, S.; Nakamura, T.; Kimura, Y.; Hanatani, S.; Yamada, T.; Ishida, T.; Yamamoto, M.; Onoue, Y.; et al. Cardiomyocyte Sirt (Sirtuin) 7 Ameliorates Stress-Induced Cardiac Hypertrophy by Interacting with and Deacetylating GATA4. Hypertension 2020, 75, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Furtado, M.B.; Costa, M.W.; Pranoto, E.A.; Salimova, E.; Pinto, A.R.; Lam, N.T.; Park, A.; Snider, P.; Chandran, A.; Harvey, R.P.; et al. Cardiogenic Genes Expressed in Cardiac Fibroblasts Contribute to Heart Development and Repair. Circ. Res. 2014, 114, 1422–1434. [Google Scholar] [CrossRef] [PubMed]
- Akazawa, H.; Komuro, I. Roles of Cardiac Transcription Factors in Cardiac Hypertrophy. Circ. Res. 2003, 92, 1079–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmanabhan, A.; Alexanian, M.; Linares-Saldana, R.; González-Terán, B.; Andreoletti, G.; Huang, Y.; Connolly, A.J.; Kim, W.; Hsu, A.; Duan, Q.; et al. BRD4 (Bromodomain-Containing Protein 4) Interacts with GATA4 (GATA Binding Protein 4) to Govern Mitochondrial Homeostasis in Adult Cardiomyocytes. Circulation 2020, 142, 2338–2355. [Google Scholar] [CrossRef]
- Jia, G.; Preussner, J.; Chen, X.; Guenther, S.; Yuan, X.; Yekelchyk, M.; Kuenne, C.; Looso, M.; Zhou, Y.; Teichmann, S.; et al. Single Cell RNA-Seq and ATAC-Seq Analysis of Cardiac Progenitor Cell Transition States and Lineage Settlement. Nat. Commun. 2018, 9, 4877. [Google Scholar] [CrossRef] [Green Version]
- Masino, A.M.; Gallardo, T.D.; Wilcox, C.A.; Olson, E.N.; Williams, R.S.; Garry, D.J. Transcriptional Regulation of Cardiac Progenitor Cell Populations. Circ. Res. 2004, 95, 389–397. [Google Scholar] [CrossRef] [Green Version]
- de Soysa, T.Y.; Ranade, S.S.; Okawa, S.; Ravichandran, S.; Huang, Y.; Salunga, H.T.; Schricker, A.; del Sol, A.; Gifford, C.A.; Srivastava, D. Single-Cell Analysis of Cardiogenesis Reveals Basis for Organ-Level Developmental Defects. Nature 2019, 572, 120–124. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhong, J.F.; Qiu, H.; MacLellan, W.R.; Marbán, E.; Wang, C. Epigenomic Reprogramming of Adult Cardiomyocyte-Derived Cardiac Progenitor Cells. Sci. Rep. 2015, 5, 17686. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, T.S.; Lee, S.T.; Wawrowsky, K.A.; Cheng, K.; Galang, G.; Malliaras, K.; Abraham, M.R.; Wang, C.; Marbán, E. Dedifferentiation and Proliferation of Mammalian Cardiomyocytes. PLoS ONE 2010, 5, e12559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, D.; Han, L.; Bauer, M.; Sanada, F.; Oikonomopoulos, A.; Hosoda, T.; Unno, K.; De Almeida, P.; Leri, A.; Wu, J.C. Dissecting the Molecular Relationship among Various Cardiogenic Progenitor Cells. Circ. Res. 2013, 112, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Chien, K.R.; Frisén, J.; Fritsche-Danielson, R.; Melton, D.A.; Murry, C.E.; Weissman, I.L. Regenerating the Field of Cardiovascular Cell Therapy. Nat. Biotechnol. 2019, 37, 232–237. [Google Scholar] [CrossRef]
- Orlic, D.; Kajstura, J.; Chimenti, S.; Bodine, D.M.; Leri, A.; Anversa, P. Bone Marrow Stem Cells Regenerate Infarcted Myocardium. Proc. Pediatr. Transplant. 2003, 7, 86–88. [Google Scholar] [CrossRef]
- Vagnozzi, R.J.; Sargent, M.A.; Lin, S.C.J.; Palpant, N.J.; Murry, C.E.; Molkentin, J.D. Genetic Lineage Tracing of Sca-1+ Cells Reveals Endothelial but Not Myogenic Contribution to the Murine Heart. Circulation 2018, 138, 2931–2939. [Google Scholar] [CrossRef]
- Müller, T.; Boileau, E.; Talyan, S.; Kehr, D.; Varadi, K.; Busch, M.; Most, P.; Krijgsveld, J.; Dieterich, C. Updated and Enhanced Pig Cardiac Transcriptome Based on Long-Read RNA Sequencing and Proteomics. J. Mol. Cell Cardiol. 2021, 150, 23–31. [Google Scholar] [CrossRef]
- Ou, Q.; Jacobson, Z.; Abouleisa, R.R.E.; Tang, X.L.; Hindi, S.M.; Kumar, A.; Ivey, K.N.; Giridharan, G.; El-Baz, A.; Brittian, K.; et al. Physiological Biomimetic Culture System for Pig and Human Heart Slices. Circ. Res. 2019, 125, 628–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathiyalagan, P.; Adamiak, M.; Mayourian, J.; Sassi, Y.; Liang, Y.; Agarwal, N.; Jha, D.; Zhang, S.; Kohlbrenner, E.; Chepurko, E.; et al. FTO-Dependent N6-Methyladenosine Regulates Cardiac Function during Remodeling and Repair. Circulation 2019, 139, 518–532. [Google Scholar] [CrossRef]
- Tu, L.N.; Timms, A.E.; Kibiryeva, N.; Bittel, D.; Pastuszko, A.; Nigam, V.; Pastuszko, P. Transcriptome Profiling Reveals Activation of Inflammation and Apoptosis in the Neonatal Striatum after Deep Hypothermic Circulatory Arrest. J. Thorac. Cardiovasc. Surg. 2019, 158, 882–890.e4. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, X.; Zheng, H.; Zhu, H.; Mai, W.; Huang, X.; Huang, Y. Multiple Roles of SFRP2 in Cardiac Development and Cardiovascular Disease. Int. J. Biol. Sci. 2020, 16, 730–738. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Cohen, E.D.; Morrisey, E.E. The Importance of Wnt Signaling in Cardiovascular Development. Pediatr. Cardiol. 2010, 31, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, Y.C.; Hodgkinson, C.P.; Gomez, J.A. The Role of Sfrp and DKK Proteins in Cardiomyocyte Development. Physiol. Rep. 2021, 9, e14678. [Google Scholar] [CrossRef]
- He, W.; Zhang, L.; Ni, A.; Zhang, Z.; Mirotsou, M.; Mao, L.; Pratt, R.E.; Dzau, V.J. Exogenously Administered Secreted Frizzled Related Protein 2 (Sfrp2) Reduces Fibrosis and Improves Cardiac Function in a Rat Model of Myocardial Infarction. Proc. Natl. Acad. Sci. USA 2010, 107, 21110–21115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtwright, A.; Siamakpour-Reihani, S.; Arbiser, J.L.; Banet, N.; Hilliard, E.; Fried, L.; Livasy, C.; Ketelsen, D.; Nepal, D.B.; Perou, C.M.; et al. Secreted Frizzle-Related Protein 2 Stimulates Angiogenesis via a Calcineurin/NFAT Signaling Pathway. Cancer Res. 2009, 69, 4621–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmeckpeper, J.; Verma, A.; Yin, L.; Beigi, F.; Zhang, L.; Payne, A.; Zhang, Z.; Pratt, R.E.; Dzau, V.J.; Mirotsou, M. Inhibition of Wnt6 by Sfrp2 Regulates Adult Cardiac Progenitor Cell Differentiation by Differential Modulation of Wnt Pathways. J. Mol. Cell Cardiol. 2015, 85, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgkinson, C.P.; Gomez, J.A.; Baksh, S.S.; Payne, A.; Schmeckpeper, J.; Pratt, R.E.; Dzau, V.J. Insights from Molecular Signature of in Vivo Cardiac C-Kit(+) Cells Following Cardiac Injury and β-Catenin Inhibition. J. Mol. Cell Cardiol. 2018, 123, 64–74. [Google Scholar] [CrossRef]
- Gomez, J.A.; Payne, A.; Pratt, R.E.; Hodgkinson, C.P.; Dzau, V.J. A Role for Sfrp2 in Cardiomyogenesis in Vivo. Proc. Natl. Acad. Sci. USA 2021, 118, e2103676118. [Google Scholar] [CrossRef]
- Deb, A.; Davis, B.H.; Guo, J.; Ni, A.; Huang, J.; Zhang, Z.; Mu, H.; Dzau, V.J. SFRP2 Regulates Cardiomyogenic Differentiation by Inhibiting a Positive Transcriptional Autofeedback Loop of Wnt3a. Stem Cells 2008, 26, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, M.; Yoshida, S.; Ueharu, H.; Chen, M.; Kato, T.; Kato, Y. PRRX1 and PRRX2 Distinctively Participate in Pituitary Organogenesis and a Cell-Supply System. Cell Tissue Res. 2014, 357, 323–335. [Google Scholar] [CrossRef]
- Leussink, B.; Brouwer, A.; El Khattabi, M.; Poelmann, R.E.; Gittenberger-de Groot, A.C.; Meijlink, F. Expression Patterns of the Paired-Related Homeobox Genes MHox/Prx1 and S8/Prx2 Suggest Roles in Development of the Heart and the Forebrain. Mech. Dev. 1995, 52, 51–64. [Google Scholar] [CrossRef]
- Bergwerff, M.; Gittenberger-De Groot, A.C.; Deruiter, M.C.; Van Iperen, L.; Meijlink, F.; Poelmann, R.E. Patterns of Paired-Related Homeobox Genes PRX1 and PRX2 Suggest Involvement in Matrix Modulation in the Developing Chick Vascular System. Dev. Dyn. 1998, 213, 59–70. [Google Scholar] [CrossRef]
- de Bakker, D.E.M.; Bouwman, M.; Dronkers, E.; Simões, F.C.; Riley, P.R.; Goumans, M.J.; Smits, A.M.; Bakkers, J. Prrx1b Restricts Fibrosis and Promotes Nrg1-Dependent Cardiomyocyte Proliferation during Zebrafish Heart Regeneration. Development 2021, 148, dev198937. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.J.; Qiu, X.B.; Wang, J.; Guo, Y.H.; Yang, C.X.; Li, L.; Gao, R.F.; Ke, Z.P.; Di, R.M.; Sun, Y.M.; et al. Prrx1 Loss-of-Function Mutations Underlying Familial Atrial Fibrillation. J. Am. Heart Assoc. 2021, 10, e023517. [Google Scholar] [CrossRef] [PubMed]
- Bosada, F.M.; Rivaud, M.R.; Uhm, J.S.; Verheule, S.; Van Duijvenboden, K.; Verkerk, A.O.; Christoffels, V.M.; Boukens, B.J. A Variant Noncoding Region Regulates Prrx1 and Predisposes to Atrial Arrhythmias. Circ. Res. 2021, 129, 420–434. [Google Scholar] [CrossRef]
- Hirose, K.; Kawashima, T.; Iwamoto, I.; Nosaka, T.; Kitamura, T. MgcRacGAP Is Involved in Cytokinesis through Associating with Mitotic Spindle and Midbody. J. Biol. Chem. 2001, 276, 5821–5828. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.M.; Fang, G. MgcRacGAP Controls the Assembly of the Contractile Ring and the Initiation of Cytokinesis. Proc. Natl. Acad. Sci. USA 2005, 102, 13158–13163. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.M.; Cao, X.Y.; He, P.; Li, J.; Feng, M.X.; Zhang, Y.L.; Zhang, X.L.; Wang, Y.H.; Yang, Q.; Zhu, L.; et al. Overexpression of Rac GTPase Activating Protein 1 Contributes to Proliferation of Cancer Cells by Reducing Hippo Signaling to Promote Cytokinesis. Gastroenterology 2018, 155, 1233–1249.e22. [Google Scholar] [CrossRef]
- Milde-Langosch, K.; Karn, T.; Müller, V.; Witzel, I.; Rody, A.; Schmidt, M.; Wirtz, R.M. Validity of the Proliferation Markers Ki67, TOP2A, and RacGAP1 in Molecular Subgroups of Breast Cancer. Breast Cancer Res. Treat 2013, 137, 57–67. [Google Scholar] [CrossRef]
- Imaoka, H.; Toiyama, Y.; Saigusa, S.; Kawamura, M.; Kawamoto, A.; Okugawa, Y.; Hiro, J.; Tanaka, K.; Inoue, Y.; Mohri, Y.; et al. RacGAP1 Expression, Increasing Tumor Malignant Potential, as a Predictive Biomarker for Lymph Node Metastasis and Poor Prognosis in Colorectal Cancer. Carcinogenesis 2015, 36, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, T.; Hirose, K.; Satoh, T.; Kaneko, A.; Ikeda, Y.; Kaziro, Y.; Nosaka, T.; Kitamura, T. MgcRacGAP Is Involved in the Control of Growth and Differentiation of Hematopoietic Cells. Blood 2000, 96, 2116–2124. [Google Scholar] [CrossRef]
- Edwards, J.J.; Rouillard, A.D.; Fernandez, N.F.; Wang, Z.; Lachmann, A.; Shankaran, S.S.; Bisgrove, B.W.; Demarest, B.; Turan, N.; Srivastava, D.; et al. Systems Analysis Implicates WAVE2 Complex in the Pathogenesis of Developmental Left-Sided Obstructive Heart Defects. JACC Basic Transl. Sci. 2020, 5, 376–386. [Google Scholar] [CrossRef]
- Hajra, K.M.; Chen, D.Y.S.; Fearon, E.R. The SLUG Zinc-Finger Protein Represses E-Cadherin in Breast Cancer. Cancer Res. 2002, 62, 1613–1618. [Google Scholar]
- Nieto, M.A.; Sargent, M.G.; Wilkinson, D.G.; Cooke, J. Control of Cell Behavior during Vertebrate Development by Slug, a Zinc Finger Gene. Science 1994, 264, 835–839. [Google Scholar] [CrossRef]
- Nieto, M.A. The Snail Superfamily of Zinc-Finger Transcription Factors. Nat. Rev Mol. Cell Biol. 2002, 3, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Takeichi, M.; Nimura, K.; Mori, M.; Nakagami, H.; Kaneda, Y. The Transcription Factors Tbx18 and Wt1 Control the Epicardial Epithelial-Mesenchymal Transition through Bi-Directional Regulation of Slug in Murine Primary Epicardial Cells. PLoS ONE 2013, 8, e57829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Wu, S.; Geng, Z.; Liu, L.; Zhang, S.; Wang, S.; Zhang, Y.; Huang, Y.; Zhang, B. LARP7 Suppresses Endothelial-to-Mesenchymal Transition by Coupling with TRIM28. Circ. Res. 2021, 129, 843–856. [Google Scholar] [CrossRef]
- Zhou, W.; Gross, K.M.; Kuperwasser, C. Molecular Regulation of Snai2 in Development and Disease. J. Cell Sci. 2019, 132, jcs235127. [Google Scholar] [CrossRef]
- Collantes, M.; Pelacho, B.; García-Velloso, M.J.; Gavira, J.J.; Abizanda, G.; Palacios, I.; Rodriguez-Borlado, L.; Álvarez, V.; Prieto, E.; Ecay, M.; et al. Non-Invasive in Vivo Imaging of Cardiac Stem/Progenitor Cell Biodistribution and Retention after Intracoronary and Intramyocardial Delivery in a Swine Model of Chronic Ischemia Reperfusion Injury. J. Transl. Med. 2017, 15, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.J.; Lewis, F.C.; Aquila, I.; Waring, C.D.; Nocera, A.; Agosti, V.; Nadal-Ginard, B.; Torella, D.; Ellison, G.M. Isolation and Characterization of Resident Endogenous C-Kit+ Cardiac Stem Cells from the Adult Mouse and Rat Heart. Nat. Protoc. 2014, 9, 1662–1681. [Google Scholar] [CrossRef]
- Dulak, J.; Szade, K.; Szade, A.; Nowak, W.; Józkowicz, A. Adult Stem Cells: Hopes and Hypes of Regenerative Medicine. Acta Biochim. Pol. 2015, 62, 329–337. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | 5′→3′ Sequence of the Forward Primer | 5′→3′ Sequence of the Reverse Primer | Product Length (bp) |
|---|---|---|---|
| ASPM | AACAGATTACGTCGTGCTGC | CTGTCTCTAGGCCGATTCGT | 205 |
| BMPR1A | ATTTGGGAAATGGCTCGTCG | CCCAGCATTCCGACATTAGC | 205 |
| CTNNA1 | CTTCTTGGCGGTCTCAGAGA | ACCCCTGGCTCATAGTTGTC | 171 |
| CTNNB1 | CCTCTCATCAAGGCTACCGT | AGGGCTCCAGTACAACCTTC | 218 |
| DDX6 | ACACTGCCTCAACACACTTT | GATTTCGATGTTCCTGCCTCA | 220 |
| DOCK7 | TGCTTACTCCACCTGCATCA | ATAGTCCTTCCGCATCAGGG | 198 |
| EIF4E | GCAAACCTTCGGCTGATCTC | ATTAGCCATCGTCCTCCTCG | 173 |
| EPAS1 | TGGGCTGGAGAGTTGAGAAG | AATAGTCTCCAGGGCTGCTG | 151 |
| FANCD2 | CAGTGTATGCCGCTCCTAGA | GAGATTGCCCAGCCAGAAAG | 246 |
| FGF2 | GCAGAAGAGAGAGGGGTTGT | CGTTCGTTTCAGTGCCACAT | 195 |
| FGFR2 | ATCGAGATTTAGCCGCCAGA | TCCCACATTAACACCCCGAA | 214 |
| FOXO1 | ATCACCAAGGCCATCGAGAG | AGTTCCTTCATTCTGCACGC | 189 |
| FRS2 | TGCCTTTAAGTGTGCTCGTG | TGGGTAAGTTCTGAGCAGCA | 182 |
| HIF1A | CTCAGTCGTCACAGTCTGGA | CCACCTCTTTTGGCAAGCAT | 212 |
| HMGA2 | ACCTCCCAATCTCCCGAAAG | GTTGTCCCTGGGCTGAAGT | 161 |
| ID4 | ACTACATCCTGGACCTGCAG | CCTCCCTCTCTAGTGCTCCT | 250 |
| KDM1A | GTCCAGTTTGTGCCACCTCT | TGCCTACATGCCCGAACAAA | 139 |
| KIT | ATTGTGAATCTTCTCGGCGC | ACTCCGGGTTTCATGTCCAT | 227 |
| KITLG | TGGCCAGTTCTATCCATGCA | TGGTCAGGGGTAAAGGCAAA | 175 |
| KLF10 | TGAGCTGCAGTTGGAAGTCT | TGTGAGGCTTGGCAGTATCT | 246 |
| LIF | GAACCTCTGAAAACTGCCGG | ACAGGAGTGATGGAAAGGGG | 151 |
| MCPH1 | CAGCCGACCATGTTCATCAG | AGTTCTCAGAGGCACAGACC | 217 |
| MED14 | CCACCATCCTCACTCACAGT | TCACTCCGGGTTCATTGGAA | 198 |
| MED21 | GTCCTCCTGCCTCTTTCAGT | CCTCCAGACATGTAGCAGCT | 228 |
| MED27 | ACTTGCATTCAGTCAACCGG | TTGTTCGACCACTTGTACGC | 172 |
| MED7 | ACTAGTAGAAGGCACGCGAA | ACTGCCCTTCACGGTGATTA | 150 |
| MTF2 | AAACTGCTGAGCCACCTTTG | TGCCTGGAAATGCTAGACGA | 172 |
| NF1 | GAATCCCCACCACAGTACCA | AAGGAGATGTGGGTGTCAGG | 226 |
| NKAP | GAGTCCCAGGAAGAGTTGCT | CATAGCTGCACCTTCACCAG | 162 |
| NOTCH2 | TTATGTCTCACCCCTGCCAG | ACTGTCCTGGAACGTCACAT | 246 |
| PAX6 | TTGCCCGAGAAAGACTAGCA | GTGGGTTGTGGGATTGGTTG | 196 |
| PRRX1 | GGACACACTACCCAGATGCT | TTTGAGGAGGGAAGCGTTCT | 155 |
| RAB10 | ATGGCGAAGAAGACGTACGA | AGGAGGTTGTGATCGTGTGA | 232 |
| RACGAP1 | ACCGCTGGAATACTGGAGTC | TGACAGGGAGCTGGATGAAG | 184 |
| RBPJ | ATGGGCAGTGGATGGAAGAA | TGTTTTGGCCGTGCAATAGT | 156 |
| RTF1 | CCAGGCGACAGTGTAAACCT | TCGCTGGCTGACTTGGAATT | 175 |
| RUNX1 | GTCCCAACTTCCTCTGCTCT | CTTCCACTCCGACCGACAAA | 226 |
| SFRP2 | GCTCCAAAGGTATGTGAAGCC | GGTCTTGCTCTTGGTCTCCA | 159 |
| SMC3 | TGGAGGACACTGAGGCAAAT | TCCTGTTGCCGCTCTAAGAA | 248 |
| SNAI2 | GCCGAGAAGTTTCAGTGCAA | GGGTCCGAATGTGCATCTTC | 169 |
| SOX5 | CAGCAGCAAGAACAGATCGC | AGCCAGTGTCCGTTGATCAG | 147 |
| SS18 | CGGATATGACCAGGGACAGT | CTTGCTGCGTTTCACCTGAT | 175 |
| STAT3 | AGCAGCAAAGAAGGAGGAGT | ACACGAGGATGTTGGTAGCA | 166 |
| TAF4B | GCCAGTCAGTTTCCTCAAGC | ACGAGTGTGCCAACCAATTC | 227 |
| TBX3 | AGGGTGTTCGATGACAGACA | GACGTGGTGGTGGAGATCTT | 233 |
| TET1 | TCTGGCAAGAAGAGAGCAGC | ATGGATGGGGTCGGTGAGTA | 248 |
| TGFB1 | ACCATGCCAATTTCTGCCTG | GAACGCACGATCATGTTGGA | 208 |
| VPS72 | TCCTTCGAGTACAAGAGCGG | GCACTTGCGCTTCTTATGGA | 188 |
| WNT2B | CAACGTGGGGACTTTGACTG | TGGCACTTACACTCCAGCTT | 185 |
| ACTB | CCCTGGAGAAGAGCTACGAG | CGTCGCACTTCATGATGGAG | 156 |
| GAPDH | CCAGAACATCATCCCTGCCT | CCTGCTTCACCACCTTCTTG | 185 |
| Step | Temperature (°C) | Time (s) | Number of Cycles | |
|---|---|---|---|---|
| Preincubation | 95 | 600 | 1 | |
| Amplification | Denaturation | 95 | 15 | 40 |
| Annealing | 58 | 15 | ||
| Elongation | 72 | 15 | ||
| Melting | 95 | 60 | 1 | |
| 40 | 60 | |||
| 70 | 1 | |||
| 95 | 1 | |||
| Cooling | 37 | 30 | 1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryl, R.; Nawrocki, M.J.; Jopek, K.; Kaczmarek, M.; Bukowska, D.; Antosik, P.; Mozdziak, P.; Zabel, M.; Dzięgiel, P.; Kempisty, B. Transcriptomic Characterization of Genes Regulating the Stemness in Porcine Atrial Cardiomyocytes during Primary In Vitro Culture. Genes 2023, 14, 1223. https://doi.org/10.3390/genes14061223
Bryl R, Nawrocki MJ, Jopek K, Kaczmarek M, Bukowska D, Antosik P, Mozdziak P, Zabel M, Dzięgiel P, Kempisty B. Transcriptomic Characterization of Genes Regulating the Stemness in Porcine Atrial Cardiomyocytes during Primary In Vitro Culture. Genes. 2023; 14(6):1223. https://doi.org/10.3390/genes14061223
Chicago/Turabian StyleBryl, Rut, Mariusz J. Nawrocki, Karol Jopek, Mariusz Kaczmarek, Dorota Bukowska, Paweł Antosik, Paul Mozdziak, Maciej Zabel, Piotr Dzięgiel, and Bartosz Kempisty. 2023. "Transcriptomic Characterization of Genes Regulating the Stemness in Porcine Atrial Cardiomyocytes during Primary In Vitro Culture" Genes 14, no. 6: 1223. https://doi.org/10.3390/genes14061223