
Molecular Mechanism of MYL4 Regulation of Skeletal Muscle Development in Pigs
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Samples
2.2. Cell Culture and Transfection
2.3. PCR
2.4. Real-Time Quantitative PCR
2.5. Cloning of 3′ Sequence of MYL4 cDNA
2.6. Cell Proliferation Assays
2.7. Western Blot
2.8. Flow Cytometry
2.9. Immunofluorescence Analysis
2.10. Bioinformatics Analysis
2.11. Statistical Analyses
3. Results
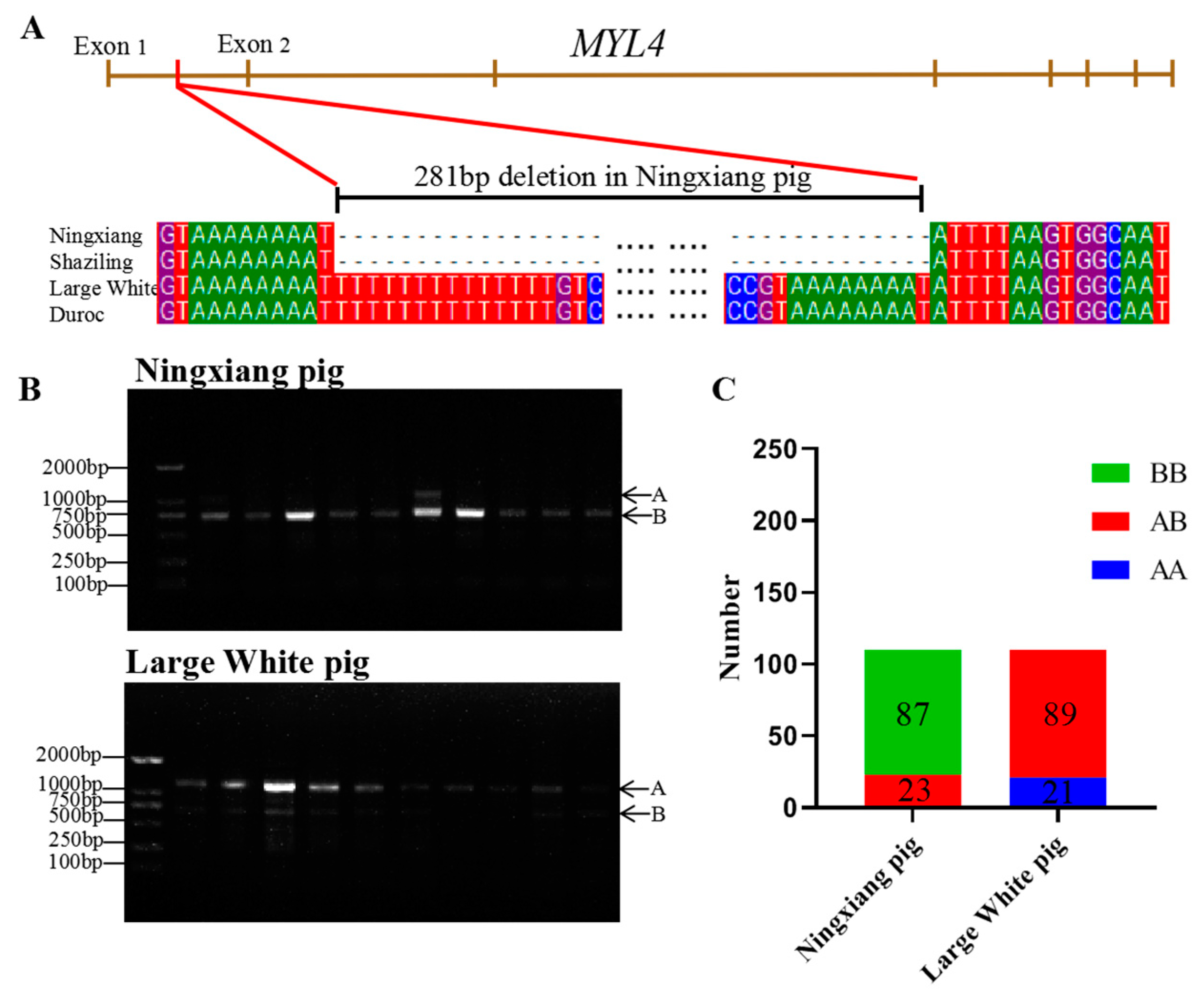
3.1. SV in MYL4 Gene of Ningxiang Pigs and Large White Pigs
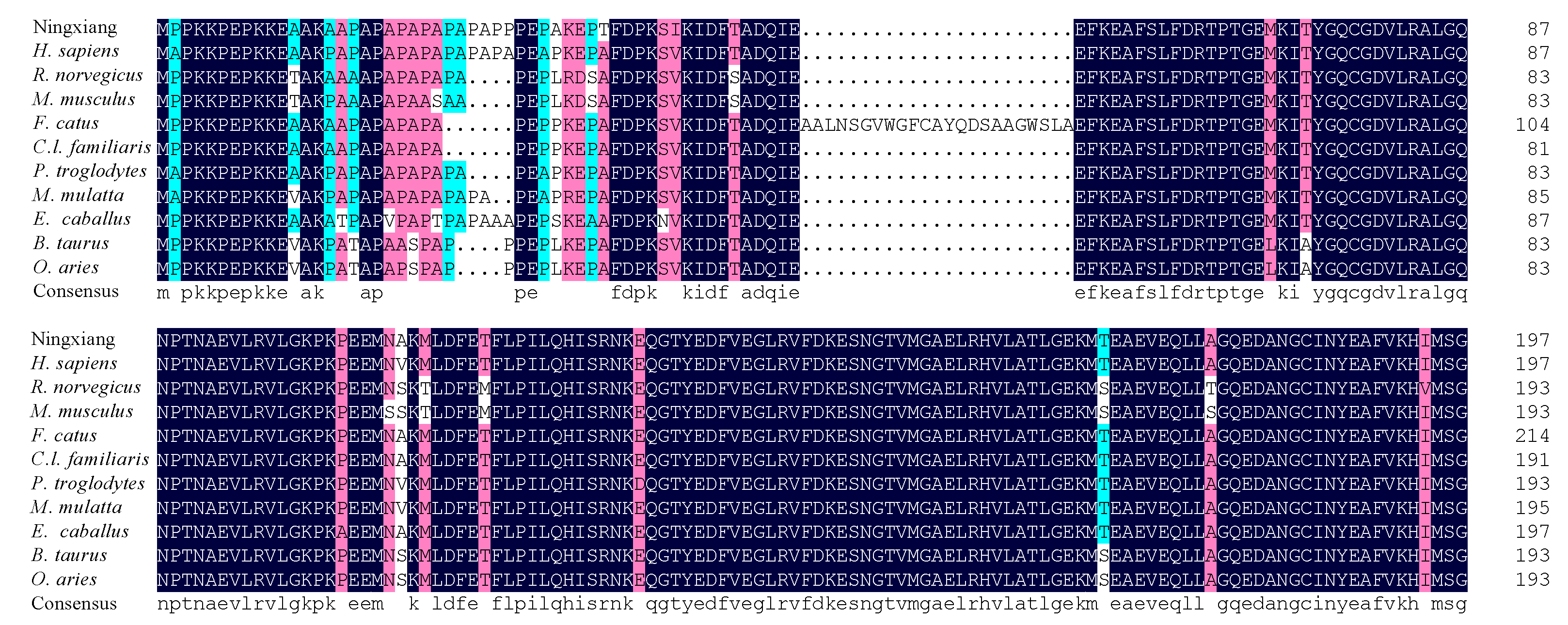
3.2. cDNA Cloning and Sequence Analysis of MYL4 in Ningxiang Pigs
3.3. Prediction of the Structures and Features of MYL4 Protein
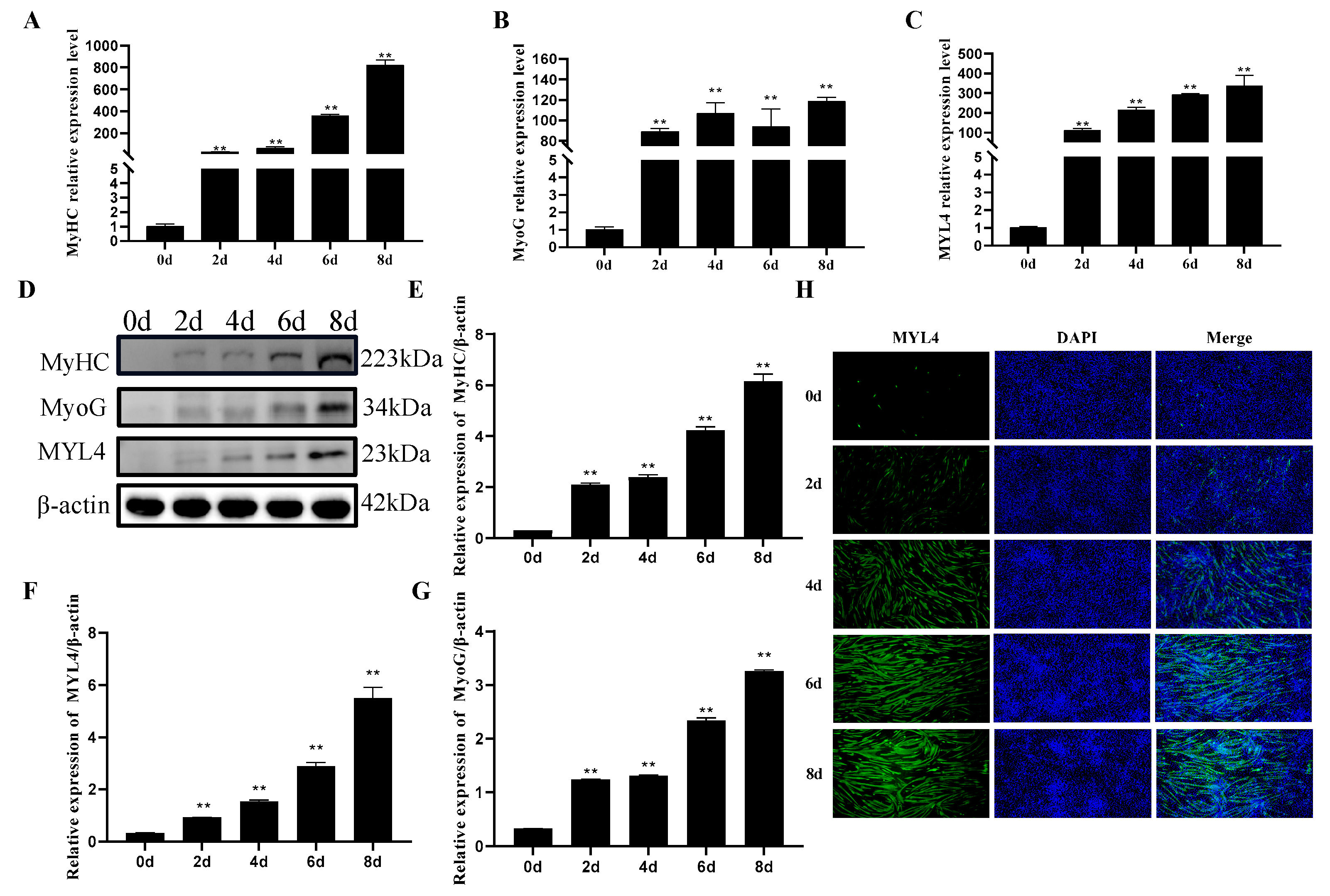
3.4. Profiles Analysis of MYL4 Gene Expression in Ningxiang Pigs and Large White Pigs
3.5. Knockdown and Overexpression of MYL4
3.6. MYL4 Inhibits C2C12 Cell Proliferation
3.7. MYL4 Promotes C2C12 Cells Apoptosis
3.8. Expression Pattern of MYL4 during the Differentiation of C2C12 Cells
3.9. MYL4 Promotes C2C12 Cells Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frontera, W.R.; Ochala, J. Skeletal Muscle: A Brief Review of Structure and Function. Calcif. Tissue Int. 2015, 96, 183–195. [Google Scholar] [CrossRef]
- Chal, J.; Pourquié, O. Making Muscle: Skeletal Myogenesis in Vivo and in Vitro. Development 2017, 144, 2104–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Lee, J.H.; Kang, N.; Kim, K.N.; Jeon, Y.J. The Effects of marine algal polyphenols, phlorotannins, on skeletal muscle growth in C2C12 muscle cells via Smad and IGF-1 signaling pathways. Mar. Drugs. 2021, 19, 266. [Google Scholar] [CrossRef] [PubMed]
- Relaix, F.; Bencze, M.; Borok, M.J.; Der Vartanian, A.; Gattazzo, F.; Mademtzoglou, D.; Perez-Diaz, S.; Prola, A.; Reyes-Fernandez, P.C.; Rotini, A.; et al. Perspectives on skeletal muscle stem cells. Nat. Commun. 2021, 12, 692. [Google Scholar] [CrossRef] [PubMed]
- Songsomboon, K.; Brenton, Z.; Heuser, J.; Kresovich, S.; Shakoor, N.; Mockler, T.; Cooper, E.A. Genomic patterns of structural variation among diverse genotypes of Sorghum bicolor and a potential role for deletions in local adaptation. G3 2021, 11, jkab154. [Google Scholar] [CrossRef]
- Conrad, D.F.; Pinto, D.; Redon, R.; Feuk, L.; Gokcumen, O.; Zhang, Y.; Aerts, J.; Andrews, T.D.; Barnes, C.; Campbell, P.; et al. Origins and Functional Impact of Copy Number Variation in the Human Genome. Nature 2010, 464, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Jiang, J.; He, J.; Liu, H.; Han, L.; Gong, Y.; Li, B.; Yu, Z.; Tang, S.; Zhang, Y.; et al. Long-read Assembly of the Chinese Indigenous Ningxiang Pig Genome and Identification of Genetic Variations in fat Metabolism Among Different Breeds. Mol. Ecol. Resour. 2022, 22, 1508–1520. [Google Scholar] [CrossRef]
- Larkin, D.M.; Astakhova, I.M.; Kozhin, A.Y.; Karamysheva, T.V.; Rubtsov, N.B.; Kuznetsov, S.B.; Zhdanova, N.S. The Structure of a Conserved Region of Porcine Genome, Represented in Human Genome by Chromosome 17. Genetika 2004, 40, 961–967. [Google Scholar] [CrossRef]
- Cohen-Haguenauer, O.; Barton, P.J.; Van Cong, N.; Cohen, A.; Masset, M.; Buckingham, M.; Frézal, J. Chromosomal Assignment of two Myosin Alkali Light-chain Genes Encoding the Ventricular/slow Skeletal Muscle Isoform and the Atrial/fetal Muscle Isoform (MYL3, MYL4). Hum. Genet. 1989, 81, 278–282. [Google Scholar] [CrossRef]
- Barton, P.J.; Robert, B.; Fiszman, M.Y.; Leader, D.P.; Buckingham, M.E. The Same Myosin Alkali Light Chain Gene is Expressed in Adult Cardiac Atria and in Fetal Skeletal Muscle. J. Muscle Res. Cell Motil. 1985, 6, 461–475. [Google Scholar] [CrossRef]
- Wang, T.Y.; Arking, D.E.; Maleszewski, J.J.; Fox-Talbot, K.; Nieuwenhuis, T.O.; Santhanam, L.; Virmani, R.; Rosenberg, A.Z.; Halushka, M.K. Human Cardiac Myosin Light Chain 4 (MYL4) Mosaic Expression Patterns Vary by Sex. Sci. Rep. 2019, 9, 12681. [Google Scholar] [CrossRef] [Green Version]
- Gudbjartsson, D.F.; Holm, H.; Sulem, P.; Masson, G.; Oddsson, A.; Magnusson, O.T.; Saemundsdottir, J.; Helgadottir, H.T.; Helgason, H.; Johannsdottir, H.; et al. A Frameshift Deletion in the Sarcomere Gene MYL4 Causes Early-onset Familial Atrial Fibrillation. Eur. Heart J. 2017, 38, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, H.; Tao, Y.; Xu, Z.; Lai, H. Relationship Between Serum miR-106 and MYL4 Levels and the Prevalence, Risk Stratification, and Prognosis of Atrial Fibrillation. J. Immunol. Res. 2022, 2022, 1069866. [Google Scholar] [CrossRef]
- Morano, I.; Hadicke, K.; Haase, H.; Böhm, M.; Erdmann, E.; Schaub, M.C. Changes in Essential Myosin Light Chain Isoform Expression Provide a Molecular Basis for Isometric Force Regulation in the Failing Human Heart. J. Mol. Cell. Cardiol. 1997, 29, 1177–1187. [Google Scholar] [CrossRef]
- Periasamy, M.; Strehler, E.E.; Garfinkel, L.I.; Gubits, R.M.; Ruiz-Opazo, N.; Nadal-Ginard, B. Fast Skeletal Muscle Myosin Light Chains 1 and 3 are Produced From a Single Gene by a Combined Process of Differential RNA Transcription and Splicing. J. Biol. Chem. 1984, 259, 13595–13604. [Google Scholar] [CrossRef]
- Ye, M.; Ye, F.; He, L.; Luo, B.; Yang, F.; Cui, C.; Zhao, X.; Yin, H.; Li, D.; Xu, H.; et al. Transcriptomic Analysis of Chicken Myozenin 3 Regulation Reveals its Potential Role in Cell Proliferation. PLoS ONE 2017, 12, e189476. [Google Scholar] [CrossRef] [Green Version]
- Ghazizadeh, Z.; Kiviniemi, T.; Olafsson, S.; Plotnick, D.; Beerens, M.E.; Zhang, K.; Gillon, L.; Steinbaugh, M.J.; Barrera, V.; Sui, S.H.; et al. Metastable Atrial State Underlies the Primary Genetic Substrate for MYL4 Mutation-Associated Atrial Fibrillation. Circulation 2020, 141, 301–312. [Google Scholar] [CrossRef]
- Dong, S.; Han, Y.; Zhang, J.; Ye, Y.; Duan, M.; Wang, K.; Wei, M.; Chamba, Y.; Shang, P. Haplotypes Within the Regulatory Region of MYL4 are Associated with Pig Muscle Fiber Size. Gene 2023, 850, 146934. [Google Scholar] [CrossRef]
- Tang, Z.; Li, Y.; Wan, P.; Li, X.; Zhao, S.; Liu, B.; Fan, B.; Zhu, M.; Yu, M.; Li, K. LongSAGE Analysis of Skeletal Muscle at Three Prenatal Stages in Tongcheng and Landrace Pigs. Genome Biol. 2007, 8, R115. [Google Scholar] [CrossRef] [Green Version]
- Nataf, Y.; Bahari, L.; Kahel-Raifer, H.; Borovok, I.; Lamed, R.; Bayer, E.A.; Sonenshein, A.L.; Shoham, Y. Clostridium Thermocellum Cellulosomal Genes are Regulated by Extracytoplasmic Polysaccharides via Alternative Sigma Factors. Proc. Natl. Acad. Sci. USA 2010, 107, 18646–18651. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.X.; Tang, Z.L.; Deng, H.; Li, Y.; Chu, M.X.; Ma, Y.H.; Li, K. Clone of MYL4 Gene and Its Expression Analysis in Pig Embryo Skeletal Muscle. J. Agric. Biotechnol. 2008, 04, 580–585. [Google Scholar]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budsuren, U.; Ulaangerel, T.; Shen, Y.; Liu, G.; Davshilt, T.; Yi, M.; Bold, D.; Zhang, X.; Bai, D.; Dorjgotov, D.; et al. MSTN Regulatory Network in Mongolian Horse Muscle Satellite Cells Revealed with miRNA Interference Technologies. Genes 2022, 13, 1836. [Google Scholar] [CrossRef] [PubMed]
- Schaub, M.C.; Hefti, M.A.; Zuellig, R.A.; Morano, I. Modulation of Contractility in Human Cardiac Hypertrophy by Myosin Essential Light Chain Isoforms. Cardiovasc. Res. 1998, 37, 381–404. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, J.; Wang, G.; Ji, Z.; Hou, L.; Liu, Z.; Chao, T. Molecular Cloning and mRNA Expression Analysis of Sheep MYL3 and MYL4 Genes. Gene 2016, 577, 209–214. [Google Scholar] [CrossRef]
- Soussi-Yanicostas, N.; Butler-Browne, G.S. Transcription of the Embryonic Myosin Light Chain Gene is Restricted to type II Muscle Fibers in Human Adult Masseter. Dev. Biol. 1991, 147, 374–380. [Google Scholar] [CrossRef]
- Chiang, C.; Scott, A.J.; Davis, J.R.; Tsang, E.K.; Li, X.; Kim, Y.; Hadzic, T.; Damani, F.N.; Ganel, L.; GTEx Consortium; et al. The Impact of Structural Variation on Human Gene Expression. Nat. Genet. 2017, 49, 692–699. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Wu, X.; Xie, C.; Xiao, D.; Zhang, B. Meat Quality and Fatty Acid Profiles of Chinese Ningxiang Pigs Following Supplementation with N-Carbamylglutamate. Animals 2020, 10, 88. [Google Scholar] [CrossRef] [Green Version]
- Velotto, S.; Claudia, V.; Antonio, C. Muscle Fibre Types, Fat Deposition and Fatty Acid Profile of Casertana Versus Large White Pig. Anim. Sci. Pap. Rep. 2012, 30, 35–44. [Google Scholar]
- Sharp, A.J.; Cheng, Z.; Eichler, E.E. Structural Variation of the Human Genome. Annu. Rev. Genomics Hum. Genet. 2006, 7, 407–442. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Li, S.T.; Yao, W.Y.; Xie, C.D.; Chen, Z.; Zeng, Z.J.; Wang, D.; Xu, K.; Shen, Z.J.; Mu, Y.; et al. The Meishan pig Genome Reveals Structural Variation-mediated Gene Expression and Phenotypic Divergence Underlying Asian Pig Domestication. Mol. Ecol. Resour. 2021, 21, 2077–2092. [Google Scholar] [CrossRef]
- Morgan, J.E.; Moore, S.E.; Walsh, F.S.; Partridge, T.A. Formation of skeletal muscle in vivo from the mouse C2 cell line. J. Cell Sci. 1992, 102, 779–787. [Google Scholar] [CrossRef]
- Lyons, G.E.; Ontell, M.; Cox, R.; Buckingham, M. The Expression of Myosin Genes in Developing Skeletal Muscle in the Mouse Embryo. J. Cell Biol. 1990, 111, 1465–1476. [Google Scholar] [CrossRef] [Green Version]
- Keller, L.R.; Emerson, C.J. Synthesis of Adult Myosin Light Chains by Embryonic Muscle Cultures. Proc. Natl. Acad. Sci. USA 1980, 77, 1020–1024. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Xie, H.; Xu, Y.; Li, X.; Deng, L.; Chen, X.; Qiu, L.; Yang, Z. An Experimental Study of the Role of Myosin Light Chain in Myogenesis in Vitro. Chin. J. Reparative Reconstr. Surg. 2008, 22, 753–758. [Google Scholar]
- Sutherland, C.J.; Esser, K.A.; Elsom, V.L.; Gordon, M.L.; Hardeman, E.C. Identification of a Program of Contractile Protein Gene Expression Initiated Upon Skeletal Muscle Differentiation. Dev. Dyn. 1993, 196, 25–36. [Google Scholar] [CrossRef]
- Sutherland, C.J.; Elsom, V.L.; Gordon, M.L.; Dunwoodie, S.L.; Hardeman, E.C. Coordination of Skeletal Muscle Gene Expression Occurs Late in Mammalian Development. Dev. Biol. 1991, 146, 167–178. [Google Scholar] [CrossRef]
- Moldovan, G.L.; Pfander, B.; Jentsch, S. PCNA, the Maestro of the Replication Fork. Cell 2007, 129, 665–679. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, A.; Filipkowski, R.K.; Rylski, M.; Wilczynski, G.M.; Konopacki, F.A.; Jaworski, J.; Ciemerych, M.A.; Sicinski, P.; Kaczmarek, L. The Critical Role of cyclin D2 in Adult Neurogenesis. J. Cell Biol. 2004, 167, 209–213. [Google Scholar] [CrossRef]
- Zhang, J.; Hua, C.; Zhang, Y.; Wei, P.; Tu, Y.; Wei, T. KAP1-associated Transcriptional Inhibitory Complex Regulates C2C12 Myoblasts Differentiation and Mitochondrial Biogenesis via miR-133a Repression. Cell Death Dis. 2020, 11, 732. [Google Scholar] [CrossRef]
- Du, C.; Jin, Y.Q.; Qi, J.J.; Ji, Z.X.; Li, S.Y.; An, G.S.; Jia, H.T.; Ni, J.H. Effects of Myogenin on Expression of Late Muscle Genes Through MyoD-dependent Chromatin Remodeling Ability of Myogenin. Mol. Cells 2012, 34, 133–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| siRNA | Forward (5′→3′) | Reverse (5′→3′) |
|---|---|---|
| siRNA1 | GCUGACCAGAUCGAAGAAUTT | AUUCUUCGAUCUGGUCAGCTT |
| siRNA2 | GCUGCGGGUCUUUGACAAATT | UUUGUCAAACACCCGCAGCTT |
| siRNA3 | GCAUCAACUAUGAAGCCUUTT | AAGGCUUCAUAGUUGAUGCTT |
| Gene | Primer Sequence (5′→3′) | Tm (°C) | Length (bp) |
|---|---|---|---|
| ssc-MYL4 | F: CAGCCCAGTCTCCCATCT | 60 | 375 |
| R: GCAGCACCTCGGCATTAG | |||
| mmu-MYL4 | F: GAAACCCGAGCCTAAGAA | 60 | 175 |
| R: AGTCCGGTCAAACAATGAA | |||
| CCNE | F: CCTCTGCTCGGGTGTTGTAG | 60 | 72 |
| R: TCTGCATCCCACACTTGCTC | |||
| CCND | F: TCAAGTGTGACCCGGACTG | 60 | 235 |
| R: GCTCCTTCCTCTTTGCGGG | |||
| PCNA | F: GCCGAGACCTTAGCCACATT | 60 | 229 |
| R: GTAGGAGACAGTGGAGTGGC | |||
| CDK4 | F: CGAGCGTAAGGCTGATGGAT | 60 | 177 |
| R: CCAGGCCGCTTAGAAACTGA | |||
| Caspase3 | F: GCTTGGAACGGTACGCTAAG | 60 | 112 |
| R: CCACTGACTTGCTCCCATGT | |||
| BAX | F: CCAGGATGCGTCCACCAA | 60 | 196 |
| R: AAAGTAGAAGAGGGCAACCAC | |||
| MyoG | F: CAATGCACTGGAGTTCGGT | 60 | 134 |
| R: CTGGGAAGGCAACAGACAT | |||
| MyHC | F: CGGTCGAAGTTGCATCCCT | 60 | 141 |
| R: GAGCCTCGATTCGCTCCTTT | |||
| Myf5 | F: CAGGAATGCCATCCGCTACA | 60 | 78 |
| R: CCCGGCAGGCTGTAATAGTT | |||
| MyoD | F: AAGACGACTCTCACGGCTTG | 60 | 169 |
| R: GCAGGTCTGGTGAGTCGAAA | |||
| Gapdh | F: AGGGCATCCTGGGCTACACT | 60 | 166 |
| R: TCCACCACCCTGTTGCTGTAG | |||
| MYL1 | F: GGGAACCCCAGCAATGAAGA | 60 | 132 |
| R: GAAGACACGCAGACCCTCAA | |||
| MYL2 | F: CTGACGTCACCGGCAATCTT | 60 | 195 |
| R: GGCAACTCCCATCTTCTCCT | |||
| MYL3 | F: GCCAAGCATCTCCCAACCAT | 60 | 115 |
| R: GGGCCAGGAAAGACTACCAC |
| Breeds | Genotype Frequency (%) | Gene Frequency (%) | χ2 | p | PIC | |||
|---|---|---|---|---|---|---|---|---|
| AA | AB | BB | A | B | ||||
| Ningxiang | 0 | 20.91 | 79.09 | 10 | 90 | 1.50 | >0.05 | 0.17 |
| Large White | 19.09 | 80.91 | 0 | 59.55 | 40.45 | 50.77 | <0.05 | 0.37 |
| Physicochemical Properties | MYL4 |
|---|---|
| Amino acid | 197 |
| Molecular weight (Da) | 21,606.64 |
| Theoretical pI | 4.98 |
| Number of atoms | 3031 |
| Asp + Glu | 32 |
| Arg + Lys | 23 |
| Extinction coefficients | 4595 |
| Aliphatic index | 67.97 |
| Instability index | 58.38 |
| Grand average of hydropathicity | −0.535 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Yu, Z.; Ai, N.; Liufu, S.; Liu, X.; Chen, B.; Li, X.; Jiang, J.; Zhang, Y.; Ma, H.; et al. Molecular Mechanism of MYL4 Regulation of Skeletal Muscle Development in Pigs. Genes 2023, 14, 1267. https://doi.org/10.3390/genes14061267
Xu X, Yu Z, Ai N, Liufu S, Liu X, Chen B, Li X, Jiang J, Zhang Y, Ma H, et al. Molecular Mechanism of MYL4 Regulation of Skeletal Muscle Development in Pigs. Genes. 2023; 14(6):1267. https://doi.org/10.3390/genes14061267
Chicago/Turabian StyleXu, Xueli, Zonggang Yu, Nini Ai, Sui Liufu, Xiaolin Liu, Bohe Chen, Xintong Li, Jun Jiang, Yuebo Zhang, Haiming Ma, and et al. 2023. "Molecular Mechanism of MYL4 Regulation of Skeletal Muscle Development in Pigs" Genes 14, no. 6: 1267. https://doi.org/10.3390/genes14061267