
Identifying Genetic Architecture of Carcass and Meat Quality Traits in a Ningxiang Indigenous Pig Population

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phenotypes and Genotyping
2.2. Genotype Imputation and Quality Control
2.3. Statistical Method
2.3.1. Estimation of Genetic Parameters
2.3.2. Principal Component Analysis
2.3.3. Genome-Wide Association Study
2.4. Linkage Disequilibrium Analysis
2.5. Candidate Genes Related to Significant SNPs
3. Results
3.1. Descriptive Statistics of Phenotypes
3.2. Estimates of Genetic Parameters
3.3. GWAS Results and Gene Annotation
3.3.1. Carcass Trait
3.3.2. Meat Quality Trait
3.4. LD Block Analysis
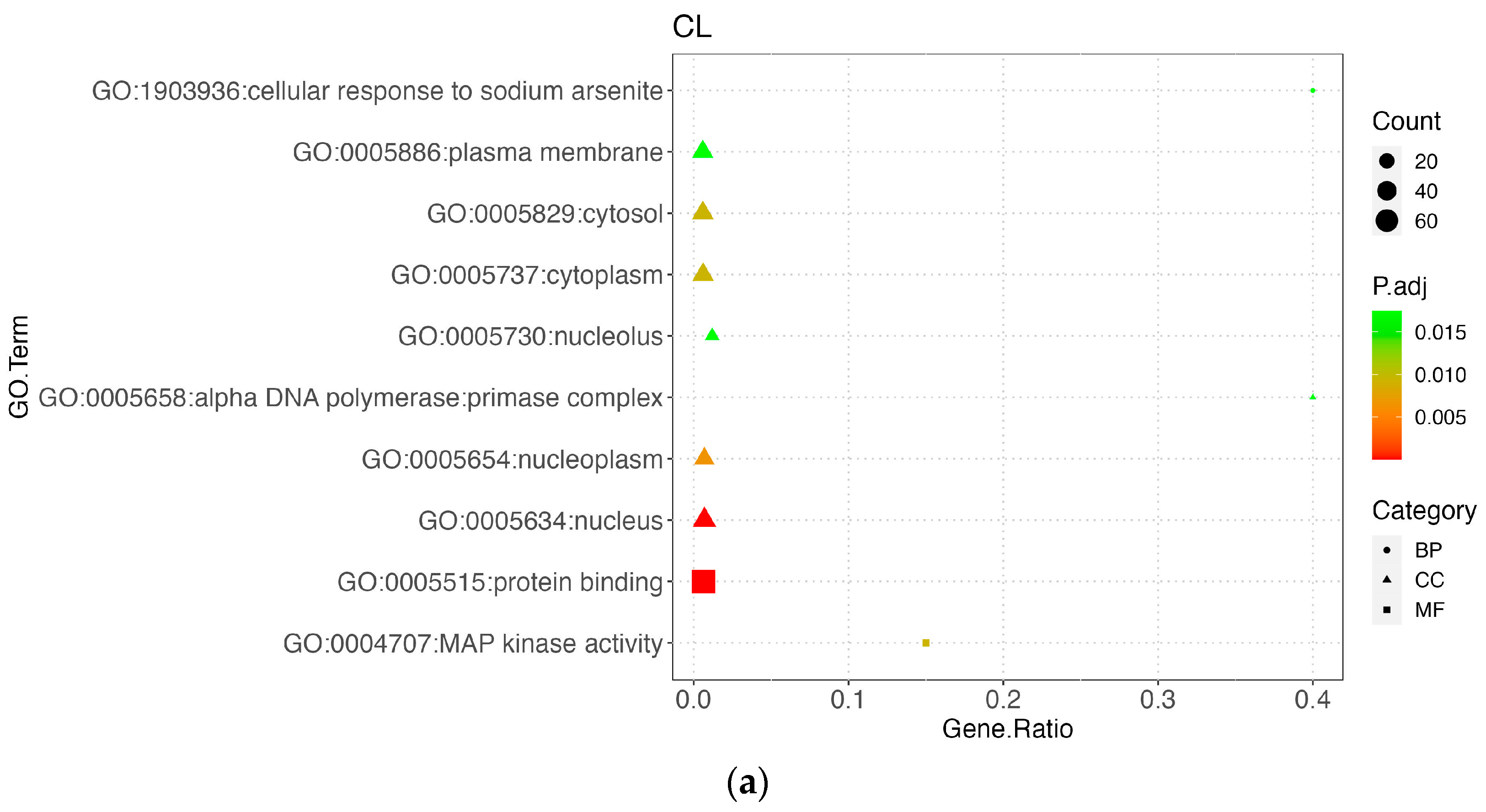
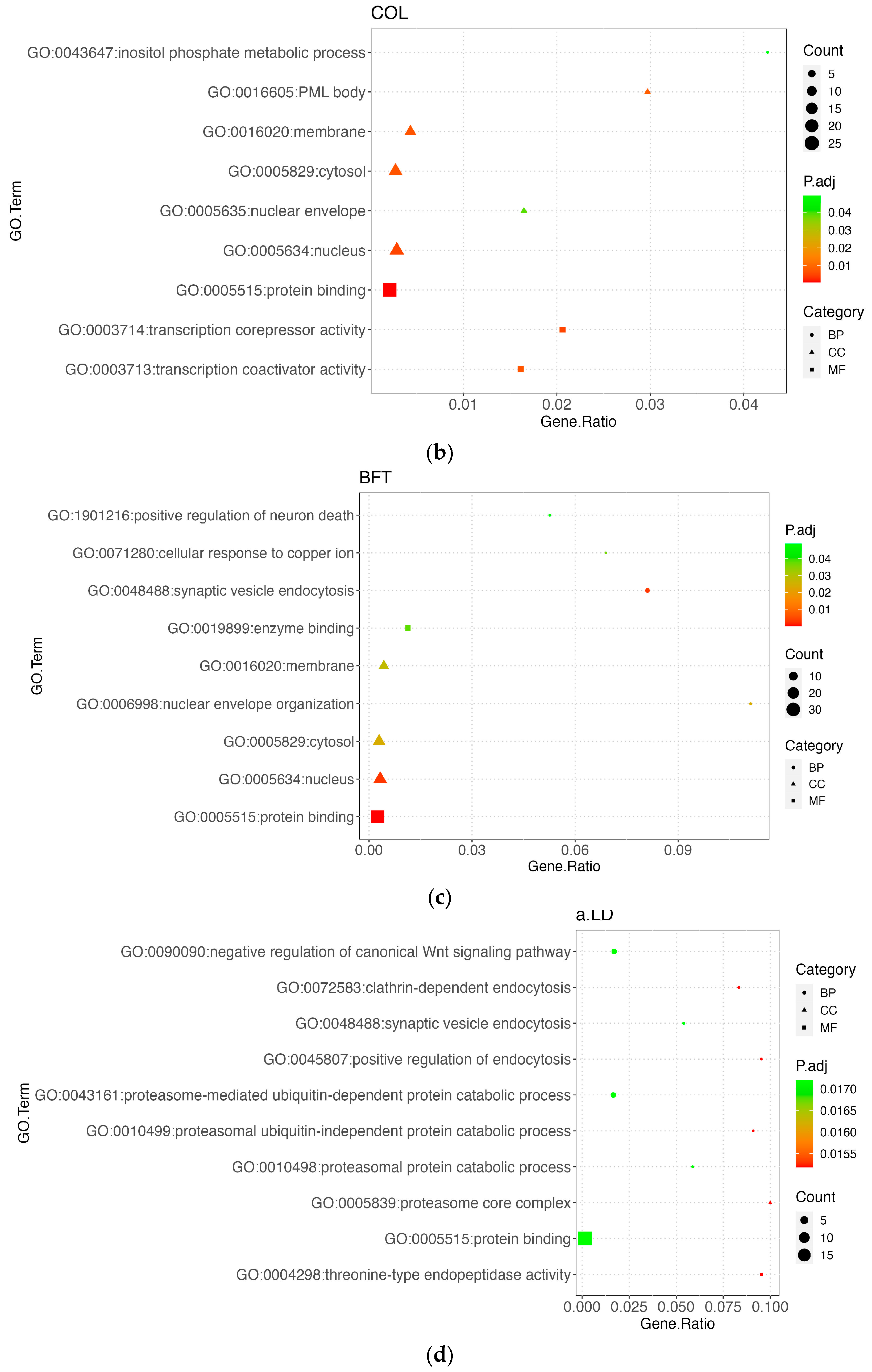
3.5. Functional Enrichment Results
3.5.1. Carcass Trait
3.5.2. Meat Quality Trait
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, J.; Huang, L.; Yang, M.; Fan, Y.; Li, L.; Fang, S.; Deng, W.; Cui, L.; Zhang, Z.; Ai, H.; et al. Possible Introgression of the VRTN Mutation Increasing Vertebral Number, Carcass Length and Teat Number from Chinese Pigs into European Pigs. Sci. Rep. 2016, 6, 19240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Zhang, W.; Cai, J.; Ni, Y.; Xiao, L.; Zhang, J. Transcriptome Analysis in Comparing Carcass and Meat Quality Traits of Jiaxing Black Pig and Duroc × Duroc × Berkshire × Jiaxing Black Pig Crosses. Gene 2022, 808, 145978. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Thakali, K.; Morse, P.; Shelby, S.; Chen, J.; Apple, J.; Huang, Y. Comparison of Growth Performance and Meat Quality Traits of Commercial Cross-Bred Pigs versus the Large Black Pig Breed. Animals 2021, 11, 200. [Google Scholar] [CrossRef] [PubMed]
- Khanal, P.; Maltecca, C.; Schwab, C.; Gray, K.; Tiezzi, F. Genetic Parameters of Meat Quality, Carcass Composition, and Growth Traits in Commercial Swine. J. Anim. Sci. 2019, 97, 3669–3683. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Barroso, M.Á.; Silió, L.; Rodríguez, C.; Palma-Granados, P.; López, A.; Caraballo, C.; Sánchez-Esquiliche, F.; Gómez-Carballar, F.; García-Casco, J.M.; Muñoz, M. Genetic Parameter Estimation and Gene Association Analyses for Meat Quality Traits in Open-air Free-range Iberian Pigs. J. Anim. Breed. Genet. 2020, 137, 581–598. [Google Scholar] [CrossRef]
- Huang, Y.; Zhou, L.; Zhang, J.; Liu, X.; Zhang, Y.; Cai, L.; Zhang, W.; Cui, L.; Yang, J.; Ji, J.; et al. A Large-Scale Comparison of Meat Quality and Intramuscular Fatty Acid Composition among Three Chinese Indigenous Pig Breeds. Meat Sci. 2020, 168, 108182. [Google Scholar] [CrossRef]
- Jiang, Y.Z.; Zhu, L.; Tang, G.Q.; Li, M.Z.; Jiang, A.A.; Cen, W.M.; Xing, S.H.; Chen, J.N.; Wen, A.X.; He, T.; et al. Carcass and Meat Quality Traits of Four Commercial Pig Crossbreeds in China. Genet. Mol. Res. 2012, 11, 4447–4455. [Google Scholar] [CrossRef]
- Jiang, Y.-Z.; Zhu, L.; Li, F.-Q.; Li, X.-W. Carcass Composition and Meat Quality of Indigenous Yanan Pigs of China. Genet. Mol. Res. 2012, 11, 166–173. [Google Scholar] [CrossRef]
- Adzitey, F.; Nurul, H. Pale Soft Exudative (PSE) and Dark Firm Dry (DFD) Meats: Causes and Measures to Reduce These Incidences: A Mini Review. Int. Food Res. J. 2011, 18, 11–20. [Google Scholar]
- Hamilton, D.N.; Ellis, M.; Miller, K.D.; McKeith, F.K.; Parrett, D.F. The Effect of the Halothane and Rendement Napole Genes on Carcass and Meat Quality Characteristics of Pigs. J. Anim. Sci. 2000, 78, 2862. [Google Scholar] [CrossRef] [Green Version]
- Scheffler, T.L.; Gerrard, D.E. Mechanisms Controlling Pork Quality Development: The Biochemistry Controlling Postmortem Energy Metabolism. Meat Sci. 2007, 77, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Timpson, N.J.; Greenwood, C.M.T.; Soranzo, N.; Lawson, D.J.; Richards, J.B. Genetic Architecture: The Shape of the Genetic Contribution to Human Traits and Disease. Nat. Rev. Genet. 2018, 19, 110–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, T.F.C. The Genetic Architecture of Quantitative Traits. Annu. Rev. Genet. 2001, 35, 303–339. [Google Scholar]
- Wang, H.; Wang, X.; Yan, D.; Sun, H.; Chen, Q.; Li, M.; Dong, X.; Pan, Y.; Lu, S. Genome-Wide Association Study Identifying Genetic Variants Associated with Carcass Backfat Thickness, Lean Percentage and Fat Percentage in a Four-Way Crossbred Pig Population Using SLAF-Seq Technology. BMC Genom. 2022, 23, 594. [Google Scholar] [CrossRef]
- Ding, R.; Zhuang, Z.; Qiu, Y.; Ruan, D.; Wu, J.; Ye, J.; Cao, L.; Zhou, S.; Zheng, E.; Huang, W.; et al. Identify Known and Novel Candidate Genes Associated with Backfat Thickness in Duroc Pigs by Large-Scale Genome-Wide Association Analysis. J. Anim. Sci. 2022, 100, skac012. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.-C.; Yoo, C.-K.; Lee, J.-B.; Jung, E.-J.; Han, S.-H.; Lee, S.-S.; Ko, M.-S.; Lim, H.-T.; Park, H.-B. Genome-Wide QTL Analysis of Meat Quality-Related Traits in a Large F2 Intercross between Landrace and Korean Native Pigs. Genet. Sel. Evol. 2015, 47, 7. [Google Scholar] [CrossRef] [Green Version]
- Suwannasing, R.; Duangjinda, M.; Boonkum, W.; Taharnklaew, R.; Tuangsithtanon, K. The Identification of Novel Regions for Reproduction Trait in Landrace and Large White Pigs Using a Single Step Genome-Wide Association Study. Asian-Australas. J. Anim. Sci. 2018, 31, 1852–1862. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ding, X.; Tan, Z.; Xing, K.; Yang, T.; Pan, Y.; Wang, Y.; Mi, S.; Sun, D.; Wang, C. Genome-Wide Association Study for Reproductive Traits in a Large White Pig Population. Anim. Genet. 2018, 49, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Browning, B.L.; Zhou, Y.; Browning, S.R. A One-Penny Imputed Genome from Next-Generation Reference Panels. Am. J. Hum. Genet. 2018, 103, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Zhang, H.; Tang, Z.; Yin, D.; Fu, Y.; Yuan, X.; Li, X.; Liu, X.; Zhao, S. HIBLUP: An Integration of Statistical Models on the BLUP Framework for Efficient Genetic Evaluation Using Big Genomic Data. Nucleic Acids Res. 2023, 51, gkad074. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, H.; Tang, Z.; Xu, J.; Yin, D.; Zhang, Z.; Yuan, X.; Zhu, M.; Zhao, S.; Li, X.; et al. rMVP: A Memory-Efficient, Visualization-Enhanced, and Parallel-Accelerated Tool for Genome-Wide Association Study. Genom. Proteom. Bioinform. 2021, 19, 619–628. [Google Scholar] [CrossRef]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Vroh Bi, I.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A Unified Mixed-Model Method for Association Mapping That Accounts for Multiple Levels of Relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef]
- Price, A.L.; Patterson, N.J.; Plenge, R.M.; Weinblatt, M.E.; Shadick, N.A.; Reich, D. Principal Components Analysis Corrects for Stratification in Genome-Wide Association Studies. Nat. Genet. 2006, 38, 904–909. [Google Scholar] [CrossRef]
- VanRaden, P.M. Efficient Methods to Compute Genomic Predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teslovich, T.M.; Musunuru, K.; Smith, A.V.; Edmondson, A.C.; Stylianou, I.M.; Koseki, M.; Pirruccello, J.P.; Ripatti, S.; Chasman, D.I.; Willer, C.J.; et al. Biological, Clinical and Population Relevance of 95 Loci for Blood Lipids. Nature 2010, 466, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.-S.; He, W.-M.; Ji, J.-J.; Zhang, C.; Guo, Y.; Yang, T.-L. LDBlockShow: A Fast and Convenient Tool for Visualizing Linkage Disequilibrium and Haplotype Blocks Based on Variant Call Format Files. Brief. Bioinform. 2021, 22, bbaa227. [Google Scholar] [CrossRef]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent Prioritization and Exploratory Visualization of Biological Functions for Gene Enrichment Analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef] [PubMed]
- Gjerlaug-Enger, E.; Aass, L.; Ødegård, J.; Vangen, O. Genetic Parameters of Meat Quality Traits in Two Pig Breeds Measured by Rapid Methods. Animal 2010, 4, 1832–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonesson, A.K.; de Greef, K.H.; Meuwissen, T.H.E. Genetic Parameters and Trends of Meat Quality, Carcass Composition and Performance Traits in Two Selected Lines of Large White Pigs. Livest. Prod. Sci. 1998, 57, 23–32. [Google Scholar] [CrossRef]
- Lee, J.-H.; Song, K.-D.; Lee, H.-K.; Cho, K.-H.; Park, H.-C.; Park, K.-D. Genetic Parameters of Reproductive and Meat Quality Traits in Korean Berkshire Pigs. Asian Australas. J. Anim. Sci. 2015, 28, 1388–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filha, W.S.A.; Bernardi, M.L.; Wentz, I.; Bortolozzo, F.P. Reproductive Performance of Gilts According to Growth Rate and Backfat Thickness at Mating. Anim. Reprod. Sci. 2010, 121, 139–144. [Google Scholar] [CrossRef]
- Gozalo-Marcilla, M.; Buntjer, J.; Johnsson, M.; Batista, L.; Diez, F.; Werner, C.R.; Chen, C.-Y.; Gorjanc, G.; Mellanby, R.J.; Hickey, J.M.; et al. Genetic Architecture and Major Genes for Backfat Thickness in Pig Lines of Diverse Genetic Backgrounds. Genet. Sel. Evol. 2021, 53, 76. [Google Scholar] [CrossRef]
- Song, B.; Zheng, C.; Zheng, J.; Zhang, S.; Zhong, Y.; Guo, Q.; Li, F.; Long, C.; Xu, K.; Duan, Y.; et al. Comparisons of Carcass Traits, Meat Quality, and Serum Metabolome between Shaziling and Yorkshire Pigs. Anim. Nutr. 2022, 8, 125–134. [Google Scholar] [CrossRef]
- Suzuki, K.; Irie, M.; Kadowaki, H.; Shibata, T.; Kumagai, M.; Nishida, A. Genetic Parameter Estimates of Meat Quality Traits in Duroc Pigs Selected for Average Daily Gain, Longissimus Muscle Area, Backfat Thickness, and Intramuscular Fat Content. J. Anim. Sci. 2005, 83, 2058–2065. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.; Lan, G.; Zhang, L.; Wang, L.; Liang, J.; Liu, X. Detection of DKK3 and CCR1 Genes Polymorphisms and Their Association with Backfat Thickness in Being Black Pigs. Acta Vet. Zootech. Sin. 2022, 53, 2083–2093. [Google Scholar]
- Kim, G.-D.; Jeong, J.-Y.; Hur, S.-J.; Yang, H.-S.; Jeon, J.-T.; Joo, S.-T. The Relationship between Meat Color (CIE L* and A*), Myoglobin Content, and Their Influence on Muscle Fiber Characteristics and Pork Quality. Korean J. Food Sci. Anim. Resour. 2010, 30, 626–633. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Hou, L.; Zhou, W.; Wang, B.; Han, P.; Gao, C.; Niu, P.; Zhang, Z.; Li, Q.; Huang, R.; et al. Genome-Wide Association Study and FST Analysis Reveal Four Quantitative Trait Loci and Six Candidate Genes for Meat Color in Pigs. Front. Genet. 2022, 13, 768710. [Google Scholar] [CrossRef]
- Tomasevic, I.; Djekic, I.; Font-i-Furnols, M.; Terjung, N.; Lorenzo, J.M. Recent Advances in Meat Color Research. Curr. Opin. Food Sci. 2021, 41, 81–87. [Google Scholar] [CrossRef]
- Liao, Y.; Gao, H.; Zhang, Y.; Yin, S.; Xu, K.; He, J. Genome-Wide Association Analysis of Post-Mortem pH and Meat Color Traits in Ningxiang Pigs. Chin. J. Anim. Sci. 2021, 57, 174–181. [Google Scholar]
- Yuan, Z.; Li, J.; Li, J.; Gao, X.; Gao, H.; Xu, S. Effects of DGAT1 Gene on Meat and Carcass Fatness Quality in Chinese Commercial Cattle. Mol. Biol. Rep. 2013, 40, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Stringer, S.; Frei, O.; Umićević Mirkov, M.; de Leeuw, C.; Polderman, T.J.C.; van der Sluis, S.; Andreassen, O.A.; Neale, B.M.; Posthuma, D. A Global Overview of Pleiotropy and Genetic Architecture in Complex Traits. Nat. Genet. 2019, 51, 1339–1348. [Google Scholar] [CrossRef]
- Rohrer, G.A.; Keele, J.W. Identification of Quantitative Trait Loci Affecting Carcass Composition in Swine: II. Muscling and Wholesale Product Yield Traits. J. Anim. Sci. 1998, 76, 2255–2262. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Jennen, D.G.J.; Tholen, E.; Juengst, H.; Kleinwächter, T.; Hölker, M.; Tesfaye, D.; Un, G.; Schreinemachers, H.-J.; Murani, E.; et al. A Genome Scan Reveals QTL for Growth, Fatness, Leanness and Meat Quality in a Duroc-Pietrain Resource Population. Anim. Genet. 2007, 38, 241–252. [Google Scholar] [CrossRef]
- Nagamine, Y.; Visscher, P.M.; Haley, C.S. QTL Detection and Allelic Effects for Growth and Fat Traits in Outbred Pig Populations. Genet. Sel. Evol. 2004, 36, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Demeure, O.; Sanchez, M.P.; Riquet, J.; Iannuccelli, N.; Demars, J.; Fève, K.; Kernaleguen, L.; Gogué, J.; Billon, Y.; Caritez, J.C.; et al. Exclusion of the Swine Leukocyte Antigens as Candidate Region and Reduction of the Position Interval for the Sus Scrofa Chromosome 7 QTL Affecting Growth and Fatness. J. Anim. Sci. 2005, 83, 1979–1987. [Google Scholar] [CrossRef] [Green Version]
- Stratz, P.; Baes, C.; Rückert, C.; Preuss, S.; Bennewitz, J. A Two-Step Approach to Map Quantitative Trait Loci for Meat Quality in Connected Porcine F(2) Crosses Considering Main and Epistatic Effects. Anim. Genet. 2013, 44, 14–23. [Google Scholar] [CrossRef]
- Stansley, B.J.; Conn, P.J. Neuropharmacological Insight from Allosteric Modulation of mGlu Receptors. Trends Pharmacol. Sci. 2019, 40, 240–252. [Google Scholar] [CrossRef]
- Pang, Y.; Zhao, J.; Fowdur, M.; Liu, Y.; Wu, H.; He, M. To Explore the Mechanism of the GRM4 Gene in Osteosarcoma by RNA Sequencing and Bioinformatics Approach. Med. Sci. Monit. Basic Res. 2018, 24, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Wan, Z.; Sun, R.; Liu, Y.-W.; Li, S.; Sun, J.; Li, J.; Zhu, J.; Moharil, P.; Zhang, B.; Ren, P.; et al. Targeting Metabotropic Glutamate Receptor 4 for Cancer Immunotherapy. Sci. Adv. 2021, 7, eabj4226. [Google Scholar] [CrossRef]
- Xiao, B.; Chen, D.; Zhou, Q.; Hang, J.; Zhang, W.; Kuang, Z.; Sun, Z.; Li, L. Glutamate Metabotropic Receptor 4 (GRM4) Inhibits Cell Proliferation, Migration and Invasion in Breast Cancer and Is Regulated by miR-328-3p and miR-370-3p. BMC Cancer 2019, 19, 891. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Li, N.; Wei, X.; Chen, B.; Zhang, Y.; Zhao, Y.; Hu, X.; Hou, S. GRM4 Inhibits the Proliferation, Migration, and Invasion of Human Osteosarcoma Cells through Interaction with CBX4. Biosci. Biotechnol. Biochem. 2020, 84, 279–289. [Google Scholar] [CrossRef]
- Kansara, M.; Teng, M.W.; Smyth, M.J.; Thomas, D.M. Translational Biology of Osteosarcoma. Nat. Rev. Cancer 2014, 14, 722–735. [Google Scholar] [CrossRef]
- Kansara, M.; Thomson, K.; Pang, P.; Dutour, A.; Mirabello, L.; Acher, F.; Pin, J.-P.; Demicco, E.G.; Yan, J.; Teng, M.W.L.; et al. Infiltrating Myeloid Cells Drive Osteosarcoma Progression via GRM4 Regulation of IL23. Cancer Discov. 2019, 9, 1511–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Wang, X.; Shan, L.; Zhou, J.; Zhang, X.; Zhu, E.; Yuan, H.; Wang, B. High-Mobility Group At-Hook 1 Mediates the Role of Nuclear Factor I/X in Osteogenic Differentiation Through Activating Canonical Wnt Signaling. Stem Cells 2021, 39, 1349–1361. [Google Scholar] [CrossRef] [PubMed]
- Chiefari, E.; Tanyolaç, S.; Iiritano, S.; Sciacqua, A.; Capula, C.; Arcidiacono, B.; Nocera, A.; Possidente, K.; Baudi, F.; Ventura, V.; et al. A Polymorphism of HMGA1 Is Associated with Increased Risk of Metabolic Syndrome and Related Components. Sci. Rep. 2013, 3, 1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.S.; Lee, J.J.; Shin, H.Y.; Choi, B.H.; Lee, C.K.; Kim, J.J.; Cho, B.W.; Kim, T.-H. Association of Melanocortin 4 Receptor (MC4R) and High Mobility Group AT-Hook 1 (HMGA1) Polymorphisms with Pig Growth and Fat Deposition Traits. Anim. Genet. 2006, 37, 419–421. [Google Scholar] [CrossRef]
- Arce-Cerezo, A.; García, M.; Rodríguez-Nuevo, A.; Crosa-Bonell, M.; Enguix, N.; Peró, A.; Muñoz, S.; Roca, C.; Ramos, D.; Franckhauser, S.; et al. HMGA1 Overexpression in Adipose Tissue Impairs Adipogenesis and Prevents Diet-Induced Obesity and Insulin Resistance. Sci. Rep. 2015, 5, 14487. [Google Scholar] [CrossRef] [Green Version]
- Foti, D.; Chiefari, E.; Fedele, M.; Iuliano, R.; Brunetti, L.; Paonessa, F.; Manfioletti, G.; Barbetti, F.; Brunetti, A.; Croce, C.M.; et al. Lack of the Architectural Factor HMGA1 Causes Insulin Resistance and Diabetes in Humans and Mice. Nat. Med. 2005, 11, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Xiao, S.; Li, W.; Huang, T.; Huang, X.; Yan, G.; Huang, Y.; Qiu, H.; Jiang, K.; Wang, X.; et al. Unravelling the Genetic Loci for Growth and Carcass Traits in Chinese Bamaxiang Pigs Based on a 1.4 Million SNP Array. J. Anim. Breed. Genet. 2019, 136, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Song, H.; Jiang, Y.; Jiang, Y.; Zhang, F.; Liu, Y.; Shi, Y.; Ding, X.; Wang, C. A Single-Step Genome Wide Association Study on Body Size Traits Using Imputation-Based Whole-Genome Sequence Data in Yorkshire Pigs. Front. Genet. 2021, 12, 629049. [Google Scholar] [CrossRef]
- Otto, G.; Roehe, R.; Looft, H.; Thoelking, L.; Knap, P.W.; Rothschild, M.F.; Plastow, G.S.; Kalm, E. Associations of DNA Markers with Meat Quality Traits in Pigs with Emphasis on Drip Loss. Meat Sci. 2007, 75, 185–195. [Google Scholar] [CrossRef]
- Zhu, X.; Qian, Y.; Li, X.; Xu, Z.; Xia, R.; Wang, N.; Liang, J.; Yin, H.; Zhang, A.; Guo, C.; et al. Structural Basis of Adhesion GPCR GPR110 Activation by Stalk Peptide and G-Proteins Coupling. Nat. Commun. 2022, 13, 5513. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, S.; Mouri, Y.; Akiyama, M.; Miyasaka, N.; Nakahama, K. GPR110, a Receptor for Synaptamide, Expressed in Osteoclasts Negatively Regulates Osteoclastogenesis. Prostaglandins Leukot. Essent. Fat. Acids 2022, 182, 102457. [Google Scholar] [CrossRef]
- Falker-Gieske, C.; Blaj, I.; Preuß, S.; Bennewitz, J.; Thaller, G.; Tetens, J. GWAS for Meat and Carcass Traits Using Imputed Sequence Level Genotypes in Pooled F2-Designs in Pigs. G3 Genes|Genomes|Genet. 2019, 9, 2823–2834. [Google Scholar] [CrossRef] [Green Version]
- Yuan, B.; Liao, F.; Shi, Z.-Z.; Ren, Y.; Deng, X.-L.; Yang, T.-T.; Li, D.-Y.; Li, R.-F.; Pu, D.-D.; Wang, Y.-J.; et al. Dihydroartemisinin Inhibits the Proliferation, Colony Formation and Induces Ferroptosis of Lung Cancer Cells by Inhibiting PRIM2/SLC7A11 Axis. OTT 2020, 13, 10829–10840. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, L.; Yan, H.; Liu, X.; Li, N.; Liang, J.; Pu, L.; Zhang, Y.; Shi, H.; Zhao, K.; et al. Genome-Wide Association Studies Identify the Loci for 5 Exterior Traits in a Large White × Minzhu Pig Population. PLoS ONE 2014, 9, e103766. [Google Scholar] [CrossRef]
- Lu, B.; Jiao, Y.; Wang, Y.; Dong, J.; Wei, M.; Cui, B.; Sun, Y.; Wang, L.; Zhang, B.; Chen, Z.; et al. A FKBP5 Mutation Is Associated with Paget’s Disease of Bone and Enhances Osteoclastogenesis. Exp. Mol. Med. 2017, 49, e336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmady, E.; Deeke, S.A.; Rabaa, S.; Kouri, L.; Kenney, L.; Stewart, A.F.R.; Burgon, P.G. Identification of a Novel Muscle A-Type Lamin-Interacting Protein (MLIP). J. Biol. Chem. 2011, 286, 19702–19713. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Zhang, M.; Yan, G.; Huang, X.; Chen, H.; Zhou, L.; Deng, W.; Zhang, Z.; Qiu, H.; Ai, H.; et al. Genome-Wide Association and Evolutionary Analyses Reveal the Formation of Swine Facial Wrinkles in Chinese Erhualian Pigs. Aging 2019, 11, 4672–4687. [Google Scholar] [CrossRef]
- Tong, X.; Li, G.; Feng, Y. TINAG Mutation as a Genetic Cause of Pectus Excavatum. Med. Hypotheses 2020, 137, 109557. [Google Scholar] [CrossRef] [PubMed]
- Heinz, L.X.; Rebsamen, M.; Rossi, D.C.; Staehli, F.; Schroder, K.; Quadroni, M.; Gross, O.; Schneider, P.; Tschopp, J. The Death Domain-Containing Protein Unc5CL Is a Novel MyD88-Independent Activator of the pro-Inflammatory IRAK Signaling Cascade. Cell Death Differ. 2012, 19, 722–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Hu, Z.; He, Z.; Jia, W.; Wang, F.; Zhou, Y.; Liu, Z.; Zhan, Q.; Liu, Y.; Yu, D.; et al. Genome-Wide Association Study Identifies Three New Susceptibility Loci for Esophageal Squamous-Cell Carcinoma in Chinese Populations. Nat. Genet. 2011, 43, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Verma, S.; Bhat, A.; Rasool Bhat, G.; Sharma, V.; Sharma, I.; Singh, H.; Kaul, S.; Rai, E.; Sharma, S. Newly Identified Genetic Variant Rs2294693 in UNC5CL Gene Is Associated with Decreased Risk of Esophageal Carcinoma in the J&K Population–India. BIOCELL 2021, 45, 665–670. [Google Scholar] [CrossRef]
- Alonso-Gil, D.; Cuadrado, A.; Giménez-Llorente, D.; Rodríguez-Corsino, M.; Losada, A. Different NIPBL Requirements of Cohesin-STAG1 and Cohesin-STAG2. Nat. Commun. 2023, 14, 1326. [Google Scholar] [CrossRef]
- Li, N. SNPS Detection of PSMC3 Gene and the Relationship with Body Measutement and Carcass Traits in Qichuan Cattle. Master’s Thesis, Northwest A&F University, Yangling, China, 2015. [Google Scholar]
- Jiang, L.; Liu, X.; Yang, J.; Wang, H.; Jiang, J.; Liu, L.; He, S.; Ding, X.; Liu, J.; Zhang, Q. Targeted Resequencing of GWAS Loci Reveals Novel Genetic Variants for Milk Production Traits. BMC Genom. 2014, 15, 1105. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.-S.; Ros-Freixedes, R.; Pena, R.N.; Baas, T.J.; Estany, J.; Rothschild, M.F. Identification of Signatures of Selection for Intramuscular Fat and Backfat Thickness in Two Duroc Populations1. J. Anim. Sci. 2015, 93, 3292–3302. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.M.; Clarke, F.M.; Purslow, P.P.; Warner, R.D. Meat Color Is Determined Not Only by Chromatic Heme Pigments but Also by the Physical Structure and Achromatic Light Scattering Properties of the Muscle. Compr. Rev. Food Sci. Food Saf. 2020, 19, 44–63. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | n | Max. | Min. | Mean ± SD | C.V. |
|---|---|---|---|---|---|
| CL (cm) | 508 | 96.40 | 68.50 | 81.35 ± 4.69 | 5.77 |
| COL (cm) | 508 | 86.50 | 34.10 | 66.11 ± 6.16 | 9.32 |
| BFT (mm) | 485 | 71.06 | 16.17 | 41.61 ± 8.28 | 19.90 |
| L.LD | 508 | 58.73 | 34.80 | 44.73 ± 3.84 | 8.58 |
| a.LD | 508 | 16.17 | 1.34 | 6.53 ± 2.61 | 39.97 |
| b.LD | 508 | 10.53 | 0.14 | 4.00 ± 1.67 | 41.75 |
| pH45min | 508 | 6.96 | 5.46 | 6.28 ± 0.31 | 4.94 |
| pH24h | 508 | 6.87 | 5.46 | 5.91 ± 0.28 | 4.74 |
| Trait | CL | COL | BFT | pH45min | pH24h | L.LD | a.LD | b.LD |
|---|---|---|---|---|---|---|---|---|
| CL | 0.80 (0.06) | 0.87 | −0.53 | −0.22 | −0.27 | 0.25 | −0.05 | 0.14 |
| COL | 0.82 *** | 0.47 (0.07) | −0.53 | 0.08 | −0.47 | 0.07 | −0.41 | 0.50 |
| BFT | −0.12 ** | −0.16 *** | 0.48 (0.08) | 0.07 | −0.08 | −0.32 | 0.07 | −0.32 |
| pH45min | 0.01 | 0.02 | −0.04 | 0.14 (0.11) | 0.10 | 0.27 | −0.39 | 0.41 |
| pH24h | −0.05 | −0.09 | 0.12 | 0.37 *** | 0.30 (0.09) | 0.45 | 0.42 | −0.37 |
| L.LD | 0.06 | −0.03 | −0.07 | −0.24 *** | −0.2 *** | 0.11 (0.07) | 0.38 | −0.08 |
| a.LD | −0.11 * | −0.28 *** | 0.18 *** | −0.07 | −0.04 | 0.31 *** | 0.44 (0.08) | −0.23 |
| b.LD | 0.27 *** | 0.32 *** | −0.18 *** | 0.09 | −0.24 *** | 0.24 *** | 0.03 | 0.19 (0.09) |
| (a) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Trait | SNP (Rsid) | CHR | POS (bp) | Consequence | MAF | PVE (%) | P-adj | Nearest Gene | DIS (bp) |
| CL | ALGA0040227 (rs80983858) | 7 | 30,176,520 | Downstream gene variant | 0.39 | 14.35 | 8.05 × 10−19 | GRM4 | 2785 |
| ALGA0040238 (rs80815545) | 7 | 30,197,014 | Intron variant | 0.36 | 12.98 | 4.72 × 10−11 | GRM4 | Within | |
| INRA0024788 (–––) | 7 | 30,31,7219 | ––– | 0.36 | 10.08 | 2.16 × 10−13 | HMGA1 | 3191 | |
| ALGA0039917 (rs81397589) | 7 | 26,737,102 | Intron variant | 0.19 | 7.02 | 1.25 × 10−9 | MLIP | Within | |
| ALGA0040777 (rs80845178) | 7 | 36,323,988 | Intergenic variant | 0.44 | 6.28 | 9.85 × 10−9 | UNC5CL | 8213 | |
| ALGA0040243 (rs80942143) | 7 | 30,213,771 | Intron variant | 0.25 | 5.69 | 4.97 × 10−8 | GRM4 | Within | |
| WU_10.2_7_48537179 (–––) | 7 | 41,877,149 | ––– | 0.42 | 5.66 | 5.39 × 10−8 | ADGRF1 | 23,492 | |
| ASGA0032589 (rs80869188) | 7 | 31,450,019 | Intron variant | 0.32 | 5.13 | 2.36 × 10−7 | FKBP5 | Within | |
| H3GA0020641 (rs80975871) | 7 | 28,521,421 | Intron variant | 0.11 | 4.92 | 4.17 × 10−7 | PRIM2 | Within | |
| ALGA0039880 (rs80928470) | 7 | 26,501,975 | Intron variant | 0.11 | 4.86 | 5.04 × 10−7 | TINAG | Within | |
| ALGA0041948 (rs80997002) | 7 | 50,283,279 | Intergenic variant | 0.47 | 4.77 | 6.30 × 10−7 | TMC3 | 99,190 | |
| ALGA0040370 (rs81397836) | 7 | 32,328,188 | Intergenic variant | 0.48 | 4.60 | 1.02 × 10−6 | SRSF3 | 29,608 | |
| M1GA0010006 (rs80946246) | 7 | 31,161,760 | Intron variant | 0.31 | 4.55 | 1.16 × 10−6 | ZNF76 | Within | |
| WU_10.2_7_36255497 (–––) | 7 | 31,181,718 | –––– | 0.31 | 4.55 | 1.16 × 10−6 | ZNF76 | Within | |
| MARC0060950 (rs80924014) | 7 | 46,569,153 | Upstream gene variant | 0.16 | 4.44 | 1.58 × 10−6 | TMEM14A | 51,421 | |
| COL | ALGA0040227 (rs80983858) | 7 | 30,176,520 | Downstream gene variant | 0.39 | 8.38 | 2.67 × 10−25 | GRM4 | 2785 |
| ALGA0040238 (rs80815545) | 7 | 30,197,014 | Intron variant | 0.36 | 7.51 | 3.17 × 10−10 | GRM4 | Within | |
| ALGA0039880 (rs80928470) | 7 | 26,501,975 | Intron variant | 0.11 | 5.75 | 4.19 × 10−7 | TINAG | Within | |
| H3GA0020641 (rs80975871) | 7 | 28,521,421 | Intron variant | 0.11 | 5.14 | 2.30 × 10−7 | PRIM2 | Within | |
| ALGA0039917 (rs81397589) | 7 | 26,737,102 | Intron variant | 0.19 | 4.88 | 4.74 × 10−7 | MLIP | Within | |
| INRA0024788 | 7 | 30,317,219 | ––– | 0.36 | 4.63 | 9.33 × 10−7 | HMGA1 | 3191 | |
| BFT | WU_10.2_18_56654365 (–––) | 18 | 51,759,775 | ––– | 0.12 | 12.66 | 5.80 × 10−16 | HECW1 | Within |
| WU_10.2_16_23509998 (–––) | 16 | 22,361,911 | ––– | 0.12 | 12.38 | 1.27 × 10−15 | NIPBL | Within | |
| WU_10.2_8_138925750 (–––) | 8 | 129,537,879 | ––– | 0.12 | 11.94 | 4.37 × 10−15 | SNCA | 266,751 | |
| ALGA0014052 (rs81360052) | 2 | 82,412,427 | Intron variant, noncoding transcript variant | 0.14 | 7.72 | 4.52 × 10−10 | TMEM174 | 75,272 | |
| ALGA0040227 (rs80983858) | 7 | 30,176,520 | Downstream gene variant | 0.39 | 5.01 | 6.14 × 10−7 | GRM4 | 2785 | |
| (b) | |||||||||
| Trait | SNP | CHR | POS (bp) | Consequence | MAF | PVE (%) | P-adj | Nearest Gene | DIS (bp) |
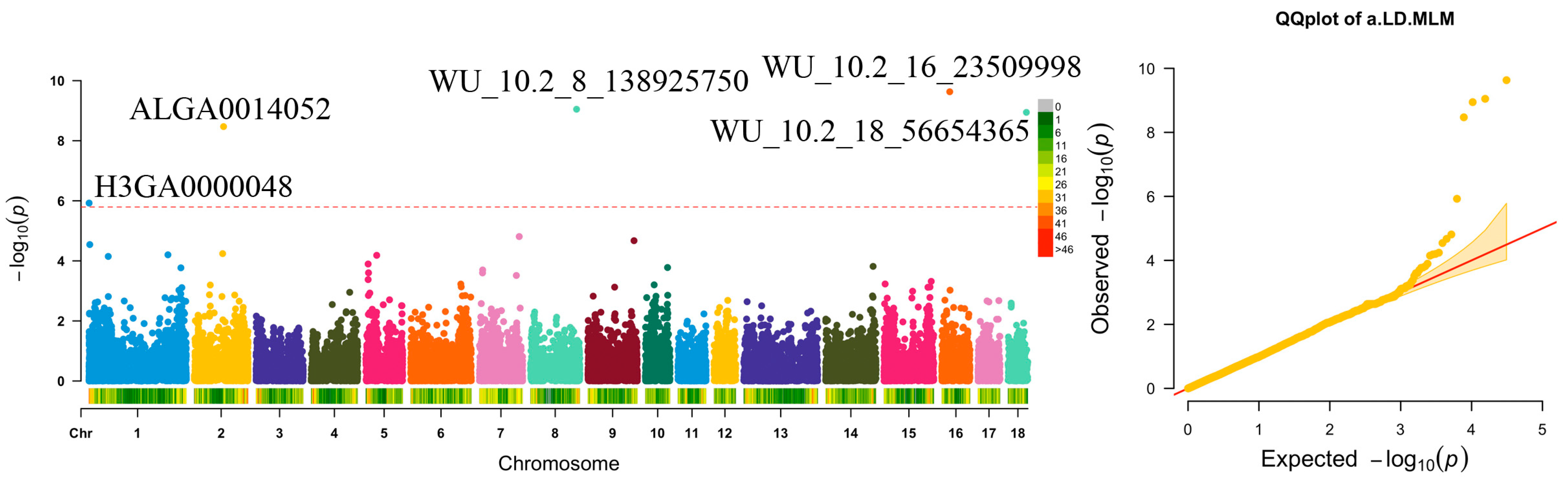
| a.LD | WU_10.2_16_23509998 (–––) | 16 | 22,361,911 | ––– | 0.12 | 7.94 | 2.34 × 10−10 | NIPBL | Within |
| WU_10.2_8_138925750 (–––) | 8 | 129,537,879 | ––– | 0.12 | 7.44 | 8.95 × 10−10 | SNCA | 266,751 | |
| WU_10.2_18_56654365 (–––) | 18 | 51,759,775 | ––– | 0.12 | 7.36 | 1.14 × 10−9 | HECW1 | Within | |
| ALGA0014052 (rs81360052) | 2 | 82,412,427 | Intron variant, noncoding transcript variant | 0.14 | 6.95 | 3.38 × 10−9 | TMEM174 | 75,272 | |
| H3GA0000048 (rs80803041) | 1 | 493,510 | Intergenic variant | 0.01 | 4.75 | 1.19 × 10−6 | ERMARD | 19,168 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, S.; Song, G.; Gao, N.; Gao, H.; Zeng, Q.; Lu, P.; Zhang, Q.; Xu, K.; He, J. Identifying Genetic Architecture of Carcass and Meat Quality Traits in a Ningxiang Indigenous Pig Population. Genes 2023, 14, 1308. https://doi.org/10.3390/genes14071308
Yin S, Song G, Gao N, Gao H, Zeng Q, Lu P, Zhang Q, Xu K, He J. Identifying Genetic Architecture of Carcass and Meat Quality Traits in a Ningxiang Indigenous Pig Population. Genes. 2023; 14(7):1308. https://doi.org/10.3390/genes14071308
Chicago/Turabian StyleYin, Shishu, Gang Song, Ning Gao, Hu Gao, Qinghua Zeng, Peng Lu, Qin Zhang, Kang Xu, and Jun He. 2023. "Identifying Genetic Architecture of Carcass and Meat Quality Traits in a Ningxiang Indigenous Pig Population" Genes 14, no. 7: 1308. https://doi.org/10.3390/genes14071308