
Transcription of WNT Genes in Hematopoietic Niche’s Mesenchymal Stem Cells in Multiple Myeloma Patients with Different Responses to Treatment
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioethics
2.2. Patients
2.3. Cell Cultures and Lines
2.4. Oligonucleotide Sequences: Probes and Primers
2.5. RNA Isolation and cDNA Preparation
2.6. Real-Time PCR
2.7. Statistical Analysis
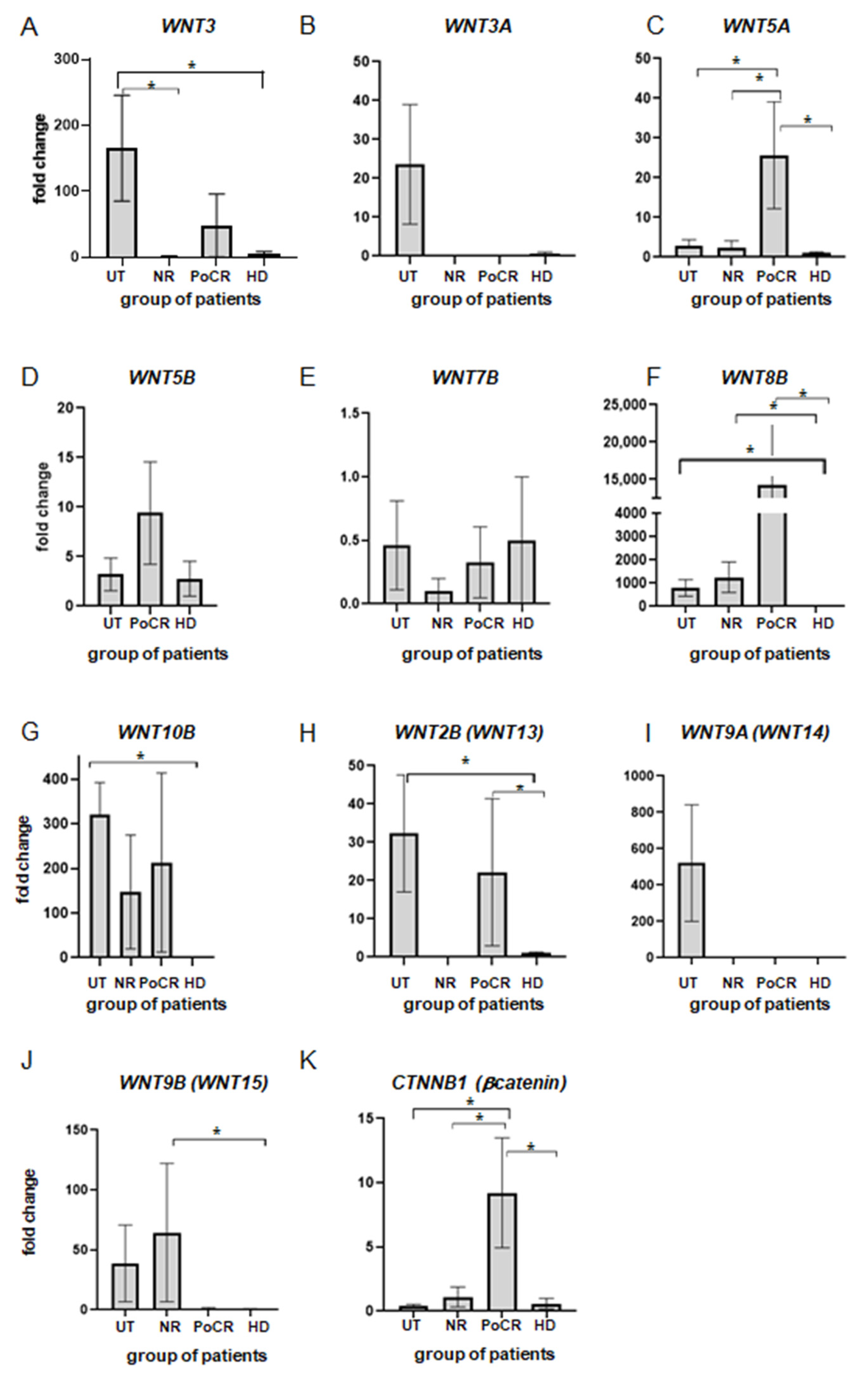
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albagoush, S.A.; Shumway, C.; Azevedo, A.M. Multiple Myeloma. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK534764/ (accessed on 26 December 2022).
- Bessmeltsev, S.S.; Abdulkadyrov, K.M. Multiple Myeloma; A Guide for Physicians; MK: Moscow, Russia, 2016. [Google Scholar]
- Baryawno, N.; Przybylski, D.; Kowalczyk, M.S.; Kfoury, Y.; Severe, N.; Gustafsson, K.; Kokkaliaris, K.D.; Mercier, F.; Tabaka, M.; Hofree, M.; et al. A Cellular Taxonomy of the Bone Marrow Stroma in Homeostasis and Leukemia. Cell 2019, 177, 1915–1932.e16. [Google Scholar] [CrossRef] [PubMed]
- Schofield, R. The Relationship between the Spleen Colony-Forming Cell and the Haemopoietic Stem Cell. Blood Cells 1978, 4, 7–25. [Google Scholar] [PubMed]
- David, T. Scadden the Stem-Cell Niche as an Entity of Action. Nature 2006, 441, 1075–1079. [Google Scholar]
- Semenova, N.Y.; Bessmel’tsev, S.S.; Rugal’, V.I. Biology of Hematopoietic Stem Cell Niche. Clin. Oncohematol. 2014, 7, 501–510. [Google Scholar]
- Dazzi, F.; Ramasamy, R.; Glennie, S.; Jones, S.P.; Roberts, I. The Role of Mesenchymal Stem Cells in Haemopoiesis. Blood Rev. 2006, 20, 161–171. [Google Scholar] [CrossRef]
- Noll, J.E.; Williams, S.A.; Purton, L.E.; Zannettino, A.C.W. Tug of War in the Haematopoietic Stem Cell Niche: Do Myeloma Plasma Cells Compete for the HSC Niche? Blood Cancer J. 2012, 2, e91. [Google Scholar] [CrossRef]
- Shi, Y.; Du, L.; Lin, L.; Wang, Y. Tumour-Associated Mesenchymal Stem/Stromal Cells: Emerging Therapeutic Targets. Nat. Rev. Drug Discov. 2016, 16, 35–52. [Google Scholar] [CrossRef]
- Man, Y.; Yao, X.; Yang, T.; Wang, Y. Hematopoietic Stem Cell Niche During Homeostasis, Malignancy, and Bone Marrow Transplantation. Front. Cell Dev. Biol. 2021, 9, 621214. [Google Scholar] [CrossRef] [PubMed]
- Schulze, M.; Weisel, K.; Grandjean, C.; Oehrlein, K.; Zago, M.; Spira, D.; Horger, M. Increasing Bone Sclerosis during Bortezomib Therapy in Multiple Myeloma Patients: Results of a Reduced-Dose Whole-Body MDCT Study. Am. J. Roentgenol. 2014, 202, 170–179. [Google Scholar] [CrossRef]
- Enukashvily, N.I.; Semenova, N.; Chubar, A.V.; Ostromyshenskii, D.I.; Gushcha, E.A.; Gritsaev, S.; Bessmeltsev, S.S.; Rugal, V.I.; Prikhodko, E.M.; Kostroma, I.; et al. Pericentromeric Non-Coding DNA Transcription Is Associated with Niche Impairment in Patients with Ineffective or Partially Effective Multiple Myeloma Treatment. Int. J. Mol. Sci. 2022, 23, 3359. [Google Scholar] [CrossRef]
- Edwards, C.M.; Zhuang, J.; Mundy, G.R. The Pathogenesis of the Bone Disease of Multiple Myeloma. Bone 2008, 42, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Vallet, S.; Filzmoser, J.M.; Pecherstorfer, M.; Podar, K. Myeloma Bone Disease: Update on Pathogenesis and Novel Treatment Strategies. Pharmaceutics 2018, 10, 202. [Google Scholar] [CrossRef] [PubMed]
- Bessmeltsev, S.S. Multiple Myeloma (Pathogenesis, Clinical Features, Diagnosis, Differential Diagnosis). Part I Clin. Oncohematol. 2013, 6, 237–257. [Google Scholar]
- Albers, J.; Keller, J.; Baranowsky, A.; Beil, F.T.; Catala-Lehnen, P.; Schulze, J.; Amling, M.; Schinke, T. Canonical Wnt Signaling Inhibits Osteoclastogenesis Independent of Osteoprotegerin. J. Cell Biol. 2013, 200, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Glass, D.A.; Bialek, P.; Ahn, J.D.; Starbuck, M.; Patel, M.S.; Clevers, H.; Taketo, M.M.; Long, F.; McMahon, A.P.; Lang, R.A.; et al. Canonical Wnt Signaling in Differentiated Osteoblasts Controls Osteoclast Differentiation. Dev. Cell 2005, 8, 751–764. [Google Scholar] [CrossRef]
- Spencer, G.J.; Utting, J.C.; Etheridge, S.L.; Arnett, T.R.; Genever, P.G. Wnt Signalling in Osteoblasts Regulates Expression of the Receptor Activator of NFκB Ligand and Inhibits Osteoclastogenesis in Vitro. J. Cell Sci. 2006, 119, 1283–1296. [Google Scholar] [CrossRef]
- Ling, L.; Nurcombe, V.; Cool, S.M. Wnt Signaling Controls the Fate of Mesenchymal Stem Cells. Gene 2009, 433, 1–7. [Google Scholar] [CrossRef]
- Maeda, K.; Kobayashi, Y.; Udagawa, N.; Uehara, S.; Ishihara, A.; Mizoguchi, T.; Kikuchi, Y.; Takada, I.; Kato, S.; Kani, S.; et al. Wnt5a-Ror2 Signaling between Osteoblast-Lineage Cells and Osteoclast Precursors Enhances Osteoclastogenesis. Nat. Med. 2012, 18, 405–412. [Google Scholar] [CrossRef]
- Pederson, L.; Ruan, M.; Westendorf, J.J.; Khosla, S.; Oursler, M.J. Regulation of Bone Formation by Osteoclasts Involves Wnt/BMP Signaling and the Chemokine Sphingosine-1-Phosphate. Proc. Natl. Acad. Sci. USA 2008, 105, 20764–20769. [Google Scholar] [CrossRef]
- Qiang, Y.-W.; Chen, Y.; Stephens, O.; Brown, N.; Chen, B.; Epstein, J.; Barlogie, B.; Shaughnessy, J.D. Myeloma-Derived Dickkopf-1 Disrupts Wnt-Regulated Osteoprotegerin and RANKL Production by Osteoblasts: A Potential Mechanism Underlying Osteolytic Bone Lesions in Multiple Myeloma. Blood 2008, 112, 196–207. [Google Scholar] [CrossRef]
- Meads, M.B.; Gatenby, R.A.; Dalton, W.S. Environment-Mediated Drug Resistance: A Major Contributor to Minimal Residual Disease. Nat. Rev. Cancer 2009, 9, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Ivanovic, Z. Hypoxia or in Situ Normoxia: The Stem Cell Paradigm. J. Cell. Physiol. 2009, 219, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Nawata, M.; Wakitani, S. Expression Profiles and Functional Analyses of Wnt-Related Genes in Human Joint Disorders. Am. J. Pathol. 2005, 167, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Kobune, M.; Chiba, H.; Kato, J.; Kato, K.; Nakamura, K.; Kawano, Y.; Takada, K.; Takimoto, R.; Takayama, T.; Hamada, H.; et al. Wnt3/RhoA/ROCK Signaling Pathway Is Involved in Adhesion-Mediated Drug Resistance of Multiple Myeloma in an Autocrine Mechanism. Mol. Cancer Ther. 2007, 6, 1774–1784. [Google Scholar] [CrossRef] [PubMed]
- Mahtouk, K.; Moreaux, J.; Hose, D.; Rème, T.; Meissner, T.; Jourdan, M.; Rossi, J.F.; Pals, S.T.; Goldschmidt, H.; Klein, B. Growth Factors in Multiple Myeloma: A Comprehensive Analysis of Their Expression in Tumor Cells and Bone Marrow Environment Using Affymetrix Microarrays. BMC Cancer 2010, 10, 198. [Google Scholar] [CrossRef]
- Bolzoni, M.; Donofrio, G.; Storti, P.; Guasco, D.; Toscani, D.; Lazzaretti, M.; Bonomini, S.; Agnelli, L.; Capocefalo, A.; Dalla Palma, B.; et al. Myeloma Cells Inhibit Non-Canonical Wnt Co-Receptor Ror2 Expression in Human Bone Marrow Osteoprogenitor Cells: Effect of Wnt5a/Ror2 Pathway Activation on the Osteogenic Differentiation Impairment Induced by Myeloma Cells. Leukemia 2013, 27, 451–463. [Google Scholar] [CrossRef]
- Qiang, Y.-W.; Shaughnessy, J.D.; Yaccoby, S. Wnt3a Signaling within Bone Inhibits Multiple Myeloma Bone Disease and Tumor Growth. Blood 2008, 112, 374–382. [Google Scholar] [CrossRef]
- Suthon, S.; Perkins, R.S.; Bryja, V.; Miranda-Carboni, G.A.; Krum, S.A. WNT5B in Physiology and Disease. Front. Cell Dev. Biol. 2021, 9, 667581. [Google Scholar] [CrossRef]
- Song, S.; Fan, G.; Li, Q.; Su, Q.; Zhang, X.; Xue, X.; Wang, Z.; Qian, C.; Jin, Z.; Li, B.; et al. IDH2 Contributes to Tumorigenesis and Poor Prognosis by Regulating M6A RNA Methylation in Multiple Myeloma. Oncogene 2021, 40, 5393–5402. [Google Scholar] [CrossRef]
- Katoh, M. Networking of WNT, FGF, Notch, BMP, and Hedgehog Signaling Pathways during Carcinogenesis. Stem Cell Rev. 2007, 3, 30–38. [Google Scholar] [CrossRef]
- Shi, C.; Chen, X.; Yin, W.; Sun, Z.; Hou, J.; Han, X. Wnt8b Regulates Myofibroblast Differentiation of Lung-Resident Mesenchymal Stem Cells via the Activation of Wnt/β-Catenin Signaling in Pulmonary Fibrogenesis. Differentiation 2022, 125, 35–44. [Google Scholar] [CrossRef]
- Zmuda, J.M.; Yerges, L.M.; Kammerer, C.M.; Cauley, J.A.; Wang, X.; Nestlerode, C.S.; Wheeler, V.W.; Patrick, A.L.; Bunker, C.H.; Moffett, S.P.; et al. Association Analysis of WNT10B with Bone Mass and Structure among Individuals of African Ancestry. J. Bone Miner. Res. 2009, 24, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Wend, P.; Wend, K.; Krum, S.A.; Miranda-Carboni, G.A. The Role of WNT10B in Physiology and Disease. Acta Physiol. 2012, 204, 34–51. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of Osteoblast and Odontoblast Differentiation by RUNX2. J. Oral Biosci. 2010, 52, 22–25. [Google Scholar] [CrossRef]
- Reya, T.; O’Riordan, M.; Okamura, R.; Devaney, E.; Willert, K.; Nusse, R.; Grosschedl, R. Wnt Signaling Regulates B Lymphocyte Proliferation through a LEF-1 Dependent Mechanism. Immunity 2000, 13, 15–24. [Google Scholar] [CrossRef]
- Bunaciu, R.P.; Tang, T.; Mao, C.D. Differential Expression of Wnt13 Isoforms during Leukemic Cell Differentiation. Oncol. Rep. 2008, 20, 195–201. [Google Scholar] [CrossRef]
- Richter, J.; Stanley, E.G.; Ng, E.S.; Elefanty, A.G.; Traver, D.; Willert, K. WNT9A Is a Conserved Regulator of Hematopoietic Stem and Progenitor Cell Development. Genes 2018, 9, 66. [Google Scholar] [CrossRef]
- Kirikoshi, H.; Sekihara, H.; Katoh, M. Expression of WNT14 and WNT14B MRNAs in Human Cancer, up-Regulation of WNT14 by IFNγ and up-Regulation of WNT14B by β-Estradiol. Int. J. Oncol. 2001, 19, 1221–1225. [Google Scholar] [CrossRef]
- Lu, S.; Yakirevich, E.; Yang, D.; Xiao, Y.; Wang, L.J.; Wang, Y. Wnt Family Member 9b (Wnt9b) Is a New Sensitive and Specific Marker for Breast Cancer. Am. J. Surg. Pathol. 2021, 45, 1633–1640. [Google Scholar] [CrossRef]
- Spaan, I.; Raymakers, R.A.; van de Stolpe, A.; Peperzak, V. Wnt Signaling in Multiple Myeloma: A Central Player in Disease with Therapeutic Potential. J. Hematol. Oncol. 2018, 11, 67. [Google Scholar] [CrossRef]
- Van Andel, H.; Kocemba, K.A.; Spaargaren, M.; Pals, S.T. Aberrant Wnt Signaling in Multiple Myeloma: Molecular Mechanisms and Targeting Options. Leukemia 2019, 33, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Frenquelli, M.; Caridi, N.; Antonini, E.; Storti, F.; Viganò, V.; Gaviraghi, M.; Occhionorelli, M.; Bianchessi, S.; Bongiovanni, L.; Spinelli, A.; et al. The WNT Receptor ROR2 Drives the Interaction of Multiple Myeloma Cells with the Microenvironment through AKT Activation. Leukemia 2020, 34, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shi, C.; Cao, H.; Chen, L.; Hou, J.; Xiang, Z.; Hu, K.; Han, X. The Hedgehog and Wnt/β-Catenin System Machinery Mediate Myofibroblast Differentiation of LR-MSCs in Pulmonary Fibrogenesis. Cell Death Dis. 2018, 9, 639. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, R.; Basu, D.; Dutta, T.K. Significance of Bone Marrow Fibrosis in Multiple Myeloma. Pathology 2007, 39, 512–515. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Sample ID | Type of M Protein | PC Infiltration of BM (Histological Examination of Trephine Biopsy) | Percentage of PC in Bone Marrow Aspirate | Microvessel Density of BM (Histological Examination of Biopsy) | Treatment Regimens * | Response to Therapy |
|---|---|---|---|---|---|---|
| 1 | Non-secretory MM | 10% | 6% | 7.2% | VCD, 4 cycles | PR |
| 2 | Ig G κ | 10% | 3% | 8.9% | VCD, 6 cycles; aHSCT | CR |
| 3 | Ig A κ | 5% | 2.4% | 10.2% | VCD, 5 cycles | PR |
| 4 | Ig G λ | 1–2% | 1% | 7.4% | VCD, 5 cycles; aHSCT | CR |
| 5 | Ig G κ | 3% | 3.6% | 8.5% | VRD, 2 cycles, +2 KRd cycles; aHSCT | VGPR |
| 6 | Ig G λ | 30% | 2.2% | 11.7% | VCD, 3 cycles; aHSCT | VGPR |
| 7 | Ig G κ | 1–2% | 1.4% | 10.6% | VCD, 5 cycles; aHSCT | PR |
| 8 | Ig G κ | 1–2% | 2.6% | 8.6% | CV, 6 cycles | CR |
| 9 | Ig G λ | 1–2% | 2.8% | 9.2% | VRD, 1 cycle; aHSCT | VGPR |
| 10 | Ig G κ | 50% | 47% | 13% | VD, 6 cycles; aHSCT | NR |
| 11 | Ig A κ | 50% | 14.4% | 12.5% | VCD, 3 cycles; aHSCT | NR |
| 12 | Ig G λ | 90% | 82.4% | 11.4% | VCD, 2 cycles | NR |
| 13 | Ig G κ | N/A | 2.4% | N/A | Untreated | UT |
| 14 | Ig A | 90% | 66% | 9.3% | Untreated | UT |
| 15 | Ig G κ | N/A | 84% | N/A | Untreated | UT |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| WNT3 | 5′-GGAGAAGCGGAAGGAAAAATG-3′ | 5′-GCACGTCGTAGATGCGAATACA-3′ |
| WNT3A | 5′-CCTGCACTCCATCCAGCTACA-3′ | 5′-GACCTCTCTTCCTACCTTTCCCTTA-3′ |
| WNT5A | 5′-GAAATGCGTGTTGGGTTGAA-3′ | 5′-ATGCCCTCTCCACAAAGTGAA-3′ |
| WNT5B | 5′-CTGCCTTTCCAGCGAGAATT-3′ | 5′-AGGTCAAATGGCCCCCTTT-3′ |
| WNT7B | 5′-CCCGGCAAGTTCTCTTTCTTC-3′ | 5′-GGCGTAGCTTTTCTGTGTCCAT-3′ |
| WNT8B | 5′-TCCCAGAAAAACTGAGGAAACTG-3′ | 5′-AACCTCTGCCTCTAGGAACCAA-3′ |
| WNT10B | 5′-CTTTTCAGCCCTTTGCTCTGAT-3′ | 5′-CCCCTAAAGCTGTTTCCAGGTA-3′ |
| WNT2B (former WNT13) | 5′-TGCCAAGGAGAAGAGGCTTAAG-3′ | 5′-GTGCGACCACAGCGGTTATT-3′ |
| WNT9A (former WNT14) | 5′-CTTAAGTACAGCAGCAAGTTCGTCAA-3′ | 5′-CCACGAGGTTGTTGTGGAAGT-3′ |
| WNT 9B (former WNT15) | 5′-CAGGTGCTGAAACTGCGCTAT-3′ | 5′-GCCCAAGGCCTCATTGGT-3′ |
| CTNNB1 (β- catenin) | 5′-CTGCTGTTTTGTTCCGAATGTC-3′ | 5′-CCATTGGCTCTGTTCTGAAGAGA-3′ |
| reference gene GAPDH | 5′-AGGTCGGAGTCAACGGATTT-3′ | 5′-TTCCCGTTCTCAGCCTTGAC-3′ |
| Gene Name | mRNA Level in MSCs (Relative to HD) | ||
|---|---|---|---|
| UT | NR | PoCR | |
| WNT3 | + | − | +/− |
| WNT3A | + | − | − |
| WNT5A | + | +/− | + |
| WNT5B | + | +/− | +/− |
| WNT7B | +/− | − | − |
| WNT8B | + | + | + |
| WNT10B | + | + | + |
| WNT11 | + | +/− | +/− |
| WNT2B (WNT13) | +/− | − | +/−− |
| WNT9A (WNT14) | + | − | − |
| WNT9B (WNT15) | + | + | +/− |
| CTNNB1 | − | +/− | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enukashvily, N.I.; Belik, L.A.; Semenova, N.Y.; Kostroma, I.I.; Motyko, E.V.; Gritsaev, S.V.; Bessmeltsev, S.S.; Sidorkevich, S.V.; Martynkevich, I.S. Transcription of WNT Genes in Hematopoietic Niche’s Mesenchymal Stem Cells in Multiple Myeloma Patients with Different Responses to Treatment. Genes 2023, 14, 1097. https://doi.org/10.3390/genes14051097
Enukashvily NI, Belik LA, Semenova NY, Kostroma II, Motyko EV, Gritsaev SV, Bessmeltsev SS, Sidorkevich SV, Martynkevich IS. Transcription of WNT Genes in Hematopoietic Niche’s Mesenchymal Stem Cells in Multiple Myeloma Patients with Different Responses to Treatment. Genes. 2023; 14(5):1097. https://doi.org/10.3390/genes14051097
Chicago/Turabian StyleEnukashvily, Natella I., Liubov. A. Belik, Natalia Yu. Semenova, Ivan I. Kostroma, Ekaterina V. Motyko, Sergey V. Gritsaev, Stanislav S. Bessmeltsev, Sergey V. Sidorkevich, and Irina S. Martynkevich. 2023. "Transcription of WNT Genes in Hematopoietic Niche’s Mesenchymal Stem Cells in Multiple Myeloma Patients with Different Responses to Treatment" Genes 14, no. 5: 1097. https://doi.org/10.3390/genes14051097