
Metabolite Profiling and Transcriptome Analysis Explain the Difference in Accumulation of Bioactive Constituents in Taxilli Herba from Two Hosts
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Metabolite Profiling Analysis of Bioactive Constituents in Taxilli Herba
2.3. Transcriptome Analysis of Taxilli Herba
2.3.1. RNA Preparation
2.3.2. Library Preparation and Sequencing
2.3.3. De Novo Assembly and Function Annotation
2.3.4. Identification and Analysis of Differentially Expressed Genes
2.3.5. qRT-PCR Verification
2.3.6. Data Processing
3. Results
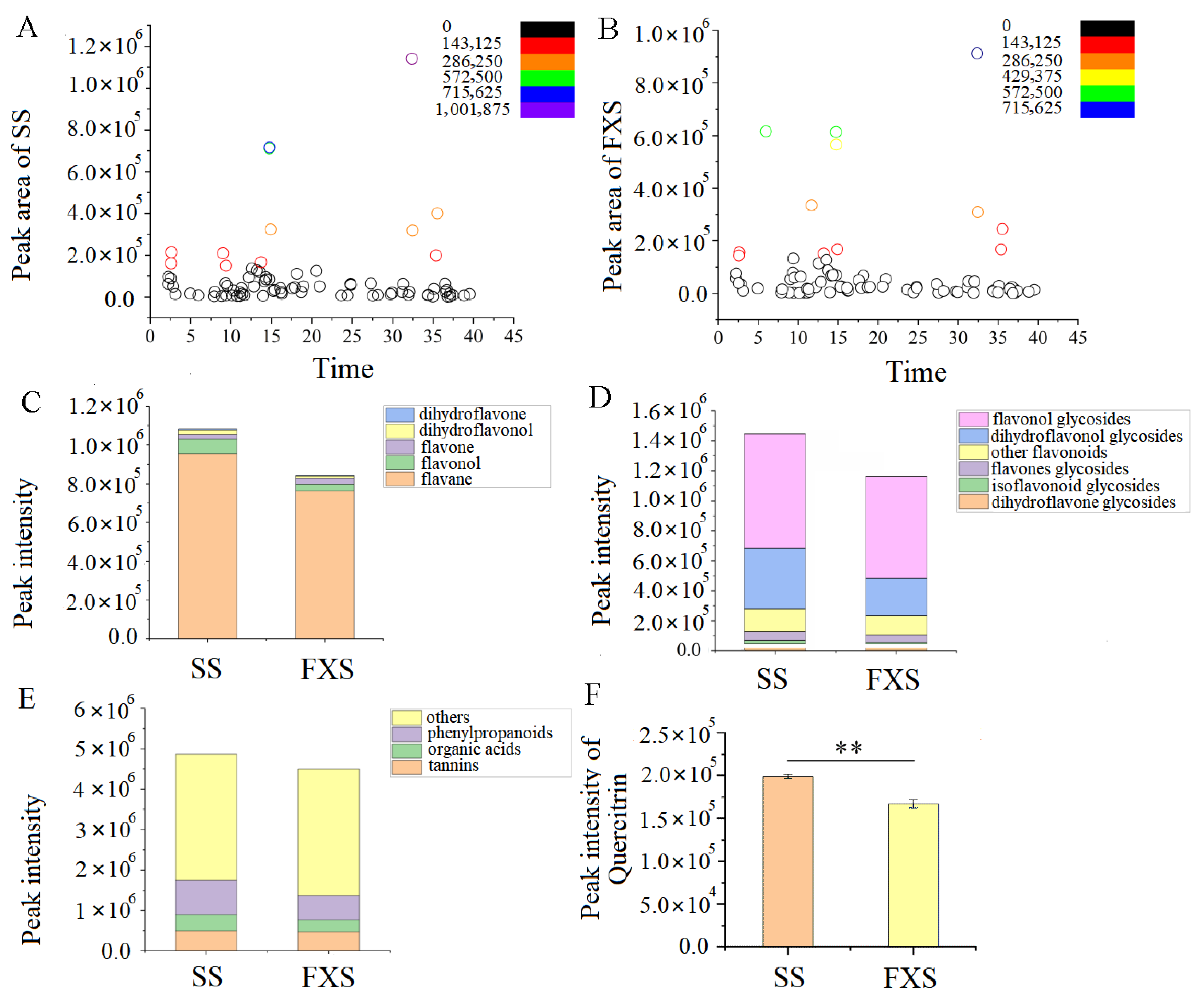
3.1. The Change in Metabolic Profiling and Metabolite Content in Taxilli Herba
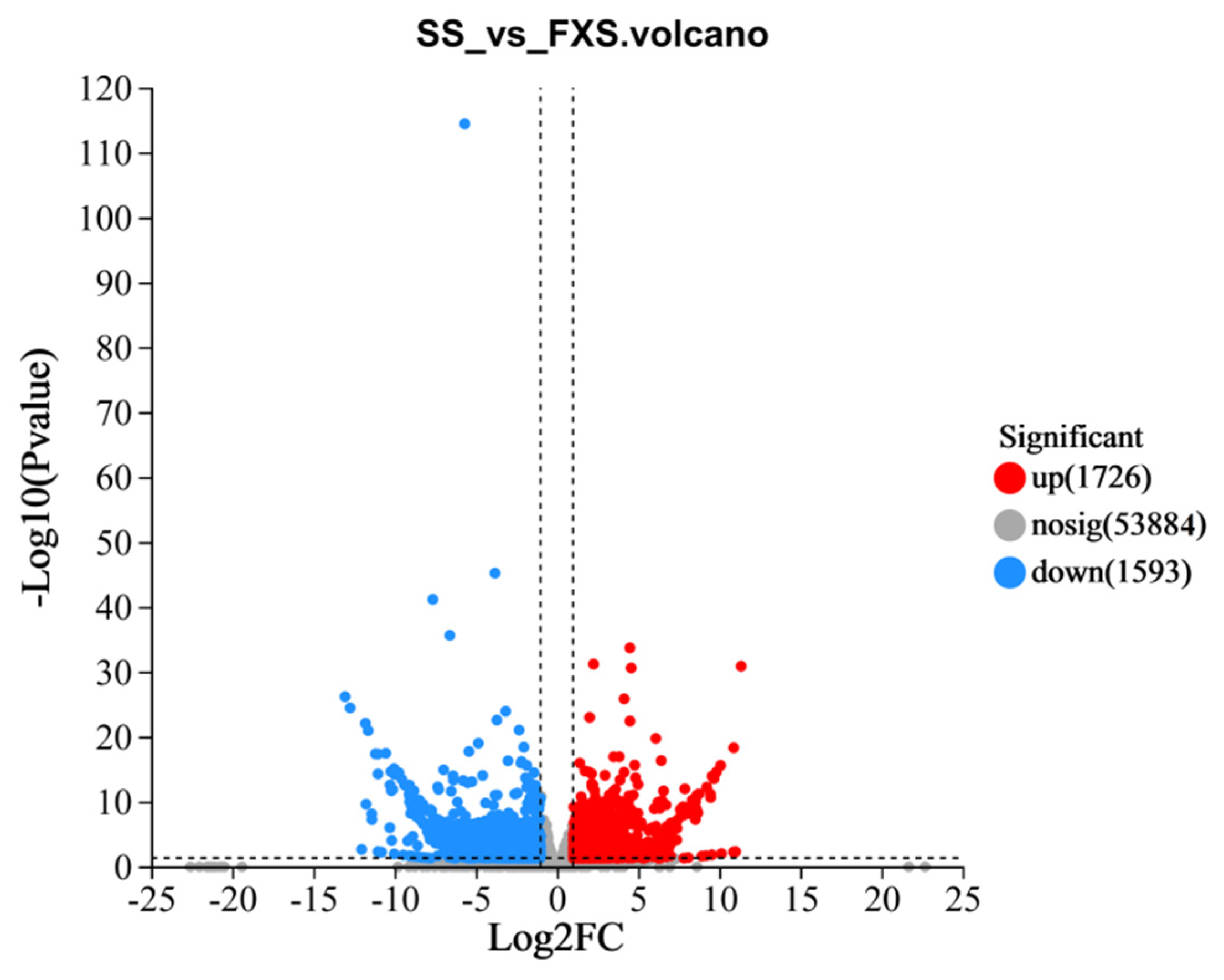
3.2. De Novo Transcriptome Assembly and Sequence Analysis
3.3. Functional Annotation and Classification
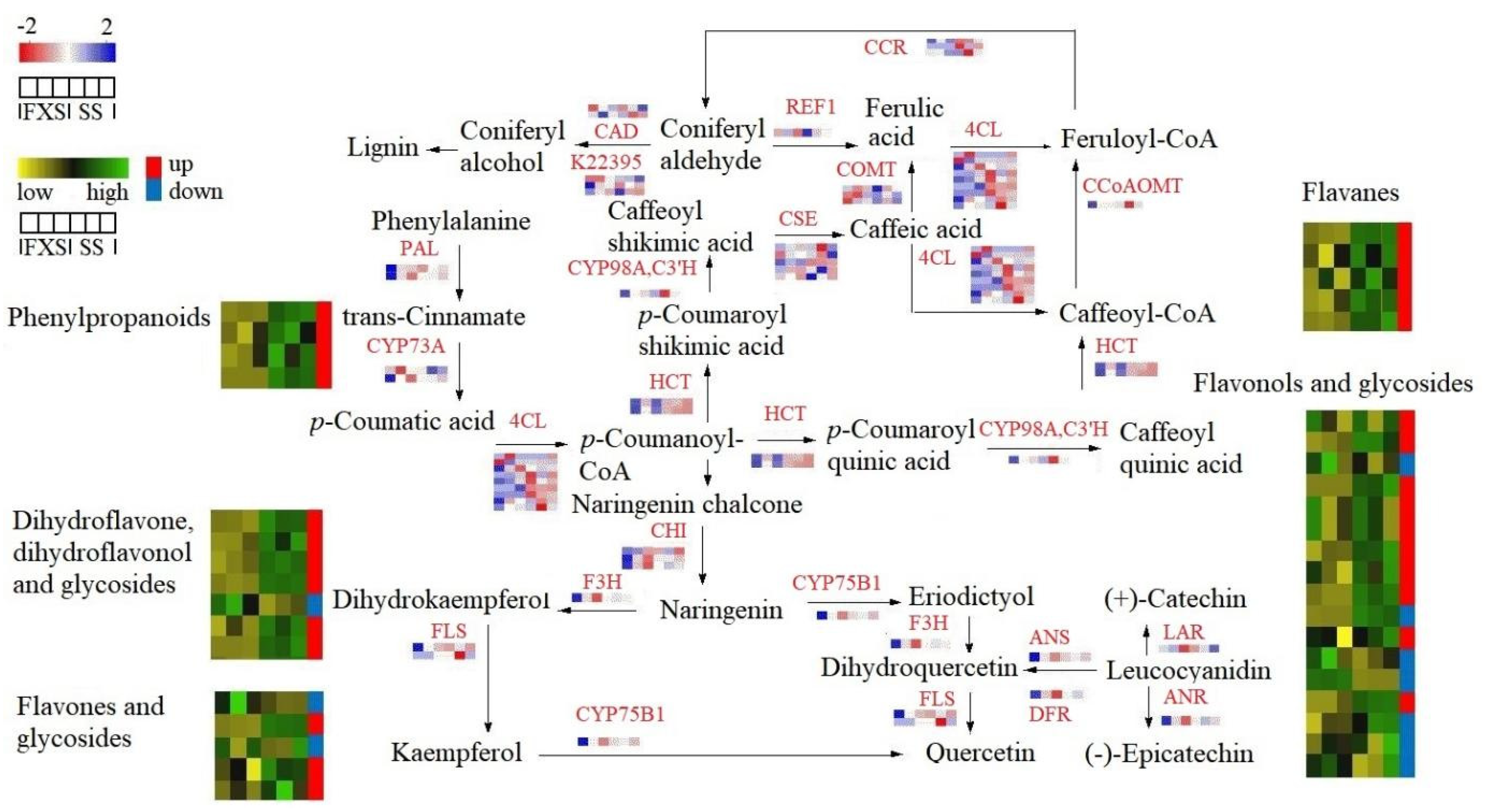
3.4. Analysis of Genes and Metabolites Involved in Flavonoid Biosynthesis in Taxilli Herba
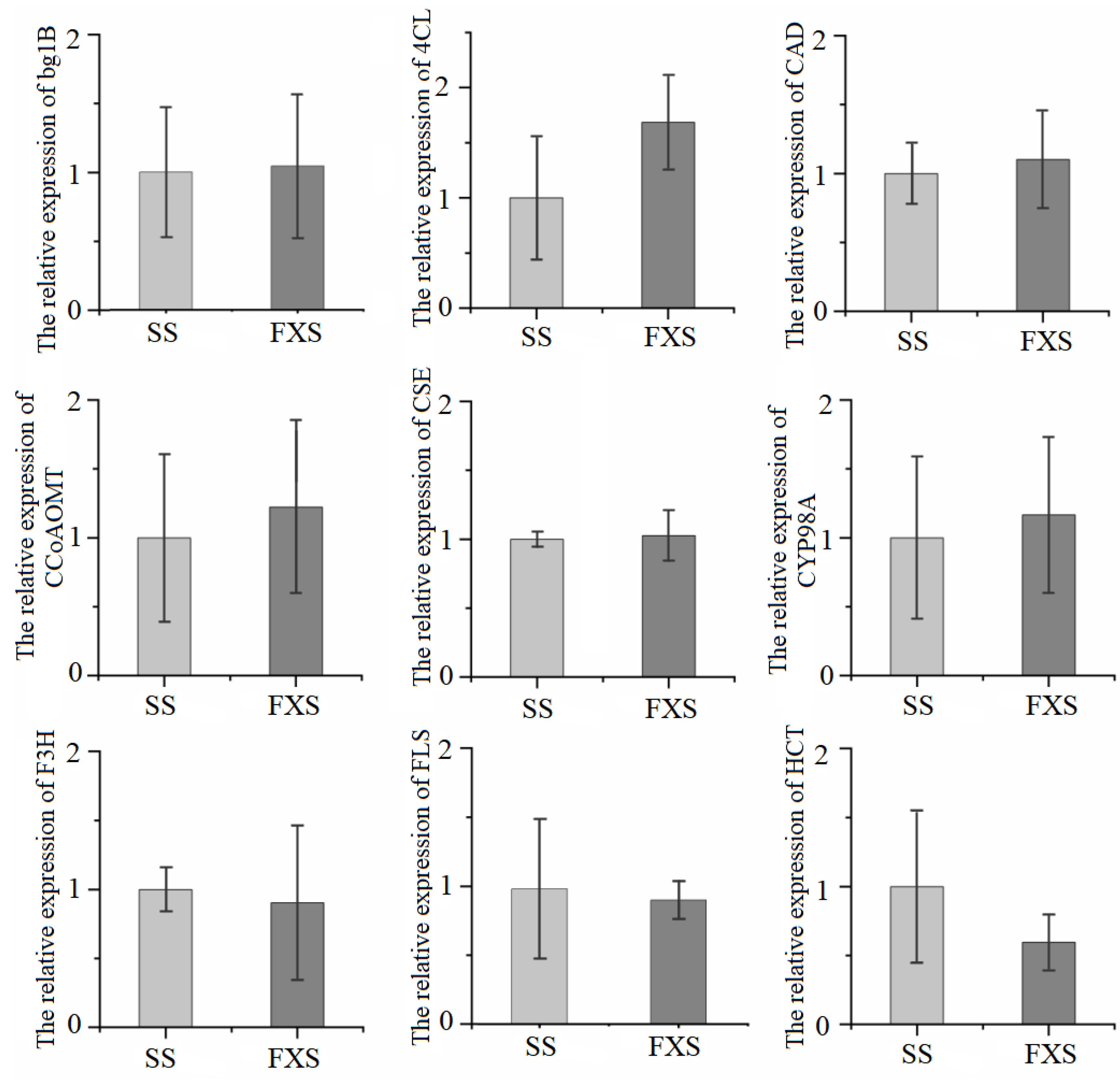
3.5. Confirmation of the Expression of Related Genes Using qRT-PCR
3.6. Transcription Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- National Pharmacopoeia Committee. Pharmacopoeia of the People’s Republic of China. Part I; Chemical Industry Press: Beijing, China, 2020; p. 312.
- Li, X.F.; Fang, R.Z.; Feng, H.; Wang, X.P. Study on the mechanism of Taxillus chinensis (DC.) Danser of tonifing liver and kidney, strengthening muscles and bones based on network pharmacology. Chin. J. Ethnomed. Ethnopharm. 2021, 30, 16–26. [Google Scholar]
- Huang, F.; Qin, Z.M.; Li, Y.H. Relationship between Taxillus chinensis Tea and the culture of south of of the five ridges. Asia-Pac. Trad. Med. 2018, 14, 3–84. [Google Scholar]
- Lv, L.; Zhu, Y.M.; Xu, D.M. Study on flavones from Taxillus chinensis (DC.) Danser and assay of its quercetin. Chin. Tradit. Pat. Med. 2004, 26, 68–70. [Google Scholar]
- Liang, Y.; Li, L.; Cai, Y.; Xu, L.B.; Xie, F.F.; Liang, D.L.; Chen, F.L. Analysis of chemical constituents in ethyl acetate extract of Taxilli Herba by UPLC-Q-Exactive-MS and screening of potential xanthine oxidase inhibitors. China J. Chin. Mater. Med. 2022, 47, 972–979. [Google Scholar]
- Huang, F.Y.; Liu, R.Y.; Chai, Z.S.; Su, B.W.; Zhu, K.X.; Lu, H.L.; Li, Y.H. Study on aromatic components in essential oils of Taxillus chinensis from different cold/hot properties hosts by GC–MS. Asia–Pac. Trad. Med. 2019, 15, 61–66. [Google Scholar]
- Zhu, K.X.; Lu, D.; Pei, H.H.; Zhao, M.; Li, Y.H. A survey on the distribution of Taxilli Herba and its host status in Guangxi. Guangxi J. Tradit. Chin. Med. 2010, 33, 59–61. [Google Scholar]
- Zhang, H.; Huang, F.Y.; Su, B.W.; Zhu, K.X.; Lu, H.L.; Yin, S.G.; Guo, M.; Li, Y.H. Impacts of different host trees on the quality of Taxillus chinensis. Mod. Tradit. Chin. Med. Mat. Medica-World Sci. Tech. 2016, 18, 1182–1187. [Google Scholar]
- Wu, N.; Yuan, J.H.; Wang, W.X.; Liu, X.H.; Li, L.; Yin, S.X.; Chen, H.J.; Xue, J.; Zhou, Y.Y.; Yang, W. Simultaneous determination of multiple active constituents in Taxilli Herba by HPLC-QTRAP-MS/MS. J. Instr. Anal. 2022, 41, 1153–1162. [Google Scholar]
- Huang, F.Y.; Liu, R.Y.; Lu, H.; Li, Y.H. Effect of different medicinal hosts on the total flavonoid content of Taxilli Herba. J. Guangxi Univ. Chin. Med. 2017, 20, 34–36. [Google Scholar]
- Su, B.W.; Wang, H.; Li, Y.H.; Pei, H.H.; Zhu, K.X.; Lu, D. Content analysis of avicularin, quercitrin and quercetin in Herba Taxilli from different host plants. J. Int. Pharm. Res. 2017, 44, 738–742. [Google Scholar]
- Liu, R.Y.; Huang, F.Y.; Guo, M.; Lu, H.L.; Zhu, K.X.; Qin, W.H.; Qin, Z.M.; Li, Y.H.; Su, B.W. Correlation study on chemical constituents of cardiac glycosides in Taxillus chinensis and its Nerium indicum host by UPLC-Q-TOF-MS/MS. China J. Chin. Mater. Med. 2019, 44, 2283–2291. [Google Scholar]
- Zhang, X.J.; Su, B.W.; Li, J.; Li, Y.H.; Lu, D.; Zhu, K.X.; Pei, H.H.; Zhao, M.H. Analysis by RP-HPLC of mangiferin component correlation between medicinal Loranthus and their mango host trees. J. Chromatogr. Sci. 2014, 52, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.P.; He, C.H.; Chai, Z.S.; Qin, W.M.; Wu, W.X.; Chen, L.Y.; Liu, J.L.; Ru, M.; Li, Y.H. Acute toxicity and hepatotox- icity of aqueous extracts of Taxilli Herba from different hosts in zebrafish. Chin. J. Exp. Tradit. Med. Form. 2021, 27, 91–97. [Google Scholar]
- Zhang, H.; Huang, F.Y.; Liu, R.Y.; Lu, H.L.; Su, B.W.; Zhu, K.X.; Li, Y.H. Effect of Taxilli Herba from different hosts on lowering blood pressure of spontaneously hypertensive rats. Chin. Tradit. Pat. Med. 2018, 40, 249–254. [Google Scholar]
- Yuan, J.H.; Cai, Z.C.; Chen, C.H.; Wu, N.; Yin, S.X.; Wang, W.X.; Chen, H.J.; Zhou, Y.Y.; Li, L.; Liu, X.H. A study for quality evaluation of Taxilli Herba from different hosts based on fingerprint-activity relationship modeling and multivariate statistical analysis. Arab. J. Chem. 2022, 15, 103933. [Google Scholar] [CrossRef]
- Lu, S.H.; Gan, Y.Y.; Tang, L.; Zhang, F.Y.; Su, B.W. Evaluation of the quality of Taxillus chinensis from different hosts sources by HPLC fingerprint combined with chemometrics. China Pharm. 2020, 31, 794–799. [Google Scholar]
- Yin, S.G.; Liu, R.Y.; Huang, F.Y.; Li, Y.H.; Li, J.Y. Study on infrared spectra of Taxillus chinensis (DC.) Danser from different host trees. Lishizhen Med. Mater. Med. Res. 2018, 29, 2629–2631. [Google Scholar]
- Xu, G.H.; Cao, J.J.; Wang, X.F.; Chen, Q.Y.; Jin, W.W.; Li, Z.; Tian, F. Evolutionary metabolomics identifies substantial metabolic divergence between Maize and Its wild ancestor, Teosinte. Plant Cell. 2019, 31, 1990–2009. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Tohge, T.; Niida, R.; Saito, K. Identification of a flavonol 7-O-rhamnosyltransferasegene determining flavonoid pattern in Arabidopsis by transcriptome coexpressionanalysis and reverse genetics. J. Biol. Chem. 2007, 282, 14932–14941. [Google Scholar] [CrossRef]
- Soltani, B.M.; Ehlting, J.; Hamberger, B.; Douglas, C.J. Multiple cis-regulatory elements regulate distinct and complex patterns of developmental and wound-induced expression of Arabidopsis thaliana 4CL gene family members. Planta 2006, 224, 1226–1238. [Google Scholar] [CrossRef]
- Tohge, T.; Nishiyama, Y.; Hirai, M.Y.; Yano, M.; Nakajima, J.; Awazuhara, M.; Inoue, E.; Takahashi, H.; Goodenowe, D.B.; Kitayama, M.; et al. Functional genomics by integrated analysis of metabolome andtranscriptome of Arabidopsis plants over-expressing an MYB transcriptionfactor. Plant J. 2005, 42, 218–235. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.C.; Zhang, H.; Zhao, D.; Wang, F.R.; Liu, X.G.; Guo, L. Purification and function exploration of transcription factor MYB54 in Arabidopsis thaliana. Acta Agric. Boreali. Sin. 2021, 4, 47–55. [Google Scholar]
- Saffer, A.M.; Irish, V.F. Flavonol rhamnosylation indirectly modifies the cell wall defects of rhamnose biosynthesis1 mutants by altering rhamnose flux. Plant J. 2018, 94, 649–660. [Google Scholar] [CrossRef]
- Tohge, T.; Fernie, A.R. Specialized metabolites of the flavonol class mediate root phototropism and growth. Mol. Plant. 2016, 9, 1554–1555. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Y.J.; Zhang, F.J.; Zhang, G.Z.; Jiang, X.Y.; Yu, H.M.; Hou, B.K. The Arabidopsis UDP-glycosyltransferases UGT79B2 and UGT79B3, contribute to cold, salt and drought stress tolerance via modulating anthocyanin accumulation. Plant J. 2017, 89, 85–103. [Google Scholar] [CrossRef]
- Peng, M.; Shahzad, R.; Gul, A.; Subthain, H.; Shen, S.; Lei, L.; Zheng, Z.; Zhou, J.; Lu, D.; Wang, S.; et al. Differentially evolved glucosyltransferases determine natural variation of rice flavone accumulation and UV-tolerance. Nat. Commun. 2017, 8, 1975–1986. [Google Scholar] [CrossRef]
- Demkura, P.V.; Ballaré, C.L. UVR8 mediates UV-B-induced Arabidopsis defense responses against botrytis cinerea by controlling sinapate accumulation. Mol. Plant. 2012, 5, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.Y.; Zeng, C.B.; Liu, C.L.; Wei, S.S.; Huang, J.Q.; Tang, H. Differential expression analysis of genes related to flesh color in Hylocereu polyrhizus and Hylocereu undatus. Mol. Plant Breed. 2021, 19, 4311–4326. [Google Scholar] [CrossRef]
- Zhu, D.; Li, L.H.; Gao, H. Identification of genes in response to drought and high temperature stress in Mentha haplocalyx seedlings based on transcriptome sequencing. Shengming Huaxue. 2020, 40, 2270–2284. [Google Scholar]
- Yang, Z.M.; Wu, Y.P.; Dai, Z.G.; Chen, X.J.; Wang, H.Q.; Yang, S.; Xie, D.W.; Tang, Q.; Cheng, C.H.; Xu, Y.; et al. Comprehensive transcriptome analysis and tissue-specific profiling of gene expression in jute (Corchorus olitorius. L). Ind. Crops. Prod. 2020, 146, 112101. [Google Scholar] [CrossRef]
- Tang, H.; Zhang, M.; Liu, J.Y.; Cai, J. Metabolomic and transcriptomic analyses reveal the characteristics of tea flavonoids and caffeine accumulationand regulation between Chinese Varieties (Camellia sinensis var. sinensis) and Assam Varieties (C. sinensis var. assamica). Genes 2022, 13, 1994–2009. [Google Scholar]
- Mei, X.; Wan, S.H.; Lin, C.Y.; Zhou, C.B.; Hu, L.H.; Deng, C.; Zhang, L.Y. Integration of metabolome and transcriptome reveals the relationship of benzenoid-phenylpropanoid pigment and aroma in purple tea flowers. Front. Plant Sci. 2021, 12, 762330. [Google Scholar] [CrossRef] [PubMed]
- Savoi, S.; Wong, D.C.; Arapitsas, P.; Miculan, M.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Transcriptome and metabolite profiling reveals that prolonged drought modulates the phenylpropanoid and terpenoid pathway in white grapes (Vitis vinifera L.). BMC Plant Biol. 2016, 16, 67–83. [Google Scholar]
- Yuan, J.H.; Li, L.; Cai, Z.C.; Wu, N.; Chen, C.H.; Yin, X.S.; Wang, W.X.; Mei, Y.Q.; Wei, L.F.; Liu, X.H.; et al. Qualitative analysis and componential differences of chemical constituents in Taxilli Herba from different hosts by UFLC-Triple TOF-MS/MS. Molecules 2021, 26, 6373. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Hong, L.; Yang, L.; Yang, H.J.; Wang, W. Research advances in AP2/ERF transcription factors in regulating plant responses to abiotic stress. Chin. Bull. Bot. 2020, 55, 481–496. [Google Scholar]
- Duan, L.P.; Xie, L.Q.; Ma, Y.M.; He, J. Regulation of secondary metabolism by transcription factors in medicinal plants. J. Chin. Me. Mater. 2021, 44, 1002–1007. [Google Scholar]
- Wang, X.P.; Niu, Y.L.; Zheng, Y. Multiple functions of MYB transcription factors in abiotic stress responses. Int. J. Mol. Sci. 2021, 22, 6125. [Google Scholar] [CrossRef]
- Liu, M.Q.; Sun, W.; Meng, X.X.; Wang, H.H.; Liu, T.X.; Sun, J.Y.; Wang, Z.; Mi, Y.; Wang, L. Identification and expression analysis of the C2H2 gene family in Cannabis sativa L. Acta Pharm. Sin. 2021, 56, 1486–1496. [Google Scholar]
- Tao, H.J. Annotation of Materia Medica; People’s Medical Publishing House: Beijing, China, 1994; pp. 254–255. [Google Scholar]
- Tang, S.W. Classified Materia Medica; China Press: Beijing, China, 1993; pp. 359–360. [Google Scholar]
- Li, S.Z. Compendium of Materia Medica; People’s Medical Publishing House: Beijing, China, 1982; Volume 2, pp. 2158–2159. [Google Scholar]
- Fernandez, C.; Monnier, Y.; Santonja, M.; Gallet, C.; Weston, L.A.; Prévosto, B.; Saunier, A.; Baldy, V.; Bousquet-Mélou, A. The impact of competition and allelopathy on the trade-off between plant defense and growth in two contrasting tree species. Front. Plant Sci. 2016, 7, 594–607. [Google Scholar] [CrossRef]
- Wang, C.C.; Chen, L.H.; Cai, Z.C.; Chen, C.H.; Liu, Z.X.; Liu, S.J.; Zou, L.S.; Tan, M.X.; Chen, J.L.; Liu, X.H.; et al. Metabolite profiling and transcriptome analysis explains difference in accumulation of bioactive constituents in Licorice (Glycyrrhiza uralensis) under salt stress. Front. Plant Sci. 2021, 12, 727882. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.Q.; Diao, P.F.; Liao, J.; Chi, Y.C.; Ha, D. Advances in molecular communication between parasitic plants and their hosts. Plant Physiol. J. 2018, 54, 519–527. [Google Scholar]
- Liu, N.; Shen, G.J.; Xu, Y.X.; Liu, H.; Zhang, J.X.; Li, S.L.; Li, J.; Zhang, C.P.; Qi, J.F.; Wang, L.; et al. Extensive inter-plant protein transfer between Cuscuta parasutes and their host plants. Mol. Plant 2019, 13, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, Y.C.; Chen, Y.; Long, Y.Q.; Tong, Q.Z.; Liu, X.D.; Zhou, R.B. Cloning and expression analysis of phenylalanine ammonia-lyase gene in Lonicera macranthoides. Chin. Tradit. Herb Drugs 2019, 50, 178–187. [Google Scholar]
- Heldt, H.W.; Piechulla, B. Phenylpropanoids comprise a multitude of plant-specialized metabolites and cell wall components. Plant Biochem. 2021, 15, 411–429. [Google Scholar]
- Wang, Y.; Yang, X.; Yang, R.J.; Wang, Y.X.; Yang, Y.X.; Xia, P.F.; Zhao, L. Advances in research of MYB transcription factors in regulating phenylpropane biosynthesis. J. Anhui Agric. Univ. 2019, 46, 859–864. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, J.; Wu, N.; Cai, Z.; Chen, C.; Zhou, Y.; Chen, H.; Xue, J.; Liu, X.; Wang, W.; Cheng, J.; et al. Metabolite Profiling and Transcriptome Analysis Explain the Difference in Accumulation of Bioactive Constituents in Taxilli Herba from Two Hosts. Genes 2023, 14, 1040. https://doi.org/10.3390/genes14051040
Yuan J, Wu N, Cai Z, Chen C, Zhou Y, Chen H, Xue J, Liu X, Wang W, Cheng J, et al. Metabolite Profiling and Transcriptome Analysis Explain the Difference in Accumulation of Bioactive Constituents in Taxilli Herba from Two Hosts. Genes. 2023; 14(5):1040. https://doi.org/10.3390/genes14051040
Chicago/Turabian StyleYuan, Jiahuan, Nan Wu, Zhichen Cai, Cuihua Chen, Yongyi Zhou, Haijie Chen, Jia Xue, Xunhong Liu, Wenxin Wang, Jianming Cheng, and et al. 2023. "Metabolite Profiling and Transcriptome Analysis Explain the Difference in Accumulation of Bioactive Constituents in Taxilli Herba from Two Hosts" Genes 14, no. 5: 1040. https://doi.org/10.3390/genes14051040