
Freezing Does Not Alter Sperm Telomere Length despite Increasing DNA Oxidation and Fragmentation
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Design
2.3. Standard Semen Analysis
2.4. Sperm Freezing and Thawing
2.5. Sperm Telomere Length Measurements
STL Measurement by qPCR
2.6. STL Measurement by Q-FISH
2.7. TUNEL Assay
2.8. 8-OHdG Immuno-Detection
2.9. Statistical Analysis
3. Results
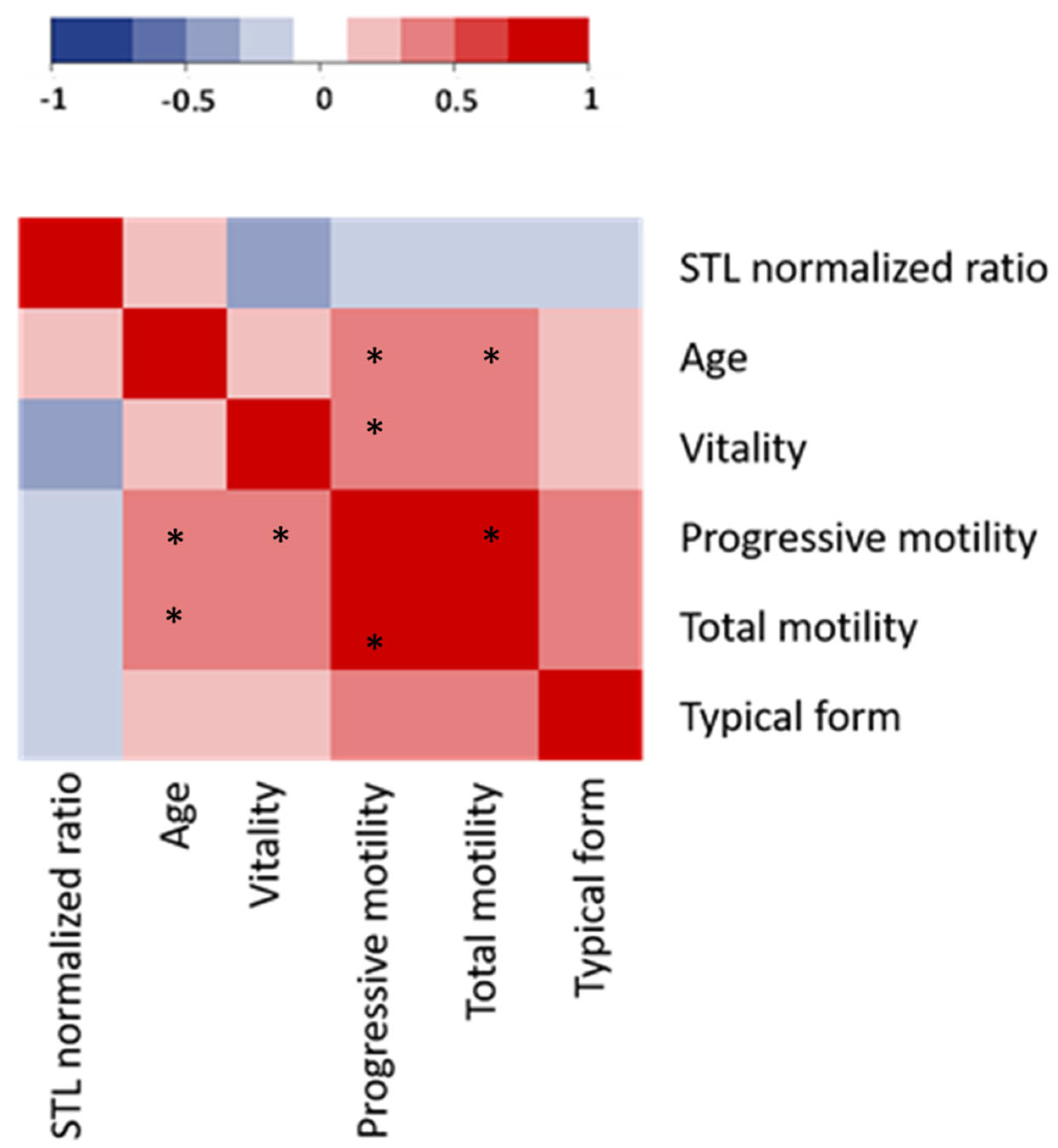
3.1. Patient Characteristics, Sperm Analysis, and STL
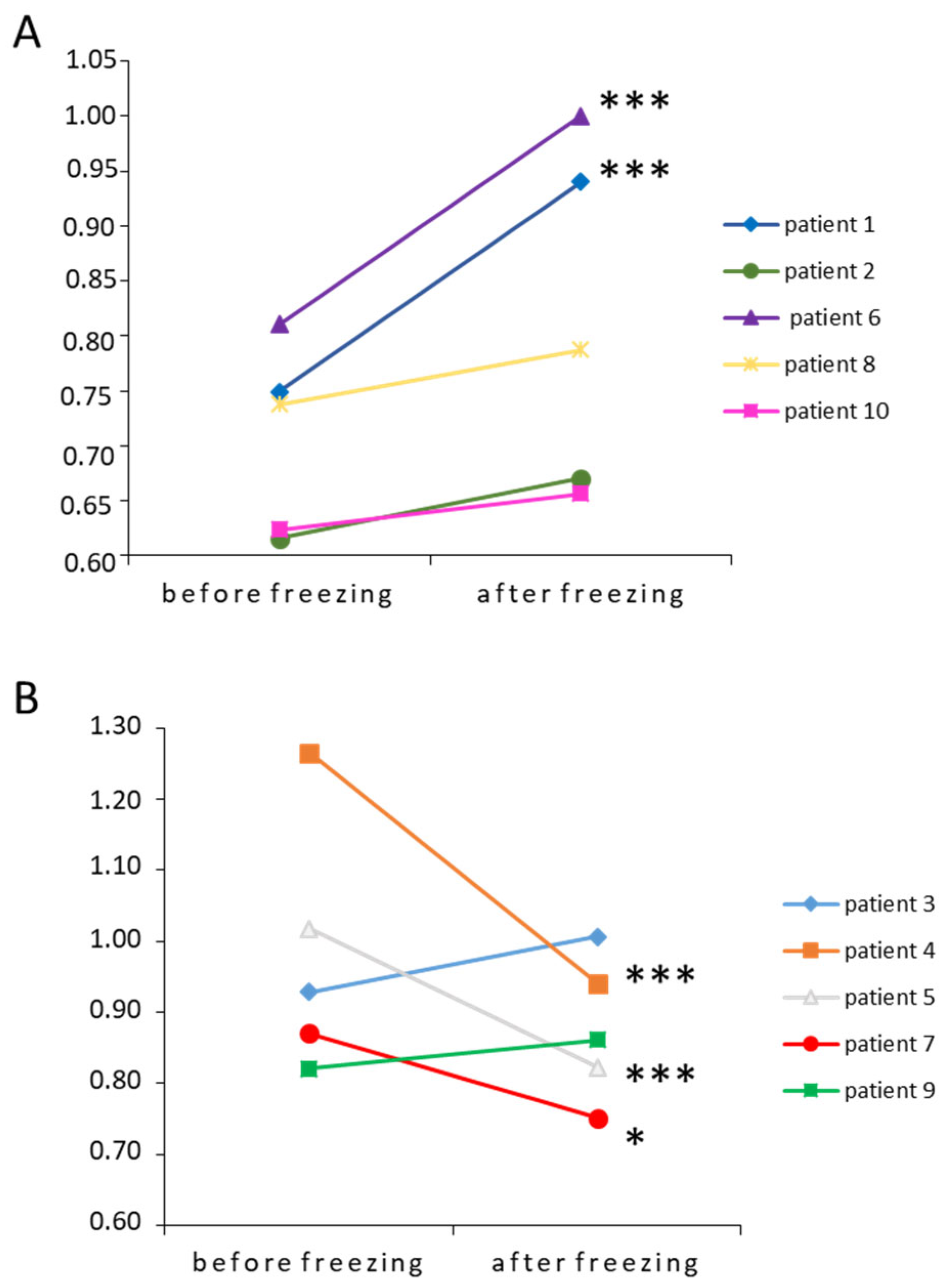
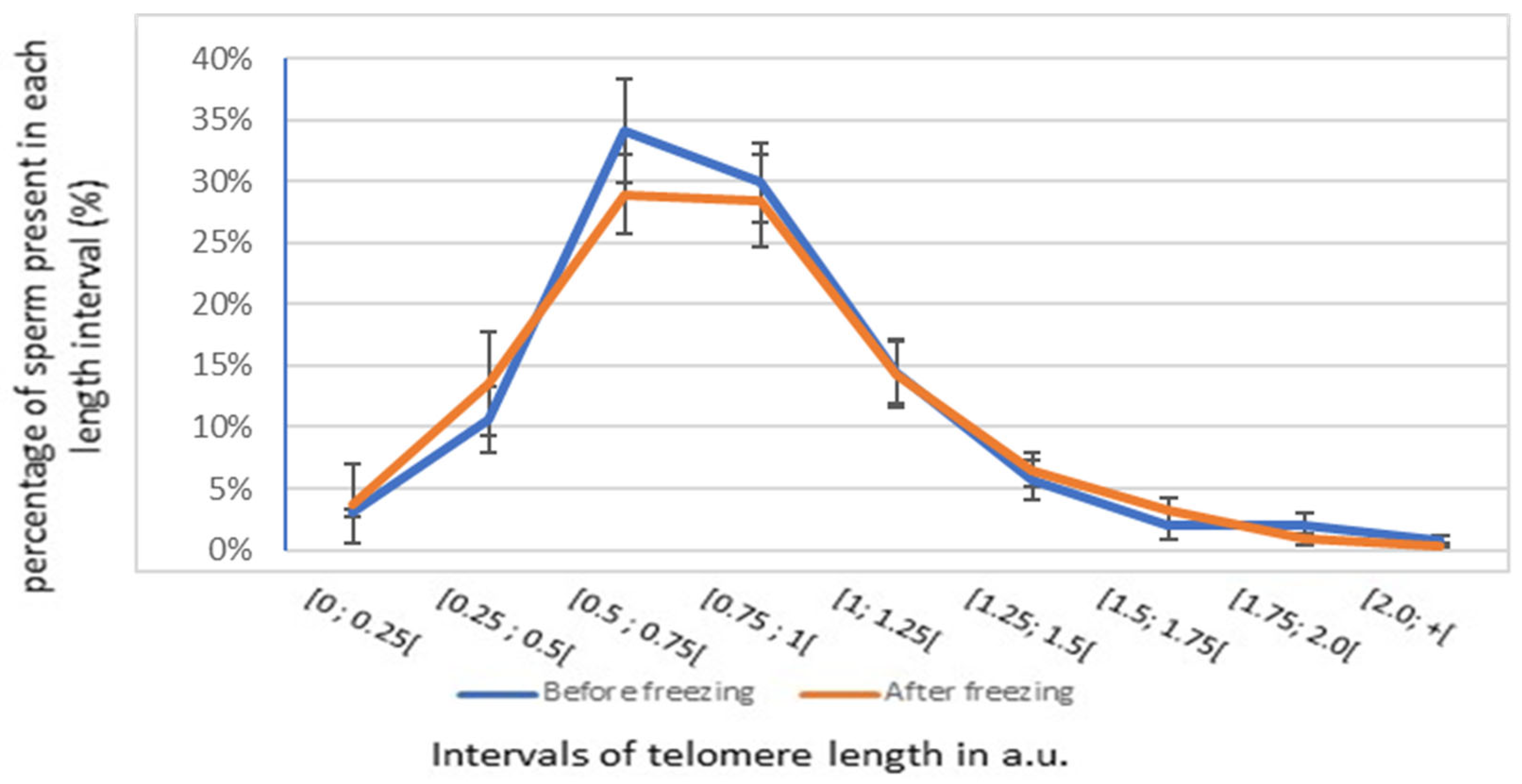
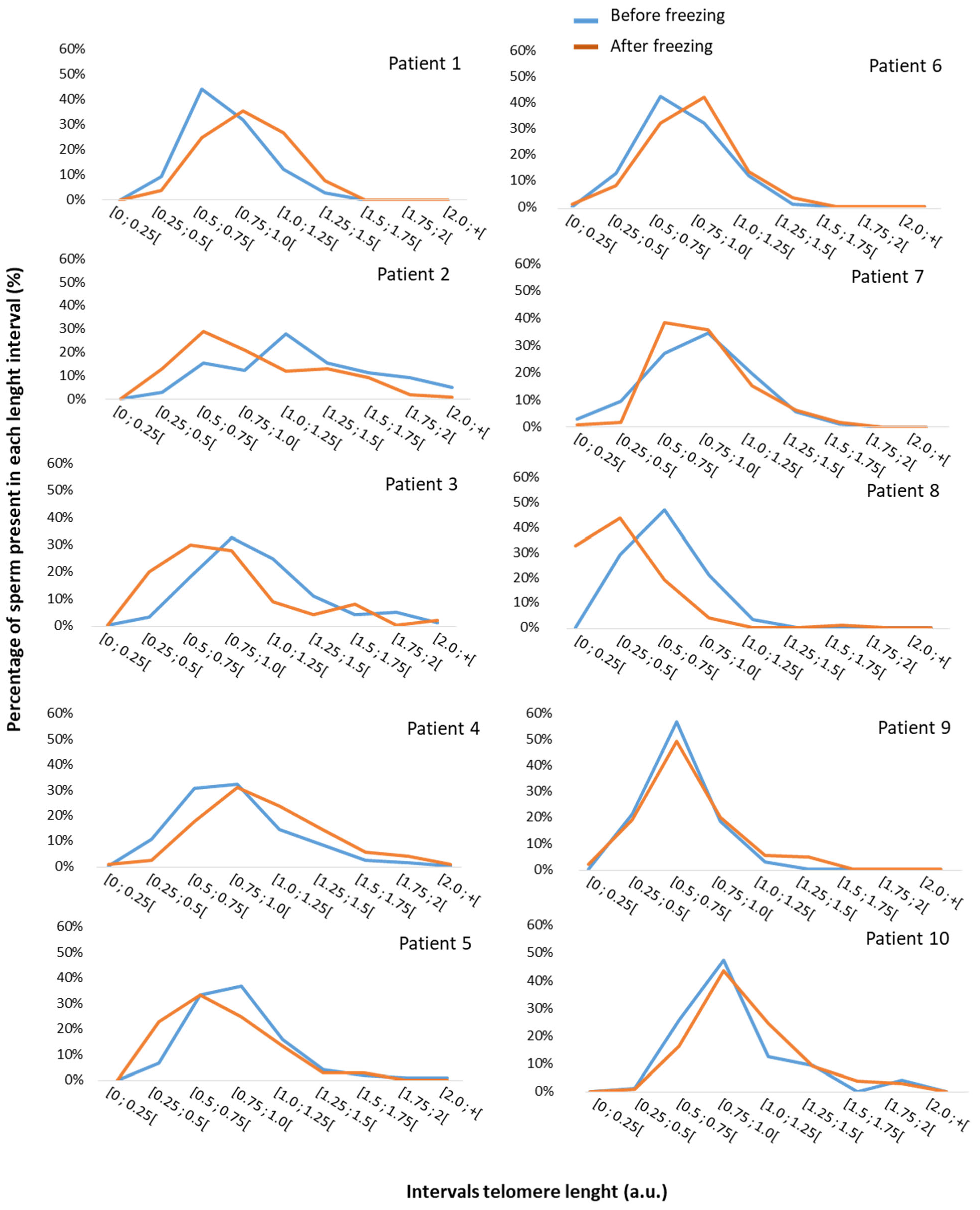
3.2. Impact of Freezing on Sperm Parameters and STL
3.3. Relationship between Sperm DNA Oxidation, DNA Fragmentation, and STL in Frozen Sperm Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samassekou, O.; Gadji, M.; Drouin, R.; Yan, J. Sizing the Ends: Normal Length of Human Telomeres. Ann. Anat. Anat. Anz. 2010, 192, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; O’Connor, M.S.; Qin, J.; Songyang, Z. Telosome, a Mammalian Telomere-Associated Complex Formed by Multiple Telomeric Proteins. J. Biol. Chem. 2004, 279, 51338–51342. [Google Scholar] [CrossRef] [PubMed]
- de Lange, T. How Telomeres Solve the End-Protection Problem. Science 2009, 326, 948. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, S. Telomerase Activity and Telomere Length in Male Germ Cells. Biol. Reprod. 2015, 92, 53. [Google Scholar] [CrossRef]
- Jørgensen, P.B.; Fedder, J.; Koelvraa, S.; Graakjaer, J. Age-Dependence of Relative Telomere Length Profiles during Spermatogenesis in Man. Maturitas 2013, 75, 380–385. [Google Scholar] [CrossRef]
- Thilagavathi, J.; Kumar, M.; Mishra, S.S.; Venkatesh, S.; Kumar, R.; Dada, R. Analysis of Sperm Telomere Length in Men with Idiopathic Infertility. Arch. Gynecol. Obstet. 2013, 287, 803–807. [Google Scholar] [CrossRef]
- Darmishonnejad, Z.; Tavalaee, M.; Izadi, T.; Tanhaei, S.; Nasr-Esfahani, M.H. Evaluation of Sperm Telomere Length in Infertile Men with Failed/Low Fertilization after Intracytoplasmic Sperm Injection. Reprod. BioMed. Online 2019, 38, 579–587. [Google Scholar] [CrossRef]
- Tahamtan, S.; Tavalaee, M.; Izadi, T.; Barikrow, N.; Zakeri, Z.; Lockshin, R.A.; Abbasi, H.; Esfahani, M.H.N. Reduced Sperm Telomere Length in Individuals with Varicocele Is Associated with Reduced Genomic Integrity. Sci. Rep. 2019, 9, 4336. [Google Scholar] [CrossRef]
- Berneau, S.C.; Shackleton, J.; Nevin, C.; Altakroni, B.; Papadopoulos, G.; Horne, G.; Brison, D.R.; Murgatroyd, C.; Povey, A.C.; Carroll, M. Associations of Sperm Telomere Length with Semen Parameters, Clinical Outcomes and Lifestyle Factors in Human Normozoospermic Samples. Andrology 2020, 8, 583–593. [Google Scholar] [CrossRef]
- Yang, Q.; Zhao, F.; Dai, S.; Zhang, N.; Zhao, W.; Bai, R.; Sun, Y. Sperm Telomere Length Is Positively Associated with the Quality of Early Embryonic Development. Hum. Reprod. 2015, 30, 1876–1881. [Google Scholar] [CrossRef]
- Ferlin, A.; Rampazzo, E.; Rocca, M.S.; Keppel, S.; Frigo, A.C.; De Rossi, A.; Foresta, C. In Young Men Sperm Telomere Length Is Related to Sperm Number and Parental Age. Hum. Reprod. 2013, 28, 3370–3376. [Google Scholar] [CrossRef] [PubMed]
- Cariati, F.; Jaroudi, S.; Alfarawati, S.; Raberi, A.; Alviggi, C.; Pivonello, R.; Wells, D. Investigation of Sperm Telomere Length as a Potential Marker of Paternal Genome Integrity and Semen Quality. Reprod. BioMed. Online 2016, 33, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Fattet, A.-J.; Chaillot, M.; Koscinski, I. Telomere Length, a New Biomarker of Male (in)Fertility? A Systematic Review of the Literature. Genes 2023, 14, 425. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Tan, Y.; Qiu, X.; Luo, H.; Li, Y.; Li, R.; Yang, X. Sperm Telomere Length as a Novel Biomarker of Male Infertility and Embryonic Development: A Systematic Review and Meta-Analysis. Front. Endocrinol. 2022, 13, 1079966. [Google Scholar] [CrossRef]
- Moskovtsev, S.I.; Willis, J.; White, J.; Mullen, J.B.M. Disruption of Telomere-Telomere Interactions Associated with DNA Damage in Human Spermatozoa. Syst. Biol. Reprod. Med. 2010, 56, 407–412. [Google Scholar] [CrossRef]
- Rocca, M.S.; Speltra, E.; Menegazzo, M.; Garolla, A.; Foresta, C.; Ferlin, A. Sperm Telomere Length as a Parameter of Sperm Quality in Normozoospermic Men. Hum. Reprod. 2016, 31, 1158–1163. [Google Scholar] [CrossRef]
- Lopes, A.C.; Oliveira, P.F.; Pinto, S.; Almeida, C.; Pinho, M.J.; Sá, R.; Rocha, E.; Barros, A.; Sousa, M. Discordance between Human Sperm Quality and Telomere Length Following Differential Gradient Separation/Swim-Up. J. Assist. Reprod. Genet. 2020, 37, 2581–2603. [Google Scholar] [CrossRef]
- Mishra, S.; Kumar, R.; Malhotra, N.; Singh, N.; Dada, R. Mild Oxidative Stress Is Beneficial for Sperm Telomere Length Maintenance. World J. Methodol. 2016, 6, 163–170. [Google Scholar] [CrossRef]
- Zhao, F.; Yang, Q.; Shi, S.; Luo, X.; Sun, Y. Semen Preparation Methods and Sperm Telomere Length: Density Gradient Centrifugation versus the Swim up Procedure. Sci. Rep. 2016, 6, 39051. [Google Scholar] [CrossRef]
- Kawanishi, S.; Oikawa, S. Mechanism of Telomere Shortening by Oxidative Stress. Ann. N. Y. Acad. Sci. 2004, 1019, 278–284. [Google Scholar] [CrossRef]
- Bui, A.D.; Sharma, R.; Henkel, R.; Agarwal, A. Reactive Oxygen Species Impact on Sperm DNA and Its Role in Male Infertility. Andrologia 2018, 50, e13012. [Google Scholar] [CrossRef] [PubMed]
- Cawthon, R.M. Telomere Measurement by Quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Cherkas, L.F.; Kato, B.S.; Demissie, S.; Hjelmborg, J.B.; Brimacombe, M.; Cupples, A.; Hunkin, J.L.; Gardner, J.P.; Lu, X.; et al. Offspring’s Leukocyte Telomere Length, Paternal Age, and Telomere Elongation in Sperm. PLoS Genet. 2008, 4, e37. [Google Scholar] [CrossRef] [PubMed]
- Perrin, J.; Fauque, P.; Paci, M.; Pons, H.; Brugnon, F. Cryoconservation of gametes: How to perform? Rev. Prat. 2018, 68, 886–890. [Google Scholar] [PubMed]
- Zribi, N.; Chakroun, N.F.; El Euch, H.; Gargouri, J.; Bahloul, A.; Keskes, L.A. Effects of Cryopreservation on Human Sperm Deoxyribonucleic Acid Integrity. Fertil. Steril. 2010, 93, 159–166. [Google Scholar] [CrossRef]
- Donnelly, E.T.; McClure, N.; Lewis, S.E.M. Cryopreservation of Human Semen and Prepared Sperm: Effects on Motility Parameters and DNA Integrity. Fertil. Steril. 2001, 76, 892–900. [Google Scholar] [CrossRef]
- WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization (Ed.) World Health Organization: Geneva, Switzerland, 2010; ISBN 978-92-4-154778-9. [Google Scholar]
- Kruger, T.F.; Ackerman, S.B.; Simmons, K.F.; Swanson, R.J.; Brugo, S.S.; Acosta, A.A. A Quick, Reliable Staining Technique for Human Sperm Morphology. Arch. Androl. 1987, 18, 275–277. [Google Scholar] [CrossRef]
- Mestres, S.; Pons-Rejraji, H.; Pereira, B.; Bouche, C.; Vega, A.; Chaput, L.; Vorilhon, S.; Janny, L.; Brugnon, F. Sperm Cryostorage in a Dry Tank: An Accurate Alternative. Cryobiology 2020, 94, 32–39. [Google Scholar] [CrossRef]
- Vorilhon, S.; Brugnon, F.; Kocer, A.; Dollet, S.; Bourgne, C.; Berger, M.; Janny, L.; Pereira, B.; Aitken, R.J.; Moazamian, A.; et al. Accuracy of Human Sperm DNA Oxidation Quantification and Threshold Determination Using an 8-OHdG Immuno-Detection Assay. Hum. Reprod. 2018, 33, 553–562. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Routledge: New York, NY, USA, 1988; ISBN 978-0-203-77158-7. [Google Scholar]
- Altman, D.G. Practical Statistics for Medical Research; Chapman and Hall/CRC: New York, NY, USA, 1990; ISBN 978-0-429-25858-9. [Google Scholar]
- Bender, R.; Lange, S. Adjusting for Multiple Testing—When and How? J. Clin. Epidemiol. 2001, 54, 343–349. [Google Scholar] [CrossRef]
- Scherthan, H. Telomeres and Meiosis in Health and Disease: Telomere Attachment and Clustering during Meiosis. Cell. Mol. Life Sci. 2007, 64, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cruz, R.; Roig, I.; Caldés, M.G. Maternal Origin of the Human Aneuploidies. Are Homolog Synapsis and Recombination to Blame? Notes (Learned) from the Underbelly. Meiosis 2009, 5, 128–136. [Google Scholar] [CrossRef]
- Eisenberg, D.T.A.; Kuzawa, C.W. The Paternal Age at Conception Effect on Offspring Telomere Length: Mechanistic, Comparative and Adaptive Perspectives. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20160442. [Google Scholar] [CrossRef] [PubMed]
- Chaput, L.; Grémeau, A.-S.; Vorilhon, S.; Pons, H.; Chabrot, C.; Grèze, V.; Pouly, J.-L.; Brugnon, F. Préservation de La Fertilité En Cancérologie. Bull. Cancer 2018, 105, 99–110. [Google Scholar] [CrossRef]
- Eustache, F. Missions and organization of the CECOS. Rev. Prat. 2018, 68, 203–206. [Google Scholar]
- Aubert, G.; Hills, M.; Lansdorp, P.M. Telomere Length Measurement-Caveats and a Critical Assessment of the Available Technologies and Tools. Mutat. Res. 2012, 730, 59–67. [Google Scholar] [CrossRef]
- Kurjanowicz, P.; Moskovtsev, S.; Librach, C. Genomic Fragmentation and Extrachromosomal Telomeric Repeats Impact Assessment of Telomere Length in Human Spermatozoa: Quantitative Experiments and Systematic Review. Hum. Reprod. 2017, 32, 2170–2177. [Google Scholar] [CrossRef]
- Raschenberger, J.; Lamina, C.; Haun, M.; Kollerits, B.; Coassin, S.; Boes, E.; Kedenko, L.; Köttgen, A.; Kronenberg, F. Influence of DNA Extraction Methods on Relative Telomere Length Measurements and Its Impact on Epidemiological Studies. Sci. Rep. 2016, 6, 25398. [Google Scholar] [CrossRef]
- Thomson, L.K.; Fleming, S.D.; Aitken, R.J.; De Iuliis, G.N.; Zieschang, J.-A.; Clark, A.M. Cryopreservation-Induced Human Sperm DNA Damage Is Predominantly Mediated by Oxidative Stress Rather than Apoptosis. Hum. Reprod. 2009, 24, 2061–2070. [Google Scholar] [CrossRef]
- Berby, B.; Bichara, C.; Rives-Feraille, A.; Jumeau, F.; Pizio, P.D.; Sétif, V.; Sibert, L.; Dumont, L.; Rondanino, C.; Rives, N. Oxidative Stress Is Associated with Telomere Interaction Impairment and Chromatin Condensation Defects in Spermatozoa of Infertile Males. Antioxidants 2021, 10, 593. [Google Scholar] [CrossRef]
- Miller, A.S.; Balakrishnan, L.; Buncher, N.A.; Opresko, P.L.; Bambara, R.A. Telomere Proteins POT1, TRF1 and TRF2 Augment Long-Patch Base Excision Repair in Vitro. Cell Cycle 2012, 11, 998–1007. [Google Scholar] [CrossRef] [PubMed]
- Sfeir, A.; Kosiyatrakul, S.T.; Hockemeyer, D.; MacRae, S.L.; Karlseder, J.; Schildkraut, C.L.; de Lange, T. Mammalian Telomeres Resemble Fragile Sites and Require TRF1 for Efficient Replication. Cell 2009, 138, 90–103. [Google Scholar] [CrossRef]
- Hezavehei, M.; Sharafi, M.; Kouchesfahani, H.M.; Henkel, R.; Agarwal, A.; Esmaeili, V.; Shahverdi, A. Sperm Cryopreservation: A Review on Current Molecular Cryobiology and Advanced Approaches. Reprod. BioMed. Online 2018, 37, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Hartshorne, G.M. Telomere Lengths in Human Pronuclei, Oocytes and Spermatozoa. Mol. Hum. Reprod. 2013, 19, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Sharqawi, M.; Hantisteanu, S.; Bilgory, A.; Aslih, N.; Shibli Abu Raya, Y.; Atzmon, Y.; Estrada, D.; Limonad, O.; Meisel-Sharon, S.; Shalom-Paz, E. The Impact of Lifestyle on Sperm Function, Telomere Length, and IVF Outcomes. Am. J. Men’s Health 2022, 16, 15579883221119932. [Google Scholar] [CrossRef]
- Torra-Massana, M.; Barragán, M.; Bellu, E.; Oliva, R.; Rodríguez, A.; Vassena, R. Sperm Telomere Length in Donor Samples Is Not Related to ICSI Outcome. J. Assist. Reprod. Genet. 2018, 35, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Aston, K.I.; Hunt, S.C.; Susser, E.; Kimura, M.; Factor-Litvak, P.; Carrell, D.; Aviv, A. Divergence of Sperm and Leukocyte Age-Dependent Telomere Dynamics: Implications for Male-Driven Evolution of Telomere Length in Humans. Mol. Hum. Reprod. 2012, 18, 517–522. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Mean ± SEM |
|---|---|
| Age (years, n = 30) | 32.8 ± 1 |
| BMI (kg/m2, n = 21) | 27 ± 1 |
| Sperm volume (mL, n = 30) | 4.7 ± 0.4 |
| Sperm concentration (M/mL, n = 30) | 108 ± 14.2 |
| PMN concentration (M/mL, n = 30) | 0.12 ± 0.06 |
| Progressive motility (%, n = 30) Total motility (%, n = 30) | 48.5 ± 2.4 54.8 ± 2.5 |
| Sperm vitality (%, n = 30) | 73.1 ± 2.7 |
| Typical forms (%, n = 30) | 6.7 ± 1 |
| STL (a.u., n = 30) | 3.2 ± 0.2 |
| Parameters | After Freezing-Thawing (Mean ± SEM) | Effect-Size [95% CI] |
|---|---|---|
| Progressive motility (%, n = 30) | 12.6 ± 2.4 | −2.8 [−3.5; −2.0] |
| Total motility (%, n = 30) | 19.9 ± 2.4 | −2.6 [−3.2; −1.9] |
| Sperm vitality (%, n = 30) | 37.8 ± 3.8 | −1.9 [−2.6; −1.3] |
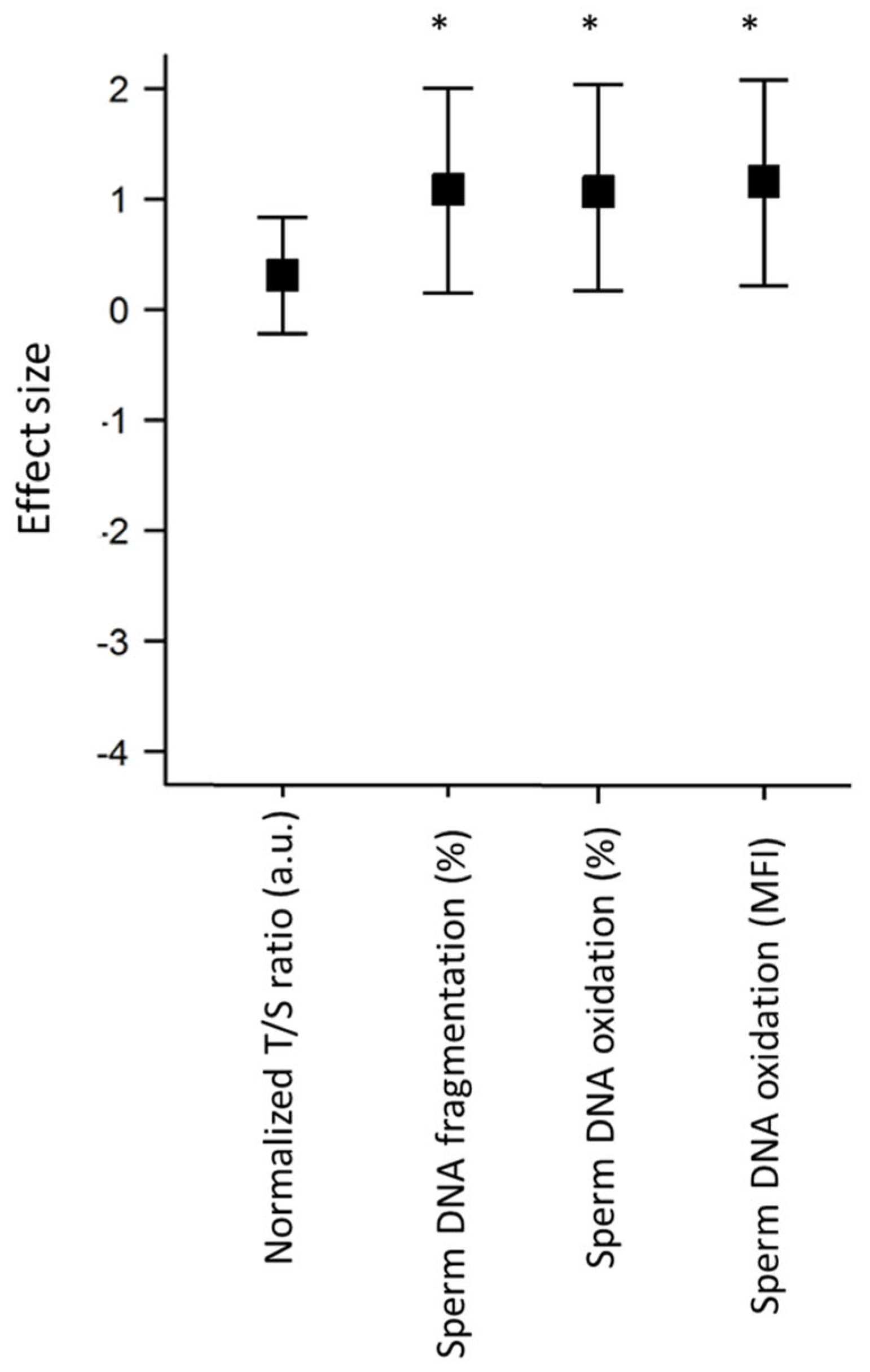
| STL (a.u., n = 30) | 3.3 ± 0.2 | 0.08 [−0.4; 0.6] |
| Parameters | Fresh (Mean ± SEM) | After Freezing-Thawing (Mean ± SEM) | Effect-Size [CI] |
|---|---|---|---|
| Sperm DNA fragmentation (%, n = 10) | 23.8 ± 5.3 | 38.8 ± 5.7 * | 0.8 [−0.05; 1.8] |
| Sperm DNA oxidation (%, n = 10) | 73.2 ± 4.7 | 85.1 ± 3.7 * | 0.8 [−0.07; 1.7] |
| Sperm DNA oxidation (MFI, a.u., n = 10) | 839.2 ± 210.6 | 1445.8 ± 202.1 * | 0.9 [−0.01; 1.8] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gouhier, C.; Pons-Rejraji, H.; Dollet, S.; Chaput, L.; Bourgne, C.; Berger, M.; Pereira, B.; Tchirkov, A.; Brugnon, F. Freezing Does Not Alter Sperm Telomere Length despite Increasing DNA Oxidation and Fragmentation. Genes 2023, 14, 1039. https://doi.org/10.3390/genes14051039
Gouhier C, Pons-Rejraji H, Dollet S, Chaput L, Bourgne C, Berger M, Pereira B, Tchirkov A, Brugnon F. Freezing Does Not Alter Sperm Telomere Length despite Increasing DNA Oxidation and Fragmentation. Genes. 2023; 14(5):1039. https://doi.org/10.3390/genes14051039
Chicago/Turabian StyleGouhier, Charlène, Hanae Pons-Rejraji, Sandra Dollet, Laure Chaput, Céline Bourgne, Marc Berger, Bruno Pereira, Andrei Tchirkov, and Florence Brugnon. 2023. "Freezing Does Not Alter Sperm Telomere Length despite Increasing DNA Oxidation and Fragmentation" Genes 14, no. 5: 1039. https://doi.org/10.3390/genes14051039