

Autophagy-Related MicroRNA: Tumor miR-125b and Thyroid Cancers

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
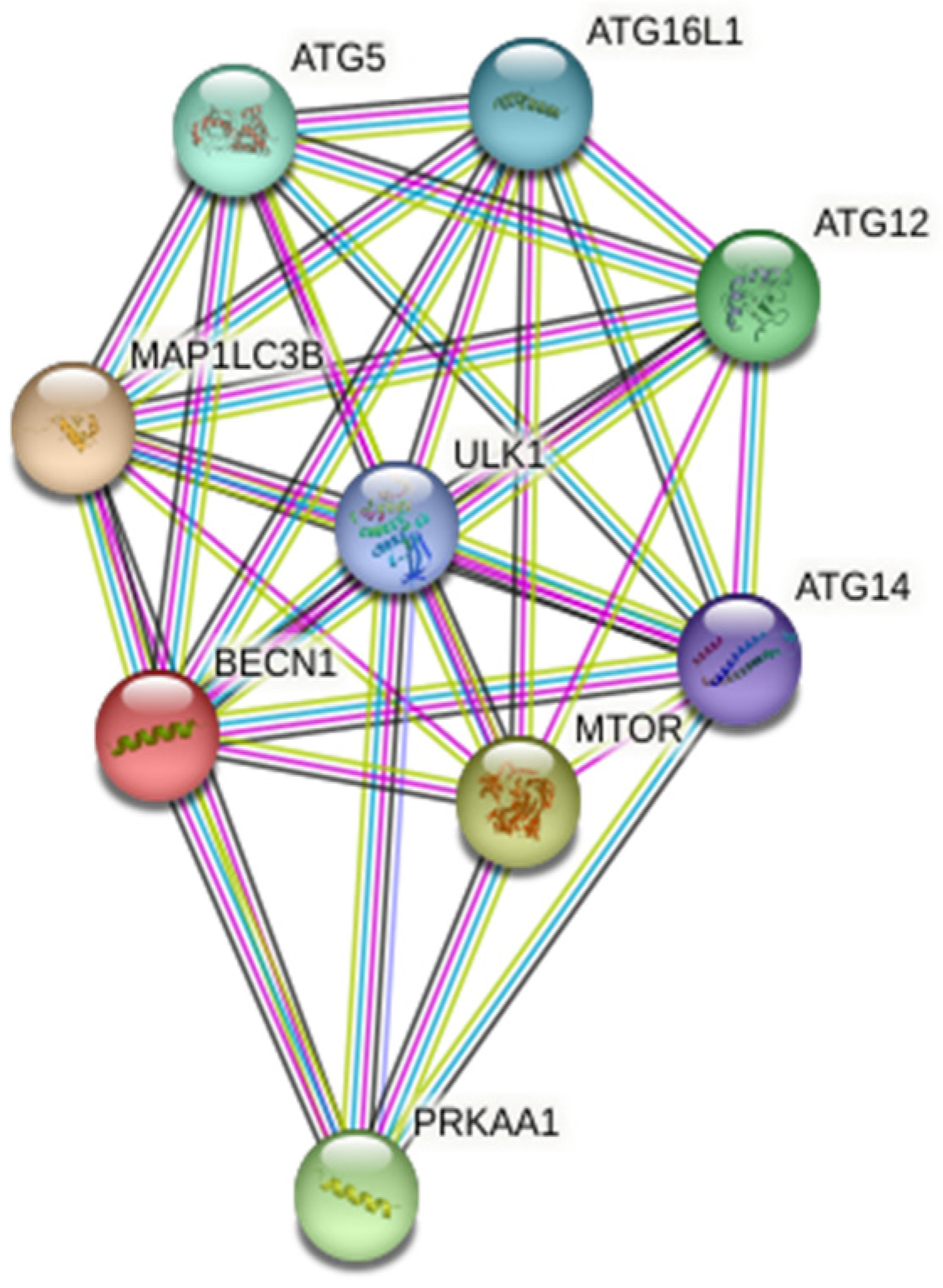
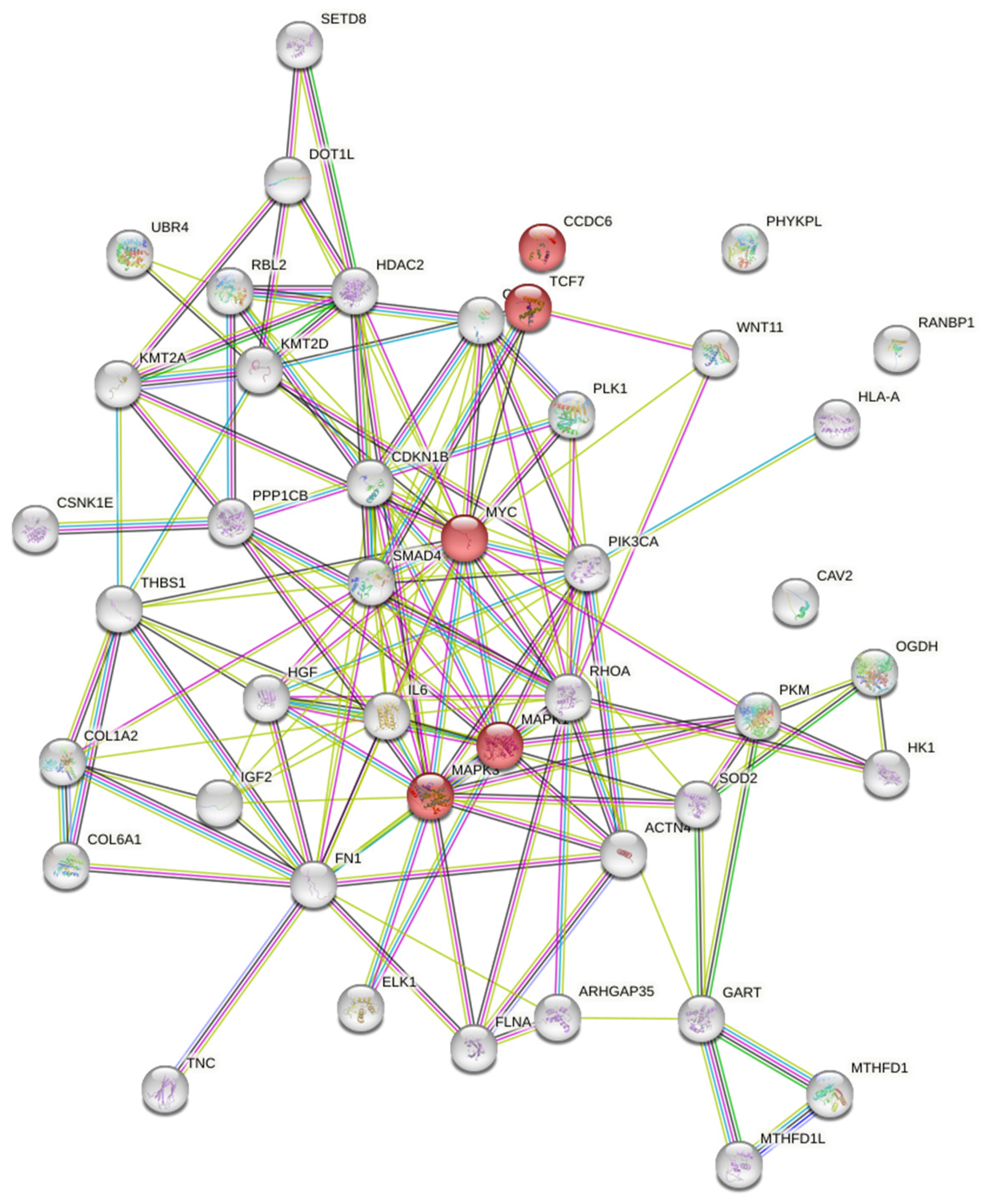
2.1. Protein-Protein Interaction (PPI) Analysis
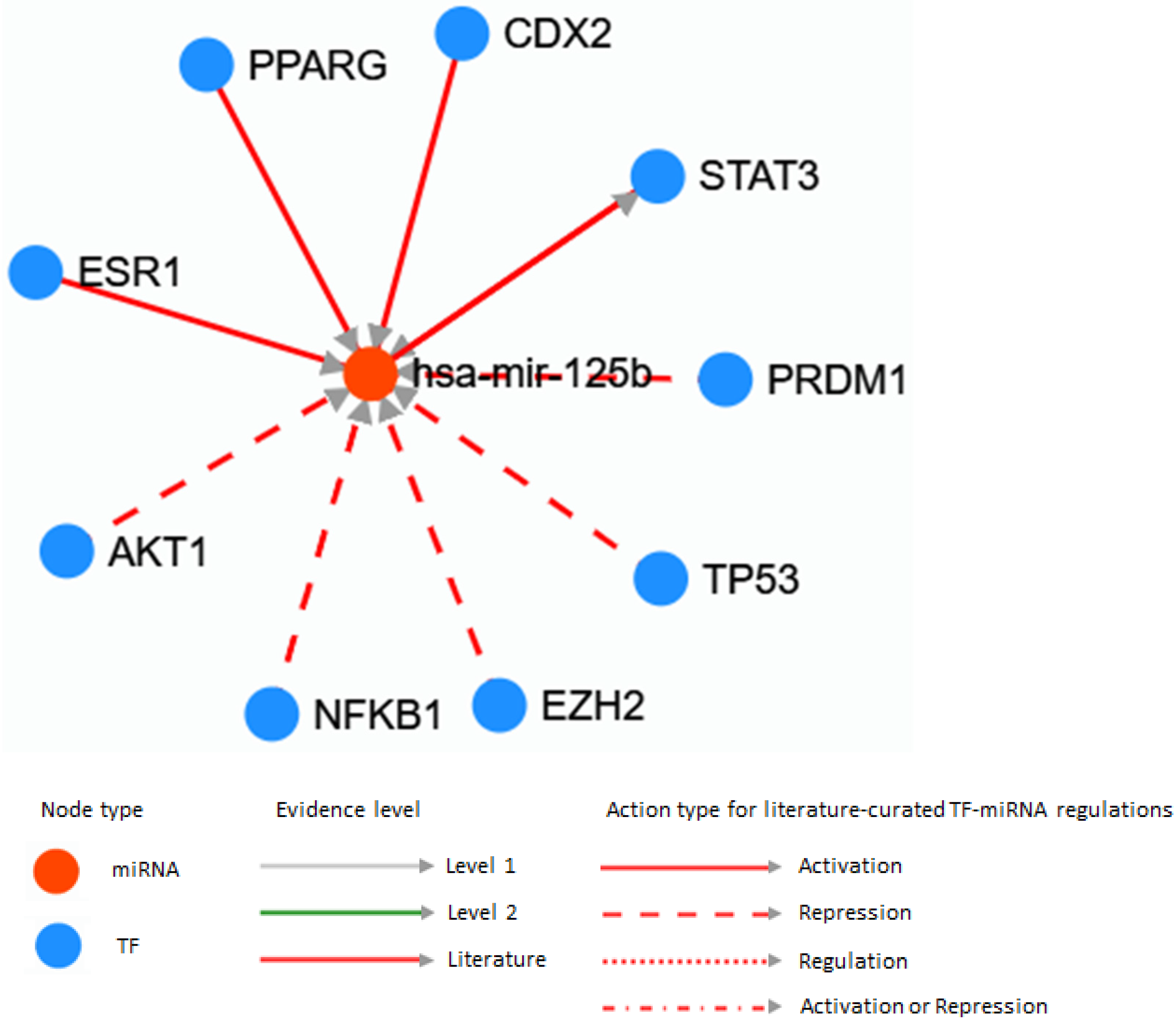
2.2. MicroRNA and Genes Associations
2.3. Clinical and Experimental Tools
2.4. DNA Extraction
2.5. BRAFV600E Mutation Analysis
2.6. Quantitative PCR with Reverse Transcription in Real Time
3. Results
3.1. Investigation of MicroRNAs Related to Autophagy
3.1.1. Investigation of MicroRNAs Regulating Proteins Associated with Autophagy
3.1.2. MicroRNAs Associated with the Thyroid Cancers
3.1.3. Targets and Signaling Cascades Associated with MicroRNA 125
3.2. MicroRNA 125b Expression in Thyroid Pathologies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oh, J.M.; Ahn, B.C. Molecular mechanisms of radioactive iodine refractoriness in differentiated thyroid cancer: Impaired sodium iodide symporter (NIS) expression owing to altered signaling pathway activity and intracellular localization of NIS. Theranostics 2021, 1, 6251–6277. [Google Scholar] [CrossRef]
- Holm, T.M.; Yeo, S.; Turner, K.M.; Guan, J.L. Targeting Autophagy in Thyroid Cancer: EMT, Apoptosis, and Cancer Stem Cells. Front. Cell Dev. Biol. 2022, 10, 821855. [Google Scholar] [CrossRef]
- Jiang, S.; Huang, Y.; Li, Y.; Gu, Q.; Jiang, C.; Tao, X.; Sun, J. Silencing FOXP2 reverses vemurafenib resistance in BRAFV600E mutant papillary thyroid cancer and melanoma cells. Endocrine 2022, 79, 86–97. [Google Scholar] [CrossRef]
- Kuo, C.Y.; Jhuang, J.Y.; Huang, W.C.; Cheng, S.P. Aberrant Expression of Thymosin Beta-4 Correlates with Advanced Disease and BRAF V600E Mutation in Thyroid Cancer. J. Histochem. Cytochem. 2022, 2, 221554221138370. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Russell, R.C.; Tian, Y.; Yuan, H.; Park, H.W.; Chang, Y.Y.; Kim, J.; Kim, H.; Neufeld, T.P.; Dillin, A.; Guan, K.L. ULK1 induces autophagy by phosphorylating Beclin-1 and activating VPS34 lipid kinase. Nat. Cell Biol. 2013, 15, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Chen, J.; Xu, H.G.; Zhou, X.; He, Q.; Li, Y.L.; Jiang, G.; Shan, Y.; Xue, B.; Zhao, R.X.; et al. MIR106B and MIR93 prevent removal of bacteria from epithelial cells by disrupting ATG16L1-mediated autophagy. Gastroenterology 2014, 146, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Stripay, J.L.; Zhang, X.; Collage, R.D.; Hulver, M.; Carchman, E.H.; Howell, G.M.; Zuckerbraun, B.S.; Lee, J.S.; Rosengart, M.R. CaMKIα regulates AMP kinase-dependent, TORC-1-independent autophagy during lipopolysaccharide-induced acute lung neutrophilic inflammation. J. Immunol. 2013, 190, 3620–3628. [Google Scholar] [CrossRef] [Green Version]
- Spirina, L.V.; Avgustinovich, A.V.; Afanas'ev, S.G.; Cheremisina, O.V.; Volkov, M.Y.; Choynzonov, E.L.; Gorbunov, A.K.; Usynin, E.A. Molecular Mechanism of Resistance to Chemotherapy in Gastric Cancers, the Role of Autophagy. Curr. Drug Targets 2020, 21, 713–721. [Google Scholar] [CrossRef]
- Sun, Y.M.; Lin, K.Y.; Chen, Y.Q. Diverse functions of miR-125 family in different cell contexts. J. Hematol. Oncol. 2013, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Peng, B.; Theng, P.Y.; Le, M.T.N. Essential functions of miR-125b in cancer. Cell Prolif. 2021, 54, 12913. [Google Scholar] [CrossRef]
- Bu, Q.; You, F.; Pan, G.; Yuan, Q.; Cui, T.; Hao, L.; Zhang, J. MiR-125b inhibits anaplastic thyroid cancer cell migration and invasion by targeting PIK3CD. Biomed. Pharmacother. 2017, 88, 443–448. [Google Scholar] [CrossRef]
- Zhang, G.; Zhou, S.; Yang, Q.; Liu, F. MicroRNA-125b reduces glucose uptake in papillary thyroid carcinoma cells. Oncol. Lett. 2020, 20, 2806–2810. [Google Scholar] [CrossRef]
- Lowery, A.J.; Miller, N.; McNeill, R.E.; Kerin, M.J. MicroRNAs as prognostic indicators and therapeutic targets: Potential effect on breast cancer management. Clin. Cancer Res. 2008, 14, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Murphy, A.J.; Guyre, P.M.; Pioli, P.A. Estradiol suppresses NF-kappa B activation through coordinated regulation of let-7a and miR-125b in primary human macrophages. J. Immunol. 2010, 184, 5029–5037. [Google Scholar] [CrossRef] [Green Version]
- Ghafouri-Fard, S.; Shirvani-Farsani, Z.; Taheri, M. The role of microRNAs in the pathogenesis of thyroid cancer. Noncoding RNA Res. 2020, 5, 88–98. [Google Scholar] [CrossRef]
- Sánchez-Sendra, B.; González-Muñoz, J.F.; Pérez-Debén, S.; Monteagudo, C. The Prognostic Value of miR-125b, miR-200c and miR-205 in Primary Cutaneous Malignant Melanoma Is Independent of BRAF Mutational Status. Cancers 2022, 14, 1532. [Google Scholar] [CrossRef]
- Wang, S.; Wu, J.; Ren, J.; Vlantis, A.C.; Li, M.Y.; Liu, S.Y.W.; Ng, E.K.W.; Chan, A.B.W.; Luo, D.C.; Liu, Z.; et al. MicroRNA-125b Interacts with Foxp3 to Induce Autophagy in Thyroid Cancer. Mol. Ther. 2018, 26, 2295–2303. [Google Scholar] [CrossRef] [Green Version]
- Plantinga, T.S.; van de Vosse, E.; Huijbers, A.; Netea, M.G.; Joosten, L.A.; Smit, J.W.; Netea-Maier, R.T. Role of genetic variants of autophagy genes in susceptibility for non-medullary thyroid cancer and patients outcome. PLoS ONE 2014, 9, 94086. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; P., L.; Wen, D.; Gao, L.; Liang, L.; Luo, Y.; Wei, Y.; Yu, H.; Yang, H.; Ma, W. Expression level of miR-146b-5p via miRNA sequencing and its potential targets in papillary thyroid cancer. Int. J. Clin. Exp. Med. 2018, 11, 17. [Google Scholar]
- Czajka, A.A.; Wójcicka, A.; Kubiak, A.; Kotlarek, M.; Bakuła-Zalewska, E.; Koperski, Ł.; Wiechno, W.; Jażdżewski, K. Family of microRNA-146 Regulates RARβ in Papillary Thyroid Carcinoma. PLoS ONE 2016, 11, 0151968. [Google Scholar] [CrossRef]
- Ramírez-Moya, J.; Wert-Lamas, L.; Riesco-Eizaguirre, G.; Santisteban, P. Impaired microRNA processing by DICER1 downregulation endows thyroid cancer with increased aggressiveness. Oncogene 2019, 38, 5486–5499. [Google Scholar] [CrossRef]
- Cong, D.; He, M.; Chen, S.; Liu, X.; Liu, X.; Sun, H. Expression profiles of pivotal microRNAs and targets in thyroid papillary carcinoma: An analysis of The Cancer Genome Atlas. OncoTargets Ther. 2015, 8, 2271–2277. [Google Scholar]
- Rosignolo, F.; Memeo, L.; Monzani, F.; Colarossi, C.; Pecce, V.; Verrienti, A.; Durante, C.; Grani, G.; Lamartina, L.; Forte, S.; et al. MicroRNA-based molecular classification of papillary thyroid carcinoma. Int. J. Oncol. 2017, 50, 1767–1777. [Google Scholar] [CrossRef] [Green Version]
- Perdas, E.; Stawski, R.; Kaczka, K.; Zubrzycka, M. Analysis of Let-7 Family miRNA in Plasma as Potential Predictive Biomarkers of Diagnosis for Papillary Thyroid Cancer. Diagnostics 2020, 10, 130. [Google Scholar] [CrossRef] [Green Version]
- Shabani, N.; Razaviyan, J.; Paryan, M.; Tavangar, S.M.; Azizi, F.; Mohammadi-Yeganeh, S.; Hedayati, M. Evaluation of miRNAs expression in medullary thyroid carcinoma tissue samples: miR-34a and miR-144 as promising overexpressed markers in MTC. Hum. Pathol. 2018, 79, 212–221. [Google Scholar] [CrossRef]
- Qiu, Z.; Li, H.; Wang, J.; Sun, C. miR-146a and miR-146b in the diagnosis and prognosis of papillary thyroid carcinoma. Oncol. Rep. 2017, 38, 2735–2740. [Google Scholar] [CrossRef] [Green Version]
- Pamedytyte, D.; Simanaviciene, V.; Dauksiene, D.; Leipute, E.; Zvirbliene, A.; Sarauskas, V.; Dauksa, A.; Verkauskiene, R.; Zilaitiene, B. Association of MicroRNA Expression and BRAFV600E Mutation with Recurrence of Thyroid Cancer. Biomolecules 2020, 10, 625. [Google Scholar] [CrossRef] [Green Version]
- Rezaei, M.; Khamaneh, A.M.; Zarghami, N.; Vosoughi, A.; Hashemzadeh, S. Evaluating pre- and post-operation plasma miRNAs of papillary thyroid carcinoma (PTC) patients in comparison to benign nodules. BMC Cancer 2019, 19, 690. [Google Scholar] [CrossRef] [Green Version]
- Le, F.; Luo, P.; Yang, Q.O.; Zhong, X.M. MiR-181a promotes growth of thyroid cancer cells by targeting tumor suppressor RB1. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5638–5647. [Google Scholar]
- Liu, Y.; Zhang, B.; Shi, T.; Qin, H. miR-182 promotes tumor growth and increases chemoresistance of human anaplastic thyroid cancer by targeting tripartite motif 8. OncoTargets Ther. 2017, 10, 1115–1122. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, G.; Dolcimascolo, A.; Torrisi, F.; Zappalà, A.; Gulino, R.; Parenti, R. MiR-19a Overexpression in FTC-133 Cell Line Induces a More De-Differentiated and Aggressive Phenotype. Int. J. Mol. Sci. 2018, 19, 3944. [Google Scholar] [CrossRef] [Green Version]
- Vosgha, H.; Ariana, A.; Smith, R.A.; Lam, A.K. miR-205 targets angiogenesis and EMT concurrently in anaplastic thyroid carcinoma. Endocr. Relat. Cancer 2018, 25, 323–337. [Google Scholar] [CrossRef] [Green Version]
- Diao, Y.; Fu, H.; Wang, Q. MiR-221 Exacerbate Cell Proliferation and Invasion by Targeting TIMP3 in Papillary Thyroid Carcinoma. Am. J. Ther. 2017, 24, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.L.; Gao, A.B.; Wang, Q.; Lou, X.E.; Zhao, J.; Lu, Q.J. MicroRNA-221 promotes papillary thyroid carcinoma cell migration and invasion via targeting RECK and regulating epithelial-mesenchymal transition. OncoTargets Ther. 2019, 12, 2323–2333. [Google Scholar] [CrossRef] [Green Version]
- Mazeh, H.; Deutch, T.; Karas, A.; Bogardus, K.A.; Mizrahi, I.; Gur-Wahnon, D.; Ben-Dov, I.Z. Next-Generation Sequencing Identifies a Highly Accurate miRNA Panel That Distinguishes Well-Differentiated Thyroid Cancer from Benign Thyroid Nodules. Cancer Epidemiol. Biomark. Prev. 2018, 27, 858–863. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Lin, L.; Li, W.; Lu, G.; Li, X. MiR-223 inhibitor suppresses proliferation and induces apoptosis of thyroid cancer cells by down-regulating aquaporin-1. J. Recept. Signal Transduct. Res. 2019, 39, 146–153. [Google Scholar] [CrossRef]
- Wen, Q.; Zhang, D.; Wang, T.; Wang, T.; Zhao, M.; Xie, Q.; Fu, Y.; Ma, Q. MiR-23a promotes cell proliferation and invasion in papillary thyroid carcinoma by targeting PTEN. Int. J. Clin. Exp. Pathol. 2016, 9, 2366–2373. [Google Scholar]
- Zhao, P.; Ma, W.; Hu, Z.; Zhang, Y.; Zhang, S.; Wang, Y. Up-regulation of miR-340-5p promotes progression of thyroid cancer by inhibiting BMP4. J. Endocrinol. Investig. 2018, 41, 1165–1172. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Zhao, R.; Zhao, Q.; Zhang, T. Upregulated miR-9-3p Promotes Cell Growth and Inhibits Apoptosis in Medullary Thyroid Carcinoma by Targeting BLCAP. Oncol. Res. 2017, 25, 1215–1222. [Google Scholar] [CrossRef]
- Mancikova, V.; Castelblanco, E.; Pineiro-Yanez, E.; Perales-Paton, J.; de Cubas, A.A.; Inglada-Perez, L.; Matias-Guiu, X.; Capel, I.; Bella, M.; Lerma, E.; et al. MicroRNA deep-sequencing reveals master regulators of follicular and papillary thyroid tumors. Mod. Pathol. 2015, 28, 748–757. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhao, L.; Zhang, Z.; Zhang, H.; Ding, C.; Su, Z. Roles of microRNA let-7b in papillary thyroid carcinoma by regulating HMGA2. Tumour Biol. 2017, 39, 1010428317719274. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.Z.; Bin, H.; Zang, Z.N. MiR-128 suppresses the growth of thyroid carcinoma by negatively regulating SPHK1. Biomed. Pharmacother. 2019, 109, 1960–1966. [Google Scholar] [CrossRef]
- Gao, X.; Chen, Z.; Li, A.; Zhang, X.; Cai, X. MiR-129 regulates growth and invasion by targeting MAL2 in papillary thyroid carcinoma. Biomed. Pharmacother. 2018, 105, 1072–1078. [Google Scholar] [CrossRef]
- Chen, X.; Li, M.; Zhou, H.; Zhang, L. miR-132 Targets FOXA1 and Exerts Tumor-Suppressing Functions in Thyroid Cancer. Oncol. Res. 2019, 27, 431–437. [Google Scholar] [CrossRef]
- Montero-Conde, C.; Graña-Castro, O.; Martín-Serrano, G.; Martínez-Montes, Á.M.; Zarzuela, E.; Muñoz, J.; Torres-Perez, R.; Pita, G.; Cordero-Barreal, A.; Leandro-García, L.J.; et al. Hsa-miR-139-5p is a prognostic thyroid cancer marker involved in HNRNPF-mediated alternative splicing. Int. J. Cancer. 2020, 146, 521–530. [Google Scholar] [CrossRef]
- Sun, J.; Shi, R.; Zhao, S.; Li, X.; Lu, S.; Bu, H.; Ma, X.; Su, C. E2F8, a direct target of miR-144, promotes papillary thyroid cancer progression via regulating cell cycle. J. Exp. Clin. Cancer Res. 2017, 36, 40. [Google Scholar] [CrossRef] [Green Version]
- Joo, L.J.S.; Weiss, J.; Gill, A.J.; Clifton-Bligh, R.; Brahmbhatt, H.; MacDiarmid, J.A.; Gild, M.L.; Robinson, B.G.; Zhao, J.T.; Sidhu, S.B. RET Kinase-Regulated MicroRNA-153-3p Improves Therapeutic Efficacy in Medullary Thyroid Carcinoma. Thyroid 2019, 29, 830–844. [Google Scholar] [CrossRef]
- Liu, C.; Xing, M.; Wang, L.; Zhang, K. miR-199a-3p downregulation in thyroid tissues is associated with invasion and metastasis of papillary thyroid carcinoma. Br. J. Biomed. Sci. 2017, 74, 90–94. [Google Scholar] [CrossRef]
- Wu, X.; Dai, L.; Zhang, Z.; Zheng, J.; Zhao, J. Overexpression of microRNA-203 can downregulate survivin and function as a potential therapeutic target in papillary thyroid cancer. Oncol. Lett. 2020, 19, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, Q.; Li, N.; Zhang, S. miR-205 targets YAP1 and inhibits proliferation and invasion in thyroid cancer cells. Mol. Med. Rep. 2018, 18, 1674–1681. [Google Scholar] [CrossRef]
- Huang, Z.; Xing, S.; Liu, M.; Deng, W.; Wang, Y.; Huang, Z.; Huang, Y.; Huang, X.; Wu, C.; Guo, X.; et al. MiR-26a-5p enhances cells proliferation, invasion, and apoptosis resistance of fibroblast-like synoviocytes in rheumatoid arthritis by regulating PTEN/PI3K/AKT pathway. Biosci. Rep. 2019, 39, BSR20182192. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, X.L.; Li, X.F.; Tang, Y.C.; Zhao, X. miR-212-3p reduced proliferation, and promoted apoptosis of fibroblast-like synoviocytes via down-regulating SOX5 in rheumatoid arthritis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 461–471. [Google Scholar]
- Lin, Y.; Cheng, K.; Wang, T.; Xie, Q.; Chen, M.; Chen, Q.; Wen, Q. miR-217 inhibits proliferation, migration, and invasion via targeting AKT3 in thyroid cancer. Biomed. Pharmacother. 2017, 95, 1718–1724. [Google Scholar] [CrossRef]
- Yin, J.J.; Cheng, X.Y. MicroRNA-23a inhibits the growth of papillary thyroid carcinoma via regulating cyclin G1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3431–3439. [Google Scholar]
- Zhou, A.; Chen, G.; Cheng, X.; Zhang, C.; Xu, H.; Qi, M.; Chen, X.; Wang, T.; Li, L. Inhibitory effects of miR-26b-5p on thyroid cancer. Mol. Med. Rep. 2019, 20, 1196–1202. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Deng, H.; Zhao, Y.; Li, C.; Liang, Y. LncRNA XIST/miR-34a axis modulates the cell proliferation and tumor growth of thyroid cancer through MET-PI3K-AKT signaling. J. Exp. Clin. Cancer Res. 2018, 37, 279. [Google Scholar] [CrossRef]
- Wu, G.; Zheng, H.; Xu, J.; Guo, Y.; Zheng, G.; Ma, C.; Hao, S.; Liu, X.; Chen, H.; Wei, S.; et al. miR-429 suppresses cell growth and induces apoptosis of human thyroid cancer cell by targeting ZEB1. Artif. Cells Nanomed. Biotechnol. 2019, 47, 548–554. [Google Scholar] [CrossRef]
- Lassalle, S.; Zangari, J.; Popa, A.; Ilie, M.; Hofman, V.; Long, E.; Patey, M.; Tissier, F.; Belléannée, G.; Trouette, H.; et al. MicroRNA-375/SEC23A as biomarkers of the in vitro efficacy of vandetanib. Oncotarget 2016, 7, 30461–30478. [Google Scholar] [CrossRef] [Green Version]
- Xue, K.C.; Hu, D.D.; Zhao, L.; Li, N.; Shen, H.Y. MiR-577 inhibits papillary thyroid carcinoma cell proliferation, migration and invasion by targeting SphK2. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 3794–3800. [Google Scholar]
- Gu, Y.; Yang, N.; Yin, L.; Feng, C.; Liu, T. Inhibitory roles of miR-9 on papillary thyroid cancer through targeting BRAF. Mol. Med. Rep. 2018, 18, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Ding, C.; Yu, H.; Shi, C.; Shi, T.; Qin, H.; Cui, Y. MiR-let-7e inhibits invasion and magration and regulates HMGB1 expression in papillary thyroid carcinoma. Biomed. Pharmacother. 2019, 110, 528–536. [Google Scholar] [CrossRef]
- Ahmed, R.; Samanta, S.; Banerjee, J.; Kar, S.S.; Dash, S.K. Modulatory role of miRNAs in thyroid and breast cancer progression and insights into their therapeutic manipulation. Curr. Res. Pharmacol. Drug Discov. 2022, 3, 100131. [Google Scholar] [CrossRef]
- Lu, M.D.; Li, H.; Nie, J.H.; Li, S.; Ye, H.S.; Li, T.T.; Wu, M.L.; Liu, J. Dual Inhibition of BRAF-MAPK and STAT3 Signaling Pathways in Resveratrol-Suppressed Anaplastic Thyroid Cancer Cells with BRAF Mutations. Int. J. Mol. Sci. 2022, 23, 14385. [Google Scholar] [CrossRef]
- Zhang, C.; Gu, H.; Liu, D.; Tong, F.; Wei, H.; Zhou, D.; Fang, J.; Dai, X.; Tian, H. The circ_FAM53B-miR-183-5p-CCDC6 axis modulates the malignant behaviors of papillary thyroid carcinoma cells. Mol. Cell. Biochem. 2022, 477, 2627–2641. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, J.; Li, C.; Peng, X.; Li, H.; Li, Z. Identification and validation of potential target genes in papillary thyroid cancer. Eur. J. Pharmacol. 2019, 843, 217–225. [Google Scholar] [CrossRef]
- Barkovskaya, A.; Buffone, A., Jr.; Žídek, M.; Weaver, V.M. Proteoglycans as Mediators of Cancer Tissue Mechanics. Front. Cell Dev. Biol. 2020, 8, 569377. [Google Scholar] [CrossRef]
- Vasioukhin, V. Adherens junctions and cancer. Subcell. Biochem. 2012, 60, 379–414. [Google Scholar]
- Long, M.; Zhu, Y.; Chen, Z.; Lin, S.; Peng, X.; Luo, D.; Li, H.; Tan, L. Lysine-Specific Demethylase 1 Affects the Progression of Papillary Thyroid Carcinoma via HIF1α and microRNA-146a. J. Clin. Endocrinol. Metab. 2020, 105, 182. [Google Scholar] [CrossRef]
- Mostafaei, S.; Keshavarz, M.; Sadri Nahand, J.; Farhadi Hassankiadeh, R.; Moradinazar, M.; Nouri, M.; Babaei, F.; Ahadi, M.; Payandeh, M.; Salari Esker, A.; et al. Viral infections and risk of thyroid cancer: A systematic review and empirical bayesian meta-analysis. Pathol. Res. Pract. 2020, 216, 152855. [Google Scholar] [CrossRef]
- Ignjatović, V.; Janković Miljuš, J.R.; Rončević, J.V.; Tatić, S.B.; Išić Denčić, T.M.; Đorić, I.Đ.; Šelemetjev, S.A. Focal adhesion kinase splicing and protein activation in papillary thyroid carcinoma progression. Histochem. Cell Biol. 2022, 157, 183–194. [Google Scholar] [CrossRef]
- Hossain, M.A.; Asa, T.A.; Rahman, M.M.; Uddin, S.; Moustafa, A.A.; Quinn, J.M.W.; Moni, M.A. Network-Based Genetic Profiling Reveals Cellular Pathway Differences Between Follicular Thyroid Carcinoma and Follicular Thyroid Adenoma. Int. J. Environ. Res. Public Health 2020, 17, 1373. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Liu, Y.; Du, X.; Wu, Y. TGF-β1 promotes the invasion and migration of papillary thyroid carcinoma cells by inhibiting the expression of lncRNA-NEF. Oncol. Lett. 2019, 17, 3125–3132. [Google Scholar] [CrossRef] [Green Version]
- Fazekas-Lavu, M.; Parker, A.; Spigelman, A.D.; Scott, R.J.; Epstein, R.J.; Jensen, M.; Samaras, K. Thyroid cancer in a patient with Lynch syndrome—Case report and literature review. Ther. Clin. Risk Manag. 2017, 13, 915–918. [Google Scholar] [CrossRef] [Green Version]
- Nieminen, T.T.; Walker, C.J.; Olkinuora, A.; Genutis, L.K.; O'Malley, M.; Wakely, P.E.; LaGuardia, L.; Koskenvuo, L.; Arola, J.; Lepistö, A.H.; et al. Thyroid Carcinomas That Occur in Familial Adenomatous Polyposis Patients Recurrently Harbor Somatic Variants in APC, BRAF, and KTM2D. Thyroid 2020, 30, 380–388. [Google Scholar] [CrossRef]
- Ciavardelli, D.; Bellomo, M.; Consalvo, A.; Crescimanno, C.; Vella, V. Metabolic Alterations of Thyroid Cancer as Potential Therapeutic Targets. Biomed. Res. Int. 2017, 2017, 2545031. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Sun, W.; Wang, Z.; Dong, W.; He, L.; Zhang, T.; Lv, C.; Zhang, H. RBM47/SNHG5/FOXO3 axis activates autophagy and inhibits cell proliferation in papillary thyroid carcinoma. Cell Death Dis. 2022, 13, 270. [Google Scholar] [CrossRef]
- Huo, N.; Cong, R.; Sun, Z.J.; Li, W.C.; Zhu, X.; Xue, C.Y.; Chen, Z.; Ma, L.Y.; Chu, Z.; Han, Y.C.; et al. STAT3/LINC00671 axis regulates papillary thyroid tumor growth and metastasis via LDHA-mediated glycolysis. Cell Death Dis. 2021, 12, 799. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, W.P.; Yao, L.; Zhang, W.; Zhu, J.; Zhang, W.C.; Zhang, Y.H.; Wang, Z.; Yan, Q.G.; Guo, Y.; et al. PRDM1 expression via human parvovirus B19 infection plays a role in the pathogenesis of Hashimoto thyroiditis. Hum. Pathol. 2015, 46, 1913–1921. [Google Scholar] [CrossRef]
- Colombo, C.; Pogliaghi, G.; Tosi, D.; Muzza, M.; Bulfamante, G.; Persani, L.; Fugazzola, L.; Cirello, V. Thyroid cancer harboring PTEN and TP53 mutations: A peculiar molecular and clinical case report. Front. Oncol. 2022, 12, 949098. [Google Scholar] [CrossRef]
- Sawicka-Gutaj, N.; Shawkat, S.; Andrusiewicz, M.; Ziółkowska, P.; Czarnywojtek, A.; Gut, P.; Ruchała, M. EZH2 and SMYD3 expression in papillary thyroid cancer. Oncol. Lett. 2021, 21, 342. [Google Scholar] [CrossRef]
- Plantinga, T.S.; Petrulea, M.S.; Oosting, M.; Joosten, L.A.B.; Piciu, D.; Smit, J.W.; Netea-Maier, R.T.; Georgescu, C.E. Association of NF-κB polymorphisms with clinical outcome of non-medullary thyroid carcinoma. Endocr. Relat. Cancer 2017, 24, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Crezee, T.; Petrulea, M.; Piciu, D.; Jaeger, M.; Smit, J.W.A.; Plantinga, T.S.; Georgescu, C.E.; Netea-Maier, R. Akt1 genetic variants confer increased susceptibility to thyroid cancer. Endocr. Connect. 2020, 9, 1065–1074. [Google Scholar] [CrossRef]
- Gong, Z.; Yang, S.; Wei, M.; Vlantis, A.C.; Chan, J.Y.K.; van Hasselt, C.A.; Li, D.; Zeng, X.; Xue, L.; Tong, M.C.F. The Isoforms of Estrogen Receptor α and β in Thyroid Cancer. Front. Oncol. 2022, 12, 916804. [Google Scholar] [CrossRef]
- Asya, O.; Yumuşakhuylu, A.C.; Bağcı, P.; Kaya, H.; Gönen, A.; Gündoğdu, Y.; Muradov, T.; Şahin, A.; Oysu, Ç. Relationship of PPARG overexpression with prognostic parameters in papillary thyroid carcinoma. Acta. Otorhinolaryngol. Ital. 2022, 42, 34–40. [Google Scholar] [CrossRef]
- Kovaleva, I.V.; Spirina, L.V.; Chizhevskaya, S.Y.; Kondakova, I.V.; Choinzonov, E.L. Expression and content of LC3B protein in tissues of papillary thyroid cancer, relationship with clinical and morphological parameters of tumors. Probl. Oncol. 2022, 68, 439–444. [Google Scholar]
- Humbert, M.; Morán, M.; de la Cruz-Ojeda, P.; Muntané, J.; Wiedmer, T.; Apostolova, N.; McKenna, S.L.; Velasco, G.; Balduini, W.; Eckhart, L.; et al. Assessing Autophagy in Archived Tissue or How to Capture Autophagic Flux from a Tissue Snapshot. Biology 2020, 9, 59. [Google Scholar] [CrossRef] [Green Version]
 —from verified databases;
—from verified databases;  —experimentally determined interactions. Predicted interactions:
—experimentally determined interactions. Predicted interactions:  —neighborhood in the genome;
—neighborhood in the genome;  —gene fusion;
—gene fusion;  —occurrence. Other:
—occurrence. Other:  —joint mention of these proteins in PubMed Abstract;
—joint mention of these proteins in PubMed Abstract;  —co-expression;
—co-expression;  —homology.
—from verified databases; —experimentally determined interactions. Predicted interactions: —neighborhood in the genome; —gene fusion; —occurrence. Other: —joint mention of these proteins in PubMed Abstract; —co-expression; —homology.
—homology.
—from verified databases; —experimentally determined interactions. Predicted interactions: —neighborhood in the genome; —gene fusion; —occurrence. Other: —joint mention of these proteins in PubMed Abstract; —co-expression; —homology. —from verified databases; —experimentally determined interactions. Predicted interactions: —neighborhood in the genome; —gene fusion; —occurrence. Other: —joint mention of these proteins in PubMed Abstract; —co-expression; —homology.
—from verified databases; —experimentally determined interactions. Predicted interactions: —neighborhood in the genome; —gene fusion; —occurrence. Other: —joint mention of these proteins in PubMed Abstract; —co-expression; —homology.
—from verified databases; —experimentally determined interactions. Predicted interactions: —neighborhood in the genome; —gene fusion; —occurrence. Other: —joint mention of these proteins in PubMed Abstract; —co-expression; —homology.
—from verified databases; —experimentally determined interactions. Predicted interactions: —neighborhood in the genome; —gene fusion; —occurrence. Other: —joint mention of these proteins in PubMed Abstract; —co-expression; —homology.


{kind=link}
{kind=link}
{kind=link}
| Change in Expression | MicroRNAs (Target/Regulator) | Reference |
|---|---|---|
| microRNAs upregulated in thyroid cancers | hsa-miR-146a-5p (TRAF and PML) | [20] |
| hsa-miR-146b-5p (RARB) | [21] | |
| hsa-miR-146b-5p (DICER1) | [22] | |
| hsa-miR-221 (AXIN, BCL2, RUNX1, CCNE2) | [23] | |
| hsa-miR-221-3p | [24] | |
| let-7p | [25] | |
| miR-144 (AXIN, BCL2, RUNX1, CCNE2, mTOR) | [23,26] | |
| miR-146a/miR-146b (IRAK1) | [27] | |
| miR-146b | [28] | |
| miR-155 | [29] | |
| miR-17-5p | [22] | |
| miR-181a (RB1) | [30] | |
| miR-181b | [29] | |
| miR-182 (TRIM8) | [31] | |
| miR-19a (PTEN, TSHr, Tg, TTF1 and Pax8, CDH1, an E-cadherin) | [32] | |
| miR-205 (VERG-A, ZEB1) | [33] | |
| miR-221 (RECK, TIMP3) | [34,35,36] | |
| miR-223 (APQ-1) | [37] | |
| miR-23a (PTEN) | [38] | |
| miR-340 (BMP4) | [39] | |
| miR-34a (AXIN, BCL2, RUNX1, CCNE2, AXL) | [23,26] | |
| miR-451a | [22] | |
| miR-9-3p (BLCAP) | [40] | |
| microRNA as oncosupressor | miR-132-3p | [22] |
| let-7a | [41] | |
| let-7b (HMGA2) | [41] | |
| miR-101 | [36] | |
| miR-1179 | [37] | |
| miR-125b (Foxo3, ATG5) | [24] | |
| miR-128 (SPHK1, Bmi-1, EGFR and E2 F3) | [42] | |
| miR-129 (MAL2) | [43] | |
| miR-130a | [12] | |
| miR-132 (FOXA2) | [44] | |
| miR-139b-5p (RICTOR, SMAD2/3 and HNRNPF) | [45] | |
| miR-144 | [36] | |
| miR-144 (E2F8) | [46] | |
| miR-146a-5p | [22] | |
| miR-146b | [36] | |
| miR-148a | [24] | |
| miR-153-3p (RPS6KB1) | [47] | |
| miR-181b | [24] | |
| miR-194 | [24] | |
| miR-199a-3p | [48] | |
| miR-203 (Survivin) | [49] | |
| miR-205 (VERG-A, ZEB1, YAP1) | [50] | |
| miR-21 | [51] | |
| miR-212 (SIRT1) | [52] | |
| miR-217 (AKT3) | [53] | |
| miR-23a (CCNG1) | [54] | |
| miR-24 | [24] | |
| miR-26b-5p (Gsk-3β and β-catenin) | [55] | |
| miR-30a | [24] | |
| miR-335-5p (ICAM-1) | [56] | |
| miR-34a | [26] | |
| miR-34a (MET, XIST) | [57] | |
| miR-424 | [24] | |
| miR-429 (ZEB1) | [58] | |
| miR-451 | [59] | |
| miR-577 (SphK2) | [60] | |
| miR-615 | [24] | |
| miR-9 (BRAF) | [61] | |
| miR-let-7e (HMGB1) | [62] |
| Target Gene | Description |
|---|---|
| ACTN4 | Alpha-actinin-4; |
| ARHGAP35 | Rho GTPase-activating protein 35; |
| CAV2 | Caveolin-2; |
| CCDC6 | Coiled-coil domain-containing protein 6 |
| CDK6 | Cyclin-dependent kinase 6; |
| CDKN1B | Cyclin-dependent kinase inhibitor 1B; |
| COL1A2 | Collagen alpha-2(I) chain; |
| COL6A1 | Collagen alpha-1(VI) chain; |
| CSNK1E | Casein kinase I isoform epsilon; |
| DOT1L | Histone-lysine N-methyltransferase; |
| ELK1 | ETS domain-containing protein Elk-1; |
| FLNA | Filamin-A; |
| FN1 | Fibronectin 1; |
| GART | Phosphoribosylamine--glycine ligase; |
| HDAC2 | Histone deacetylase 1/2; |
| HGF | Hepatocyte growth factor; |
| HK1 | Hexose kinase. |
| HLA-A | HLA class I histocompatibility antigen; |
| IGF2 | Insulin-like growth factor II; |
| IL6 | Interleukin-6; |
| KMT2A | [histone h3]-lysine4 n-trimethyltransferase; |
| KMT2D | Histone methyltransferase. Methylates ‘Lys-4’ of histone H3 (H3K4me). |
| MAPK1 | Mitogen-activated protein kinase 1; |
| MAPK3 | Mitogen-activated protein kinase 3; |
| MTHFD1 | C-1-tetrahydrofolate synthase; |
| MTHFD1L | Monofunctional C1-tetrahydrofolate synthase; |
| MYC | Myc proto-oncogene protein; |
| OGDH | 2-oxoglutarate dehydrogenase; |
| PHYKPL | 5-phosphohydroxy-L-lysine phospho-lyase; |
| PIK3CA | Phosphatidylinositol 4,5-bisphosphate 3-kinase; |
| PKM | Pyruvate kinase m1/2; Pyruvate kinase PKM; |
| PLK1 | Serine/threonine-protein kinase PLK1; |
| PPP1CB | Serine/threonine-protein phosphatase PP1-beta catalytic subunit; |
| RANBP1 | Ran-specific GTPase-activating protein; |
| RBL2 | Rb transcriptional corepressor like 2; |
| RHOA | Ras homolog gene family, member a; |
| SETD8 | [histone h4]-lysine20 n-methyltransferase setd8; |
| SMAD4 | Mothers against decapentaplegic homolog 4; |
| SOD2 | Superoxide dismutase [Mn], mitochondrial; |
| TCF7 | Transcription factor 7; |
| THBS1 | Thrombospondin-1; |
| TNC | Tenascin; |
| UBR4 | E3 ubiquitin-protein ligase UBR4; |
| WNT11 | Wingless-type mmtv integration site family, member 11 |
| Signaling Cascade | Components |
|---|---|
| ECM-receptor interaction | THBS1 |
| COL6A1 | |
| COL1A2 | |
| FN1 | |
| TNC | |
| LAMA4 | |
| Proteoglycans in cancer | THBS1 |
| RHOA | |
| CAV2 | |
| MYC | |
| IGF2 | |
| FLNA | |
| MAPK3 | |
| WNT11 | |
| PIK3CA | |
| FN1 | |
| HGF | |
| MAPK1 | |
| ELK1 | |
| PPP1CB | |
| Lysine degradation | OGDH |
| SETD8 | |
| PHYKPL | |
| KMT2D | |
| DOT1L | |
| KMT2A | |
| Adherens junction | PVRL2 |
| RHOA | |
| TJP1 | |
| SMAD4 | |
| MAPK3 | |
| ACTN4 | |
| EP300 | |
| TCF7 | |
| MAPK1 | |
| Viral carcinogenesis | RBL2 |
| RANBP1 | |
| PKM | |
| CDKN1B | |
| RHOA | |
| CDK6 | |
| HDAC2 | |
| MAPK3 | |
| ACTN4 | |
| EP300 | |
| PIK3CA | |
| MAPK1 | |
| HLA-A | |
| UBR4 | |
| Thyroid cancer | MYC |
| MAPK3 | |
| TCF7 | |
| MAPK1 | |
| CCDC6 | |
| Focal adhesion | THBS1 |
| RHOA | |
| CAV2 | |
| COL6A1 | |
| ARHGAP35 | |
| FLNA | |
| MAPK3 | |
| COL1A2 | |
| ACTN4 | |
| PIK3CA | |
| FN1 | |
| TNC | |
| HGF | |
| MAPK1 | |
| ELK1 | |
| PPP1CB | |
| LAMA4 | |
| One carbon pool by folate | MTHFD1L |
| MTHFD1 | |
| GART | |
| TGF-beta signaling pathway | THBS1 |
| RHOA | |
| SMAD4 | |
| MYC | |
| MAPK3 | |
| EP300 | |
| MAPK1 | |
| Colorectal cancer | RHOA |
| SMAD4 | |
| MYC | |
| MAPK3 | |
| PIK3CA | |
| TCF7 | |
| MAPK1 | |
| Central carbon metabolism in cancer | PKM |
| MYC | |
| MAPK3 | |
| PIK3CA | |
| HK1 | |
| MAPK1 | |
| FoxO signaling pathway | RBL2 |
| CDKN1B | |
| SMAD4 | |
| MAPK3 | |
| CSNK1E | |
| EP300 | |
| SOD2 | |
| PIK3CA | |
| PLK1 | |
| IL6 | |
| MAPK1 |
| Indicator | Hsa-miR-125-b Expression Level (Relative Units) | |
|---|---|---|
| Patients with thyroid pathology | Benign thyroid pathology | 6.75 (1.15; 18.38) |
| PTC | 1.00 (0.19; 4.59) # | |
| Tumor size | T1-2N0M0 | 1.15 (0.07; 10.56) |
| T3-4N0-1M0 | 1.00 (0.62; 3.03) | |
| Regional metastases | T1-2N0M0 | 1.07 (0.14; 4.59) |
| T3-4N1M0 | 0.81 (0.19; 18.38) | |
| Invasion | − | 1.00 (0.14; 3.03) |
| + | 2.63 (0.19; 24.25) * | |
| Histological subtype | Classical subtype | 1.11 (0.33; 10.56) |
| Follicular subtype | 0.03 (0; 0.14) *** | |
| BRAFV600E mutational status | − | 2.66 (0.87; 32.00) |
| + | 0.21 (0.01; 0.66) ** | |
| Risk of disease recurrence after treatment (ATA) | Low risk | 0.11 (0.02; 1.24) |
| Intermediate risk | 11.78 (2.39; 28.13) **** | |
| High risk | 1.00 (0.62; 1.52) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spirina, L.V.; Kovaleva, I.V.; Chizhevskaya, S.Y.; Chebodaeva, A.V.; Tarasenko, N.V. Autophagy-Related MicroRNA: Tumor miR-125b and Thyroid Cancers. Genes 2023, 14, 685. https://doi.org/10.3390/genes14030685
Spirina LV, Kovaleva IV, Chizhevskaya SY, Chebodaeva AV, Tarasenko NV. Autophagy-Related MicroRNA: Tumor miR-125b and Thyroid Cancers. Genes. 2023; 14(3):685. https://doi.org/10.3390/genes14030685
Chicago/Turabian StyleSpirina, Liudmila V., Irina V. Kovaleva, Svetlana Yu. Chizhevskaya, Anastasiya V. Chebodaeva, and Nataliya V. Tarasenko. 2023. "Autophagy-Related MicroRNA: Tumor miR-125b and Thyroid Cancers" Genes 14, no. 3: 685. https://doi.org/10.3390/genes14030685