
The Adaptive Evolution in the Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) Revealed by the Diversity of Larval Gut Bacteria

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Collection and Laboratory Feeding
2.2. Processing of S. frugiperda Larvae
2.3. DNA Extraction and 16S rDNA Sequencing
2.4. Statistical Analyses
3. Results
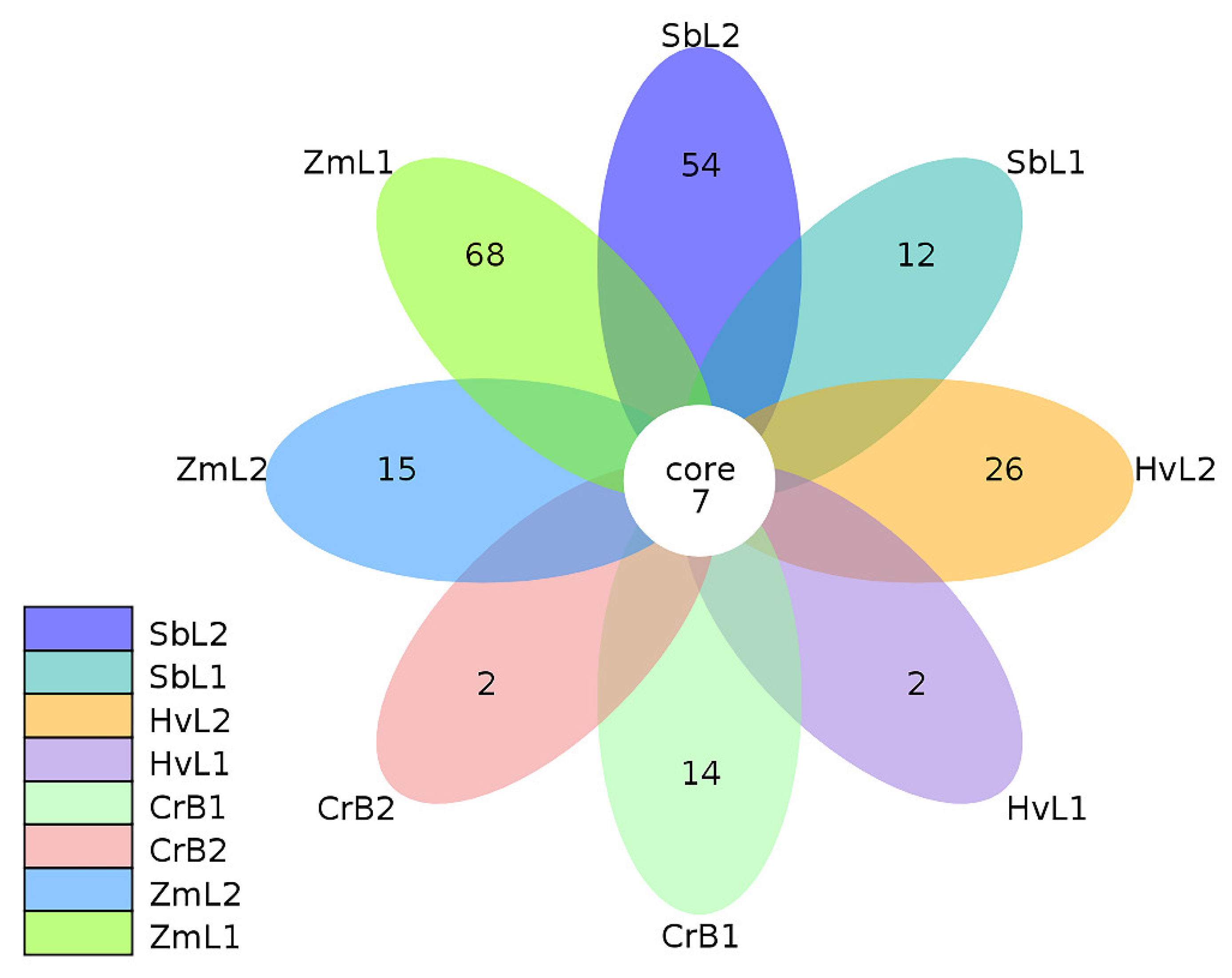
3.1. Sequence Analysis
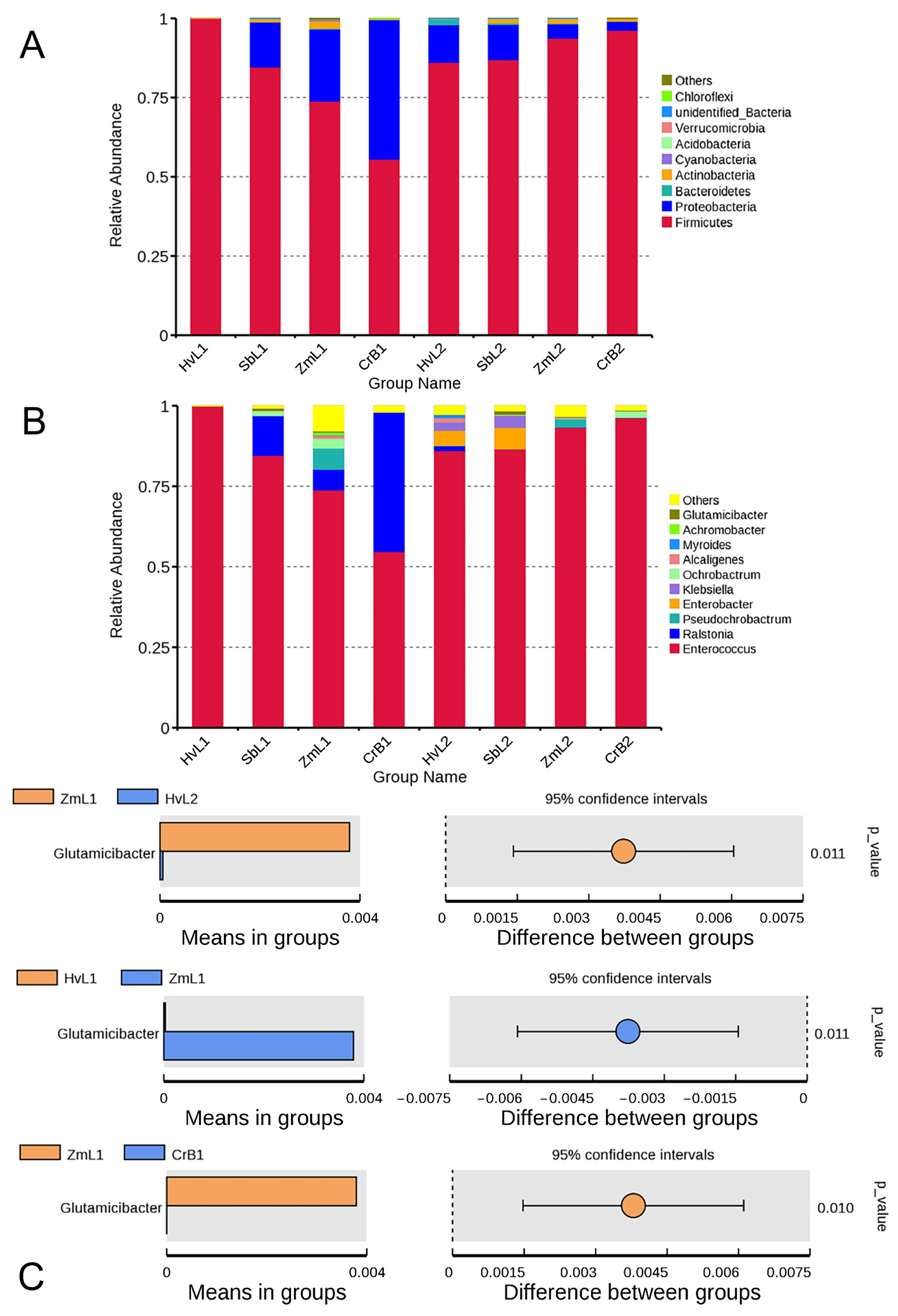
3.2. Taxa Annotation and Relative Abundance
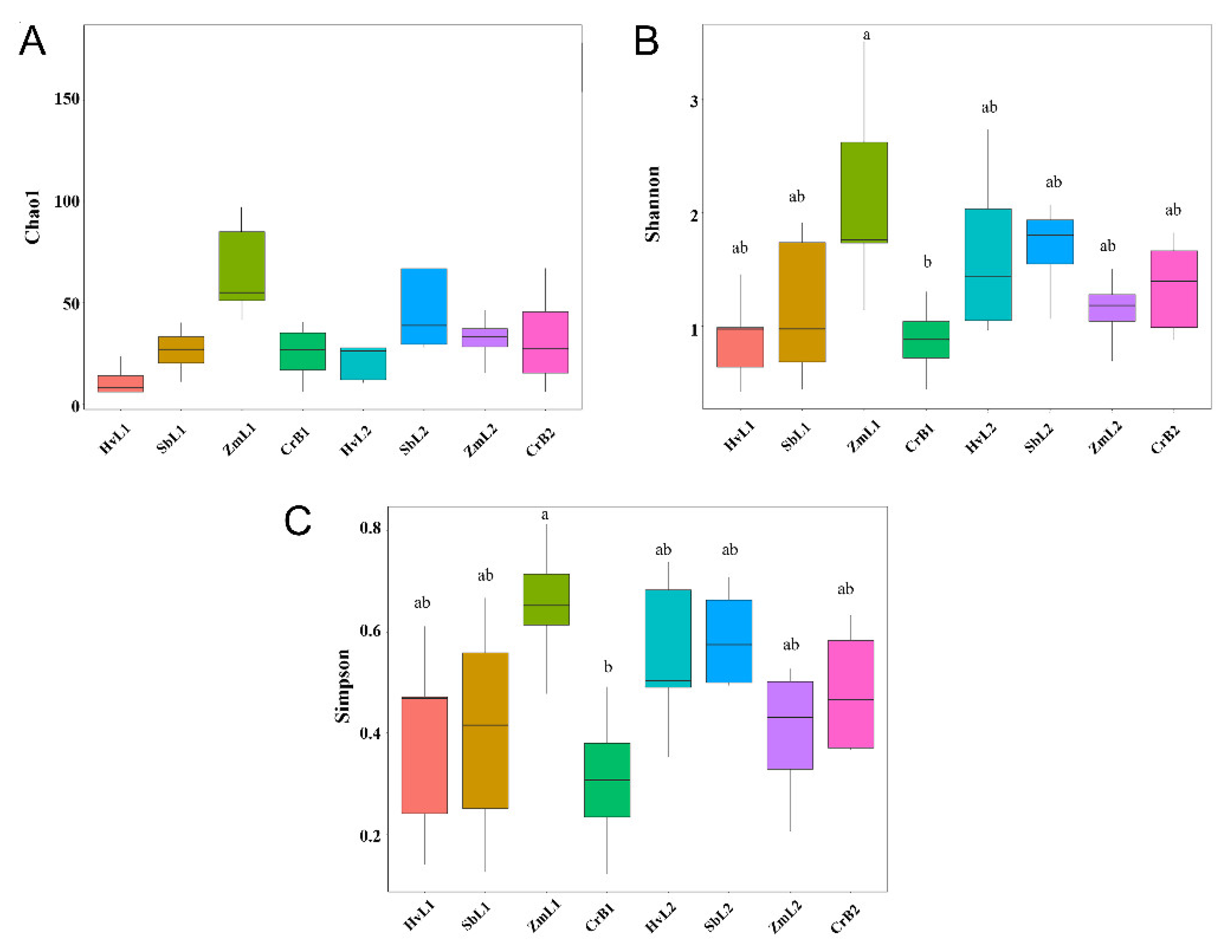
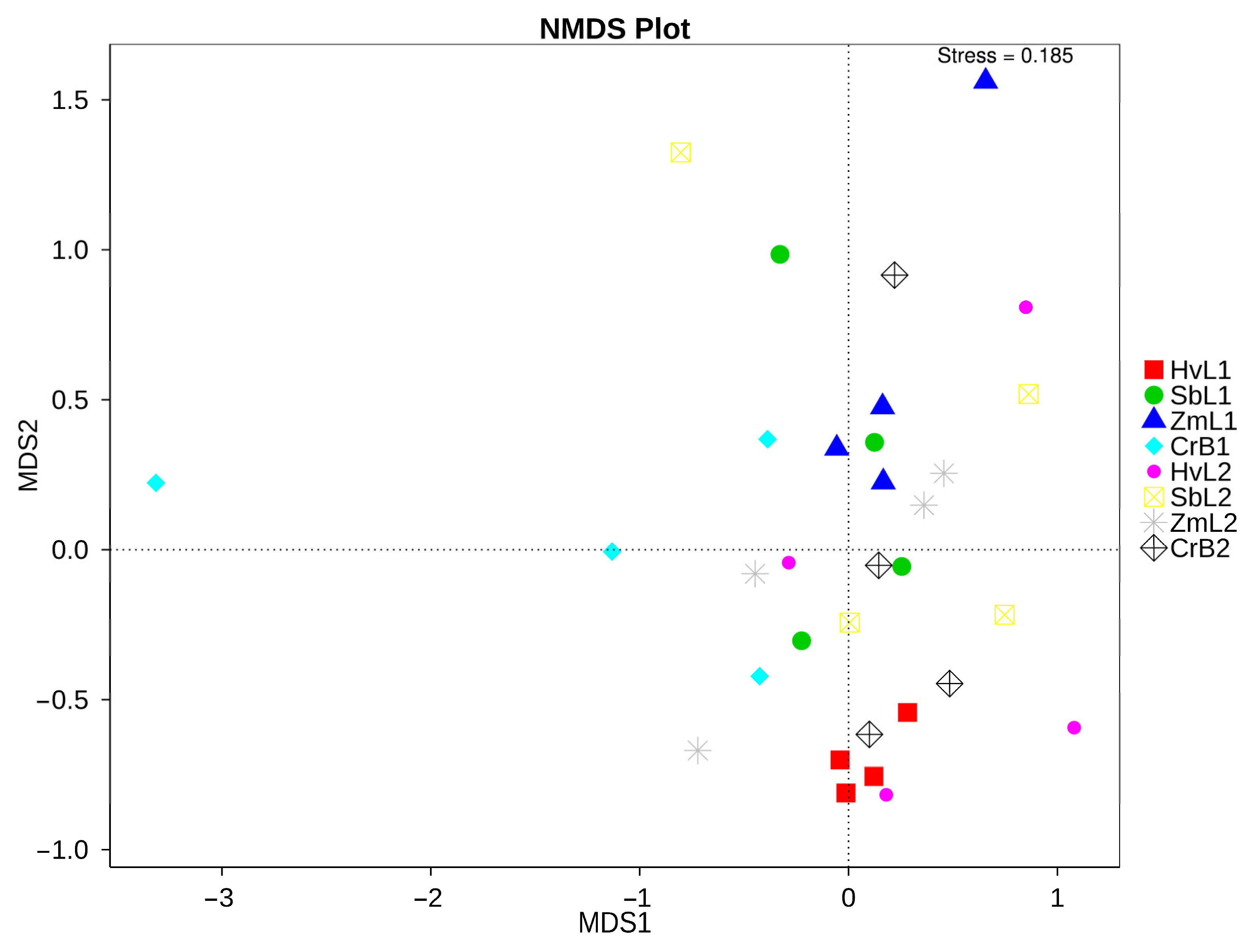
3.3. Diversity of Gut Bacteria
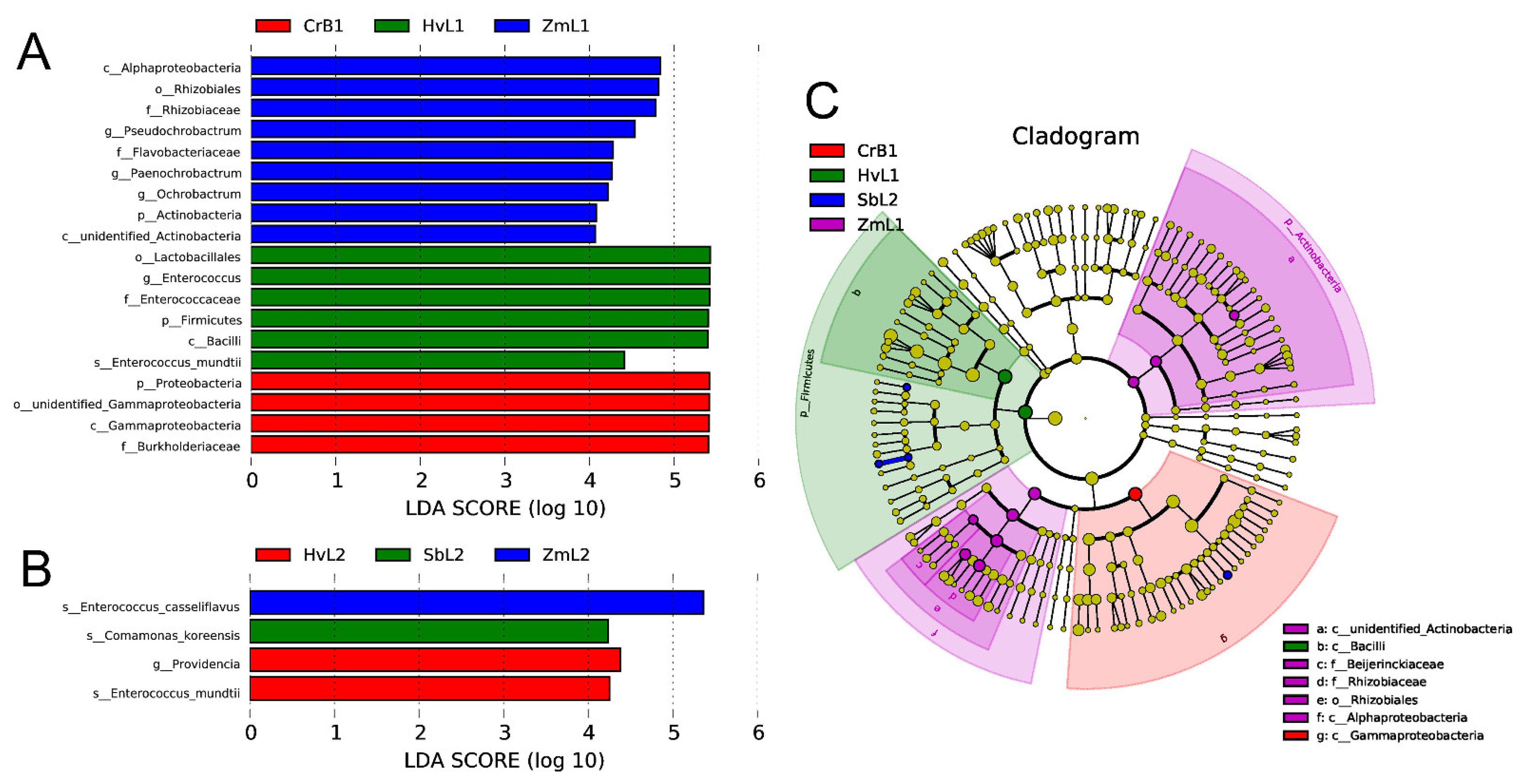
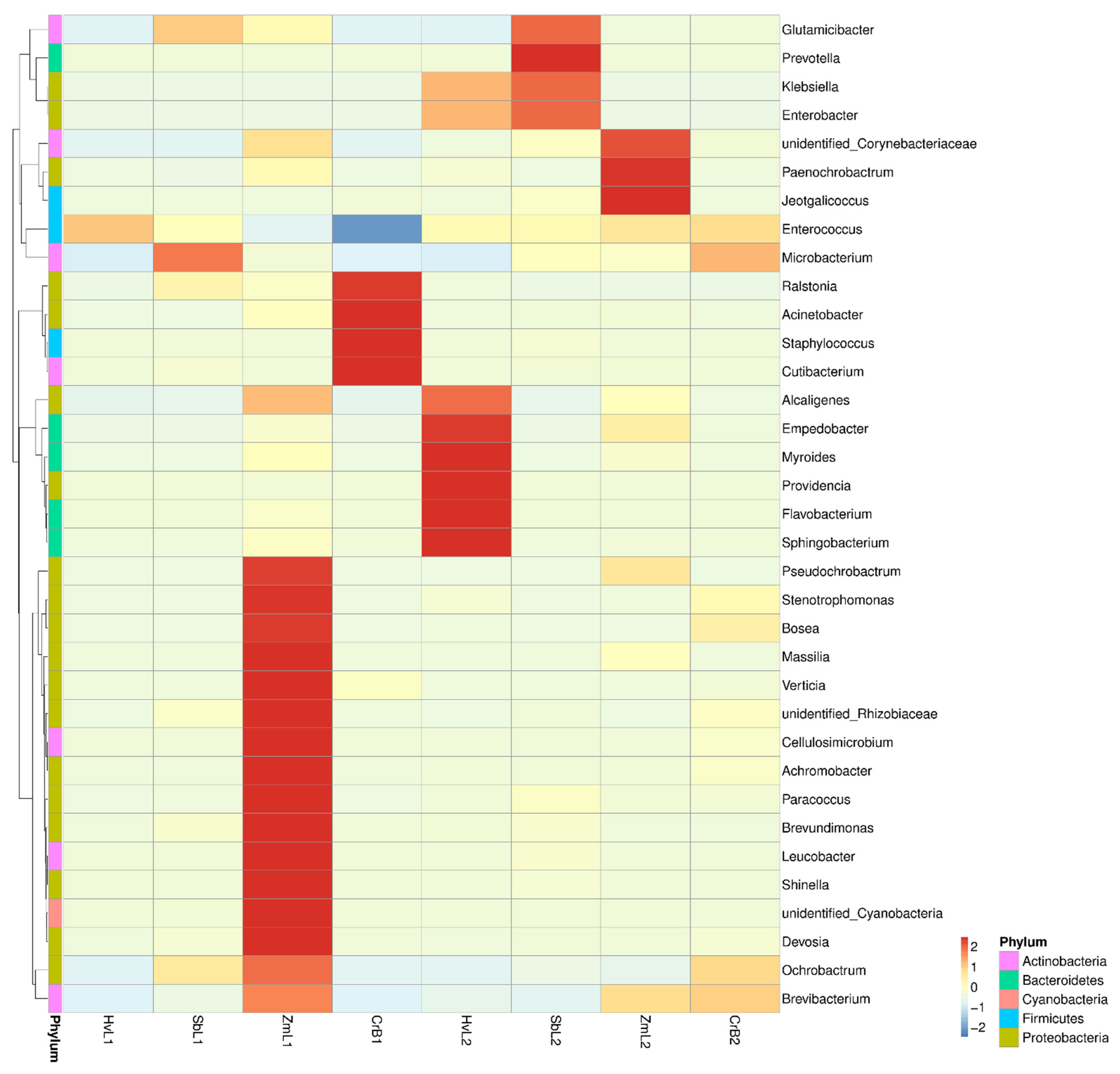
3.4. Cluster Analysis of Predominant Bacteria
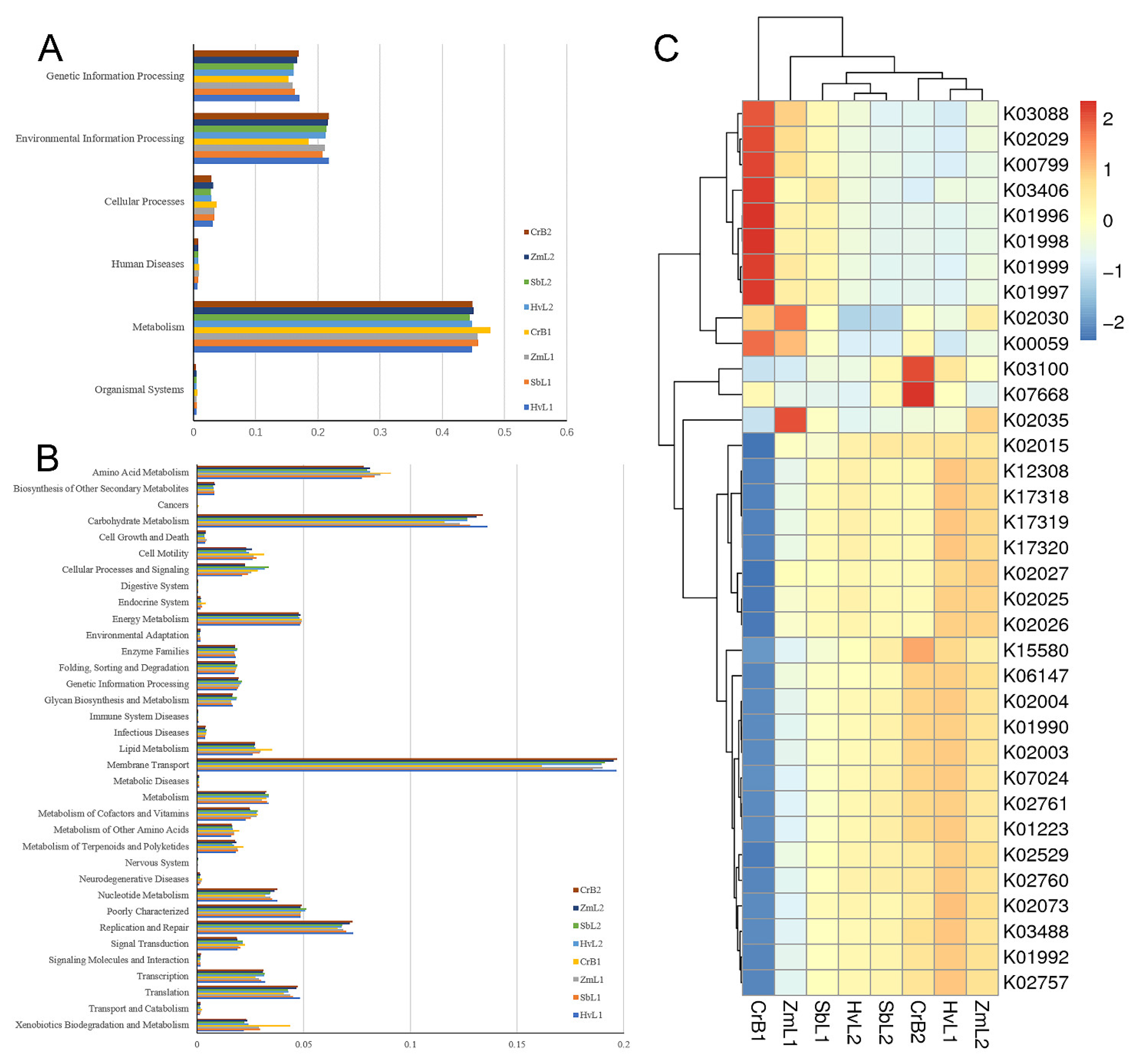
3.5. Prediction of Bacterial Functions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, J.E.; Abbott, J. The Natural History of the Rarer Lepidopterous Insects of Georgia: Including their Systematic Characters, the Particulars of Their Several Metamorphoses, and the Plants on Which They Feed; Missouri Botanical Garden Press: London, UK, 1797. [Google Scholar] [CrossRef]
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Johnson, S.J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the western hemisphere. Int. J. Trop. Insect Sci. 1987, 8, 543–549. [Google Scholar] [CrossRef]
- Todd, E.L.; Poole, R.W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the western hemisphere. Ann. Entomol. Soc. Am. 1980, 73, 722–738. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in west and central Africa. PLoS ONE 2016, 11, e165632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cock, M.; Beseh, P.K.; Buddie, A.G.; Cafá, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef] [Green Version]
- Sharanabasappa, C.; Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, H.M.M.; Maruthi, M.S.; Pavithra, H.B.; Hegbe, K.; Navi, S.; Prabhu, S.T.; Goergen, G.E. First report of the fall armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- Yang, P.; Chang, X. The occurrence and impact of Spodoptera frugiperda in Asia and Africa and its prevention and control strategies. China Plant Prot. Guide 2019, 39, 88–90. [Google Scholar]
- Jiang, Y.; Liu, J.; Zhu, X. Analysis of the occurrence and future trends of the invasion of Spodoptera frugiperda in my country. China Plant Prot. 2019, 39, 33–35. [Google Scholar]
- Jiang, Y.; Liu, J.; Xie, M.; Li, Y.; Yang, J.; Zhang, M.; Qiu, K. Observation on law of diffusion damage of Spodoptera frugiperda in China in 2019. Plant Prot. 2019, 45, 10–19. [Google Scholar]
- Sun, X.; Hu, C.; Jia, H.; Wu, Q.; Shen, X.; Zhao, S.; Jiang, Y.; Wu, K. Case study on the first immigration of fall armyworm, Spodoptera frugiperda invading into China. J. Integr. Agric. 2021, 20, 664–672. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Specht, A.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Wyckhuys, K.; Jia, X.; Nie, F.; Wu, K. Fall armyworm invasion heightens pesticide expenditure among Chinese smallholder farmers. J. Environ. Manag. 2021, 282, 111949. [Google Scholar] [CrossRef]
- Silver, K.S.; Soderlund, D.M. Action of pyrazoline-type insecticides at neuronal target sites. Pestic. Biochem. Phys. 2005, 81, 136–143. [Google Scholar] [CrossRef]
- Zhang, Y. Natural enemy insect resources of Spodoptera frugiperda and domestic research progress. Agric. Technol. Serv. 2020, 37, 38–39. [Google Scholar]
- Sharon, G.; Segal, D.; Ringo, J.M.; Hefetz, A.; Zilber-Rosenberg, I.; Rosenberg, E. Commensal bacteria play a role in mating preference of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2010, 107, 20051–20056. [Google Scholar] [CrossRef] [Green Version]
- Sharon, G.; Segal, D.; Zilber-Rosenberg, I.; Rosenberg, E. Symbiotic bacteria are responsible for diet-induced mating preference in Drosophila melanogaster, providing support for the hologenome concept of evolution. Gut Microbes 2011, 2, 190–192. [Google Scholar] [CrossRef] [Green Version]
- Vollaard, E.J.; Clasener, H.A. Colonization resistance. Antimicrob. Agents Chemother. 1994, 38, 409–414. [Google Scholar] [CrossRef] [Green Version]
- Takatsuka, J.; Kunimi, Y. Intestinal bacteria affect growth of Bacillus thuringiensis in larvae of the oriental tea tortrix, Homona magnanima diakonoff (Lepidoptera: Tortricidae). J. Invertebr. Pathol. 2000, 76, 222–226. [Google Scholar] [CrossRef]
- Paramasiva, I.; Sharma, H.C.; Krishnayya, P.V. Antibiotics influence the toxicity of the delta endotoxins of Bacillus thuringiensis towards the cotton bollworm, Helicoverpa armigera. BMC Microbiol. 2014, 14, 200. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Li, N.; Yue, Y.X.; Yan, F.F.; Wang, N.N.; Huo, G.C. Research progress on the effects of dietary main components in diet on intestinal microbial composition and metabolism. Food Ind. Sci. Technol. 2019, 40, 354–360. [Google Scholar]
- Liu, C. The Effects of Different Sugar Source Feeds on The Gut Health and Gut Microflora of Overwintering Honeybees; Shandong Agricultural University: Taian, China, 2017. [Google Scholar]
- Acevedo, F.E.; Peiffer, M.; Tan, C.W.; Stanley, B.A.; Stanley, A.; Wang, J.; Jones, A.G.; Hoover, K.; Rosa, C.; Luthe, D.; et al. Fall armyworm-associated gut bacteria modulate plant defense responses. Mol. Plant Microbe Interact. 2017, 30, 127–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broderick, N.A.; Buchon, N.; Lemaitre, B. Microbiota-induced changes in Drosophila melanogaster host gene expression and gut morphology. mBio 2014, 5, e01117-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaturvedi, S.; Rego, A.; Lucas, L.K.; Gompert, Z. Sources of variation in the gut microbial community of Lycaeides melissa Caterpillars. Sci. Rep. 2017, 7, 11335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhou, H.; Lai, Y.; Chen, Q.; Yu, X.Q.; Wang, X. Gut microbiota dysbiosis influences metabolic homeostasis in Spodoptera frugiperda. Front. Microbiol. 2021, 12, 727434. [Google Scholar] [CrossRef] [PubMed]
- Hood, G.R.; Forbes, A.A.; Powell, T.H.; Egan, S.P.; Hamerlinck, G.; Smith, J.J.; Feder, J.L. Sequential divergence and the multiplicative origin of community diversity. Proc. Natl. Acad. Sci. USA 2015, 112, E5980–E5989. [Google Scholar] [CrossRef] [Green Version]
- Ceja-Navarro, J.A.; Vega, F.E.; Karaoz, U.; Hao, Z.; Jenkins, S.; Lim, H.C.; Kosina, P.; Infante, F.; Northen, T.R.; Brodie, E.L. Gut microbiota mediate caffeine detoxification in the primary insect pest of coffee. Nat. Commun. 2015, 6, 7618. [Google Scholar] [CrossRef] [Green Version]
- Zhu-Salzman, K.; Koiwa, H.; Salzman, R.A.; Shade, R.E.; Ahn, J.E. Cowpea bruchid Callosobruchus maculatus uses a three-component strategy to overcome a plant defensive cysteine protease inhibitor. Insect Mol. Biol. 2003, 12, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Després, L.; David, J.P.; Gallet, C. The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol. Evol. 2007, 22, 298–307. [Google Scholar] [CrossRef]
- Berenbaum, M.R. Allelochemicals in insect-microbe-plant Interactions; agents provocateurs in the coevolutionary arms race. In Novel Aspects of Insect-Plant Interactions; Barbosa, P., Letourneau, D.K., Eds.; Wiley-Interscience: New York, NY, USA, 1988; pp. 97–123. [Google Scholar]
- Feldhaar, H. Bacterial symbionts as mediators of ecologically important traits of insect hosts. Ecol. Entomol. 2011, 36, 533–543. [Google Scholar] [CrossRef]
- Hansen, A.K.; Moran, N.A. The impact of microbial symbionts on host plant utilization by herbivorous insects. Mol. Ecol. 2014, 23, 1473–1496. [Google Scholar] [CrossRef]
- Semova, I.; Carten, J.D.; Stombaugh, J.; Mackey, L.C.; Knight, R.; Farber, S.A.; Rawls, J.F. Microbiota regulate intestinal absorption and metabolism of fatty acids in the zebrafish. Cell Host Microbe 2012, 12, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Ellegaard, K.M.; Engel, P. Genomic diversity landscape of the honey bee gut microbiota. Nat. Commun. 2019, 10, 446. [Google Scholar] [CrossRef] [Green Version]
- Moran, N.A.; Hansen, A.K.; Powell, J.E.; Sabree, Z.L. Distinctive gut microbiota of honey bees assessed using deep sampling from individual worker bees. PLoS ONE 2012, 7, e36393. [Google Scholar] [CrossRef] [Green Version]
- Bing, X.; Gerlach, J.; Loeb, G.; Buchon, N. Nutrient-Dependent Impact of Microbes on Drosophila suzukii Development. mBio 2018, 9, e02199-17. [Google Scholar] [CrossRef] [Green Version]
- Erkosar, B.; Kolly, S.; van der Meer, J.R.; Kawecki, T.J. Adaptation to chronic nutritional stress leads to reduced dependence on microbiota in Drosophila melanogaster. MBio 2017, 8, e01496-17. [Google Scholar] [CrossRef] [Green Version]
- Kudo, R.; Masuya, H.; Endoh, R.; Kikuchi, T.; Ikeda, H. Gut bacterial and fungal communities in ground-dwelling beetles are associated with host food habit and habitat. ISME J. 2019, 13, 676–685. [Google Scholar] [CrossRef] [Green Version]
- Rossmassler, K.; Dietrich, C.; Thompson, C.; Mikaelyan, A.; Nonoh, J.O.; Scheffrahn, R.H.; Sillam-Dusses, D.; Brune, A. Metagenomic analysis of the microbiota in the highly compartmented hindguts of six wood- or soil-feeding higher termites. Microbiome 2015, 3, 56. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Cobas, A.E.; Maiques, E.; Angelova, A.; Carrasco, P.; Moya, A.; Latorre, A. Diet shapes the gut microbiota of the omnivorous cockroach Blattella germanica. FEMS Microbiol. Ecol. 2015, 91, fiv022. [Google Scholar] [CrossRef] [Green Version]
- Broderick, N.A.; Raffa, K.F.; Goodman, R.M.; Handelsman, J. Census of the bacterial community of the gypsy moth larval midgut by using culturing and culture-independent methods. Appl. Environ. Microbiol. 2004, 70, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Xia, X.; Chen, J.; Geoff, M.G.; You, M. Effects of different diets on the diversity of larval gut bacteria of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Acta Entomol. 2019, 62, 1172–1185. [Google Scholar]
- Yuan, X.; Zhang, X.; Liu, X.; Dong, Y.; Yan, Z.; Lv, D.; Wang, P.; Li, Y. Comparison of gut bacterial communities of Grapholita molesta (Lepidoptera: Tortricidae) reared on different host plants. Int. J. Mol. Sci. 2021, 22, 6843. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wu, Y.; Gu, R.; Zou, X.; Zhang, Z.; Niu, X.; Wang, Z.; Chen, J.; Wu, Y.; Li, T.; et al. Isolation and identification of gut bacteria of Spodoptera frugiperda that migrated to Chongqing area. J. Southwest Univ. Nat. Sci. Ed. 2019, 41, 8–14. [Google Scholar] [CrossRef]
- Wei, J.; Tang, Y.; Gu, R.; Wu, Y.; Zou, X.; Zhang, Z.; Chen, J.; Li, C.; Pan, G.; Zhou, Z. Isolation and identification of endophytic bacteria of maize leaf in Chongqing area and comparison between maize microbes and gut bacteria of Spodoptera frugiperda. J. Southwest Univ. Nat. Sci. Ed. 2019, 41, 17–23. [Google Scholar]
- Li, Q.; Tang, Y.; Jiang, R.; Zhang, Y.; Feng, Z.; Bai, X.; Gu, R.; Wu, Y.; Wu, Y.; Chen, J.; et al. Isolation and identification of gut bacteria of Spodoptera frugiperda feeding on maize in Yunnan, China. J. Southwest Univ. Nat. Sci. Ed. 2020, 42, 1–8. [Google Scholar]
- Xu, T.M.; Fu, C.Y.; Su, Z.T.; Xiao, G.L.; Li, W.W.; Chen, B. Composition and diversity of gut bacteria community of Spodoptera frugiperda from the first invasion site and the dispersal area in Yunnan province. Plant Prot. 2020, 46, 116–125. [Google Scholar]
- Wang, S.; Zhu, Q.; Tan, Y.; Ma, Q.; Wang, R.; Zhang, M.; Xu, H.; Zhang, Z. Artificial diets and rearing technique of Spodoptera frugiperda (J. E. Smith) in laboratory. J. Environ. Entomol. 2019, 41, 742–747. [Google Scholar]
- Franzén, O.; Hu, J.; Bao, X.; Itzkowitz, S.H.; Peter, I.; Bashir, A. Improved OTU-picking using long-read 16S rRNA gene amplicon sequencing and generic hierarchical clustering. Microbiome 2015, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Mason, C.J.; Hoover, K.; Felton, G.W. Effects of maize (Zea mays) genotypes and microbial sources in shaping fall armyworm (Spodoptera frugiperda) gut bacterial communities. Sci. Rep. 2021, 11, 4429. [Google Scholar] [CrossRef]
- Lü, D.; Dong, Y.; Yan, Z.; Liu, X.; Zhang, Y.; Yang, D.; He, K.; Wang, Z.; Wang, P.; Yuan, X.; et al. Dynamics of gut microflora across the life cycle of Spodoptera frugiperda and its effects on the feeding and growth of larvae. Pest Manag. Sci. 2023, 79, 173–182. [Google Scholar] [CrossRef]
- Chen, Y.P.; Li, Y.H.; Sun, Z.X.; Du, E.W.; Lu, Z.H.; Li, H.; Gui, F.R. Effects of host plants on bacterial community structure in larvae midgut of Spodoptera frugiperda. Insects 2022, 13, 373. [Google Scholar] [CrossRef]
- Li, Y.; Liu, L.; Cai, X.; Yang, X.; Lin, J.; Shu, B. The bacterial and fungal communities of the larval midgut of Spodoptera frugiperda (Lepidoptera: Noctuidae) varied by feeding on two cruciferous vegetables. Sci. Rep. 2022, 12, 13063. [Google Scholar]
- Gomes, A.F.F.; Omoto, C.; Cônsoli, F.L. Gut bacteria of field-collected larvae of Spodoptera frugiperda undergo selection and are more diverse and active in metabolizing multiple insecticides than laboratory-selected resistant strains. J. Pest Sci. 2020, 93, 833–851. [Google Scholar] [CrossRef]
- Jones, A.G.; Mason, C.J.; Felton, G.W.; Hoover, K. Host plant and population source drive diversity of microbial gut communities in two polyphagous insects. Sci. Rep. 2019, 9, 2792. [Google Scholar] [CrossRef] [Green Version]
- Gichuhi, J.; Sevgan, S.; Khamis, F.; Van den Berg, J.; du Plessis, H.; Ekesi, S.; Herren, J.K. Diversity of fall armyworm, Spodoptera frugiperda and their gut bacterial community in Kenya. PeerJ 2020, 8, e8701. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Wu, Q.; Gao, X.; Wu, K. Population life tables for the invasive fall armyworm, Spodoptera frugiperda fed on major oil crops planted in China. J. Integr. Agric. 2021, 20, 745–754. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, K.A.; Lee, W.J. Microbiota, gut physiology, and insect immunity. Adv. Insect Physiol. 2017, 52, 111–138. [Google Scholar] [CrossRef]
- Strano, C.P.; Malacrinò, A.; Campolo, O.; Palmeri, V. Influence of host plant on Thaumetopoea pityocampa gut bacterial community. Microb. Ecol. 2018, 75, 487–494. [Google Scholar] [CrossRef]
- Tinker, K.A.; Ottesen, E.A. The core gut microbiome of the American cockroach, Periplaneta americana, is stable and resilient to dietary shifts. Appl. Environ. Microbiol. 2016, 82, 6603–6610. [Google Scholar] [CrossRef] [Green Version]
- Lv, D.; Liu, X.; Dong, Y.; Yan, Z.; Zhang, X.; Wang, P.; Yuan, X.; Li, Y. Comparison of gut bacterial communities of fall armyworm (Spodoptera frugiperda) reared on different host plants. Int. J. Mol. Sci. 2021, 22, 11266. [Google Scholar] [CrossRef]
- Liu, Y.J.; Shen, Z.J.; Yu, J.M.; Li, Z.; Liu, X.X.; Xu, H.L. Comparison of gut bacterial communities and their associations with host diets in four fruit borers. Pest Manag. Sci. 2020, 76, 1353–1362. [Google Scholar] [CrossRef]
- Hübner, J. Zuträge zur Sammlung Exotischer Schmettlinge [sic]; Jacob Hübner: Augsburg, Germany, 1808. [Google Scholar]
- Presa-Parra, E.; Lasa, R.; Reverchon, F.; Simón, O.; Williams, T. Use of biocides to minimize microbial contamination in Spodoptera exigua multiple nucleopolyhedrovirus preparations. Biol. Control 2020, 151, 104408. [Google Scholar] [CrossRef]
- Xu, J.; Feng, J. Research progress on genetic diversity and pathogenic genomics of R. solanacearum. Chin. Agric. Sci. 2013, 46, 2902–2909. [Google Scholar]
- Gomes, S.I.; Kielak, A.M.; Hannula, S.E.; Heinen, R.; Jongen, R.; Keesmaat, I.; De Long, J.R.; Bezemer, T.M. Microbiomes of a specialist caterpillar are consistent across different habitats but also resemble the local soil microbial communities. Anim. Microbiome 2020, 2, 37. [Google Scholar] [CrossRef] [PubMed]
- Paulson, A.R.; von Aderkas, P.; Perlman, S.J. Bacterial associates of seed-parasitic wasps (Hymenoptera: Megastigmus). BMC Microbiol. 2014, 14, 224. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Liang, B.; Li, F.; Li, S. Recent advances in the biodegradation of chlorothalonil. Curr. Microbiol. 2011, 63, 450–457. [Google Scholar] [CrossRef]
- Li, D.D.; Li, J.Y.; Hu, Z.Q.; Liu, T.X.; Zhang, S.Z. Fall armyworm gut bacterial diversity associated with different developmental stages, environmental habitats, and diets. Insects 2022, 13, 762. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [Green Version]
- Dillon, R.J.; Dillon, V.M. The gut bacteria of insects: Nonpathogenic interactions. Annu. Rev. Entomol. 2004, 49, 71–92. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, 60. [Google Scholar] [CrossRef] [Green Version]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Higgins, C.F. ABC transporters: From microorganisms to man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef]
- Marger, M.D.; Saier, M.H. A major superfamily of transmembrane facilitators that catalyze uniport, symport and antiport. Trends Biochem Sci. 1993, 18, 13–20. [Google Scholar] [CrossRef]
- Paulsen, I.T.; Skurray, R.A.; Tam, R.; Saier, M.H., Jr.; Turner, R.J.; Weiner, J.H.; Goldberg, E.B.; Grinius, L.L. The SMR family: A novel family of multidrug efflux proteins involved with the efflux of lipophilic drugs. Mol. Microbiol. 1996, 19, 1167–1175. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Raw Reads * | Clean Reads * | Base (nt) | AvgLen (nt) * | Effective (%) * |
|---|---|---|---|---|---|
| HvL1.1 | 18,434 | 16,712 | 24,823,416 | 1485 | 90.66 |
| HvL1.2 | 13,733 | 12,297 | 18,259,102 | 1484 | 89.54 |
| HvL1.3 | 12,142 | 10,472 | 15,560,783 | 1485 | 86.25 |
| HvL1.4 | 16,263 | 14,725 | 21,871,158 | 1485 | 90.54 |
| SbL1.1 | 16,291 | 14,668 | 21,773,693 | 1484 | 90.04 |
| SbL1.2 | 20,232 | 18,538 | 27,167,447 | 1465 | 91.63 |
| SbL1.3 | 23,156 | 19,666 | 29,188,401 | 1484 | 84.93 |
| SbL1.4 | 23,467 | 21,067 | 31,264,901 | 1484 | 89.77 |
| ZmL1.1 | 17,639 | 15,895 | 23,578,489 | 1483 | 90.11 |
| ZmL1.2 | 10,148 | 9020 | 13,191,119 | 1462 | 88.88 |
| ZmL1.3 | 12,649 | 10,831 | 15,562,182 | 1436 | 85.63 |
| ZmL1.4 | 13,328 | 12,007 | 17,818,003 | 1483 | 90.09 |
| CrB1.1 | 26,445 | 23,292 | 34,155,876 | 1466 | 88.08 |
| CrB1.2 | 22,652 | 20,542 | 30,485,052 | 1484 | 90.69 |
| CrB1.3 | 18,589 | 16,033 | 23,804,488 | 1484 | 86.25 |
| CrB1.4 | 14,176 | 12,932 | 18,909,984 | 1462 | 91.22 |
| HvL2.1 | 12,065 | 11,046 | 16,295,806 | 1475 | 91.55 |
| HvL2.2 | 15,120 | 13,924 | 20,641,745 | 1482 | 92.09 |
| HvL2.3 | 25,269 | 21,263 | 31,586,883 | 1485 | 84.15 |
| HvL2.4 | 21,157 | 19,133 | 28,414,494 | 1485 | 90.43 |
| SbL2.1 | 15,491 | 13,452 | 19,895,164 | 1478 | 86.84 |
| SbL2.2 | 15,980 | 14,028 | 20,809,492 | 1483 | 87.78 |
| SbL2.3 | 18,850 | 16,365 | 24,214,434 | 1479 | 86.82 |
| SbL2.4 | 25,471 | 22,426 | 33,299,817 | 1484 | 88.05 |
| ZmL2.1 | 12,346 | 11,205 | 16,617,495 | 1483 | 90.76 |
| ZmL2.2 | 19,422 | 17,006 | 25,168,430 | 1479 | 87.56 |
| ZmL2.3 | 17,704 | 16,464 | 24,431,320 | 1483 | 93 |
| ZmL2.4 | 13,393 | 12,090 | 17,803,176 | 1472 | 90.27 |
| CrB2.1 | 25,248 | 21,817 | 32,396,921 | 1484 | 86.41 |
| CrB2.2 | 13,162 | 11,940 | 17,728,360 | 1484 | 90.72 |
| CrB2.3 | 16,103 | 14,615 | 21,684,846 | 1483 | 90.76 |
| CrB2.4 | 22,175 | 20,180 | 29,957,818 | 1484 | 91 |
| Total | 568,300 | 505,651 | 748,360,295 | 47319 | 2852.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-P.; Liu, X.; Yi, C.-Y.; Chen, X.-Y.; Liu, C.-H.; Zhang, C.-C.; Chen, Q.-D.; Chen, S.; Liu, H.-L.; Pu, D.-Q. The Adaptive Evolution in the Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) Revealed by the Diversity of Larval Gut Bacteria. Genes 2023, 14, 321. https://doi.org/10.3390/genes14020321
Wang Y-P, Liu X, Yi C-Y, Chen X-Y, Liu C-H, Zhang C-C, Chen Q-D, Chen S, Liu H-L, Pu D-Q. The Adaptive Evolution in the Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) Revealed by the Diversity of Larval Gut Bacteria. Genes. 2023; 14(2):321. https://doi.org/10.3390/genes14020321
Chicago/Turabian StyleWang, Yan-Ping, Xu Liu, Chun-Yan Yi, Xing-Yu Chen, Chang-Hua Liu, Cui-Cui Zhang, Qing-Dong Chen, Song Chen, Hong-Ling Liu, and De-Qiang Pu. 2023. "The Adaptive Evolution in the Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) Revealed by the Diversity of Larval Gut Bacteria" Genes 14, no. 2: 321. https://doi.org/10.3390/genes14020321