
Comprehensive Analysis of miRNA and mRNA Expression Profiles during Muscle Development of the Longissimus Dorsi Muscle in Gannan Yaks and Jeryaks
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Total RNA Extraction
2.3. Construction and Sequencing of Small Ribonucleic Acid Library
2.4. Differentially Expressed miRNA Analysis
2.5. Target Gene Prediction of Differentially Expressed miRNAs and KEGG and GO Analysis
2.6. miRNA–mRNA Interaction Network Construction
2.7. Real-Time qPCR Validation of Differentially Expressed miRNAs
3. Results
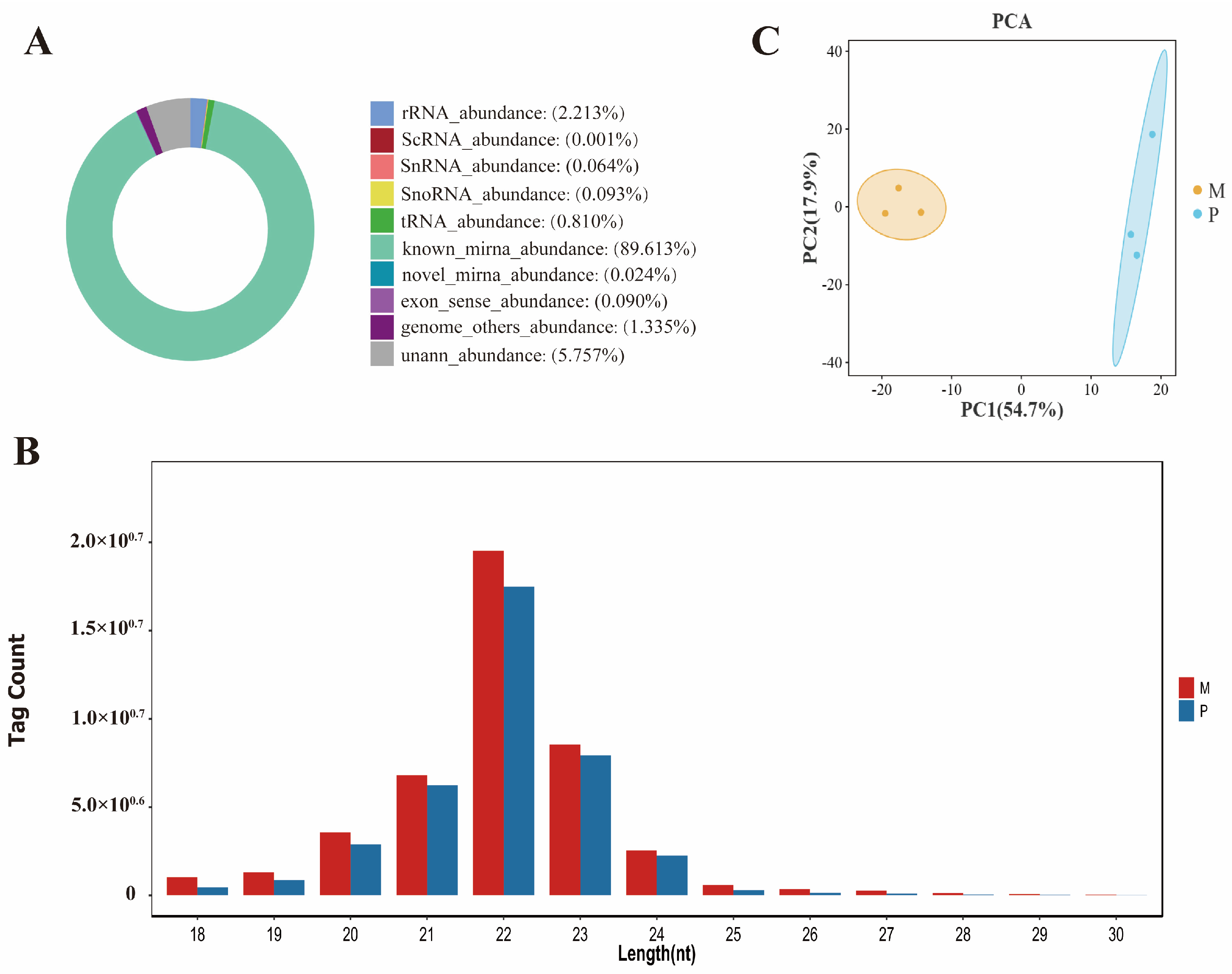
3.1. Summary of Sequencing Small RNA
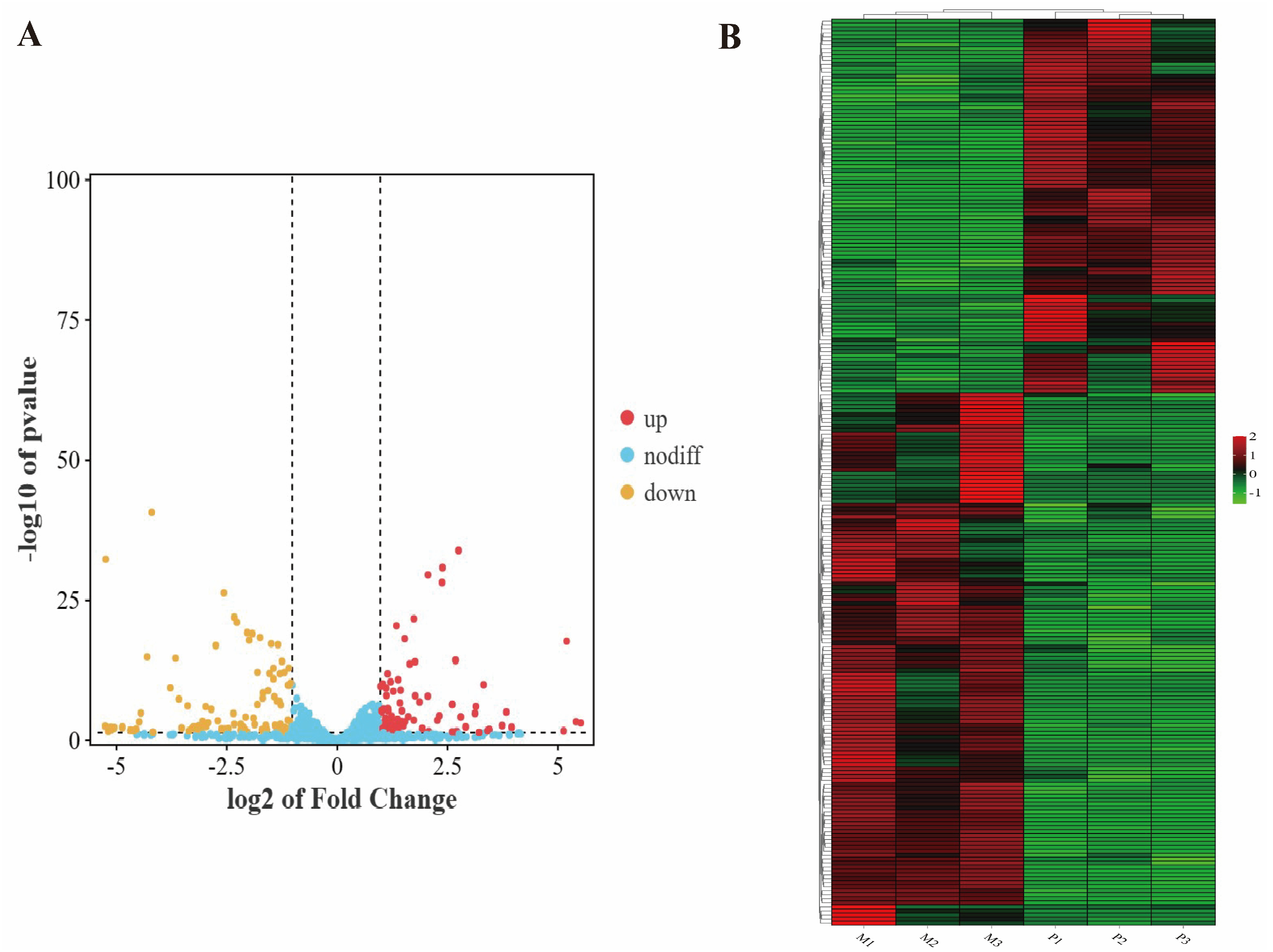
3.2. Differential Expression Analysis of miRNAs
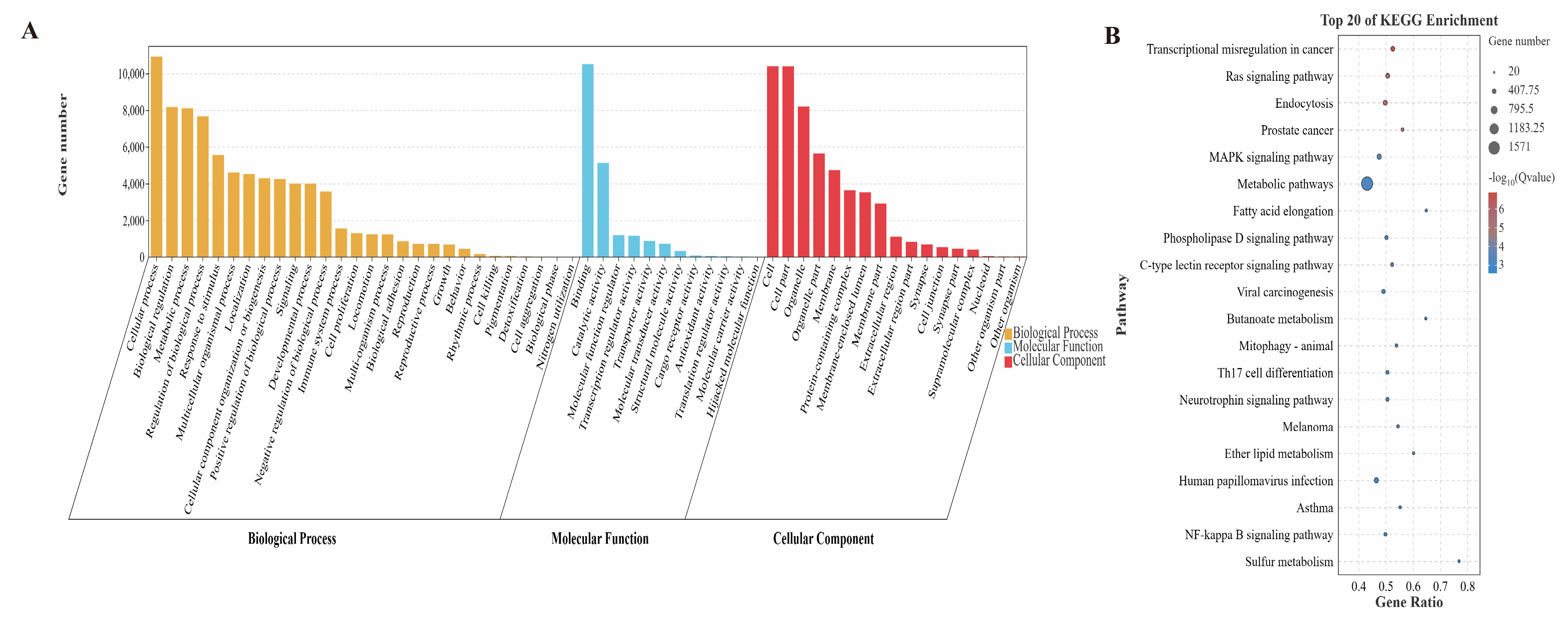
3.3. Target Gene Prediction of Differentially Expressed miRNAs and KEGG and GO Analysis
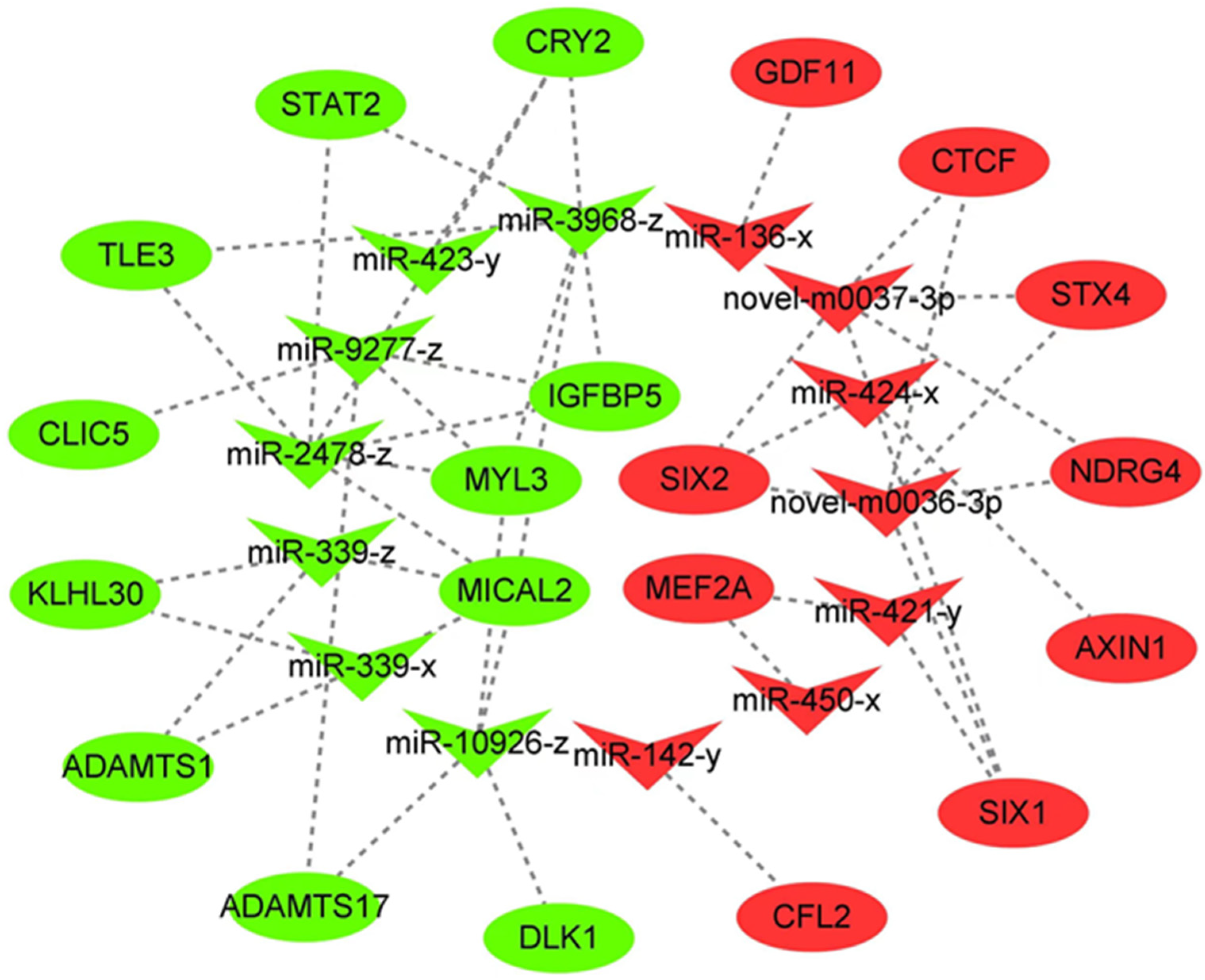
3.4. Building the Network for miRNA–mRNA Interactions
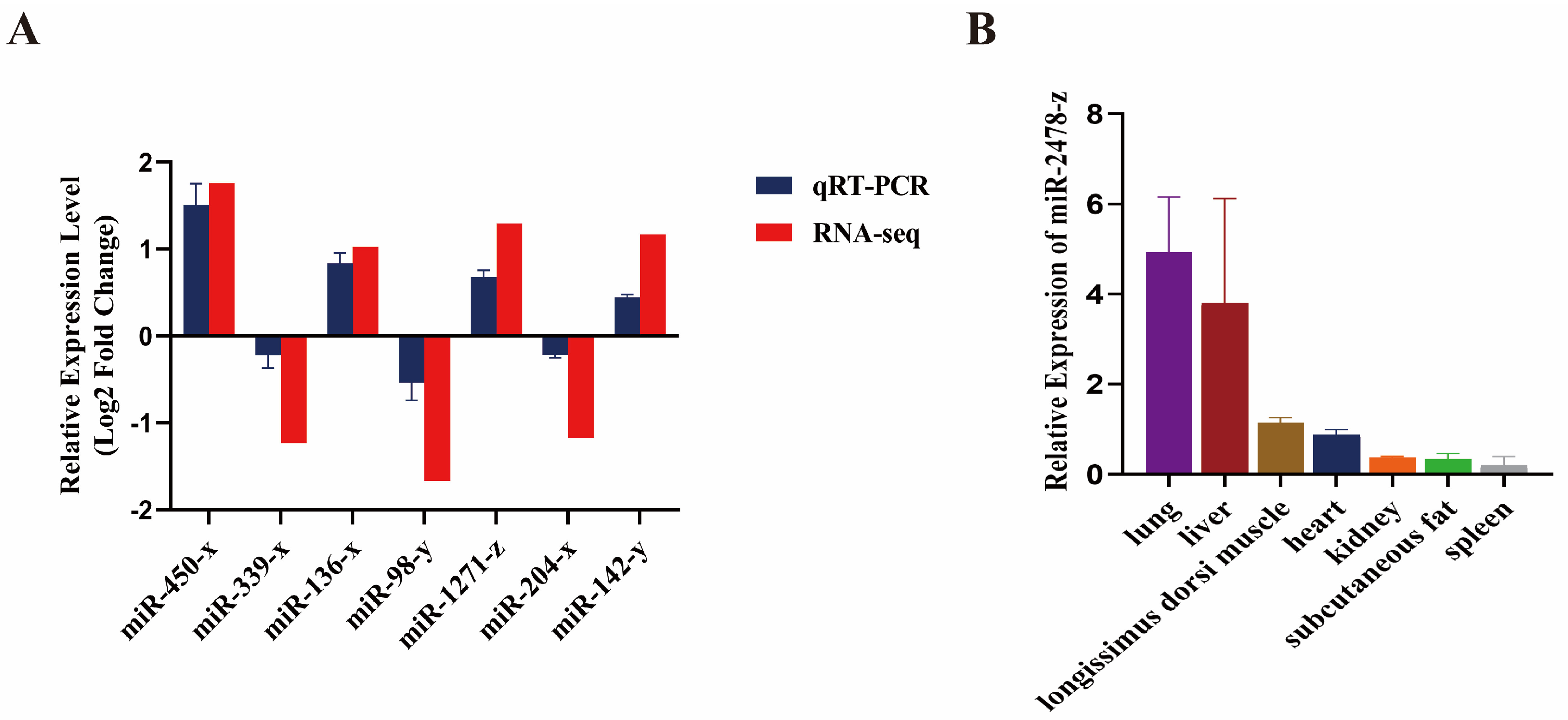
3.5. Validation of Differentially Expressed miRNAs by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, S.Z.; Ma, D.L.; Li, B.M.; Baozhaxi, J.C.; Zhang, T.Y.; Xu, G.Q.; Zhang, H.X.; Wang, L.B.; Lamao, J.B.; Wang, W.B.; et al. Determination of growth and development indicators of Jeryak in Gannan alpine pasture area. Chin. Herbiv. Sci. 2019, 39, 73–75. [Google Scholar]
- Wei, Y.P.; Xu, J.T. Current status of cattle-yak production and research in Qinghai Province. Chin. Bov. Sci. 2010, 36, 60–62. [Google Scholar]
- Yin, H.; He, H.; Cao, X.; Shen, X.; Han, S.; Cui, C.; Zhao, J.; Wei, Y.; Chen, Y.; Xia, L.; et al. MiR-148a-3p Regulates Skeletal Muscle Satellite Cell Differentiation and Apoptosis via the PI3K/AKT Signaling Pathway by Targeting Meox2. Front. Genet. 2020, 11, 512. [Google Scholar] [CrossRef]
- Liu, J.; Li, F.; Hu, X.; Cao, D.; Liu, W.; Han, H.; Zhou, Y.; Lei, Q. Deciphering the miRNA transcriptome of breast muscle from the embryonic to post-hatching periods in chickens. BMC Genom. 2021, 22, 64. [Google Scholar] [CrossRef]
- Luo, W.; Abdalla, B.A.; Nie, Q.; Zhang, X. The genetic regulation of skeletal muscle development:insights from chicken studies. Front. Agric. Sci. Eng. 2017, 4, 295–304. [Google Scholar] [CrossRef]
- Hernandez-Hernandez, J.M.; Garcia-Gonzalez, E.G.; Brun, C.E.; Rudnicki, M.A. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.V.; Hughes, S.M. Mef2 and the skeletal muscle differentiation program. Semin. Cell Dev. Biol. 2017, 72, 33–44. [Google Scholar] [CrossRef]
- Zhang, R.M.; Pan, Y.; Zou, C.X.; An, Q.; Cheng, J.R.; Li, P.J.; Zheng, Z.H.; Pan, Y.; Feng, W.Y.; Yang, S.F.; et al. CircUBE2Q2 promotes differentiation of cattle muscle stem cells and is a potential regulatory molecule of skeletal muscle development. BMC Genom. 2022, 23, 267. [Google Scholar] [CrossRef]
- Buckingham, M.; Relaix, F. PAX3 and PAX7 as upstream regulators of myogenesis. Semin. Cell Dev. Biol. 2015, 44, 115–125. [Google Scholar] [CrossRef]
- Gao, L.; Yang, M.; Wei, Z.; Gu, M.; Yang, L.; Bai, C.; Wu, Y.; Li, G. MSTN Mutant Promotes Myogenic Differentiation by Increasing Demethylase TET1 Expression via the SMAD2/SMAD3 Pathway. Int. J. Biol. Sci. 2020, 16, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pan, M.; Huang, D.; Guo, Y.; Yang, M.; Zhang, W.; Mai, K. Myostatin-1 Inhibits Cell Proliferation by Inhibiting the mTOR Signal Pathway and MRFs, and Activating the Ubiquitin-Proteasomal System in Skeletal Muscle Cells of Japanese Flounder Paralichthys olivaceus. Cells 2020, 9, 2376. [Google Scholar] [CrossRef] [PubMed]
- Tajbakhsh, S.; Borello, U.; Vivarelli, E.; Kelly, R.; Papkoff, J.; Duprez, D.; Buckingham, M.; Cossu, G. Differential activation of Myf5 and MyoD by different Wnts in explants of mouse paraxial mesoderm and the later activation of myogenesis in the absence of Myf5. Development 1998, 125, 4155–4162. [Google Scholar] [CrossRef] [PubMed]
- Elia, D.; Madhala, D.; Ardon, E.; Reshef, R.; Halevy, O. Sonic hedgehog promotes proliferation and differentiation of adult muscle cells: Involvement of MAPK/ERK and PI3K/Akt pathways. Biochim. Biophys. Acta 2007, 1773, 1438–1446. [Google Scholar] [CrossRef]
- Otto, A.; Schmidt, C.; Luke, G.; Allen, S.; Valasek, P.; Muntoni, F.; Lawrence-Watt, D.; Patel, K. Canonical Wnt signalling induces satellite-cell proliferation during adult skeletal muscle regeneration. J. Cell Sci. 2008, 121, 2939–2950. [Google Scholar] [CrossRef]
- Abu-Elmagd, M.; Robson, L.; Sweetman, D.; Hadley, J.; Francis-West, P.; Munsterberg, A. Wnt/Lef1 signaling acts via Pitx2 to regulate somite myogenesis. Dev. Biol. 2010, 337, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Gao, F.; Wen, L.; Ouyang, M.; Wang, Y.; Wang, Q.; Luo, L.; Jian, Z. Osteocalcin Induces Proliferation via Positive Activation of the PI3K/Akt, P38 MAPK Pathways and Promotes Differentiation Through Activation of the GPRC6A-ERK1/2 Pathway in C2C12 Myoblast Cells. Cell. Physiol. Biochem. 2017, 43, 1100–1112. [Google Scholar] [CrossRef]
- Kornasio, R.; Riederer, I.; Butler-Browne, G.; Mouly, V.; Uni, Z.; Halevy, O. Beta-hydroxy-beta-methylbutyrate (HMB) stimulates myogenic cell proliferation, differentiation and survival via the MAPK/ERK and PI3K/Akt pathways. Biochim. Biophys. Acta 2009, 1793, 755–763. [Google Scholar] [CrossRef]
- Ghini, F.; Rubolino, C.; Climent, M.; Simeone, I.; Marzi, M.J.; Nicassio, F. Endogenous transcripts control miRNA levels and activity in mammalian cells by target-directed miRNA degradation. Nat. Commun. 2018, 9, 3119. [Google Scholar] [CrossRef]
- Kooshapur, H.; Choudhury, N.R.; Simon, B.; Muhlbauer, M.; Jussupow, A.; Fernandez, N.; Jones, A.N.; Dallmann, A.; Gabel, F.; Camilloni, C.; et al. Structural basis for terminal loop recognition and stimulation of pri-miRNA-18a processing by hnRNP A1. Nat. Commun. 2018, 9, 2479. [Google Scholar] [CrossRef]
- Khatri, B.; Seo, D.; Shouse, S.; Pan, J.H.; Hudson, N.J.; Kim, J.K.; Bottje, W.; Kong, B.C. MicroRNA profiling associated with muscle growth in modern broilers compared to an unselected chicken breed. BMC Genom. 2018, 19, 683. [Google Scholar] [CrossRef]
- Li, Z.; Abdalla, B.A.; Zheng, M.; He, X.; Cai, B.; Han, P.; Ouyang, H.; Chen, B.; Nie, Q.; Zhang, X. Systematic transcriptome-wide analysis of mRNA-miRNA interactions reveals the involvement of miR-142-5p and its target (FOXO3) in skeletal muscle growth in chickens. Mol. Genet. Genom. 2018, 293, 69–80. [Google Scholar] [CrossRef]
- Xu, S.; Chang, Y.; Wu, G.; Zhang, W.; Man, C. Potential role of miR-155-5p in fat deposition and skeletal muscle development of chicken. Biosci. Rep. 2020, 40, BSR20193796. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M.; Shan, Y.; Ji, G.; Ju, X.; Tu, Y.; Sheng, Z.; Xie, J.; Zou, J.; Shu, J. miRNA-mRNA network regulation in the skeletal muscle fiber phenotype of chickens revealed by integrated analysis of miRNAome and transcriptome. Sci. Rep. 2020, 10, 10619. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhang, S.; Xu, Z.; Gao, J.; Mishra, S.K.; Zhu, Q.; Zhao, X.; Wang, Y.; Yin, H.; Fan, X.; et al. MiRNA Profiling in Pectoral Muscle Throughout Pre- to Post-Natal Stages of Chicken Development. Front. Genet. 2020, 11, 570. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F.; Tao, Y.; Li, J.; Deng, Z.; Yan, Z.; Xiao, X.; Wang, D.Z. microRNA-1 and microRNA-206 regulate skeletal muscle satellite cell proliferation and differentiation by repressing Pax7. J. Cell Biol. 2010, 190, 867–879. [Google Scholar] [CrossRef]
- Nakasa, T.; Ishikawa, M.; Shi, M.; Shibuya, H.; Adachi, N.; Ochi, M. Acceleration of muscle regeneration by local injection of muscle-specific microRNAs in rat skeletal muscle injury model. J. Cell. Mol. Med. 2010, 14, 2495–2505. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Ran, L.; Lang, H.; Zhou, M.; Yu, L.; Yi, L.; Zhu, J.; Liu, L.; Mi, M. Myricetin improves endurance capacity by inducing muscle fiber type conversion via miR-499. Nutr. Metab. 2019, 16, 27. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cao, X.; Ge, L.; Gu, Y.; Lv, X.; Getachew, T.; Mwacharo, J.M.; Haile, A.; Sun, W. MiR-22-3p Inhibits Proliferation and Promotes Differentiation of Skeletal Muscle Cells by Targeting IGFBP3 in Hu Sheep. Animals 2022, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Lu, S.; Bai, M.; Xiang, L.; Li, J.; Jia, C.; Jiang, H. Integrative microRNA-mRNA Analysis of Muscle Tissues in Qianhua Mutton Merino and Small Tail Han Sheep Reveals Key Roles for oar-miR-655-3p and oar-miR-381-5p. DNA Cell Biol. 2019, 38, 423–435. [Google Scholar] [CrossRef]
- Friedlander, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Betel, D.; Koppal, A.; Agius, P.; Sander, C.; Leslie, C. Comprehensive modeling of microRNA targets predicts functional non-conserved and non-canonical sites. Genome Biol. 2010, 11, R90. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef]
- Huang, C.; Ge, F.; Ma, X.; Dai, R.; Dingkao, R.; Zhaxi, Z.; Burenchao, G.; Bao, P.; Wu, X.; Guo, X.; et al. Comprehensive Analysis of mRNA, lncRNA, circRNA, and miRNA Expression Profiles and Their ceRNA Networks in the Longissimus Dorsi Muscle of Cattle-Yak and Yak. Front. Genet. 2021, 12, 772557. [Google Scholar] [CrossRef]
- Campos, C.F.; Costa, T.C.; Rodrigues, R.; Guimaraes, S.; Moura, F.H.; Silva, W.; Chizzotti, M.L.; Paulino, P.; Benedeti, P.; Silva, F.F.; et al. Proteomic analysis reveals changes in energy metabolism of skeletal muscle in beef cattle supplemented with vitamin A. J. Sci. Food Agric. 2020, 100, 3536–3543. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Pei, J.; Xiong, L.; Guo, S.; Wang, X.; Kang, Y.; Guo, X. Analysis of Chromatin Openness in Testicle Tissue of Yak and Cattle-Yak. Int. J. Mol. Sci. 2022, 23, 15810. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.Z.; Zhang, H.X.; Lamao, J.B.; Fan, J.F.; Wang, J.F.; Yao, X.H.; Bao, Y.Q.; Ma, D.L.; Li, B.M.; Wang, W.B. Determination of slaughtering performance and meat quality of Jeryak in alpine pasture area. Chin. Herbiv. Sci. 2019, 39, 4. [Google Scholar]
- Zhao, S.; Sun, W.; Chen, S.Y.; Li, Y.; Wang, J.; Lai, S.; Jia, X. The exploration of miRNAs and mRNA profiles revealed the molecular mechanisms of cattle-yak male infertility. Front. Vet. Sci. 2022, 9, 974703. [Google Scholar] [CrossRef]
- Mitin, N.; Kudla, A.J.; Konieczny, S.F.; Taparowsky, E.J. Differential effects of Ras signaling through NFkappaB on skeletal myogenesis. Oncogene 2001, 20, 1276–1286. [Google Scholar] [CrossRef]
- Yue, J.; Lopez, J.M. Understanding MAPK Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [PubMed]
- Bengal, E.; Aviram, S.; Hayek, T. p38 MAPK in Glucose Metabolism of Skeletal Muscle: Beneficial or Harmful? Int. J. Mol. Sci. 2020, 21, 6480. [Google Scholar] [CrossRef] [PubMed]
- Galpin, A.J.; Raue, U.; Jemiolo, B.; Trappe, T.A.; Harber, M.P.; Minchev, K.; Trappe, S. Human skeletal muscle fiber type specific protein content. Anal. Biochem. 2012, 425, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Gao, P.; Li, Z.; Dai, A.; Yang, M.; Chen, S.; Su, J.; Deng, Z.; Li, L. Forkhead Box O Signaling Pathway in Skeletal Muscle Atrophy. Am. J. Pathol. 2022, 192, 1648–1657. [Google Scholar] [CrossRef] [PubMed]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Stavnezer, E. Ski regulates muscle terminal differentiation by transcriptional activation of Myog in a complex with Six1 and Eya3. J. Biol. Chem. 2009, 284, 2867–2879. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.X.; Gong, Z.A.; Zhang, W.T.; Liu, K.; Li, T.; Shao, S.L.; Zhang, W.W. [Effects of transcription factor SIX2 gene on the proliferation of bovine skeletal muscle satellite cells]. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2022, 38, 622–627. [Google Scholar]
- Zhu, M.; Zheng, R.; Guo, Y.; Zhang, Y.; Zuo, B. NDRG4 promotes myogenesis via Akt/CREB activation. Oncotarget 2017, 8, 101720–101734. [Google Scholar] [CrossRef]
- Yoo, M.; Kim, B.G.; Lee, S.J.; Jeong, H.J.; Park, J.W.; Seo, D.W.; Kim, Y.K.; Lee, H.Y.; Han, J.W.; Kang, J.S.; et al. Syntaxin 4 regulates the surface localization of a promyogenic receptor Cdo thereby promoting myogenic differentiation. Skelet. Muscle 2015, 5, 28. [Google Scholar] [CrossRef]
- Fedoriw, A.M.; Stein, P.; Svoboda, P.; Schultz, R.M.; Bartolomei, M.S. Transgenic RNAi reveals essential function for CTCF in H19 gene imprinting. Science 2004, 303, 238–240. [Google Scholar] [CrossRef]
- Delgado-Olguin, P.; Brand-Arzamendi, K.; Scott, I.C.; Jungblut, B.; Stainier, D.Y.; Bruneau, B.G.; Recillas-Targa, F. CTCF promotes muscle differentiation by modulating the activity of myogenic regulatory factors. J. Biol. Chem. 2011, 286, 12483–12494. [Google Scholar] [CrossRef] [PubMed]
- Krek, A.; Grun, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; Macmenamin, P.; Da, P.I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNAs | Forward (5′ → 3′) | Reverse (5′ → 3′) |
|---|---|---|
| miR-450-x | TTTTGCAATATGTTCCTGAAT | |
| miR-136-x | ACTCCATTTGTTTTGATGATGG | |
| miR-1271-z | CTTGGCACCTAGTAAGTACTCAA | |
| miR-142-y | TGTAGTGTTTCCTACTTTATGG | |
| miR-204-x | TTCCCTTTGTCATCCTATGCCT | |
| miR-98-y | CTATACAACTTACTACTTTCCT | |
| miR-339-x | TCCCTGTCCTCCAGGAGCTCACT | |
| U6 | ACGGACAGGATTGACAGATT | TCGCTCCACCAACTAAGA |
| Samples | Clean Reads | High Quality | 3′ Adapter Null | Insert Null | 5′ Adapter Contaminants | PolyA (%) | Clean Tags |
|---|---|---|---|---|---|---|---|
| M1 | 17,408,914 (100%) | 17,260,011 (99.1447%) | 9612 (0.0557%) | 108,519 (0.6287%) | 31,981 (0.1853%) | 321 (0.0019%) | 16,255,077 (93.3721%) |
| M2 | 14,015,050 (100%) | 13,899,769 (99.1774%) | 6900 (0.0496%) | 54,327 (0.3908%) | 13,958 (0.1004%) | 162 (0.0012%) | 13,362,108 (95.3411%) |
| M3 | 16,597,591 (100%) | 16,448,657 (99.1027%) | 10,968 (0.0667%) | 83,221 (0.5059%) | 20,095 (0.1222%) | 247 (0.0015%) | 15,172,291 (91.4126%) |
| P1 | 14,318,748 (100%) | 14,151,985 (98.8354%) | 9475 (0.0670%) | 59,453 (0.4201%) | 7319 (0.0517%) | 126 (0.0009%) | 13,782,038 (96.2517%) |
| P2 | 9,195,586 (100%) | 9,071,110 (98.6464%) | 76,171 (0.8397%) | 46,305 (0.5105%) | 3837 (0.0423%) | 76 (0.0008%) | 8,756,687 (95.2271%) |
| P3 | 16,740,804 (100%) | 16,610,388 (99.2210%) | 66,607 (0.4010%) | 74,568 (0.4489%) | 6918 (0.0416%) | 134 (0.0008%) | 16,180,410 (96.6525%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, D.; Wei, Y.; Li, X.; Bai, Y.; Liu, Z.; Li, J.; Chen, Z.; Shi, B.; Zhang, X.; Zhao, Z.; et al. Comprehensive Analysis of miRNA and mRNA Expression Profiles during Muscle Development of the Longissimus Dorsi Muscle in Gannan Yaks and Jeryaks. Genes 2023, 14, 2220. https://doi.org/10.3390/genes14122220
Guo D, Wei Y, Li X, Bai Y, Liu Z, Li J, Chen Z, Shi B, Zhang X, Zhao Z, et al. Comprehensive Analysis of miRNA and mRNA Expression Profiles during Muscle Development of the Longissimus Dorsi Muscle in Gannan Yaks and Jeryaks. Genes. 2023; 14(12):2220. https://doi.org/10.3390/genes14122220
Chicago/Turabian StyleGuo, Dashan, Yali Wei, Xupeng Li, Yanbin Bai, Zhanxin Liu, Jingsheng Li, Zongchang Chen, Bingang Shi, Xiaolan Zhang, Zhidong Zhao, and et al. 2023. "Comprehensive Analysis of miRNA and mRNA Expression Profiles during Muscle Development of the Longissimus Dorsi Muscle in Gannan Yaks and Jeryaks" Genes 14, no. 12: 2220. https://doi.org/10.3390/genes14122220