
Advancements in Research on Prevention and Control Strategies for Maize White Spot Disease

Abstract
:1. Introduction
Occurrence, Influencing Factors and Severity of MWS in Maize
2. Pathogen and Characteristics of MWS
2.1. Pathogens
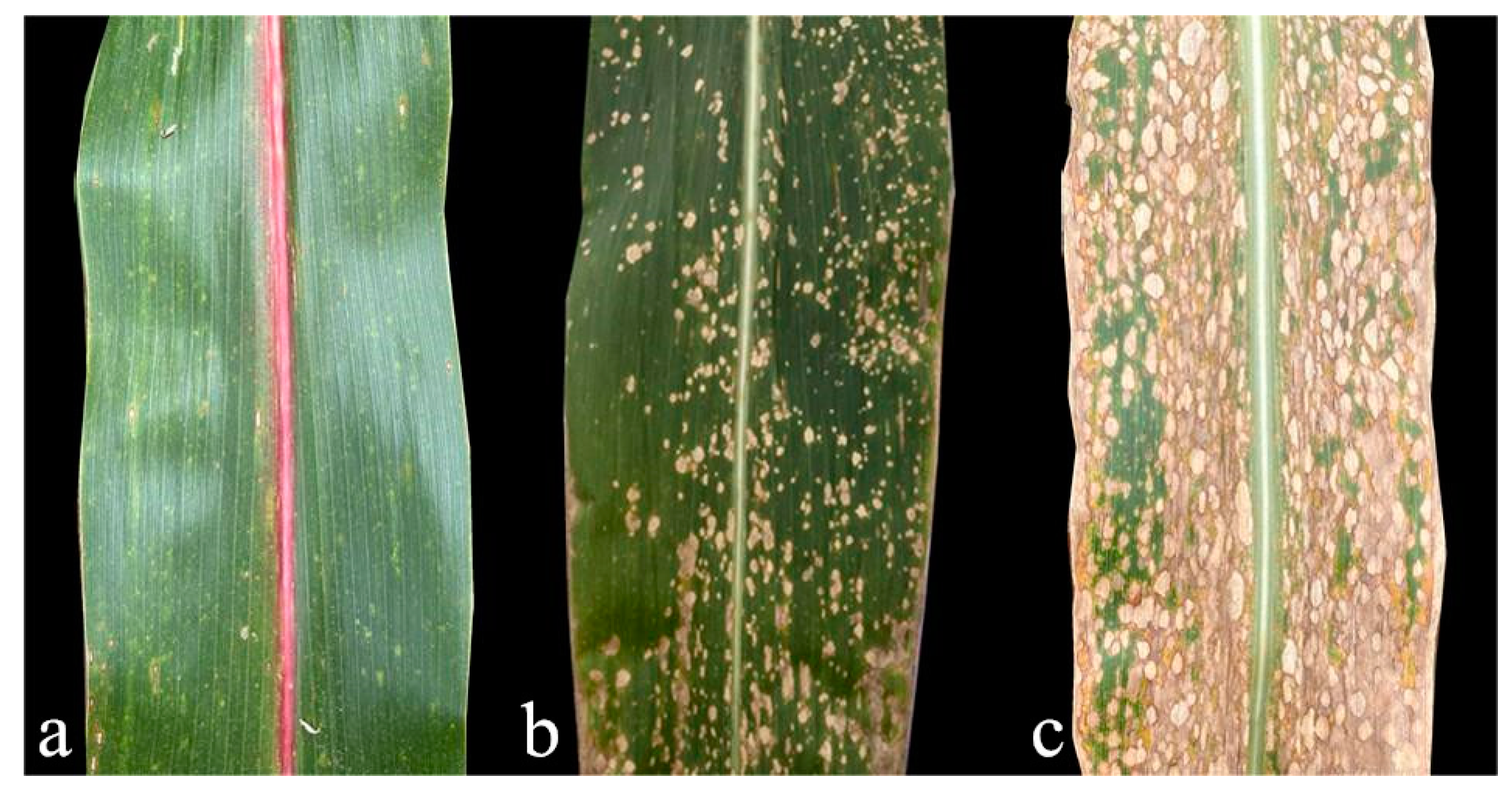
2.2. Features of the Lesions
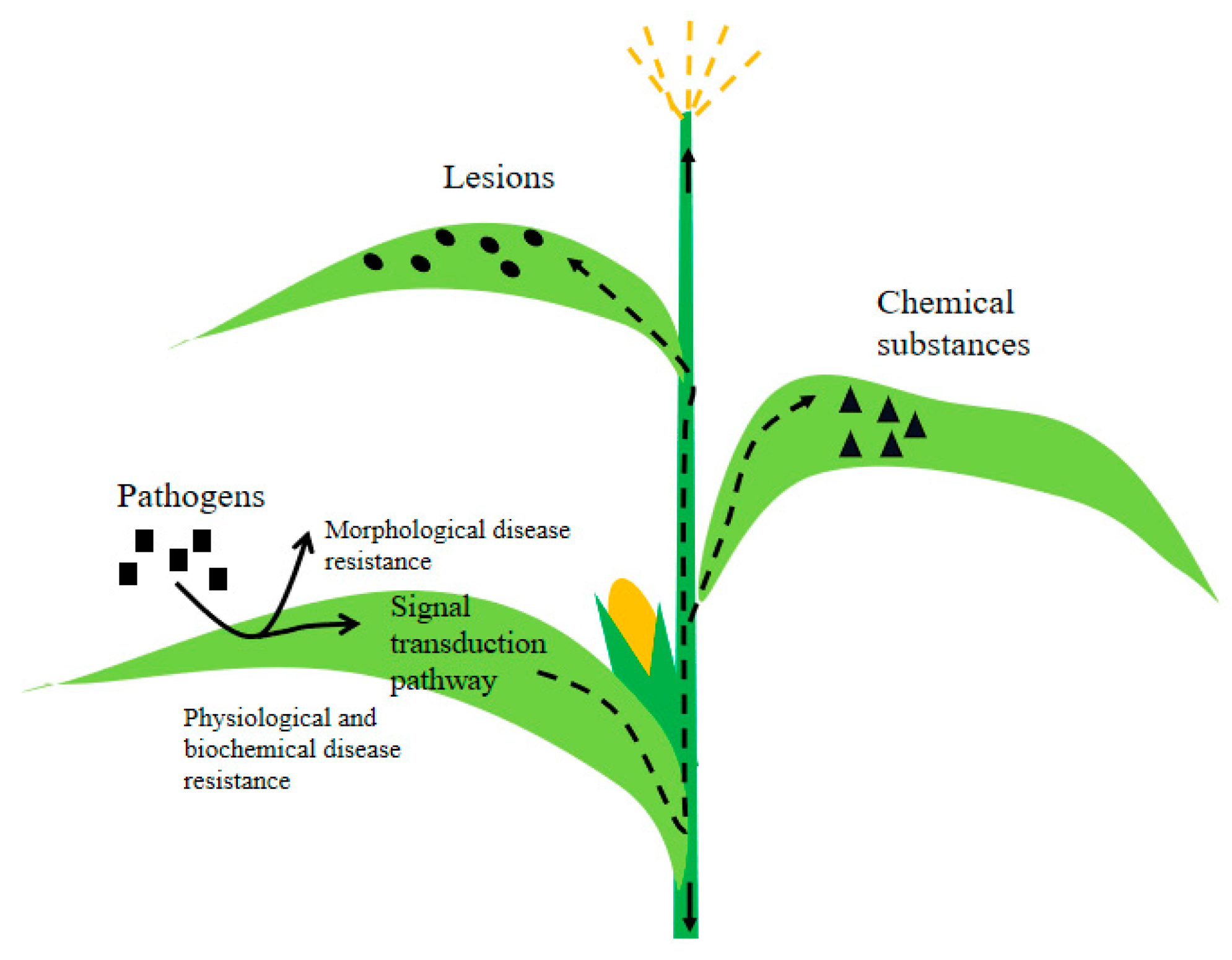
3. Plant Immunity against Pathogens
4. Research Progress on the Identification of Resistance Germplasm and Mapping of Resistance Genes
4.1. Resistant Inbred Lines
4.2. Mapping of Resistance Genes
5. Route of Transmission and Control Strategies for MWS Disease
5.1. Transmission Routes
5.2. Prevention and Control Strategies
6. Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rane, M.S.; Payak, M.M.; Renfro, B.L. A Phaeosphaeria leaf spot of maize. Indian Phytopathol. 1966, 3, 8–10. [Google Scholar]
- Paccola-Meirelles; Ferreira; Meirelles; Marriel; Casela. Detection of a Bacterium Associated with a Leaf Spot Disease of Maize in Brazil. J. Phytopathol. 2001, 149, 275–279. [Google Scholar] [CrossRef]
- Carson, M.L. Vulnerability of U.S. Maize Germ Plasm to Phaeosphaeria Leaf Spot. Plant Dis. 1999, 83, 462–464. [Google Scholar] [CrossRef]
- Derera, J.; Tongoona, P.; Vivek, B.S.; Van Rij, N.; Laing, M.D. Gene Action Determining Phaeosphaeria Leaf Spot Disease Resistance in Experimental Maize Hybrids. S. Afr. J. Plant Soil 2007, 24, 138–143. [Google Scholar] [CrossRef]
- Adam, E.; Deng, H.; Odindi, J.; Abdel-Rahman, E.M.; Mutanga, O. Detecting the Early Stage of Phaeosphaeria Leaf Spot Infestations in Maize Crop Using In Situ Hyperspectral Data and Guided Regularized Random Forest Algorithm. J. Spectrosc. 2017, 2017, 6961387. [Google Scholar] [CrossRef]
- Sibiya, J.; Tongoona, P.; Derera, J.; Van Rij, N.; Makanda, I. Combining Ability Analysis for Phaeosphaeria Leaf Spot Resistance and Grain Yield in Tropical Advanced Maize Inbred Lines. Field Crops Res. 2011, 120, 86–93. [Google Scholar] [CrossRef]
- Pérez-y-Terrón, R.; Villegas, M.C.; Cuellar, A.; Muñoz-Rojas, J.; Castañeda-Lucio, M.; Hernández-Lucas, I.; Bustillos-Cristales, R.; Bautista-Sosa, L.; Munive, J.A.; Caicedo-Rivas, R.; et al. Detection of Pantoea ananatis, causal agent of leaf spot disease of maize, in Mexico. Australas Plant Dis. Notes 2009, 4, 96–99. [Google Scholar] [CrossRef]
- Alippi, A.M.; López, A.C. First Report of Leaf Spot Disease of Maize Caused by Pantoea ananatis in Argentina. Plant Dis. 2010, 94, 487. [Google Scholar] [CrossRef]
- Krawczyk, K.; Kamasa, J.; Zwolinska, A.; Pospieszny, H. First Report of Pantoea ananatis Associated with Leaf Spot Disease of Maize in Poland. J. Plant Pathol. 2010, 92, 807–811. [Google Scholar]
- Toaza, A.; Caiza, R.B.; Garrido, A.D.; Moreno, C.; Guevara, J.L.; Regalado, H.; Ramos, L.; Flores, F.; Garrido, P. First report of Pantoea ananatis causing leaf spot disease of maize in Ecuador. Plant Dis. 2021, 105, 3286. [Google Scholar] [CrossRef]
- Wang, D.; Liu, P.F.; Guo, X.Y.; Wang, A.G.; Zhu, Y.F.; Wu, X.; Chen, Z.H. Research progress on maize white spot disease and its resistance gene mapping [J/OL]. Mol. Plant Breed. 2023, 1–21. Available online: https://kns.cnki.net/kcms/detail/46.1068.S.20220124.2027.016.html (accessed on 1 October 2023).
- Fan, J.J.; Liang, X.G.; Lou, Y.; Du, Y.; Li, X.Y.; Zhang, S.H. Preliminary report on the occurrence of corn white spot and cloud spot in Wenshan Prefecture. Yunnan Agric. Sci. Technol. 2022, S1, 12–13. [Google Scholar]
- Zhang, S.; Zhu, Z.P.; He, P.B.; Li, Y.M.; He, P.F.; Chen, Q.; Chang, W.; Wu, Y.X.; Liu, Y.L.; Tan, W.Z.; et al. Identification of Pathogenic Bacteria and Evaluation of Field Resistance of Maize White Spot Disease in Yunnan Province. J. Plant Prot. 2022, 49, 840–847. [Google Scholar]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.M.; Figueiredo, J.E.F.; Chaves, C.L.; Ruas, E.A.; Balbi-Peña, M.I.; Colauto, N.B.; Paccola-Meirelles, L.D. Genomic Variability of Pantoea Ananatis in Maize White Spot Lesions Assessed by AFLP Markers. Genet. Mol. Res. 2016, 15, 15049452. [Google Scholar] [CrossRef]
- Carson, M.L. Yield Loss Potential of Phaeosphaeria Leaf Spot of Maize Caused by Phaeosphaeria maydis in the United States. Plant Dis. 2005, 89, 986–988. [Google Scholar] [CrossRef]
- Wisser, R.J.; Balint-Kurti, P.J.; Nelson, R.J. The Genetic Architecture of Disease Resistance in Maize: A Synthesis of Published Studies. Phytopathology 2006, 96, 120–129. [Google Scholar] [CrossRef]
- Moreira, J.U.V.; Bento, D.A.V.; De Souza, A.P.; De Souza, C.L. QTL Mapping for Reaction to Phaeosphaeria Leaf Spot in a Tropical Maize Population. Theor. Appl. Genet. 2009, 119, 1361–1369. [Google Scholar] [CrossRef]
- Pinto, N. Controle químico de doenças foliares em milho. Rev. Bras. De Milho E Sorgo 2004, 3, 134–138. [Google Scholar] [CrossRef]
- Sauer, A.V.; Rocha, K.R.; Gonçalves, R.M.; Meirelles, W.F.; Figueiredo, J.E.F.; Marriel, I.E.; Paccola-Meirelles, L.D. Survival of Pantoea Ananatis, Causal Agent of Maize White Spot Disease in Crop Debris. ASB J. 2015, 1, 21. [Google Scholar] [CrossRef]
- Bomfeti, C.A.; Souza-Paccola, E.A.; Massola Júnior, N.S.; Júnior, M.; Marriel, I.; Meirelles, W.; Casela, C.; Paccola-Meirelles, L. Localization of Pantoea ananatis inside lesions of maize white spot disease using transmission electron microscopy and molecular techniques. Trop. Plant Pathol. 2008, 33, 63–66. [Google Scholar] [CrossRef]
- Capucho, A.S.; Zambolim, L.; Duarte, H.S.S.; Parreira, D.F.; Ferreira, P.A.; Lanza, F.E.; Costa, R.V.; Casela, C.R.; Cota, L.V. Influence of leaf position that correspond to whole plant severity and diagrammatic scale for white spot of corn. Crop Prot. 2010, 29, 1015–1020. [Google Scholar] [CrossRef]
- Pegoraro, D.G.; Vacaro, E.; Nuss, C.N.; Dal Soglio, F.K.; Sereno, M.J.C.D.M.; Barbosa Neto, J.F. Efeito de época de semeadura e adubação na mancha-foliar de Phaeosphaeria em milho. Pesq. Agropec. Bras. 2001, 36, 1037–1042. [Google Scholar] [CrossRef]
- Tirado, M.C.; Clarke, R.; Jaykus, L.A.; McQuatters-Gollop, A.; Frank, J.M. Climate Change and Food Safety: A Review. Food Res. Int. 2010, 43, 1745–1765. [Google Scholar] [CrossRef]
- Vieira, R.A.; Tessmann, D.J.; Scapim, C.A.; Hata, F.T.; Rodovalho, M.A.; Barreto, R.R. Genetic Resistance of New Popcorn Hybrids to Foliar Diseases. Crop Breed. Appl. Biotechnol. 2009, 9, 140–146. [Google Scholar] [CrossRef]
- Chakraborty, S.; Tiedemann, A.V.; Teng, P.S. Climate Change: Potential Impact on Plant Diseases. Environ. Pollut. 2000, 108, 317–326. [Google Scholar] [CrossRef]
- Cervelatti, E.P.; Paiva, E.; Meirelles, W.F.; Casela, C.R.; Fernandes, F.T.; Teixeira, F.F.; Paccola-Meirelles, L.D. Characterization of Fungal Isolates from Pycnidia and Pseudothecia from Lesions of Phaeosphaeria Leaf Spot in Maize. Rev. Bras. Milho E Sorgo 2002, 1, 30–37. [Google Scholar] [CrossRef]
- Bomfeti, C.A.; Meirelles, W.F.; Souza-Paccola, E.A.; Casela, C.; Ferreira, A.; Marriel, I.; Paccola-Meirelles, L.D. Avaliação de produtos químicos comerciais, in vitro e in vivo, no controle da doença foliar, mancha branca do milho, causada por Pantoea ananatis. Summa Phytopathol. 2007, 33, 63–67. [Google Scholar] [CrossRef]
- Gonçalves, R.M.; Figueiredo, J.E.; Pedro, E.S.; Meirelles, W.F.; Leite Junior, R.P.; Sauer, A.V.; Paccola-Meirelles, L.D. Etiology of Phaeosphaeria leaf spot disease of maize. J. Plant Pathol. 2013, 95, 559–569. [Google Scholar]
- Lanza, F.E. Mancha-Branca do Milho: Etiologia e Resistência de Genótipos. Master’s Thesis, Universidade Federal de Viçosa, Minas Gerais, Brazil, 2009. [Google Scholar]
- Pomini, A.M.; Paccola-Meirelles, L.D.; Marsaioli, A.J. Acyl-Homoserine Lactones Produced by Pantoea sp. Isolated from the “Maize White Spot” Foliar Disease. J. Agric. Food Chem. 2007, 55, 1200–1204. [Google Scholar] [CrossRef]
- Do Amaral, A.L.; Dal Soglio, F.K.; De Carli, M.L.; Neto, J.F.B. Pathogenic Fungi Causing Symptoms Similar to Phaeosphaeria Leaf Spot of Maize in Brazil. Plant Dis. 2005, 89, 44–49. [Google Scholar] [CrossRef]
- Zou, C.J.; Wang, J.; Li, X.; Zhang, H.Y.; Zhang, Z.Y.; Cui, L.N.; Liu, S.J. Epidemic warning of maize new leaf spot disease in southwestern and southern regions. Sichuan Agric. Sci. Technol. 2021, 5, 37–39. [Google Scholar]
- Escanferla, M.; Wysmierski, P.; Meirelles, W.; Meirelles, L. Viability and Dissemination of Pantoea Ananatis, Etiological Agent of Maize White Spot Disease. ASB J. 2018, 4, 52. [Google Scholar] [CrossRef]
- Lana, U.G.D.P.; Gomes, E.A.; Silva, D.D.; Costa, R.V.; Cota, L.V.; Parreira, D.F.; Souza, I.R.P.; Guimarães, C.T. Detection and Molecular Diversity of Pantoea ananatis Associated with White Spot Disease in Maize, Sorghum and Crabgrass in Brazil. J. Phytopathol. 2012, 160, 441–448. [Google Scholar] [CrossRef]
- De Maayer, P.; Chan, W.; Rubagotti, E.; Venter, S.N.; Toth, I.K.; Birch, P.R.J.; Coutinho, T.A. Analysis of the Pantoea Ananatis Pan-Genome Reveals Factors Underlying Its Ability to Colonize and Interact with Plant, Insect and Vertebrate Hosts. BMC Genom. 2014, 15, 404. [Google Scholar] [CrossRef] [PubMed]
- Walcott, R.R.; Gitaitis, R.D.; Castro, A.C.; Sanders, F.H.; Diaz-Perez, J.C. Natural Infestation of Onion Seed by Pantoea ananatis, Causal Agent of Center Rot. Plant Dis. 2002, 86, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Gitaitis, R.; Walcott, R.; Culpepper, S.; Sanders, H.; Zolobowska, L.; Langston, D. Recovery of Pantoea Ananatis, Causal Agent of Center Rot of Onion, from Weeds and Crops in Georgia, USA. Crop Prot. 2002, 21, 983–989. [Google Scholar] [CrossRef]
- Kido, K.; Hasegawa, M.; Matsumoto, H.; Kobayashi, M.; Takikawa, Y. Pantoea Ananatis Strains Are Differentiated into Three Groups Based on Reactions of Tobacco and Welsh Onion and on Genetic Characteristics. J. Gen. Plant Pathol. 2010, 76, 208–218. [Google Scholar] [CrossRef]
- Cota, L.V.; Costa, R.V.; Silva, D.D.; Parreira, D.F.; Lana, U.G.P.; Casela, C.R. First Report of Pathogenicity of Pantoea Ananatis in Sorghum (Sorghum bicolor) in Brazil. Austral. Plant Disease Notes 2010, 5, 120. [Google Scholar] [CrossRef]
- Cother, E.J.; Noble, D.H.; Van De Ven, R.J.; Lanoiselet, V.; Ash, G.; Vuthy, N.; Visarto, P.; Stodart, B. Bacterial Pathogens of Rice in the Kingdom of Cambodia and Description of a New Pathogen Causing a Serious Sheath Rot Disease. Plant Pathol. 2010, 59, 944–953. [Google Scholar] [CrossRef]
- Zhang, F.Y.; Zeng, Y.J. Research progress on plant resistance. For. Sci. Technol. Inf. 2019, 1, 1–3. [Google Scholar]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Nguyen, C.T.; Liang, Y.; Cao, Y.; Stacey, G. Role of LysM Receptors in Chitin-Triggered Plant Innate Immunity. Plant Signal. Behav. 2013, 8, e22598. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.H.; Yang, J.W.; Wang, Y.; Li, G.F.; Duan, C.X.; Jia, X.; Wang, J.J. Research progress in gene mining and molecular breeding utilization of corn resistance to fungal diseases. J. Crops 2023, 3, 1–11. [Google Scholar]
- Qin, C.X.; Li, Y.; Zhang, Y.X.; Dai, S.J. The role of LysM protein in plant immunity. J. Shanghai Norm. Univ. Nat. Sci. Ed. 2022, 51, 7. [Google Scholar] [CrossRef]
- Chiang, Y.H.; Coaker, G. Effector triggered immunity: NLR immune perception and downstream defense responses. Arab. Book 2015, 13, e0183. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Ding, Y.; Zheng, J.Q.; Wan, B.J.; Cao, T.; Liu, X.H. Research progress on immune signaling pathways mediated by plant PRRs and NLRs. Jiangsu Agric. Sci. 2023, 51, 43–50. [Google Scholar] [CrossRef]
- Falk, A.; Feys, B.J.; Frost, L.N.; Jones, J.D.G.; Daniels, M.J.; Parker, J.E. EDS1, an Essential Component of R Gene-Mediated Disease Resistance in Arabidopsis Has Homology to Eukaryotic Lipases. Proc. Natl. Acad. Sci. USA 1999, 96, 3292–3297. [Google Scholar] [CrossRef]
- Parker, J.E.; Feys, B.J.; Van Der Biezen, E.A.; Noël, L.; Aarts, N.; Austin, M.J.; Botella, M.A.; Frost, L.N.; Daniels, M.J.; Jones, J.D.G. Unravelling R Gene-mediated Disease Resistance Pathways in Arabidopsis. Mol. Plant Pathol. 2000, 1, 17–24. [Google Scholar] [CrossRef]
- Fang, X. Functional Study on a Plant Disease Resistance Gene. Master’s Thesis, Hefei University of Technology, Hefei, China, 2023. [Google Scholar]
- Kourelis, J.; Van Der Hoorn, R.A.L. Defended to the Nines: 25 Years of Resistance Gene Cloning Identifies Nine Mechanisms for R Protein Function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef]
- Johal, G.S.; Briggs, S.P. Reductase Activity Encoded by the HM1 Disease Resistance Gene in Maize. Science 1992, 258, 985–987. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kars, I.; Essenstam, B.; Liebrand, T.W.H.; Wagemakers, L.; Elberse, J.; Tagkalaki, P.; Tjoitang, D.; Van Den Ackerveken, G.; Van Kan, J.A.L. Fungal Endopolygalacturonases Are Recognized as Microbe-Associated Molecular Patterns by the Arabidopsis Receptor-Like Protein Responsiveness to Botrytis Polygalacturonases1. Plant Physiol. 2014, 164, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, R.N.; Schwessinger, B.; Joe, A.; Thomas, N.; Liu, F.; Albert, M.; Robinson, M.R.; Chan, L.J.G.; Luu, D.D.; Chen, H.; et al. The Rice Immune Receptor XA21 Recognizes a Tyrosine-Sulfated Protein from a Gram-Negative Bacterium. Sci. Adv. 2015, 1, e1500245. [Google Scholar] [CrossRef]
- De Jonge, R.; Peter Van Esse, H.; Maruthachalam, K.; Bolton, M.D.; Santhanam, P.; Saber, M.K.; Zhang, Z.; Usami, T.; Lievens, B.; Subbarao, K.V.; et al. Tomato Immune Receptor Ve1 Recognizes Effector of Multiple Fungal Pathogens Uncovered by Genome and RNA Sequencing. Proc. Natl. Acad. Sci. USA 2012, 109, 5110–5115. [Google Scholar] [CrossRef]
- Luderer, R.; Takken, F.L.W.; Wit, P.J.G.M.D.; Joosten, M.H.A.J. Cladosporium fulvum Overcomes Cf-2 -mediated Resistance by Producing Truncated AVR2 Elicitor Proteins. Mol. Microbiol. 2002, 45, 875–884. [Google Scholar] [CrossRef]
- Lozano-Torres, J.L.; Wilbers, R.H.P.; Gawronski, P.; Boshoven, J.C.; Finkers-Tomczak, A.; Cordewener, J.H.G.; America, A.H.P.; Overmars, H.A.; Van ‘T Klooster, J.W.; Baranowski, L.; et al. Dual Disease Resistance Mediated by the Immune Receptor Cf-2 in Tomato Requires a Common Virulence Target of a Fungus and a Nematode. Proc. Natl. Acad. Sci. USA 2012, 109, 10119–10124. [Google Scholar] [CrossRef] [PubMed]
- Krasileva, K.V.; Dahlbeck, D.; Staskawicz, B.J. Activation of an Arabidopsis Resistance Protein Is Specified by the in Planta Association of Its Leucine-Rich Repeat Domain with the Cognate Oomycete Effector. Plant Cell 2010, 22, 2444–2458. [Google Scholar] [CrossRef] [PubMed]
- Caplan, J.L.; Mamillapalli, P.; Burch-Smith, T.M.; Czymmek, K.; Dinesh-Kumar, S.P. Chloroplastic Protein NRIP1 Mediates Innate Immune Receptor Recognition of a Viral Effector. Cell 2008, 132, 449–462. [Google Scholar] [CrossRef]
- Qi, D.; Dubiella, U.; Kim, S.H.; Sloss, D.I.; Dowen, R.H.; Dixon, J.E.; Innes, R.W. Recognition of the Protein Kinase Avrpphb Susceptible1 by the Disease Resistance Protein Resistance to Pseudomonas Syringae5 Is Dependent on S-Acylation and an Exposed Loop in Avrpphb Susceptible1. Plant Physiol. 2014, 164, 340–351. [Google Scholar] [CrossRef]
- Maqbool, A.; Saitoh, H.; Franceschetti, M.; Stevenson, C.; Uemura, A.; Kanzaki, H.; Kamoun, S.; Terauchi, R.; Banfield, M. Structural Basis of Pathogen Recognition by an Integrated HMA Domain in a Plant NLR Immune Receptor. eLife 2015, 4, e08709. [Google Scholar] [CrossRef]
- Wang, H.; Hou, J.; Ye, P.; Hu, L.; Huang, J.; Dai, Z.; Zhang, B.; Dai, S.; Que, J.; Min, H.; et al. A Teosinte-Derived Allele of a MYB Transcription Repressor Confers Multiple Disease Resistance in Maize. Mol. Plant 2021, 14, 1846–1863. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; He, Y.; Kabahuma, M.; Chaya, T.; Kelly, A.; Borrego, E.; Bian, Y.; El Kasmi, F.; Yang, L.; Teixeira, P.; et al. A Gene Encoding Maize Caffeoyl-CoA O-Methyltransferase Confers Quantitative Resistance to Multiple Pathogens. Nat. Genet. 2017, 49, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Carson, M.L. Inheritance of Resistance to Phaeosphaeria Leaf Spot of Maize. Plant Dis. 2001, 85, 798–800. [Google Scholar] [CrossRef]
- Lopes, M.T.G.; Lopes, R.; Brunelli, K.R.; Silva, H.P.D.; Matiello, R.R.; Camargo, L.E.A. Controle genético da resistência à mancha-de-Phaeosphaeria em milho. Cienc. Rural 2007, 37, 605–611. [Google Scholar] [CrossRef]
- Guimarães, P.S.; Zagatto Paterniani, M.; Dudienas, C.; Lüders, R.R.; Gallo, P.B. Capacidade combinatória para resistência àmancha branca em linhagens endogâmicas de milho. Summa Phytopathol. 2009, 35, 282–287. [Google Scholar] [CrossRef]
- Paterniani, M.E.A.G.Z.; Sawazaki, E.; Dudienas, C.; Duarte, A.P.; Gallo, P.B. Diallel Crosses among Maize Lines with Emphasis on Resistance to Foliar Diseases. Genet. Mol. Biol. 2000, 23, 381–385. [Google Scholar] [CrossRef]
- Pegoraro, D.G.; Barbosa Neto, J.F.; Dal Soglio, F.K.; Vacaro, E.; Nuss, C.N.; Conceição, L. Inheritance of the resistance to phaeosphaeria leaf spot in maize. Pesqui. Agropecuária Bras. 2002, 37, 329–336. [Google Scholar] [CrossRef]
- Silva, H.P.D.; Moro, J.R. Diallel Analysis of Maize Resistance to Phaeosphaeria Maydis. Sci. Agric. 2004, 61, 36–42. [Google Scholar] [CrossRef]
- Wang, D.; He, Y.; Nie, L.; Guo, S.; Tu, L.; Guo, X.; Wang, A.; Liu, P.; Zhu, Y.; Wu, X.; et al. Integrated IBD Analysis, GWAS Analysis and Transcriptome Analysis to Identify the Candidate Genes for White Spot Disease in Maize. IJMS 2023, 24, 10005. [Google Scholar] [CrossRef]
- Sawazaki, E.; Dudienas, C.; Paterniani, M.E.; Galvao, J.; Castro, J.L.; Pereira, J. Reação de cultivares de milho à mancha de Phaeosphaeria no Estado de São Paulo. Pesqui. Agropecuária Bras. 1997, 32, 585–589. [Google Scholar]
- Carson, M.L.; Stuber, C.W.; Senior, M.L. Quantitative Trait Loci Conditioning Resistance to Phaeosphaeria Leaf Spot of Maize Caused by Phaeosphaeria maydis. Plant Dis. 2005, 89, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Lanza, F.E.; Zambolim, L.; Casela, C.R.; Costa, R.V.; Cota, L.V.; Silva, D.D.; Figueiredo, J.E.F. Etiology and Epidemiological Variables Associated with Maize Resistance to White Spot Disease. J. Plant Pathol. 2013, 95, 349–359. [Google Scholar]
- Lopes, M.T.G.; Vieira, M.L.C.; Lopes, R.; Brunelli, K.R.; Matiello, R.R.; Da Silva, H.P.; Camargo, L.E.A. Progeny Evaluation for Resistance to Phaeosphaeria Leaf Spot in Tropical Maize. Can. J. Plant Pathol. 2011, 33, 49–53. [Google Scholar] [CrossRef]
- Lana, U.G.D.P.; Prazeres De Souza, I.R.; Noda, R.W.; Pastina, M.M.; Magalhaes, J.V.; Guimaraes, C.T. Quantitative Trait Loci and Resistance Gene Analogs Associated with Maize White Spot Resistance. Plant Dis. 2017, 101, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.S.; Kuki, M.C.; Pinto, R.J.B.; Scapim, C.A.; Faria, M.V.; De Leon, N. Genomic-Wide Association Study for White Spot Resistance in a Tropical Maize Germplasm. Euphytica 2020, 216, 15. [Google Scholar] [CrossRef]
- Kistner, M.B.; Galiano-Carneiro, A.L.; Kessel, B.; Presterl, T.; Miedaner, T. Multi-parental QTL Mapping of Resistance to White Spot of Maize (Zea mays) in Southern Brazil and Relationship to QTLs of Other Foliar Diseases. Plant Breed. 2021, 140, 801–811. [Google Scholar] [CrossRef]
- Duan, X.H.; Shao, L.B.; Wang, X.Z. Investigation and analysis of corn white spot disease in Lincang and Baoshan cities. Agric. Sci. Technol. Commun. 2022, 02, 209–211+253. [Google Scholar]
- Mamede, M.C.; Tebaldi, N.D.; Mota, L.C.B.M.; Martins, O.M.; Coelho, L. Detection of Pantoea Ananatis in Corn Seeds on Semi-Selective Medium. Trop. Plant Pathol. 2018, 43, 254–256. [Google Scholar] [CrossRef]
- Rijavec, T.; Lapanje, A.; Dermastia, M.; Rupnik, M. Isolation of Bacterial Endophytes from Germinated Maize Kernels. Can. J. Microbiol. 2007, 53, 802–808. [Google Scholar] [CrossRef]
- Azad, H.R.; Holmes, G.J.; Cooksey, D.A. A New Leaf Blotch Disease of Sudangrass Caused by Pantoea ananas and Pantoea stewartii. Plant Dis. 2000, 84, 973–979. [Google Scholar] [CrossRef]
- Dornelas, G.A.; Pozza, E.A.; Souza, P.E.D.; Costa, R.V.D.; Pozza, A.A.A.; Santos, L.A. Nitrogen and Potassium Fertilization on the Yield and Intensity of the Maize White Spot. Rev. Ceres 2015, 62, 351–359. [Google Scholar] [CrossRef]
- Brito, A.H.; Pereira, J.L.A.R.; Pinho, R.G.V.; Balestre, M. Controle Químico de Doenças Foliares e Grãos Ardidos em Milho (Zea mays L.). Rev. Bras. Milho E Sorgo 2012, 11, 49–59. [Google Scholar] [CrossRef]
- Berghetti, J.; Sheidt, B.T.; Casa, R.T.; Coelho, A.E.; Sangoi, L.; Gonçalves, M.J.; Leolato, L. Nitrogen Rates and Sowing Dates Influence The Severity of White Spot Disease and Grain Yield of Maize. Rbms 2020, 19, 13. [Google Scholar] [CrossRef]
- Duarte, R.P.; Juliatti, F.C.; de Freitas, P.T. Eficácia de Diferentes Fungicidas Na Cultura do Milho. Biosci. J. 2009, 25, 101–111. [Google Scholar]
- Holland, J. Genetic Architecture of Complex Traits in Plants. Curr. Opin. Plant Biol. 2007, 10, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Li, H.Y.; Hu, G.H.; Liu, X.Y.; Li, C.H.; Zhang, D.F.; Li, Y.; Wang, T.Y. Construction and Application of Core Germplasm for Corn Application. J. Plant Genet. Resour. 2023, 24, 911–916. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial Number | Country | Units | Resistant Material |
|---|---|---|---|
| 1 | Brazil | Home of Agriculture | CO 42, IAC Taiúba, P3041, AGM 2007, C 805, P3051, C425, Dina 70, C 701, Dina 170, XL 380 [72] |
| 2 | USA | North Carolina State University | Inbred lines with C103 or Mo17 as paternal, A619, NC258, LH213 [3,73] |
| 3 | USA | Campinas Agricultural Research Institute | L5 and L8 of CIMMYT [68] |
| 4 | Brazil | Faculty of Agronomy, Federal University of Rio Grande do Sul | LA06, LA30, AS3466, AS3477 [23,69] |
| 5 | Brazil | Syngenta Seeds LTD | DAS95, DAS41, DAS86, DAS72, DAS2 [70] |
| 6 | Brazil | Faculty of Agricultural Sciences, Federal University of Amazon | DAS95, DAS72 [66] |
| 7 | South Africa | University of KwaZulu-Natal African Centre for Crop Improvement | B23, B22, B16, CML488, CML444 [4] |
| 8 | USA | Campinas Agricultural Research Institute | PM518, IP4035, IP398 [67] |
| 9 | Brazil | Department of Agronomy, National Agricultural University of Maringa | 13 hybrids (27, 22, 05, 01, 25, 26, 18, 17, 16, 20,13, 19 and 23), IAC 112, AUDPC [25] |
| 10 | Brazil | University of Sao Paulo | L08-05F [18] |
| 11 | Brazil | Federal University of Visosa | BRS1030, BRS1035, BRS1010, L2 [30,74] |
| 12 | South Africa | University of KwaZulu-Natal African Centre for Crop Improvement | A1220-4, N3-2-3-3, CML312, CML488 [6] |
| 13 | China | Yunnan Agricultural University | Tiandan 206, DS917, Fuyu 1388, Darwin 5 [13] |
| 14 | China | Institute of Dryland Food Crops, Guizhou Academy of Agricultural Sciences | QR27, QB512 [71] |
| Test Materials | Methods | QTL/QTN |
|---|---|---|
| 158 of B73 × Mo17 F2:7 inbred lines | Composite Interval Mapping (CIM) | bin 1.06, bin 4.07, bin 7.01, bin 7.03, bin 8.07/8.08 [73] |
| F2 population of The L14-04B × L08-05F | Multiple Interval Mapping (MIM) | bin 1.03, bin 3.07~3.08, bin 4.08, bin 6.06–6.07, bin 8.00–8.02, bin 8.06–8.07 [18] |
| F2:3 population of L31.2.1.2 × L726 | Multiple Interval Mapping (MIM) | qMWS1.06, qMWS2.06, qMWS2.07, qMWS3.08, qMWS4.05, qMWS4.09, qMWS4.10, qMWS8.03, qMWS8.05 [76] |
| 183 Tropical popcorn inbred lines | Mixed Linear Models | bin 1.01, bin 1.05, bin 3.04, bin 4.02, bin 4.02, bin 5.03, bin 6.05, bin 7.02, bin 8.03 [77] |
| Hybrid offspring of 7 parents | Composite Interval Mapping (CIM) | qMws1.03, qMws1.04, qMws6.02, qMws8.05, qMws10.03, qMws10.06 [78] |
| 143 inbred lines | Mixed Linear Models | SYNGENTA14387, PHM13619.5, PZE-101040783, SYN11249, PZE-101151153, SYN10891, SYN10137, PZA00131.14, SYN37674, PZE-102031753, PZE-103084298, ZM012464-0529, SYN30108, PZE-108057528, SYN4935 [71] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, E.; Fan, X.; Jiang, F.; Zhang, Y. Advancements in Research on Prevention and Control Strategies for Maize White Spot Disease. Genes 2023, 14, 2061. https://doi.org/10.3390/genes14112061
Xing E, Fan X, Jiang F, Zhang Y. Advancements in Research on Prevention and Control Strategies for Maize White Spot Disease. Genes. 2023; 14(11):2061. https://doi.org/10.3390/genes14112061
Chicago/Turabian StyleXing, Enyun, Xingming Fan, Fuyan Jiang, and Yudong Zhang. 2023. "Advancements in Research on Prevention and Control Strategies for Maize White Spot Disease" Genes 14, no. 11: 2061. https://doi.org/10.3390/genes14112061