
Characterization of Glycosyltransferase Family 1 (GT1) and Their Potential Roles in Anthocyanin Biosynthesis in Maize

Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Maize GT1s
2.2. Multiple Sequence Alignment of GT1s
2.3. Phylogenetic Analysis of GT1s
2.4. Gene Structure and Conserved Motif Analysis
2.5. Chromosomal Localization and Collinearity Analysis for Duplicated Genes
2.6. Expression Profile Analysis
2.7. Molecular Docking of GT1s
3. Results
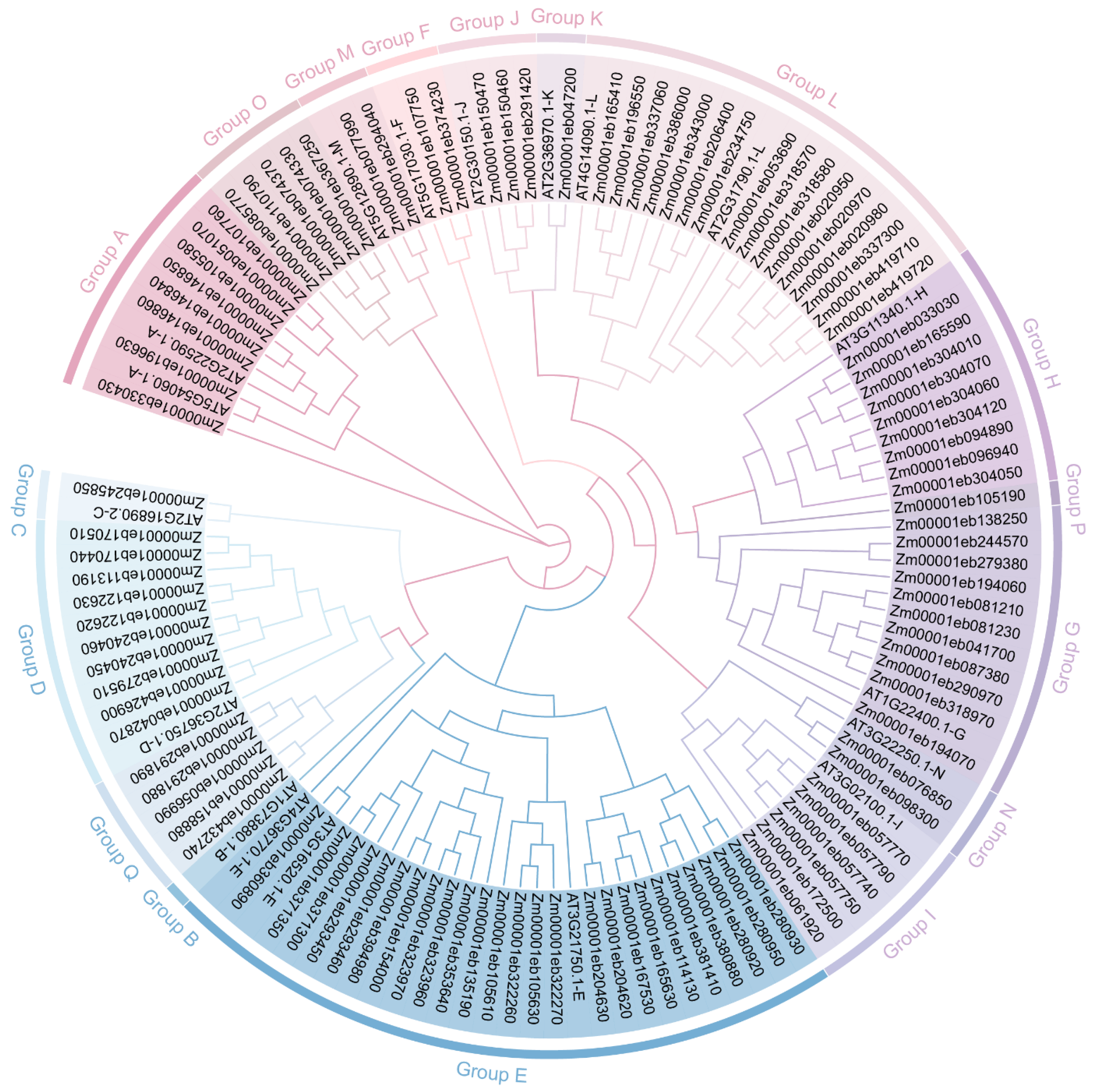
3.1. Identification and Phylogenetic Analysis of Maize GT1s
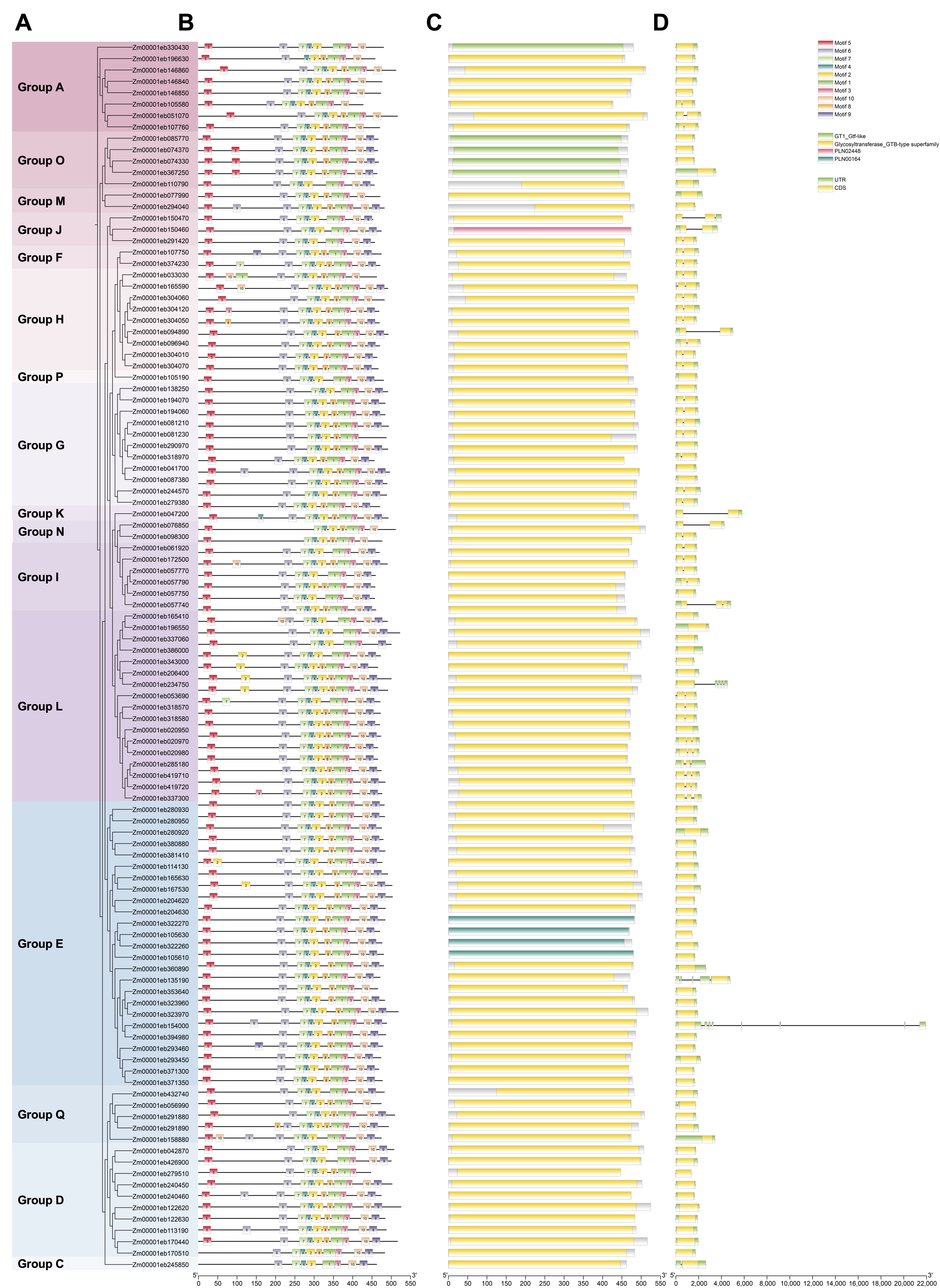
3.2. Motif Composition, Conserved Domain, and Gene Structure Analyses of GT1 Genes
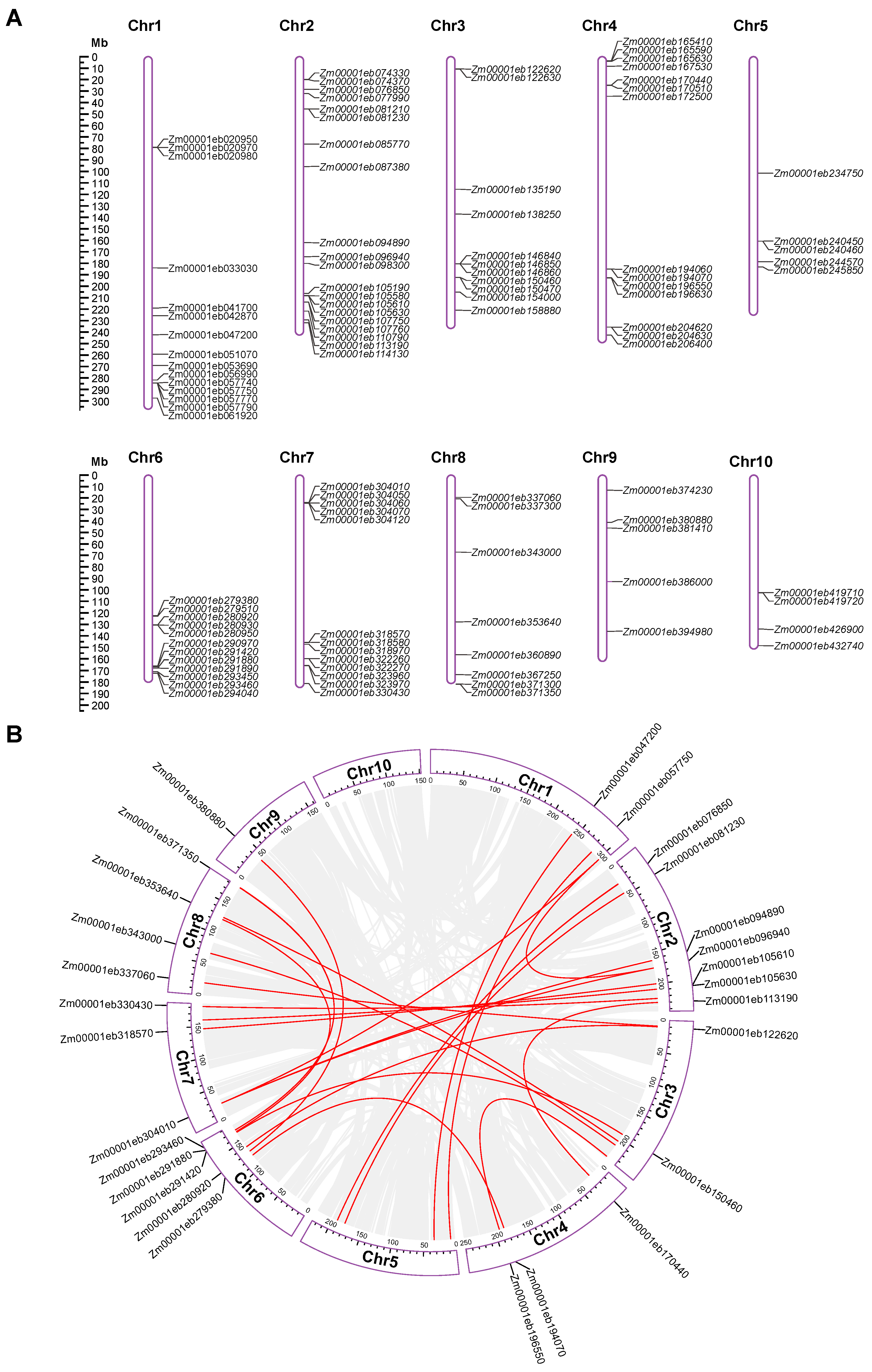
3.3. Chromosomal Distribution and Gene Duplication of GT1 Genes
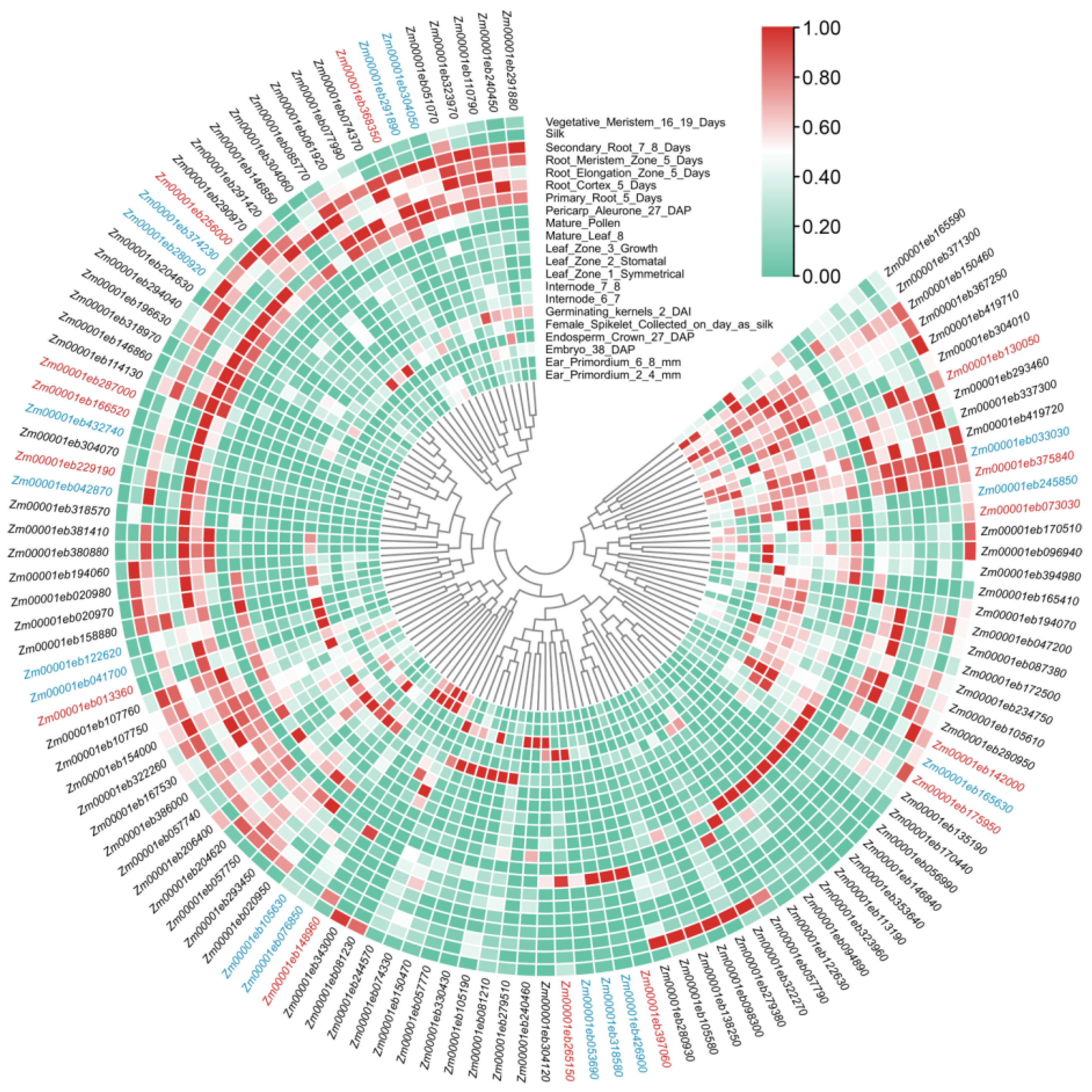
3.4. Expression Analysis of GT1 Genes
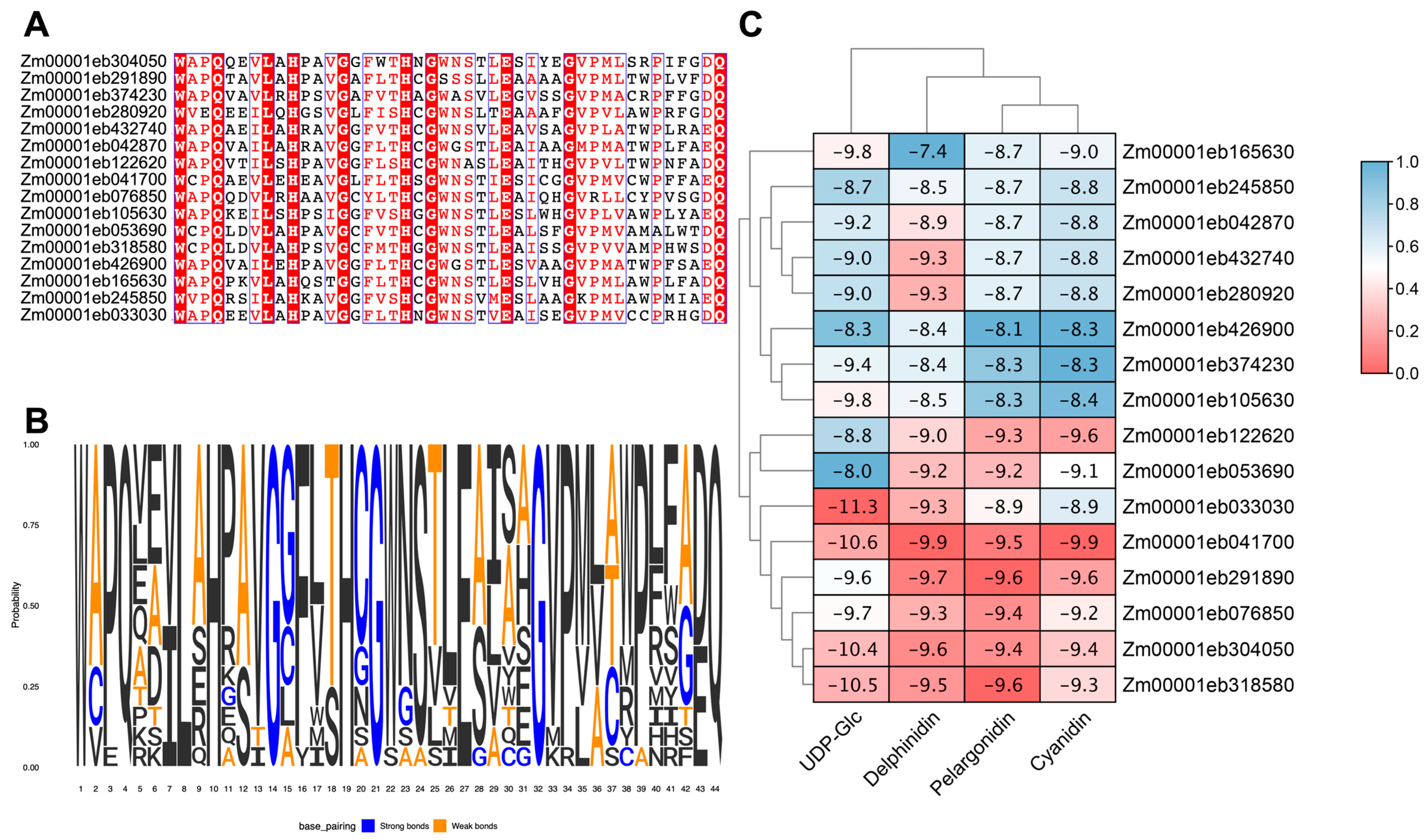
3.5. Candidate GT1 Genes in Anthocyanin Biosynthesis
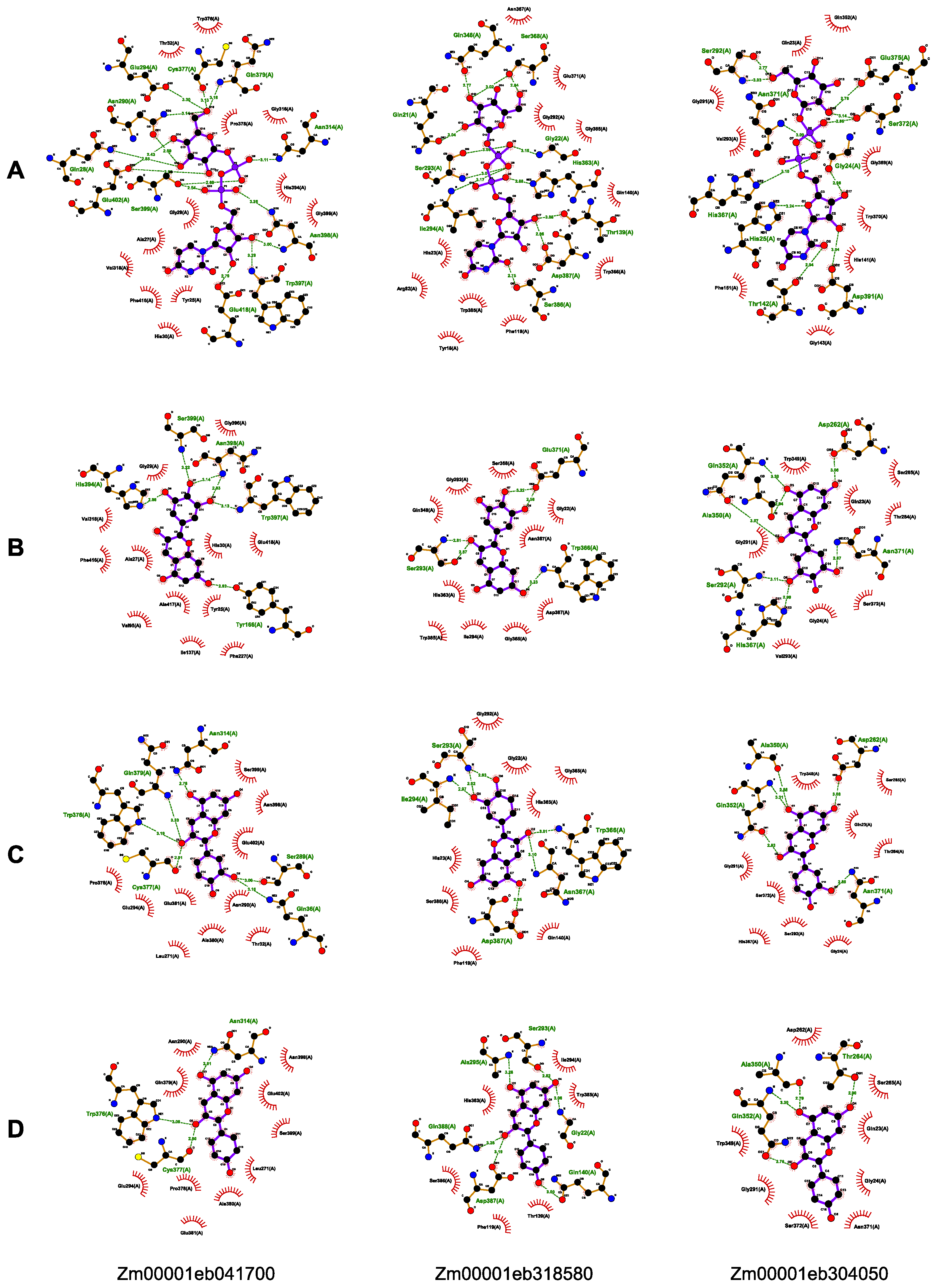
3.6. Molecular Docking of GT1s
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Strasser, R. Plant protein glycosylation. Glycobiology 2016, 26, 926–939. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.Z.; Jin, S.H.; Jiang, X.Y.; Dong, R.R.; Li, P.; Li, Y.J.; Hou, B.K. Ectopic expression of UGT75D1, a glycosyltransferase preferring indole-3-butyric acid, modulates cotyledon development and stress tolerance in seed germination of Arabidopsis thaliana. Plant Mol. Biol. 2016, 90, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hou, B. Glycosyltransferases: Key players involved in the modification of plant secondary metabolites. Front. Biol. China 2009, 4, 39–46. [Google Scholar] [CrossRef]
- Jones, P.; Messner, B.; Nakajima, J.; Schäffner, A.R.; Saito, K. UGT73C6 and UGT78D1, glycosyltransferases involved in flavonol glycoside biosynthesis in Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 43910–43918. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Han, K.; Heller, W.; Albert, A.; Dobrev, P.I.; Zažímalová, E.; Schäffner, A.R. Kaempferol 3-O-rhamnoside-7-O-rhamnoside is an endogenous flavonol inhibitor of polar auxin transport in Arabidopsis shoots. New Phytol. 2014, 201, 466–475. [Google Scholar] [CrossRef]
- Hofer, B. Recent developments in the enzymatic O-glycosylation of flavonoids. Appl. Microbiol. Biotechnol. 2016, 100, 4269–4281. [Google Scholar] [CrossRef]
- Liu, Z.; Yan, J.P.; Li, D.K.; Luo, Q.; Yan, Q.; Liu, Z.B.; Ye, L.M.; Wang, J.M.; Li, X.F.; Yang, Y. UDP-glucosyltransferase71c5, a major glucosyltransferase, mediates abscisic acid homeostasis in Arabidopsis. Plant Physiol. 2015, 167, 1659–1670. [Google Scholar] [CrossRef]
- Poppenberger, B.; Fujioka, S.; Soeno, K.; George, G.L.; Vaistij, F.E.; Hiranuma, S.; Seto, H.; Takatsuto, S.; Adam, G.; Yoshida, S.; et al. The UGT73C5 of Arabidopsis thaliana glucosylates brassinosteroids. Proc. Natl. Acad. Sci. USA 2005, 102, 15253–15258. [Google Scholar] [CrossRef]
- Tanaka, K.; Hayashi, K.; Natsume, M.; Kamiya, Y.; Sakakibara, H.; Kawaide, H.; Kasahara, H. UGT74D1 catalyzes the glucosylation of 2-oxindole-3-acetic acid in the auxin metabolic pathway in Arabidopsis. Plant Cell Physiol. 2014, 55, 218–228. [Google Scholar] [CrossRef]
- Wang, J.; Ma, X.M.; Kojima, M.; Sakakibara, H.; Hou, B.K. Glucosyltransferase UGT76C1 finely modulates cytokinin responses via cytokinin N-glucosylation in Arabidopsis thaliana. Plant Physiol. Biochem. 2013, 65, 9–16. [Google Scholar] [CrossRef]
- George Thompson, A.M.; Iancu, C.V.; Neet, K.E.; Dean, J.V.; Choe, J.Y. Differences in salicylic acid glucose conjugations by UGT74F1 and UGT74F2 from Arabidopsis thaliana. Sci. Rep. 2017, 7, 46629. [Google Scholar] [CrossRef]
- Tognetti, V.B.; Van Aken, O.; Morreel, K.; Vandenbroucke, K.; van de Cotte, B.; De Clercq, I.; Chiwocha, S.; Fenske, R.; Prinsen, E.; Boerjan, W.; et al. Perturbation of indole-3-butyric acid homeostasis by the UDP-glucosyltransferase UGT74E2 modulates Arabidopsis architecture and water stress tolerance. Plant Cell 2010, 22, 2660–2679. [Google Scholar] [CrossRef]
- Von Saint Paul, V.; Zhang, W.; Kanawati, B.; Geist, B.; Faus-Kessler, T.; Schmitt-Kopplin, P.; Schäffner, A.R. The Arabidopsis glucosyltransferase UGT76B1 conjugates isoleucic acid and modulates plant defense and senescence. Plant Cell 2011, 23, 4124–4145. [Google Scholar] [CrossRef]
- Gharabli, H.; Della Gala, V.; Welner, D.H. The function of UDP-glycosyltransferases in plants and their possible use in crop protection. Biotechnol. Adv. 2023, 67, 108182. [Google Scholar] [CrossRef]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S.G. Glycosyltransferases: Structures, functions, and mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Hanada, K. An evolutionary view of functional diversity in family 1 glycosyltransferases. Plant J. 2011, 66, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Bowles, D.; Lim, E.K.; Poppenberger, B.; Vaistij, F.E. Glycosyltransferases of lipophilic small molecules. Annu. Rev. Plant Biol. 2006, 57, 567–597. [Google Scholar] [CrossRef]
- Liang, D.M.; Liu, J.H.; Wu, H.; Wang, B.B.; Zhu, H.J.; Qiao, J.J. Glycosyltransferases: Mechanisms and applications in natural product development. Chem. Soc. Rev. 2015, 44, 8350–8374. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, Z.; Zhang, L.; Wang, J.; Wu, C. Glycosyltransferase GT1 family: Phylogenetic distribution, substrates coverage, and representative structural features. Comput. Struct. Biotechnol. J. 2020, 18, 1383–1390. [Google Scholar] [CrossRef]
- Caputi, L.; Malnoy, M.; Goremykin, V.; Nikiforova, S.; Martens, S. A genome-wide phylogenetic reconstruction of family 1 UDP-glycosyltransferases revealed the expansion of the family during the adaptation of plants to life on land. Plant J. 2012, 69, 1030–1042. [Google Scholar] [CrossRef]
- Sun, G.; Liao, J.; Kurze, E.; Hoffmann, T.D.; Steinchen, W.; McGraphery, K.; Habegger, R.; Marek, L.; Catici, D.A.M.; Ludwig, C.; et al. Apocarotenoids are allosteric effectors of a dimeric plant glycosyltransferase involved in defense and lignin formation. New Phytol. 2023, 238, 2080–2098. [Google Scholar] [CrossRef]
- Wang, Y.W.; Wang, W.C.; Jin, S.H.; Wang, J.; Wang, B.; Hou, B.K. Over-expression of a putative poplar glycosyltransferase gene, PtGT1, in tobacco increases lignin content and causes early flowering. J. Exp. Bot. 2012, 63, 2799–2808. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xue, X.; Wang, X.; Zhang, C.; Zheng, D.; Song, W.; Zhao, J.; Wei, J.; Wu, Z.; Zhang, Z. Natural variation of ZmCGT1 is responsible for isoorientin accumulation in maize silk. Plant J. 2022, 109, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T.; Jones, P. Glycosyltransferases in plant natural product synthesis: Characterization of a supergene family. Trends Plant Sci. 2000, 5, 380–386. [Google Scholar] [CrossRef]
- Li, Y.; Li, P.; Wang, Y.; Dong, R.; Yu, H.; Hou, B. Genome-wide identification and phylogenetic analysis of Family-1 UDP glycosyltransferases in maize (Zea mays). Planta 2014, 239, 1265–1279. [Google Scholar] [CrossRef]
- Lu, L.; Hou, Q.; Wang, L.; Zhang, T.; Zhao, W.; Yan, T.; Zhao, L.; Li, J.; Wan, X. Genome-Wide Identification and Characterization of Polygalacturonase Gene Family in Maize (Zea mays L.). Int. J. Mol. Sci. 2021, 22, 722. [Google Scholar] [CrossRef]
- Zhu, T.; Wu, S.; Zhang, D.; Li, Z.; Xie, K.; An, X.; Ma, B.; Hou, Q.; Dong, Z.; Tian, Y.; et al. Genome-wide analysis of maize GPAT gene family and cytological characterization and breeding application of ZmMs33/ZmGPAT6 gene. Theor. Appl. Genet. 2019, 132, 2137–2154. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Ma, B.; Hou, Q.; Wan, X. Phylogeny and Functions of LOB Domain Proteins in Plants. Int. J. Mol. Sci. 2020, 21, 2278. [Google Scholar] [CrossRef]
- Hou, Q.; Wang, L.; Qi, Y.; Yan, T.; Zhang, F.; Zhao, W.; Wan, X. A systematic analysis of the subtilase gene family and expression and subcellular localization investigation of anther-specific members in maize. Plant Physiol. Biochem. 2023, 203, 108041. [Google Scholar] [CrossRef]
- Fang, C.; Wu, S.; Li, Z.; Pan, S.; Wu, Y.; An, X.; Long, Y.; Wei, X.; Wan, X. A Systematic Investigation of Lipid Transfer Proteins Involved in Male Fertility and Other Biological Processes in Maize. Int. J. Mol. Sci. 2023, 24, 1660. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef]
- Hufford, M.B.; Seetharam, A.S.; Woodhouse, M.R.; Chougule, K.M.; Ou, S.; Liu, J.; Ricci, W.A.; Guo, T.; Olson, A.; Qiu, Y.; et al. De novo assembly, annotation, and comparative analysis of 26 diverse maize genomes. Science 2021, 373, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res 2014, 42, W320–W324. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Li, Y.; Baldauf, S.; Lim, E.K.; Bowles, D.J. Phylogenetic analysis of the UDP-glycosyltransferase multigene family of Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 4338–4343. [Google Scholar] [CrossRef]
- Walley, J.W.; Sartor, R.C.; Shen, Z.; Schmitz, R.J.; Wu, K.J.; Urich, M.A.; Nery, J.R.; Smith, L.G.; Schnable, J.C.; Ecker, J.R.; et al. Integration of omic networks in a developmental atlas of maize. Science 2016, 353, 814–818. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Hirai, M.Y.; Yonekura-Sakakibara, K. Decoding genes with coexpression networks and metabolomics-‘majority report by precogs’. Trends Plant Sci. 2008, 13, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, D.; Peng, Y.; Cai, M.; Liang, Z.; Yuan, Z.; Du, X.; Wang, J.; Schnable, P.S.; Gu, R.; et al. The anthocyanin accumulation related ZmBZ1, facilitates seedling salinity stress tolerance via ROS scavenging. Int. J. Mol. Sci. 2022, 23, 16123. [Google Scholar] [CrossRef]
- Magadum, S.; Banerjee, U.; Murugan, P.; Gangapur, D.; Ravikesavan, R. Gene duplication as a major force in evolution. J. Genet. 2013, 92, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Chr | Chromosomal Position | Length (aa) | Gene Symbol | Classification | Number of Introns | Gene Type 1 |
|---|---|---|---|---|---|---|---|
| Zm00001eb020950 | Chr1 | 79115933–79117889 | 473 | L | 0 | Proximal | |
| Zm00001eb020970 | Chr1 | 79160471–79162703 | 465 | L | 2 | Tandem | |
| Zm00001eb020980 | Chr1 | 79196751–79198788 | 465 | L | 2 | Tandem | |
| Zm00001eb033030 | Chr1 | 184391611–184393452 | 462 | BX9 | H | 1 | Dispersed |
| Zm00001eb041700 | Chr1 | 219547437–219549217 | 496 | G | 0 | Dispersed | |
| Zm00001eb042870 | Chr1 | 226122699–226124455 | 507 | D | 0 | Dispersed | |
| Zm00001eb047200 | Chr1 | 242648507–242654443 | 492 | K | 1 | WGD or Segmental | |
| Zm00001eb051070 | Chr1 | 259782541–259784813 | 516 | A | 1 | Dispersed | |
| Zm00001eb053690 | Chr1 | 269552894–269554690 | 471 | IAGLU1 | L | 2 | Dispersed |
| Zm00001eb056990 | Chr1 | 282218667–282220404 | 474 | Q | 1 | Dispersed | |
| Zm00001eb057740 | Chr1 | 284753755–284758573 | 460 | I | 2 | Tandem | |
| Zm00001eb057750 | Chr1 | 284764856–284766611 | 457 | I | 0 | WGD or Segmental | |
| Zm00001eb057770 | Chr1 | 284774974–284776823 | 459 | I | 1 | Tandem | |
| Zm00001eb057790 | Chr1 | 284779663–284781743 | 458 | I | 1 | Tandem | |
| Zm00001eb061920 | Chr1 | 298006146–298007982 | 469 | I | 1 | Dispersed | |
| Zm00001eb074330 | Chr2 | 19866206–19867838 | 467 | CZOG1 | O | 0 | Proximal |
| Zm00001eb074370 | Chr2 | 20057564–20059103 | 465 | O | 0 | Proximal | |
| Zm00001eb076850 | Chr2 | 28481546–28485790 | 511 | SK1 | N | 1 | WGD or Segmental |
| Zm00001eb077990 | Chr2 | 32158225–32160541 | 471 | M | 0 | Dispersed | |
| Zm00001eb081210 | Chr2 | 45506320–45508428 | 493 | G | 1 | Proximal | |
| Zm00001eb081230 | Chr2 | 45701694–45703531 | 487 | G | 1 | WGD or Segmental | |
| Zm00001eb085770 | Chr2 | 76160840–76162497 | 466 | O | 0 | Dispersed | |
| Zm00001eb087380 | Chr2 | 95946783–95948674 | 489 | G | 0 | Dispersed | |
| Zm00001eb094890 | Chr2 | 162271068–162276069 | 492 | UGT5174 | H | 1 | WGD or Segmental |
| Zm00001eb096940 | Chr2 | 174552351–174554493 | 470 | H | 1 | WGD or Segmental | |
| Zm00001eb098300 | Chr2 | 180463741–180465530 | 476 | N | 1 | Dispersed | |
| Zm00001eb105190 | Chr2 | 206674564–206676436 | 480 | P | 0 | Dispersed | |
| Zm00001eb105580 | Chr2 | 208529035–208530699 | 427 | SM2 | A | 1 | Dispersed |
| Zm00001eb105610 | Chr2 | 208610550–208612222 | 480 | E | 0 | WGD or Segmental | |
| Zm00001eb105630 | Chr2 | 208612412–208613837 | 470 | E | 0 | WGD or Segmental | |
| Zm00001eb107750 | Chr2 | 214236761–214238763 | 474 | F | 1 | Dispersed | |
| Zm00001eb107760 | Chr2 | 214252312–214254275 | 470 | A | 1 | Dispersed | |
| Zm00001eb110790 | Chr2 | 222058692–222060708 | 456 | M | 0 | Dispersed | |
| Zm00001eb113190 | Chr2 | 229706824–229708687 | 487 | D | 0 | WGD or Segmental | |
| Zm00001eb114130 | Chr2 | 232266306–232268281 | 476 | E | 0 | Dispersed | |
| Zm00001eb122620 | Chr3 | 10681696–10683755 | 525 | D | 0 | WGD or Segmental | |
| Zm00001eb122630 | Chr3 | 10830992–10832903 | 484 | D | 0 | Tandem | |
| Zm00001eb135190 | Chr3 | 115806978–115812646 | 471 | E | 4 | Dispersed | |
| Zm00001eb138250 | Chr3 | 137388625–137390462 | 491 | G | 0 | Dispersed | |
| Zm00001eb146840 | Chr3 | 180787206–180789032 | 476 | A | 0 | Tandem | |
| Zm00001eb146850 | Chr3 | 180819236–180820720 | 473 | A | 0 | Tandem | |
| Zm00001eb146860 | Chr3 | 180824047–180826007 | 512 | A | 0 | Tandem | |
| Zm00001eb150460 | Chr3 | 192562189–192565830 | 474 | J | 1 | WGD or Segmental | |
| Zm00001eb150470 | Chr3 | 192567391–192571400 | 452 | J | 2 | Tandem | |
| Zm00001eb154000 | Chr3 | 205428339–205450268 | 488 | E | 9 | Tandem | |
| Zm00001eb158880 | Chr3 | 221304007–221307439 | 474 | Q | 0 | Dispersed | |
| Zm00001eb165410 | Chr4 | 3542471–3544431 | 490 | L | 0 | Dispersed | |
| Zm00001eb165590 | Chr4 | 4060833–4062879 | 491 | BX8 | H | 2 | Dispersed |
| Zm00001eb165630 | Chr4 | 4116762–4118560 | 491 | E | 0 | Dispersed | |
| Zm00001eb167530 | Chr4 | 8333244–8335422 | 502 | E | 1 | Dispersed | |
| Zm00001eb170440 | Chr4 | 24818567–24820532 | 516 | D | 0 | WGD or Segmental | |
| Zm00001eb170510 | Chr4 | 25081389–25083426 | 483 | D | 0 | Proximal | |
| Zm00001eb172500 | Chr4 | 34588042–34589834 | 490 | I | 1 | Dispersed | |
| Zm00001eb194060 | Chr4 | 185323500–185325440 | 484 | G | 1 | Proximal | |
| Zm00001eb194070 | Chr4 | 185374820–185376744 | 484 | G | 1 | WGD or Segmental | |
| Zm00001eb196550 | Chr4 | 192838263–192841161 | 522 | L | 0 | WGD or Segmental | |
| Zm00001eb196630 | Chr4 | 193168606–193170298 | 458 | UFGT4 | A | 0 | Dispersed |
| Zm00001eb204620 | Chr4 | 236014681–236017119 | 503 | CEP2 | E | 0 | Tandem |
| Zm00001eb204630 | Chr4 | 236082248–236084078 | 485 | E | 0 | Tandem | |
| Zm00001eb206400 | Chr4 | 243100226–243107731 | 500 | L | 0 | WGD or Segmental | |
| Zm00001eb234750 | Chr5 | 101616810–101621419 | 491 | L | 4 | WGD or Segmental | |
| Zm00001eb240450 | Chr5 | 161029842–161031557 | 502 | D | 0 | Tandem | |
| Zm00001eb240460 | Chr5 | 161031522–161033144 | 474 | D | 0 | Tandem | |
| Zm00001eb244570 | Chr5 | 178888288–178890479 | 488 | G | 1 | Dispersed | |
| Zm00001eb245850 | Chr5 | 183441650–183444276 | 462 | C | 1 | Tandem | |
| Zm00001eb279380 | Chr6 | 122762448–122764350 | 470 | G | 1 | WGD or Segmental | |
| Zm00001eb279510 | Chr6 | 123180906–123182259 | 447 | D | 0 | Dispersed | |
| Zm00001eb280920 | Chr6 | 130691016–130693844 | 475 | CGT1 | E | 0 | WGD or Segmental |
| Zm00001eb280930 | Chr6 | 130704890–130706769 | 482 | UGT1 | E | 0 | Tandem |
| Zm00001eb280950 | Chr6 | 130831473–130833292 | 483 | CGT2 | E | 0 | Proximal |
| Zm00001eb290970 | Chr6 | 166328839–166330747 | 491 | G | 0 | Dispersed | |
| Zm00001eb291420 | Chr6 | 167093344–167095148 | 457 | J | 1 | WGD or Segmental | |
| Zm00001eb291880 | Chr6 | 168153151–168154910 | 509 | Q | 0 | WGD or Segmental | |
| Zm00001eb291890 | Chr6 | 168199823–168201808 | 493 | Q | 0 | Tandem | |
| Zm00001eb293450 | Chr6 | 171371429–171373586 | 473 | E | 0 | Tandem | |
| Zm00001eb293460 | Chr6 | 171373527–171375229 | 478 | E | 0 | WGD or Segmental | |
| Zm00001eb294040 | Chr6 | 172699717–172701409 | 482 | M | 0 | Dispersed | |
| Zm00001eb304010 | Chr7 | 23801316–23803037 | 463 | UGT9250 | H | 1 | WGD or Segmental |
| Zm00001eb304050 | Chr7 | 24029052–24030857 | 469 | UFGT2 | H | 1 | Tandem |
| Zm00001eb304060 | Chr7 | 24154958–24156803 | 482 | H | 1 | Proximal | |
| Zm00001eb304070 | Chr7 | 24181111–24183054 | 466 | H | 1 | Proximal | |
| Zm00001eb304120 | Chr7 | 24588945–24591016 | 468 | CNGT1 | H | 1 | Proximal |
| Zm00001eb318570 | Chr7 | 146013215–146015100 | 472 | L | 1 | WGD or Segmental | |
| Zm00001eb318580 | Chr7 | 146096773–146098587 | 470 | L | 1 | Tandem | |
| Zm00001eb318970 | Chr7 | 147684571–147686411 | 456 | G | 1 | Dispersed | |
| Zm00001eb322260 | Chr7 | 160217163–160219108 | 476 | E | 0 | Tandem | |
| Zm00001eb322270 | Chr7 | 160220531–160222324 | 484 | E | 0 | Tandem | |
| Zm00001eb323960 | Chr7 | 165932228–165934061 | 483 | E | 0 | Tandem | |
| Zm00001eb323970 | Chr7 | 166034566–166036489 | 518 | UMC2716 | E | 0 | Tandem |
| Zm00001eb330430 | Chr7 | 181720552–181722435 | 480 | A | 0 | WGD or Segmental | |
| Zm00001eb337060 | Chr8 | 19469860–19471775 | 500 | L | 0 | WGD or Segmental | |
| Zm00001eb337300 | Chr8 | 20579174–20581428 | 476 | L | 2 | Dispersed | |
| Zm00001eb343000 | Chr8 | 67212322–67213925 | 465 | L | 0 | WGD or Segmental | |
| Zm00001eb353640 | Chr8 | 128038314–128040087 | 465 | E | 0 | WGD or Segmental | |
| Zm00001eb360890 | Chr8 | 156736301–156738932 | 480 | E | 0 | Dispersed | |
| Zm00001eb367250 | Chr8 | 173936706–173940214 | 463 | CZOG2 | O | 0 | Dispersed |
| Zm00001eb371300 | Chr8 | 182117851–182119448 | 468 | E | 0 | Tandem | |
| Zm00001eb371350 | Chr8 | 182219335–182220996 | 477 | E | 0 | WGD or Segmental | |
| Zm00001eb374230 | Chr9 | 13120306–13122164 | 471 | BZ1 | F | 1 | Dispersed |
| Zm00001eb380880 | Chr9 | 41151938–41153717 | 479 | E | 0 | WGD or Segmental | |
| Zm00001eb381410 | Chr9 | 46177110–46178902 | 484 | E | 0 | Dispersed | |
| Zm00001eb386000 | Chr9 | 92897492–92899854 | 472 | UMC2700 | L | 0 | Tandem |
| Zm00001eb394980 | Chr9 | 136330935–136332755 | 486 | E | 0 | Dispersed | |
| Zm00001eb419710 | Chr10 | 102460083–102462158 | 474 | L | 2 | Tandem | |
| Zm00001eb419720 | Chr10 | 102642521–102644377 | 484 | L | 2 | Tandem | |
| Zm00001eb426900 | Chr10 | 134433367–134435275 | 500 | D | 0 | Dispersed | |
| Zm00001eb432740 | Chr10 | 148886364–148888273 | 482 | Q | 0 | Dispersed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Li, Y.; Wang, X.; Jiao, Z.; Zhang, W.; Long, Y. Characterization of Glycosyltransferase Family 1 (GT1) and Their Potential Roles in Anthocyanin Biosynthesis in Maize. Genes 2023, 14, 2099. https://doi.org/10.3390/genes14112099
Li H, Li Y, Wang X, Jiao Z, Zhang W, Long Y. Characterization of Glycosyltransferase Family 1 (GT1) and Their Potential Roles in Anthocyanin Biosynthesis in Maize. Genes. 2023; 14(11):2099. https://doi.org/10.3390/genes14112099
Chicago/Turabian StyleLi, Huangai, Yiping Li, Xiaofang Wang, Ziwei Jiao, Wei Zhang, and Yan Long. 2023. "Characterization of Glycosyltransferase Family 1 (GT1) and Their Potential Roles in Anthocyanin Biosynthesis in Maize" Genes 14, no. 11: 2099. https://doi.org/10.3390/genes14112099