
Identification of Differentially Expressed Genes and Prediction of Expression Regulation Networks in Dysfunctional Endothelium

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Extraction Based on GEO Database
2.2. Differential Gene Screening and Data Processing
2.3. Construction of the miRNA–Target Gene and Target Gene Transcription Factor (TF) Regulatory Network
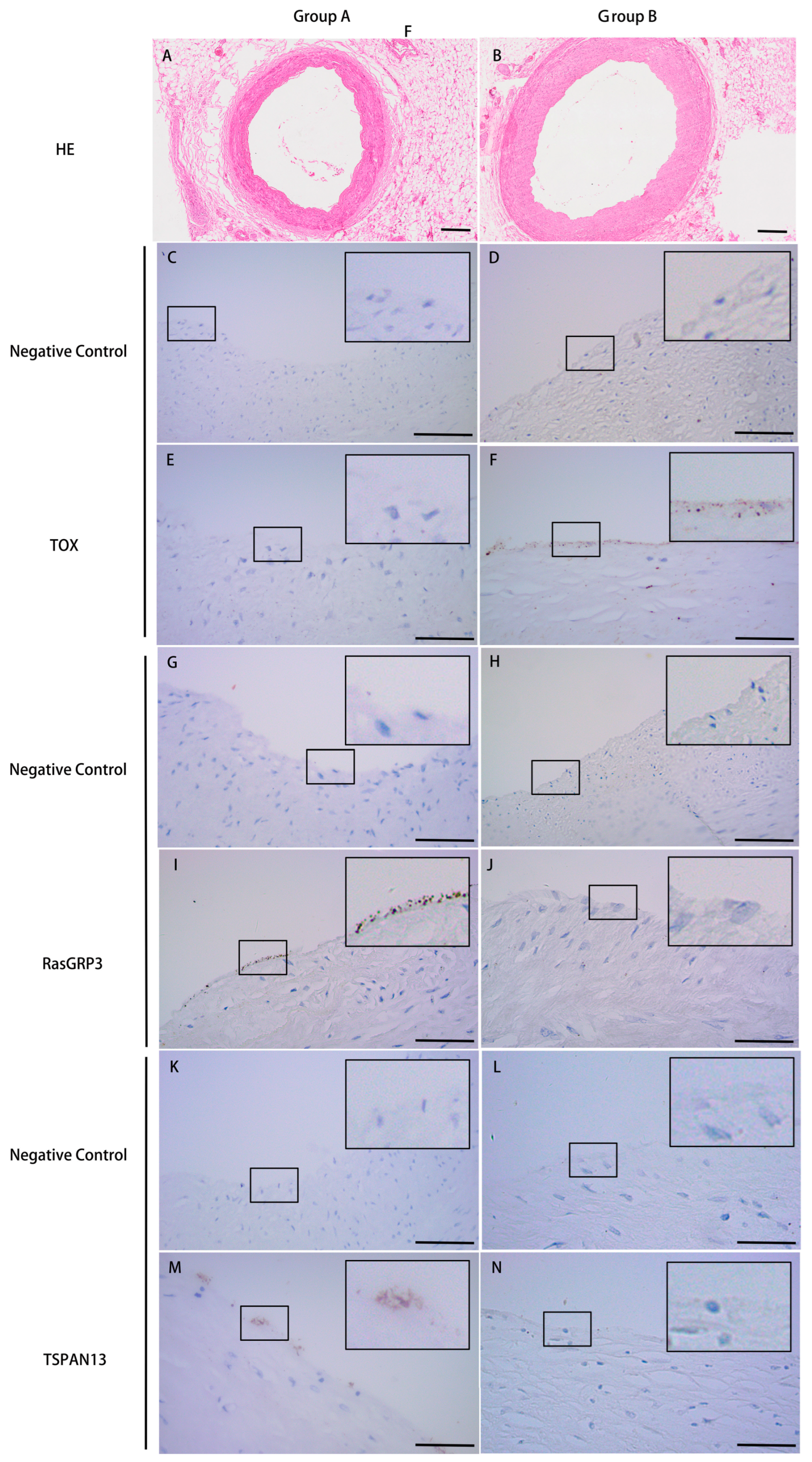
2.4. Immunohistochemistry Validation
2.5. Statistical analysis
3. Results
3.1. Identification and Correlation Analysis of Significant DEGs in ECA
3.2. Construction of the miRNA–Target Gene and Target Gene–TF Regulatory Network
3.3. Validation of the Microarray Data by IHC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Han, C.; Kang, K.-W.; Kim, T.Y.; Uhm, J.-S.; Park, J.-W.; Jung, I.H.; Kim, M.; Bae, S.; Lim, H.-S.; Yoon, D. Artificial Intelligence-Enabled ECG Algorithm for the Prediction of Coronary Artery Calcification. Front. Cardiovasc. Med. 2022, 9, 849223. [Google Scholar] [CrossRef] [PubMed]
- Ference, B.A.; Mahajan, N. The Role of Early LDL Lowering to Prevent the Onset of Atherosclerotic Disease. Curr. Atheroscler. Rep. 2013, 15, 312. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Narang, R.; Sreenivas, V.; Rastogi, V.; Bhatia, J.; Saluja, D.; Srivastava, K. Circulatory miR-133b and miR-21 as Novel Biomarkers in Early Prediction and Diagnosis of Coronary Artery Disease. Genes 2020, 11, 164. [Google Scholar] [CrossRef] [PubMed]
- Zorc-Pleskovič, R.; Pleskovič, A.; Vraspir-Porenta, O.; Zorc, M.; Milutinović, A. Immune cells and vasa vasorum in the tunica media of atherosclerotic coronary arteries. Bosn. J. Basic Med Sci. 2018, 18, 240–245. [Google Scholar] [CrossRef]
- Guarner-Lans, V.; Ramírez-Higuera, A.; Rubio-Ruiz, M.; Castrejón-Téllez, V.; Soto, M.; Pérez-Torres, I. Early Programming of Adult Systemic Essential Hypertension. Int. J. Mol. Sci. 2020, 21, 1203. [Google Scholar] [CrossRef]
- Ditano-Vázquez, P.; Torres-Peña, J.D.; Galeano-Valle, F.; Pérez-Caballero, A.I.; Demelo-Rodríguez, P.; Lopez-Miranda, J.; Katsiki, N.; Delgado-Lista, J.; Alvarez-Sala-Walther, L.A. The Fluid Aspect of the Mediterranean Diet in the Prevention and Management of Cardiovascular Disease and Diabetes: The Role of Polyphenol Content in Moderate Consumption of Wine and Olive Oil. Nutrients 2019, 11, 2833. [Google Scholar] [CrossRef]
- Yagi, H.; Sumino, H.; Aoki, T.; Tsunekawa, K.; Araki, O.; Kimura, T.; Makoto, N.; Takayuki, O.; Masami, M. Impaired blood rheology is associated with endothelial dysfunction in patients with coronary risk factors. Clin. Hemorheol. Microcirc. 2016, 62, 139–150. [Google Scholar] [CrossRef]
- Stary, H. Natural history and histological classification of atherosclerotic lesions: An update. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1177–1178. [Google Scholar] [CrossRef]
- Reddy, K.G.; Nair, R.N.; Sheehan, H.M.; Hodgson, J.M. Evidence that selective endothelial dysfunction may occur in the absence of angiographic or ultrasound atherosclerosis in patients with risk factors for atherosclerosis. J. Am. Coll. Cardiol. 1994, 23, 833–843. [Google Scholar] [CrossRef]
- Bonetti, P.O.; Lerman, L.O.; Lerman, A. Endothelial dysfunction: A marker of atherosclerotic risk. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 168–175. [Google Scholar] [CrossRef]
- van Rooij, E.; Olson, E.N. MicroRNAs: Powerful new regulators of heart disease and provocative therapeutic targets. J. Clin. Investig. 2007, 117, 2369–2376. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Schulte, C.; Zeller, T. microRNA-based diagnostics and therapy in cardiovascular disease-Summing up the facts. Cardiovasc. Diagn. Ther. 2015, 5, 17–36. [Google Scholar]
- Lambert, M.; Jambon, S.; Depauw, S.; David-Cordonnier, M.-H. Targeting Transcription Factors for Cancer Treatment. Molecules 2018, 23, 1479. [Google Scholar] [CrossRef]
- Zhou, X.-G.; Huang, X.-L.; Liang, S.-Y.; Tang, S.-M.; Wu, S.-K.; Huang, T.-T.; Mo, Z.-N.; Wang, Q.-Y. Identifying miRNA and gene modules of colon cancer associated with pathological stage by weighted gene co-expression network analysis. Onco Targets Ther. 2018, 11, 2815–2830. [Google Scholar] [CrossRef]
- Hebbel, R.P.; Wei, P.; Milbauer, L.; Corban, M.T.; Solovey, A.; Kiley, J.; Pattee, J.; Lerman, L.O.; Pan, W.; Lerman, A. Abnormal Endothelial Gene Expression Associated With Early Coronary Atherosclerosis. J. Am. Heart Assoc. 2020, 9, e016134. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, H.; Yao, C.; Shuai, J.; Sun, X. Effect of Dynamic Interaction between microRNA and Transcription Factor on Gene Expression. BioMed Res. Int. 2016, 2016, 1–10. [Google Scholar] [CrossRef]
- Janky, R.; Verfaillie, A.; Imrichová, H.; Van de Sande, B.; Standaert, L.; Christiaens, V.; Hulselmans, G.; Herten, K.; Sanchez, M.N.; Potier, D.; et al. iRegulon: From a Gene List to a Gene Regulatory Network Using Large Motif and Track Collections. PLoS Comput. Biol. 2014, 10, e1003731. [Google Scholar] [CrossRef]
- Mayer, F.; Pröpper, S.; Ritz-Timme, S. Dityrosine, a protein product of oxidative stress, as a possible marker of acute myo-cardial infarctions. Int. J. Legal Med. 2014, 128, 787–794. [Google Scholar] [CrossRef]
- Aliahmad, P.; Seksenyan, A.; Kaye, J. The many roles of TOX in the immune system. Curr. Opin. Immunol. 2012, 24, 173–177. [Google Scholar] [CrossRef]
- Wilkinson, B.; Chen, J.Y.-F.; Han, P.; Rufner, K.M.; Goularte, O.D.; Kaye, J. TOX: An HMG box protein implicated in the regulation of thymocyte selection. Nat. Immunol. 2002, 3, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Arora, M.; Kumari, S.; Singh, J.; Chopra, A.; Chauhan, S.S. Expression pattern, regulation, and clinical significance of TOX in breast cancer. Cancer Immunol. Immunother. 2021, 70, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Aliahmad, P.; O’Flaherty, E.; Han, P.; Goularte, O.D.; Wilkinson, B.; Satake, M.; Molkentin, J.; Kaye, J. TOX Provides a Link Between Calcineurin Activation and CD8 Lineage Commitment. J. Exp. Med. 2004, 199, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Chi, T.H.; Wan, M.; Zhao, K.; Taniuchi, I.; Chen, L.; Littman, D.R.; Crabtree, G.R. Reciprocal regulation of CD4/CD8 expression by SWI/SNF-like BAF complexes. Nature 2002, 418, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Aliahmad, P.; De La Torre, B.; Kaye, J. Shared dependence on the DNA-binding factor TOX for the development of lymphoid tissue–inducer cell and NK cell lineages. Nat. Immunol. 2010, 11, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Perez-Potti, A.; Nguyen, S.; Gorin, J.B.; Wu, V.H.; Gostick, E.; Llewellyn-Lacey, S.; Hammer, Q.; Falck-Jones, S.; Vangeti, S.; et al. TOX is expressed by exhausted and polyfunctional human effector memory CD8(+) T cells. Sci. Immunol. 2020, 5, eaba7918. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Shao, Z.; Chen, L.; Zheng, Q.; Zhang, Q.; Ding, W.; Zhang, M.; Yu, Q.; Gao, D. Role, function and regulation of the thymocyte selection-associated high mobility group box protein in CD8(+) T cell exhaustion. Immunol. Lett. 2021, 229, 1–7. [Google Scholar] [CrossRef]
- Seo, H.; Chen, J.; González-Avalos, E.; Samaniego-Castruita, D.; Das, A.; Wang, Y.H.; López-Moyado, I.F.; Georges, R.O.; Zhang, W.; Onodera, A.; et al. TOX and TOX2 transcription factors cooperate with NR4A transcription factors to impose CD8(+) T cell exhaustion. Proc. Natl. Acad. Sci. USA 2019, 116, 12410–12415. [Google Scholar] [CrossRef]
- Wolf, D.; Ley, K. Immunity and Inflammation in Atherosclerosis. Circ. Res. 2019, 124, 315–327. [Google Scholar] [CrossRef]
- Leistner, D.M.; Kränkel, N.; Meteva, D.; Abdelwahed, Y.S.; Seppelt, C.; Stähli, B.E.; Rai, H.; Skurk, C.; Lauten, A.; Mochmann, H.-C.; et al. Differential immunological signature at the culprit site distinguishes acute coronary syndrome with intact from acute coronary syndrome with ruptured fibrous cap: Results from the prospective translational OPTICO-ACS study. Eur. Heart J. 2020, 41, 3549–3560. [Google Scholar] [CrossRef]
- Heim, K.; Binder, B.; Wieland, D.; Hensel, N.; Llewellyn-Lacey, S.; Gostick, E.; Price, D.A.; Emmerich, F.; Vingerhoet, H.; Kraft, A.R.; et al. TOX defines the degree of CD8+ T cell dysfunction in distinct phases of chronic HBV infection. Gut 2020, 70, 1550–1560. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Li, X.; Liu, R.; Chen, Y.; Ren, C.; Du, S. TOX correlates with prognosis, immune infiltration, and T cells exhaustion in lung adenocarcinoma. Cancer Med. 2020, 9, 6694–6709. [Google Scholar] [CrossRef]
- Mitin, N.; Rossman, K.L.; Der, C.J. Signaling Interplay in Ras Superfamily Function. Curr. Biol. 2005, 15, R563–R574. [Google Scholar] [CrossRef]
- Lambert, Q.T.; Reuther, G.W. Activation of Ras Proteins by Ras Guanine Nucleotide Releasing Protein Family Members. Methods Enzymol. 2006, 407, 82–98. [Google Scholar] [CrossRef]
- Roberts, D.M.; Anderson, A.L.; Hidaka, M.; Swetenburg, R.L.; Patterson, C.; Stanford, W.L.; Bautch, V.L. A Vascular Gene Trap Screen Defines RasGRP3 as an Angiogenesis-Regulated Gene Required for the Endothelial Response to Phorbol Esters. Mol. Cell. Biol. 2004, 24, 10515–10528. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Z.; Kovács, I.; Török, M.; Tóth, D.; Vereb, G.; Buzás, K.; Juhász, I.; Blumberg, P.M.; Bíró, T.; Czifra, G. Function of RasGRP3 in the formation and progression of human breast cancer. Mol. Cancer 2014, 13, 96. [Google Scholar] [CrossRef]
- Chen, X.; Wu, Q.; Depeille, P.; Chen, P.; Thornton, S.; Kalirai, H.; Coupland, S.E.; Roose, J.P.; Bastian, B.C. RasGRP3 Mediates MAPK Pathway Activation in GNAQ Mutant Uveal Melanoma. Cancer Cell 2017, 31, 685–696. [Google Scholar] [CrossRef]
- Chen, M.-C.; Chang, T.-H.; Chang, J.-P.; Huang, H.-D.; Ho, W.-C.; Lin, Y.-S.; Pan, K.-L.; Liu, W.-H.; Huang, Y.-K. Circulating miR-148b-3p and miR-409-3p as biomarkers for heart failure in patients with mitral regurgitation. Int. J. Cardiol. 2016, 222, 148–154. [Google Scholar] [CrossRef]
- Charrin, S.; Jouannet, S.; Boucheix, C.; Rubinstein, E. Tetraspanins at a glance. J. Cell Sci. 2014, 127, 3641–3648. [Google Scholar] [CrossRef]
- Huang, H.; Sossey-Alaoui, K.; Beachy, S.H.; Geradts, J. The tetraspanin superfamily member NET-6 is a new tumor suppressor gene. J. Cancer Res. Clin. Oncol. 2007, 133, 761–769. [Google Scholar] [CrossRef]
- Huang, Y.; Zucker, B.; Zhang, S.; Elias, S.; Zhu, Y.; Chen, H.; Ding, T.; Li, Y.; Sun, Y.; Lou, J.; et al. Migrasome formation is mediated by assembly of micron-scale tetraspanin macrodomains. Nat. Cell Biol. 2019, 21, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-N.; Chen, J.-J.; Wu, J.-H.; Chung, Y.-T.; Chen, J.-W.; Chiu, C.-H.; Liu, C.-J.; Liu, M.-T.; Chang, Y.-C.; Li, C.; et al. Association between Exosomal miRNAs and Coronary Artery Disease by Next-Generation Sequencing. Cells 2021, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, J.; Wu, M.; Xu, H. Identification of key miRNAs and mRNAs related to coronary artery disease by meta-analysis. BMC Cardiovasc. Disord. 2021, 21, 443. [Google Scholar] [CrossRef] [PubMed]
- Hall, I.F.; Climent, M.; Viviani Anselmi, C.; Papa, L.; Tragante, V.; Lambroia, L.; Farina, F.M.; Kleber, M.E.; März, W.; Biguori, C.; et al. rs41291957 controls miR-143 and miR-145 expression and impacts coronary artery disease risk. EMBO Mol. Med. 2021, 13, e14060. [Google Scholar] [CrossRef]
- Saadatian, Z.; Nariman-Saleh-Fam, Z.; Khaheshi, I.; Mansoori, Y.; Daraei, A.; Ghaderian SM, H.; Omrani, M.D. Peripheral Blood Mononuclear Cells Expression Levels of miR-196a and miR-100 in Coronary Artery Disease Patients. Immunol. Investig. 2021, 50, 914–924. [Google Scholar] [CrossRef]
- Neiburga, K.D.; Vilne, B.; Bauer, S.; Bongiovanni, D.; Ziegler, T.; Lachmann, M.; Wengert, S.; Hawe, J.S.; Güldener, U.; Westerlund, A.M.; et al. Vascular Tissue Specific miRNA Profiles Reveal Novel Correlations with Risk Factors in Coronary Artery Disease. Biomolecules 2021, 11, 1683. [Google Scholar] [CrossRef]
- Du, Y.; Hu, Y.; Wen, N.; Fu, S.; Zhang, G.; Li, L.; Liu, T.; Lv, X.; Zhang, W. Abnormal expression of TGFBR2, EGF, LRP10, and IQGAP1 is involved in the pathogenesis of coronary artery disease. Rev. Cardiovasc. Med. 2021, 22, 947–958. [Google Scholar] [CrossRef]
- Mechelinck, M.; Peschel, M.; Habigt, M.A.; Kroy, D.; Lehrke, M.; Helmedag, M.J.; Rossaint, R.; Barton, M.; Hein, M. Serum from Patients with Severe Al-coholic Liver Cirrhosis Inhibits Proliferation and Migration of Human Coronary Artery Smooth Muscle Cells. J. Clin. Med. 2021, 10, 5471. [Google Scholar] [CrossRef]
- Kim, M.; Kim, H.B.; Park, D.S.; Cho, K.H.; Hyun, D.Y.; Kee, H.J.; Hong, Y.J.; Jeong, M.H. A model of atherosclerosis using nicotine with balloon overdilation in a porcine. Sci. Rep. 2021, 11, 13695. [Google Scholar] [CrossRef] [PubMed]
- Kassiteridi, C.; Cole, J.E.; Griseri, T.; Falck-Hansen, M.; Goddard, M.E.; Seneviratne, A.N.; Green, P.A.; Park, I.; Shami, A.G.; Pattarabanjird, T.; et al. CD200 Limits Monopoiesis and Monocyte Recruitment in Atherosclerosis. Circ. Res. 2021, 129, 280–295. [Google Scholar] [CrossRef]
- Liu, S.; Gao, J.; Wang, S. HOXA9 inhibitors promote microcirculation of coronary arteries in rats via downregulating E-selectin/VCAM-1. Exp. Ther. Med. 2021, 22, 871. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | No. | Sex | Age | PMI | Cause of Death | Score of IHC Staining | ||
|---|---|---|---|---|---|---|---|---|
| (years) | (days) | TOX | RasGRP3 | TSPAN13 | ||||
| A | 1 | M | 30 | 11 | poisoning | 1 | 1 | 2 |
| A | 2 | M | 25 | 2 | accident | 0 | 1 | 1 |
| A | 3 | F | 34 | 4 | cardiomyopathy | 1 | 2 | 2 |
| A | 4 | M | 22 | 1 | head injury | 2 | 1 | 1 |
| A | 5 | M | 30 | 5 | electric shock | 1 | 1 | 1 |
| A | 6 | M | 26 | 4 | pneumonia | 1 | 1 | 2 |
| A | 7 | M | 37 | 3 | head injury | 0 | 2 | 1 |
| B | 8 | M | 46 | 1 | head injury | 1 | 1 | 1 |
| B | 9 | M | 56 | 23 | cardiomyopathy | 2 | 1 | 1 |
| B | 10 | M | 44 | 2 | chest injury | 1 | 0 | 1 |
| B | 11 | M | 57 | 3 | pulmonary embolism | 1 | 1 | 1 |
| B | 12 | F | 47 | 5 | cardiomyopathy | 2 | 0 | 1 |
| B | 13 | M | 52 | 1 | liver cancer | 3 | 0 | 1 |
| B | 14 | M | 29 | 8 | alcoholism | 2 | 1 | 1 |
| B | 15 | M | 49 | 8 | rupture of spleen | 3 | 1 | 1 |
| Probe_id | Gene Symbol | Log2FC | p Value | adj. p Value | t | B | Change |
|---|---|---|---|---|---|---|---|
| 205572_at | ANGPT2 | −1.853 | 0.002 | 0.363 | −3.613 | −1.228 | down |
| 204948_s_at | FST | 1.430 | 0.003 | 0.379 | 3.356 | −1.690 | up |
| 204529_s_at | TOX | 1.248 | 0.004 | 0.391 | 3.221 | −1.932 | up |
| 202391_at | BASP1 | 1.255 | 0.002 | 0.369 | 3.566 | −1.314 | up |
| 213110_s_at | COL4A5 | −1.160 | 0.001 | 0.363 | −3.683 | −1.103 | down |
| 204235_s_at | GULP1 | 1.084 | 0.003 | 0.372 | 3.413 | −1.589 | up |
| 205801_s_at | RASGRP3 | −1.104 | 0.004 | 0.391 | −3.201 | −1.966 | down |
| 207808_s_at | PROS1 | −1.030 | 0.003 | 0.372 | −3.420 | −1.575 | down |
| 217979_at | TSPAN13 | −1.018 | 0.002 | 0.363 | −3.643 | −1.174 | down |
| 204992_s_at | PFN2 | 1.026 | 0.005 | 0.393 | 3.151 | −2.056 | up |
| 209230_s_at | NUPR1 | −1.073 | 0.006 | 0.394 | −3.085 | −2.172 | down |
| 207526_s_at | IL1RL1 | 1.578 | 0.006 | 0.394 | 3.068 | −2.202 | up |
| 212950_at | ADGRF5 | −1.511 | 0.011 | 0.404 | −2.803 | −2.664 | down |
| 206710_s_at | EPB41L3 | 1.029 | 0.012 | 0.404 | 2.755 | −2.746 | up |
| 202345_s_at | FABP5 | −1.027 | 0.012 | 0.404 | −2.747 | −2.760 | down |
| 209277_at | TFPI2 | −1.515 | 0.015 | 0.410 | −2.658 | −2.909 | down |
| 203980_at | FABP4 | −1.740 | 0.018 | 0.424 | −2.575 | −3.049 | down |
| 202546_at | VAMP8 | 1.418 | 0.021 | 0.428 | 2.502 | −3.170 | up |
| 219148_at | PBK | 1.063 | 0.027 | 0.441 | 2.396 | −3.343 | up |
| 206029_at | ANKRD1 | 1.131 | 0.029 | 0.447 | 2.356 | −3.407 | up |
| 205680_at | MMP10 | −1.153 | 0.030 | 0.447 | −2.339 | −3.434 | down |
| 204438_at | MRC1 | −1.376 | 0.040 | 0.463 | −2.201 | −3.650 | down |
| 205110_s_at | FGF13 | −1.013 | 0.049 | 0.480 | −2.090 | −3.819 | down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, F.; Zeng, Y.; Zhao, M.; Zhu, Y.; Li, J.; Tang, R. Identification of Differentially Expressed Genes and Prediction of Expression Regulation Networks in Dysfunctional Endothelium. Genes 2022, 13, 1563. https://doi.org/10.3390/genes13091563
Cheng F, Zeng Y, Zhao M, Zhu Y, Li J, Tang R. Identification of Differentially Expressed Genes and Prediction of Expression Regulation Networks in Dysfunctional Endothelium. Genes. 2022; 13(9):1563. https://doi.org/10.3390/genes13091563
Chicago/Turabian StyleCheng, Fang, Yujie Zeng, Minzhu Zhao, Ying Zhu, Jianbo Li, and Renkuan Tang. 2022. "Identification of Differentially Expressed Genes and Prediction of Expression Regulation Networks in Dysfunctional Endothelium" Genes 13, no. 9: 1563. https://doi.org/10.3390/genes13091563