
Expression and Prognostic Value of Chromobox Family Proteins in Esophageal Cancer
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Materials
2.1. Oncomine
2.2. GEPIA2
2.3. UALCAN
2.4. Kaplan–Meier Plotter
2.5. cBioPortal
2.6. Metascape
2.7. TIMER2
2.8. TISIDB
2.9. Cell Culture and Transfection
2.10. RNA Isolation and Real-Time Quantitative PCR
2.11. Western Blot Assay
2.12. Colony Formation and Cell Viability Assays
2.13. Wound Healing and Transwell Assays
2.14. Statistical Analysis
3. Results
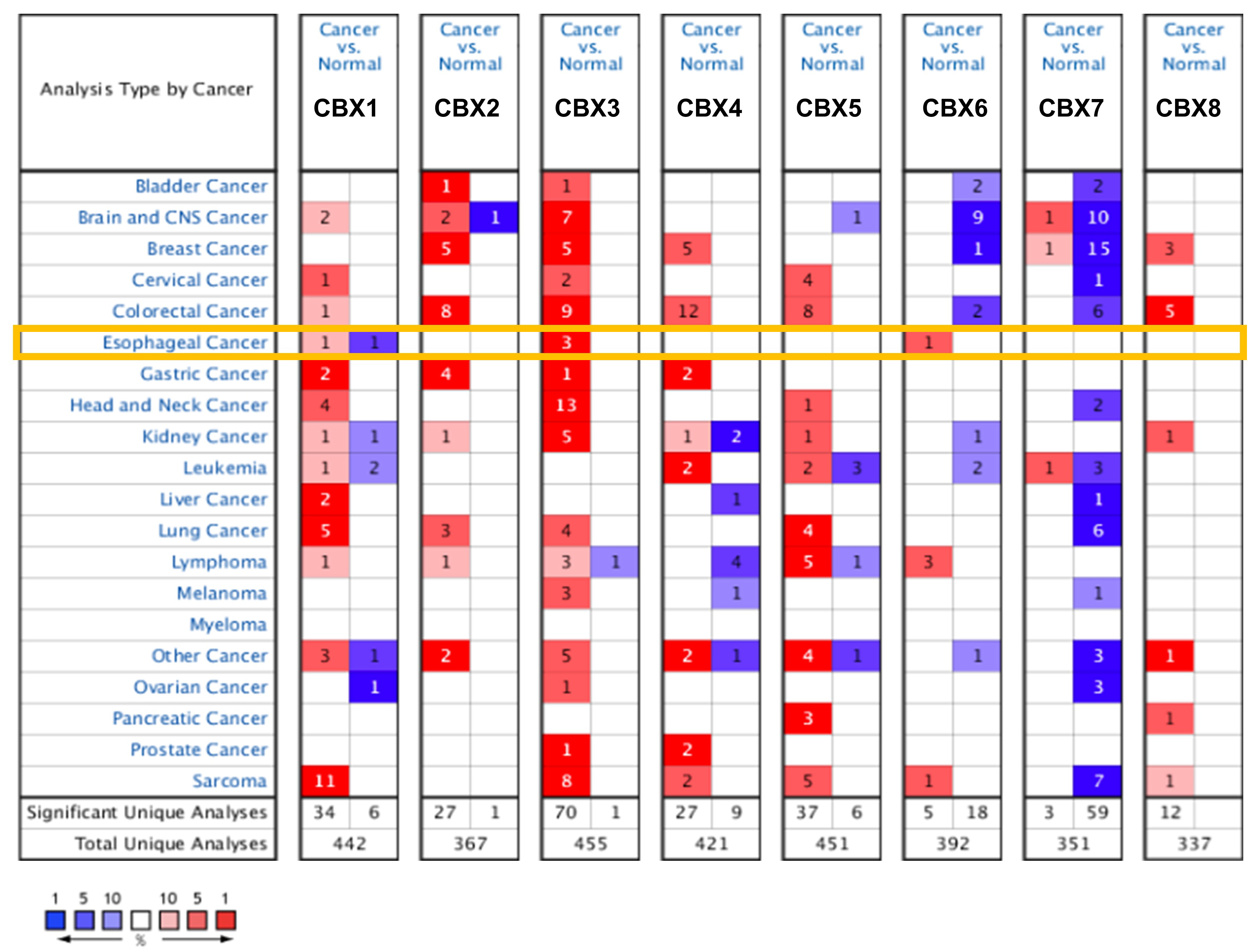
3.1. Expression Profile of CBX Family Proteins in EC
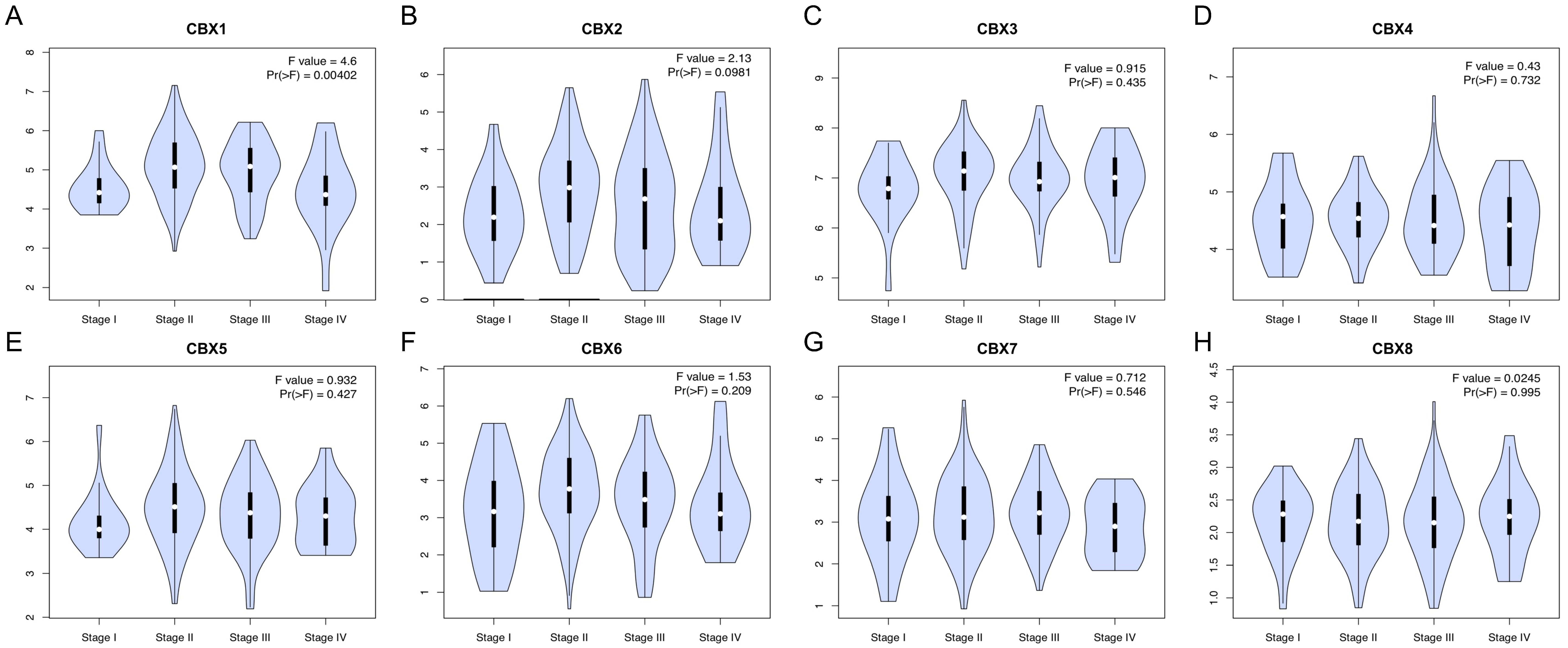
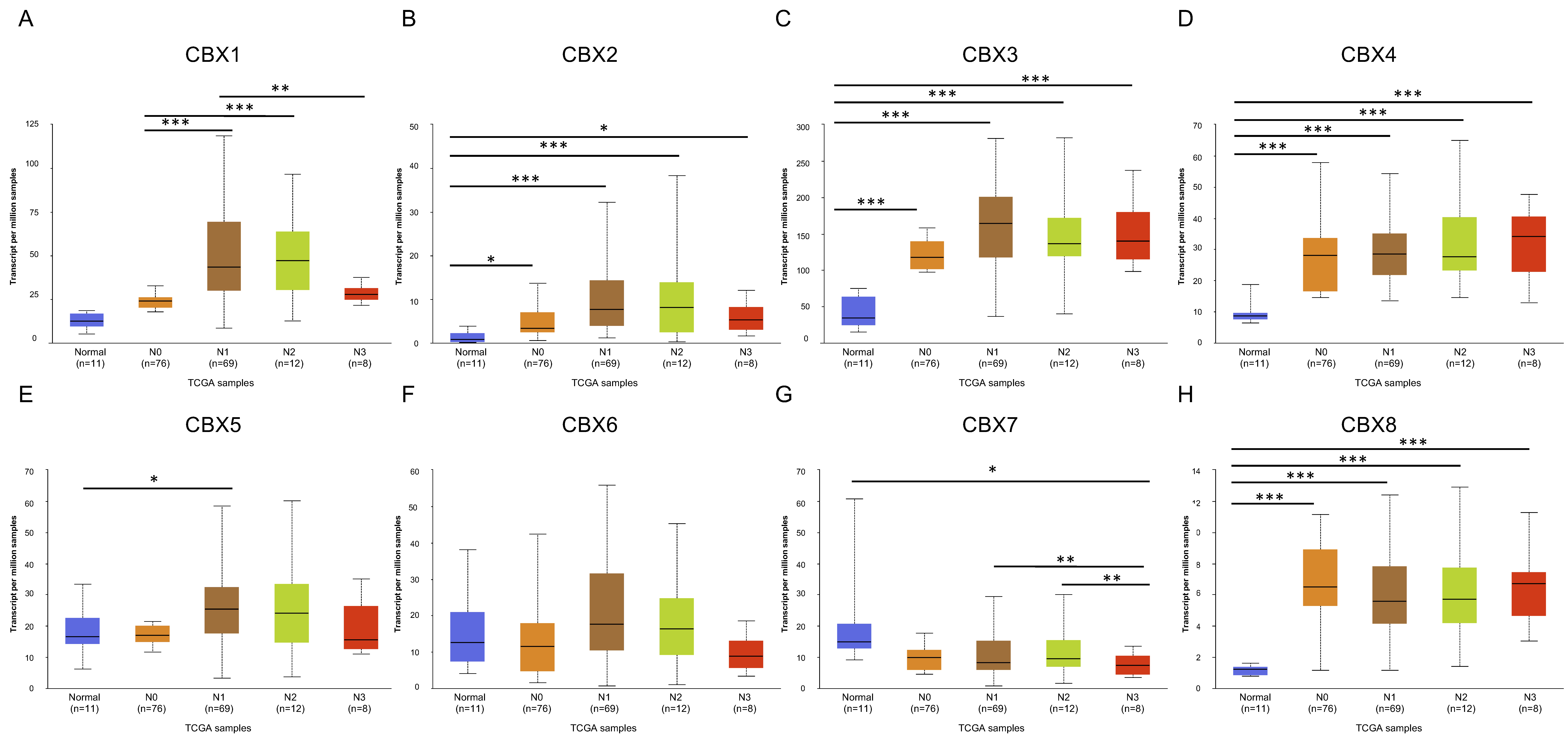
3.2. Relationships between CBX Family Proteins and Clinicopathologic Variables in EC
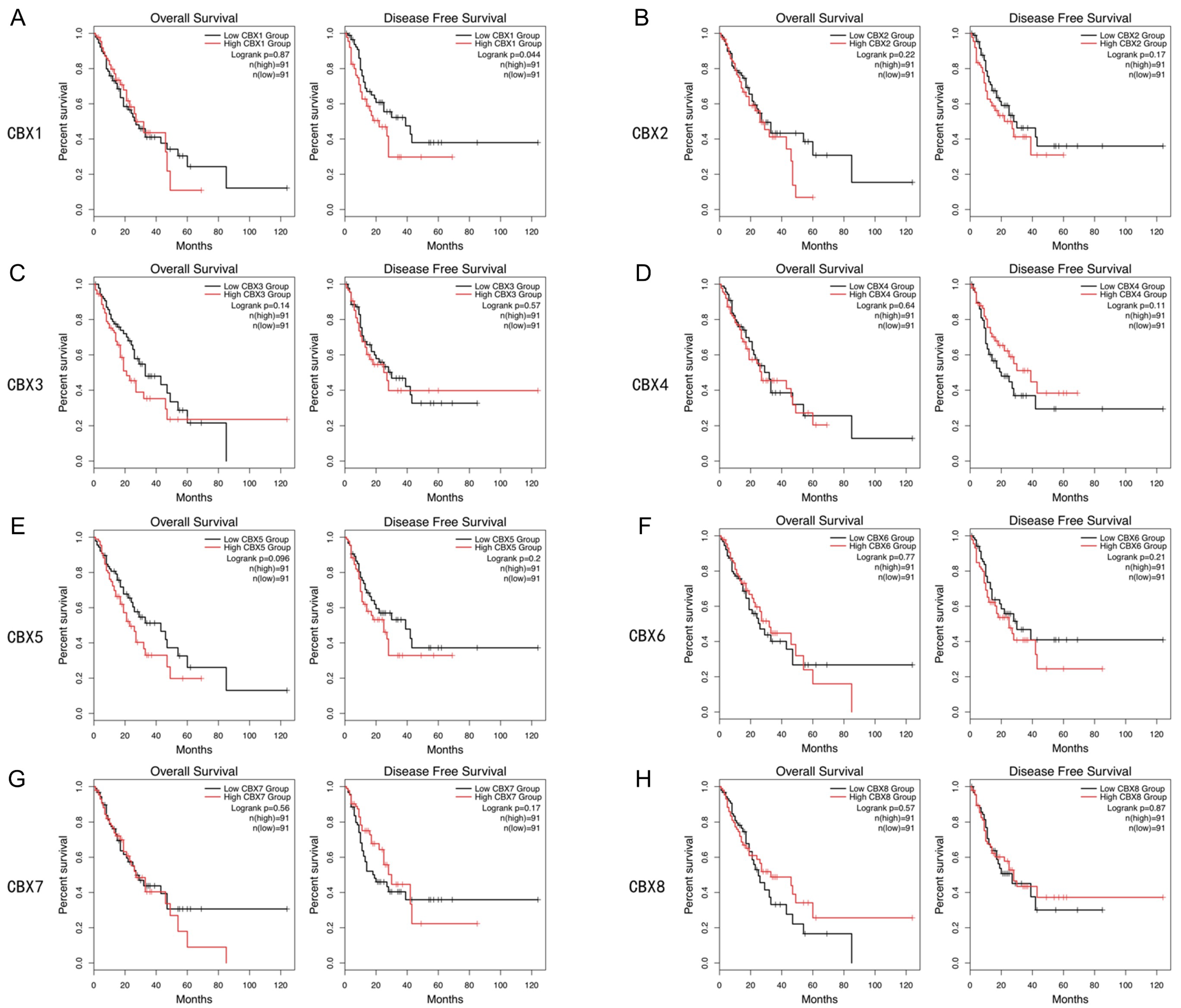
3.3. Prognostic Roles of CBX Family Proteins in EC
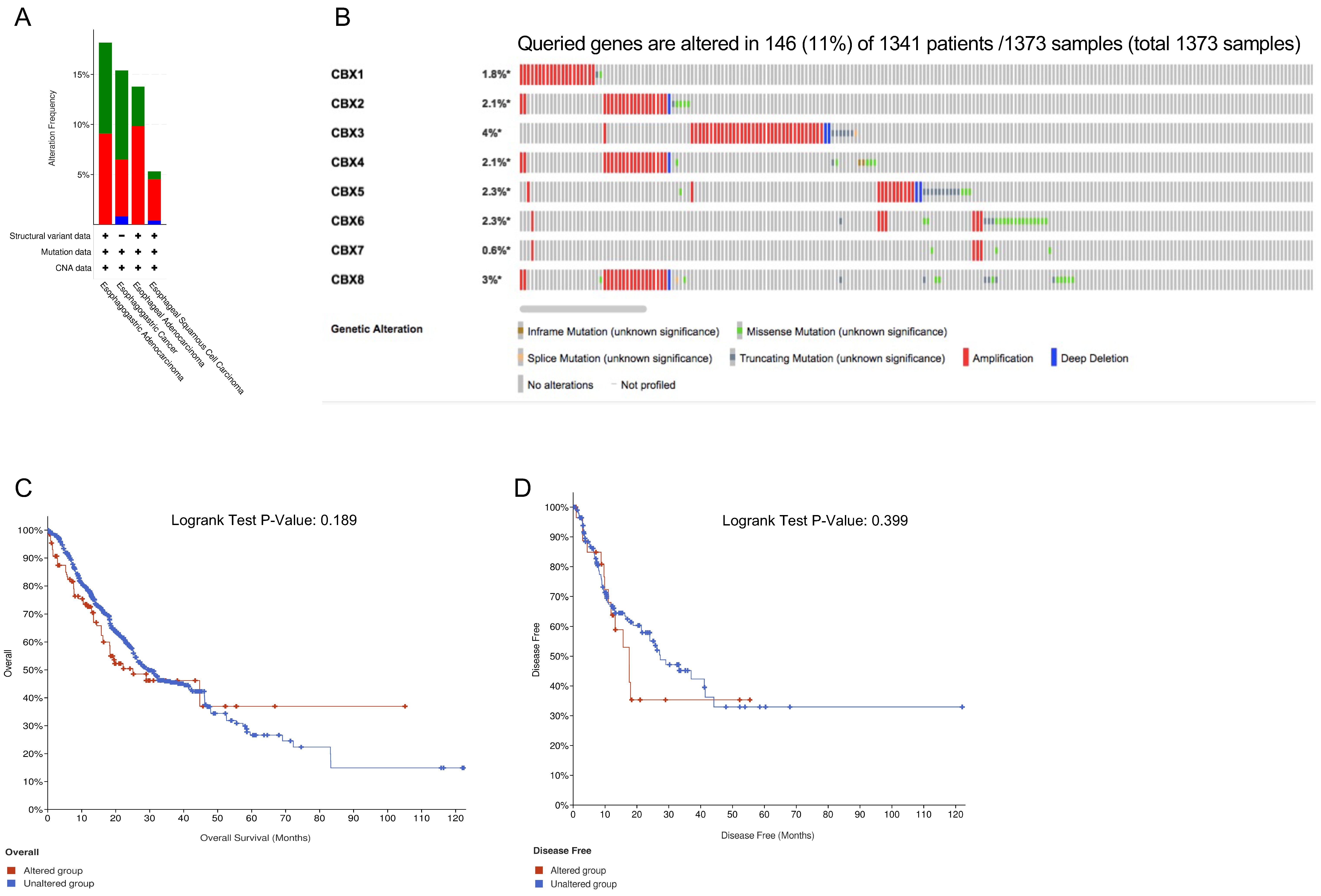
3.4. Genetic Alterations of CBX Family Proteins and the Related Prognostic Value in EC
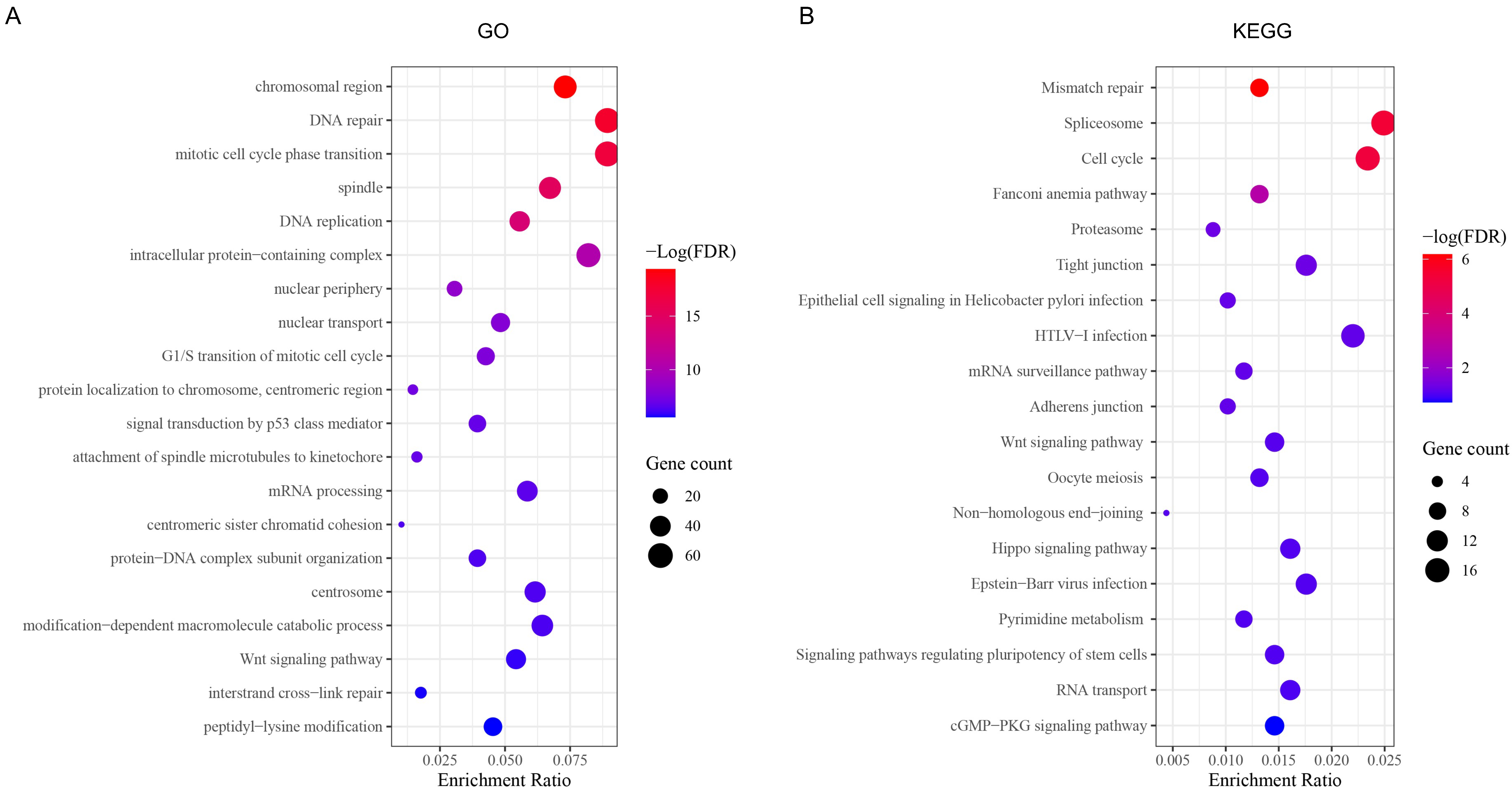
3.5. GO and KEGG Analysis
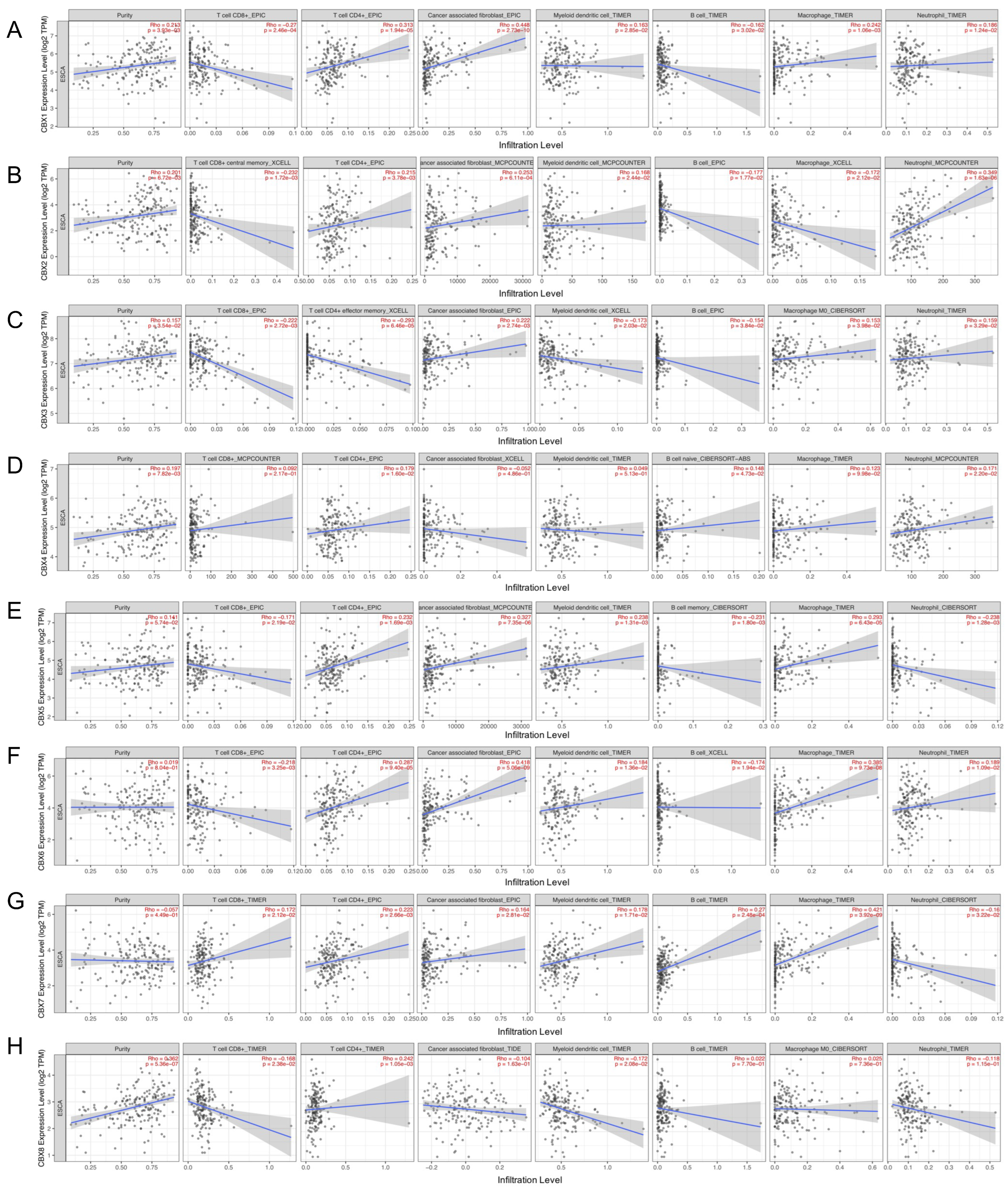
3.6. Relationships between CBX Family Proteins and Immune Cells Infiltration Level and Immune Regulators in EC
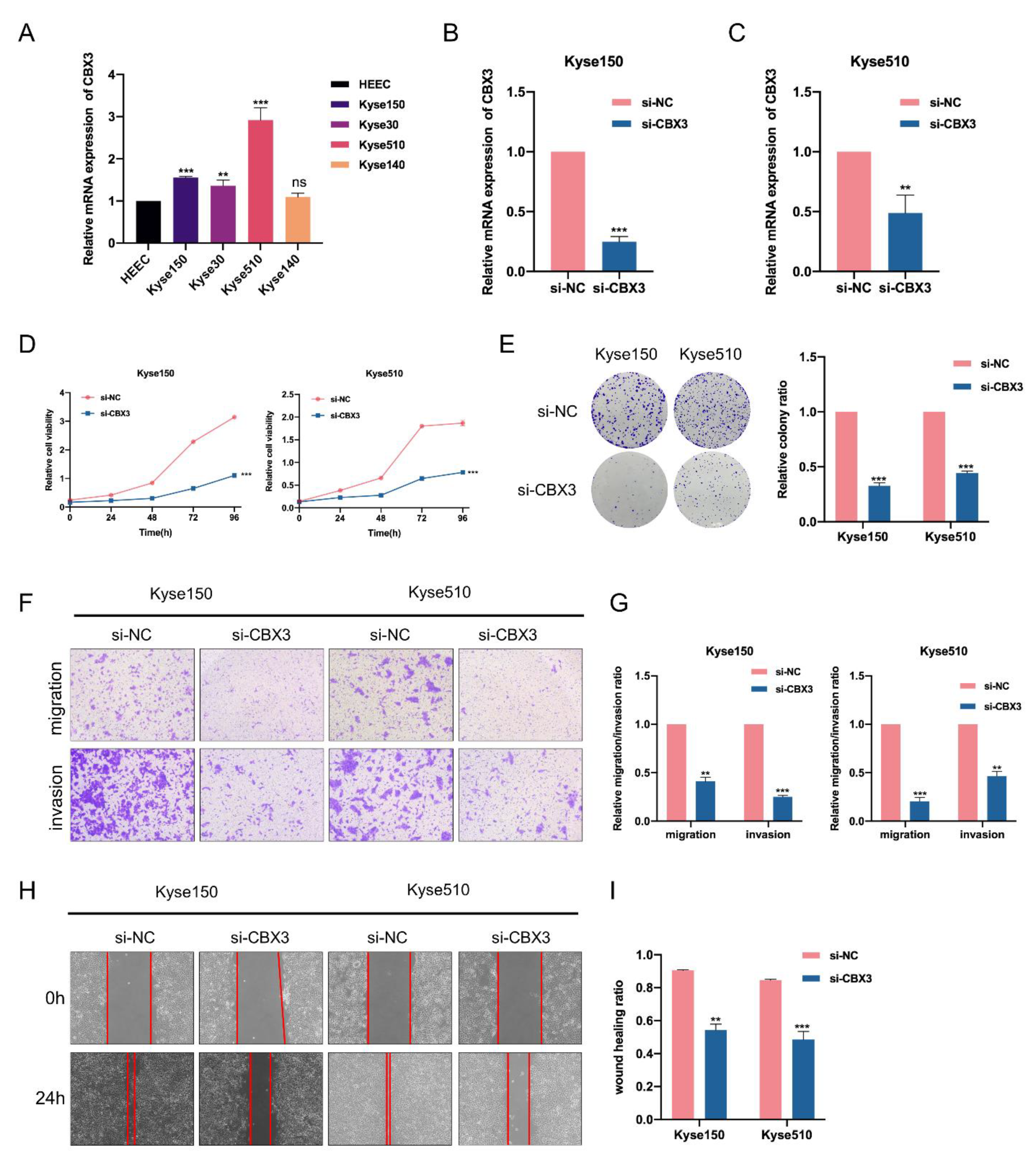
3.7. CBX3 Promotes Cell Proliferation, Migration and Invasion of EC Cells In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Smyth, E.C.; Lagergren, J.; Fitzgerald, R.C.; Lordick, F.; Shah, M.A.; Lagergren, P.; Cunningham, D. Oesophageal cancer. Nat. Rev. Dis. Primers 2017, 3, 17048. [Google Scholar] [CrossRef] [PubMed]
- Huynh, J.; Patel, K.; Gong, J.; Cho, M.; Malla, M.; Parikh, A.; Klempner, S. Immunotherapy in Gastroesophageal Cancers: Current Evidence and Ongoing Trials. Curr. Treat. Options Oncol. 2021, 22, 100. [Google Scholar] [CrossRef]
- Yang, Y.-M.; Hong, P.; Xu, W.W.; He, Q.-Y.; Li, B. Advances in targeted therapy for esophageal cancer. Signal Transduct. Target. Ther. 2020, 5, 229. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.-M.; Shen, L.; Shah, M.A.; Enzinger, P.; Adenis, A.; Doi, T.; Kojima, T.; Metges, J.-P.; Li, Z.; Kim, S.-B.; et al. Pembrolizumab plus chemotherapy versus chemotherapy alone for first-line. Lancet 2021, 398, 759–771. [Google Scholar] [CrossRef]
- Lu, Z.; Wang, J.; Shu, Y.; Liu, L.; Kong, L.; Yang, L.; Wang, B.; Sun, G.; Ji, Y.; Cao, G.; et al. Sintilimab versus placebo in combination with chemotherapy as first line. BMJ 2022, 377, e068714. [Google Scholar] [CrossRef]
- Kojima, T.; Shah, M.A.; Muro, K.; Francois, E.; Adenis, A.; Hsu, C.-H.; Doi, T.; Moriwaki, T.; Kim, S.-B.; Lee, S.-H.; et al. Randomized Phase III KEYNOTE-181 Study of Pembrolizumab Versus Chemotherapy in. J. Clin. Oncol. 2020, 38, 4138–4148. [Google Scholar] [CrossRef]
- Yamashina, T.; Ishihara, R.; Nagai, K.; Matsuura, N.; Matsui, F.; Ito, T.; Fujii, M.; Yamamoto, S.; Hanaoka, N.; Takeuchi, Y.; et al. Long-term outcome and metastatic risk after endoscopic resection of superficial. Am. J. Gastroenterol. 2013, 108, 544–551. [Google Scholar] [CrossRef]
- Visaggi, P.; Barberio, B.; Ghisa, M.; Ribolsi, M.; Savarino, V.; Fassan, M.; Vamasoni, M.; Marchi, S.; de Bortoli, N.; Savarino, E. Modern Diagnosis of Early Esophageal Cancer: From Blood Biomarkers to Advanced. Cancers 2021, 13, 3162. [Google Scholar] [CrossRef]
- Jiang, W.; Xia, T.; Liu, C.; Li, J.; Zhang, W.; Sun, C. Remodeling the Epigenetic Landscape of Cancer-Application Potential of Flavonoids in. Front. Oncol. 2021, 11, 2331. [Google Scholar]
- Ma, R.-G.; Zhang, Y.; Sun, T.-T.; Cheng, B. Epigenetic regulation by polycomb group complexes: Focus on roles of CBX proteins. J. Zhejiang Univ. Sci. B 2014, 15, 412–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolton, D.; Merrill, J.C. Pc2 and SUMOylation. Biochem. Soc. Trans. 2007, 35 Pt 6, 1401–1404. [Google Scholar]
- Vincenz, C.; Kerppola, T.K. Different polycomb group CBX family proteins associate with distinct regions of. Proc. Natl. Acad. Sci. USA 2008, 105, 16572–16577. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhang, Z.; Pan, L.; Zhou, Y. Identification of miR-758-3p as Potential Modulator of CBX5 Expression in Gastric Cancer. Technol. Cancer Res. Treat. 2018, 17, 1533033818816061. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-W.; Zhang, L.; Qin, W.; Yao, X.-H.; Zheng, L.-Z.; Liu, X.; Li, J.; Guo, W.-J. Oncogenic role of the chromobox protein CBX7 in gastric cancer. J. Exp. Clin. Cancer Res. 2010, 29, 144. [Google Scholar] [CrossRef] [PubMed]
- Ni, S.-J.; Zhao, L.-Q.; Wang, X.-F.; Wu, Z.-H.; Hua, R.-X.; Wan, C.-H.; Zhang, J.-Y.; Zhang, X.-W.; Huang, M.-Z.; Gan, L.; et al. CBX7 regulates stem cell-like properties of gastric cancer cells via p16 and. J. Hematol. Oncol. 2018, 11, 17. [Google Scholar] [CrossRef]
- Yang, Y.F.; Pan, Y.H.; Tian, Q.H.; Wu, D.C.; Su, S.G. CBX1 Indicates Poor Outcomes and Exerts Oncogenic Activity in Hepatocellular Carcinoma. Transl. Oncol. 2018, 11, 1110–1118. [Google Scholar] [CrossRef]
- Liang, Y.K.; Lin, H.Y.; Chen, C.F.; Zeng, D. Prognostic values of distinct CBX family members in breast cancer. Oncotarget 2017, 8, 92375–92387. [Google Scholar] [CrossRef]
- Hu, C.; Zhang, Q.; Tang, Q.; Zhou, H.; Liu, W.; Huang, J.; Liu, Y.; Wang, Q.; Zhang, J.; Zhou, M.; et al. CBX4 promotes the proliferation and metastasis via regulating BMI-1 in lung cancer. J. Cell. Mol. Med. 2020, 24, 618–631. [Google Scholar] [CrossRef]
- Zhou, H.; Xiong, Y.; Liu, Z.; Hou, S.; Zhou, T. Expression and prognostic significance of CBX2 in colorectal cancer: Database mining for CBX family members in malignancies and vitro analyses. Cancer Cell Int. 2021, 21, 402. [Google Scholar] [CrossRef]
- Chen, L.Y.; Cheng, C.S.; Qu, C.; Wang, P.; Chen, H.; Meng, Z.Q.; Chen, Z. CBX3 promotes proliferation and regulates glycolysis via suppressing FBP1 in pancreatic cancer. Biochem. Biophys. Res. Commun. 2018, 500, 691–697. [Google Scholar] [PubMed]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pander, A.; Chinnaiyan, A.M. ONCOMINE: A Cancer Microarray Database and Integrated Data-Mining Platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Nagy, A.; Lanczky, A.; Menyhart, O.; Gyorffy, B. Validation of miRNA prognostic power in hepatocellular carcinoma using expression data of independent datasets. Sci. Rep. 2018, 8, 9227. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Ru, B.; Wong, C.N.; Tong, Y.; Zhong, J.Y.; Zhong, S.S.W.; Wu, W.C.; Chu, K.C.; Wong, C.Y.; Lau, C.Y.; Chen, I.; et al. TISIDB: An integrated repository portal for tumor-immune system interactions. Bioinformatics 2019, 35, 4200–4202. [Google Scholar] [CrossRef]
- Xie, H.; Li, J.; Ying, Y.; Yan, H.; Jin, K.; Ma, X.; He, L.; Xu, X.; Liu, B.; Wang, X.; et al. METTL3/YTHDF2 m6A axis promotes tumorigenesis by degrading SETD7 and KLF4. J. Cell. Mol. Med. 2020, 24, 4092–4104. [Google Scholar] [CrossRef]
- Sun, J.; Ma, X.; Ying, Y.; Wang, W.; Shen, H.; Wang, S.; Xie, H.; Yi, J.; Zhan, W.; Li, J.; et al. SMAD3 and FTO are involved in miR-5581-3p-mediated inhibition of cell migration. Cell Death Discov. 2022, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Ying, Y.; Sun, J.; Xie, H.; Li, J.; He, L.; Wang, W.; Chen, S.; Shen, H.; Yi, J.; et al. circKDM4C enhances bladder cancer invasion and metastasis through. Cell Death Discov. 2021, 7, 365. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Kan, A.; Zhang, W.; Zhou, J.; Zhang, H.; Chen, J.; Tang, S. CBX3/HP1γ promotes tumor proliferation and predicts poor survival in. Aging 2019, 11, 5483–5497. [Google Scholar] [CrossRef]
- Ma, C.; Nie, X.G.; Wang, Y.L.; Liu, X.H.; Liang, X.; Zhou, Q.L.; Wu, D.P. CBX3 predicts an unfavorable prognosis and promotes tumorigenesis in osteosarcoma. Mol. Med. Rep. 2019, 19, 4205–4212. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Huang, F.; Zhang, D.; Ju, J.; Wu, X.B.; Wang, Y.; Wang, Y.; Wu, Y.; Nie, M.; Li, Z.; et al. Heterochromatin protein HP1gamma promotes colorectal cancer progression and is regulated by miR-30a. Cancer Res. 2015, 75, 4593–4604. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Liu, J.; He, W.; Qu, M.; Huang, X.; Deng, Y.; Shen, L.; Zhao, X.; Guo, H.; Jiang, J.; et al. A regulatory circuit HP1γ/miR-451a/c-Myc promotes prostate cancer progression. Oncogene 2018, 37, 415–426. [Google Scholar] [CrossRef]
- Shiota, M.; Song, Y.; Yokomizo, A.; Tada, Y.; Kuroiwa, K.; Eto, M.; Oda, Y.; Inokuchi, J.; Uchiumi, T.; Fujimoto, N.; et al. Human heterochromatin protein 1 isoform HP1beta enhances androgen receptor activity and is implicated in prostate cancer growth. Endocr. Relat. Cancer. 2010, 17, 455–467. [Google Scholar] [CrossRef]
- Tretiakova, M.S.; Bond, S.D.; Wheeler, D.; Contreras, A.; Kocherginsky, M.; Kroll, T.G.; Hale, T.K. Heterochromatin protein 1 expression is reduced in human thyroid malignancy. Lab. Investig. 2014, 94, 788–795. [Google Scholar] [CrossRef]
- Lin, H.; Lian, J.; Xia, L.; Guan, G.; You, J. CBX3 Promotes Gastric Cancer Progression and Affects Factors Related to Immunotherapeutic Responses. Cancer Manag. Res. 2020, 12, 10113–10125. [Google Scholar] [CrossRef]
- Chang, S.-C.; Lai, Y.-C.; Chen, Y.-C.; Wang, N.-K.; Wang, W.-S.; Lai, J.-I. CBX3/heterochromatin protein 1 gamma is significantly upregulated in patients. Asia-Pac. J. Clin. Oncol. 2018, 14, e283–e288. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, X.; Zha, Z.; Zhu, Y.; Zhang, G.; Li, G. CBX3 regulated by miR-139 promotes the development of HCC by regulating cell. Cell Cycle 2022, 21, 1740–1752. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, W.; Wang, P.; Zhang, Q.; Cong, A.; Yang, X.; Sang, K. LncRNA SNHG11 aggravates cell proliferation and migration in triple-negative breast cancer via sponging miR-2355-5p and targeting CBX5. Exp. Ther. Med. 2021, 22, 892. [Google Scholar] [CrossRef] [PubMed]
- Pongas, G.; Kim, M.K.; Min, D.J.; House, C.D.; Jordan, E.; Caplen, N.; Chakka, S.; Ohiri, J.; Kruhlak, M.J.; Annunziata, C.M. BRD4 facilitates DNA damage response and represses CBX5/Heterochromatin protein 1. Oncotarget 2017, 8, 51402–51415. [Google Scholar] [CrossRef]
- Wheeler, L.J.; Watson, Z.L.; Qamar, L.; Yamamoto, T.M.; Post, M.D.; Berning, A.A.; Spillman, M.A.; Behbakht, K.; Bitler, B.G. CBX2 identified as driver of anoikis escape and dissemination in high grade serous ovarian cancer. Oncogenesis 2018, 7, 92. [Google Scholar] [CrossRef]
- Zheng, S.; Lv, P.; Su, J.; Miao, L.; Xu, H.; Li, M. Overexpression of CBX2 in breast cancer promotes tumor progression through the. Am. J. Transl. Res. 2019, 11, 1668–1682. [Google Scholar]
- Han, Q.; Li, C.; Cao, Y.; Bao, J.; Li, K.; Song, R.; Chen, X.; Li, J.; Wu, X. CBX2 is a functional target of miRNA let-7a and acts as a tumor promoter in osteosarcoma. Cancer Med. 2019, 8, 3981–3991. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qin, G.; Liang, X.; Wang, W.; Wang, Z.; Liao, D.; Zhong, L.; Zhang, R.; Zeng, Y.X.; Wu, Y.; et al. Targeting the CK1alpha/CBX4 axis for metastasis in osteosarcoma. Nat. Commun. 2020, 11, 1141. [Google Scholar] [CrossRef]
- Zeng, J.S.; Zhang, Z.D.; Pei, L.; Bai, Z.Z.; Yang, Y.; Yang, H.; Tian, Q.H. CBX4 exhibits oncogenic activities in breast cancer via Notch1 signaling. Int. J. Biochem. Cell Biol. 2018, 95, 1–8. [Google Scholar] [CrossRef]
- Zhao, W.; Ma, B.; Tian, Z.; Han, H.; Tang, J.; Dong, B.; An, G.; Cao, B.; Wang, B. Inhibiting CBX4 efficiently protects hepatocellular carcinoma cells against sorafenib resistance. Br. J. Cancer 2021, 124, 1237–1248. [Google Scholar] [CrossRef]
- Deng, H.; Guan, X.; Gong, L.; Zeng, J.; Zhang, H.; Chen, M.Y.; Li, G. CBX6 is negatively regulated by EZH2 and plays a potential tumor suppressor role in breast cancer. Sci. Rep. 2019, 9, 197. [Google Scholar] [CrossRef]
- Sakai, K.; Nishiuchi, T.; Tange, S.; Suzuki, Y.; Yano, S.; Terashima, M.; Suzuki, T.; Matsumoto, K. Proteasomal degradation of polycomb-group protein CBX6 confers MMP-2 expression essential for mesothelioma invasion. Sci. Rep. 2020, 10, 16678. [Google Scholar] [CrossRef] [PubMed]
- Pallante, P.; Terracciano, L.; Carafa, V.; Schneider, S.; Zlobec, I.; Lugli, A.; Bianco, M.; Ferraro, A.; Sacchetti, S.; Troncone, G.; et al. The loss of the CBX7 gene expression represents an adverse prognostic marker for. Eur. J. Cancer 2010, 46, 2304–2313. [Google Scholar] [CrossRef] [PubMed]
- Karamitopoulou, E.; Pallante, P.; Zlobec, I.; Tornillo, L.; Carafa, V.; Schaffner, T.; Borner, M.; Diamantis, I.; Esposito, F.; Brunner, T.; et al. Loss of the CBX7 protein expression correlates with a more aggressive phenotype in pancreatic cancer. Eur. J. Cancer 2010, 46, 1438–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallante, P.; Federico, A.; Berlingieri, M.T.; Bianco, M.; Ferraro, A.; Forzati, F.; Iaccarino, A.; Russo, M.; Pierantoni, G.M.; Leone, V.; et al. Loss of the CBX7 gene expression correlates with a highly malignant phenotype in thyroid cancer. Cancer Res. 2008, 68, 6770–6778. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gou, J.; Li, H.; Yang, X. Bioinformatic analysis of the expression and prognostic value of chromobox family proteins in human breast cancer. Sci. Rep. 2020, 10, 17739. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Park, J.H.; Won, H.Y.; Lee, J.Y.; Kong, G. CBX7 inhibits breast tumorigenicity through DKK-1-mediated suppression of the Wnt/β-catenin pathway. FASEB J. 2014, 29, 300–313. [Google Scholar] [CrossRef]
- Ni, S.; Wang, H.; Zhu, X.; Wang, C.; Xu, J.; Lu, C.; Xiao, L.; He, J.; Jiang, C.; Wang, W.; et al. CBX7 suppresses cell proliferation, migration, and invasion through the inhibition. Oncotarget 2017, 8, 8010–8021. [Google Scholar] [CrossRef]
- Bernard, D.; Martinez-Leal, J.F.; Rizzo, S.; Martinez, D.; Hudson, D.; Visakorpi, T.; Peters, G.; Carnero, A.; Beach, D.; Gil, J. CBX7 controls the growth of normal and tumor-derived prostate cells by repressing. Oncogene 2005, 24, 5543–5551. [Google Scholar] [CrossRef]
- Scott, C.L.; Gil, J.; Hernando, E.; Teruya-Feldstein, J.; Narita, M.; Martínez, D.; Visakorpi, T.; Mu, D.; Cordon-Cardo, C.; Peters, G.; et al. Role of the chromobox protein CBX7 in lymphomagenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 5389–5394. [Google Scholar] [CrossRef]
- Tang, J.; Wang, G.; Zhang, M.; Li, F.-Y.; Sang, Y.; Wang, B.; Hu, K.; Wu, Y.; Luo, R.; Liao, D.; et al. Paradoxical role of CBX8 in proliferation and metastasis of colorectal cancer. Oncotarget 2014, 5, 10778–10790. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Y.; Cheng, C.; Cui, H.; Cheng, L.; Kong, P.; Wang, J.; Li, Y.; Chen, W.; Song, B.; et al. Genomic analyses reveal mutational signatures and frequently altered genes in esophageal squamous cell carcinoma. Am. J. Hum. Genet. 2015, 96, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Tang, J.; Zhan, W.; Zhang, R.; Zhang, M.; Liao, D.; Wang, X.; Wu, Y.; Kang, T. CBX8 Suppresses Tumor Metastasis via Repressing Snail in Esophageal Squamous Cell Carcinoma. Theranostics 2017, 7, 3478–3488. [Google Scholar] [CrossRef]
- Hanahan, D.; Coussens, L.M. Accessories to the crime: Functions of cells recruited to the tumor microenvironment. Cancer Cell 2012, 21, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBleu, V.S. Imaging the Tumor Microenvironment. Cancer J. 2015, 21, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y. Tumor microenvironment and cancer therapy resistance. Cancer Lett. 2016, 380, 205–215. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, Z.; Zou, M.; Wan, R.; Wu, T.; Luo, Y.; Wu, G.; Wang, W.; Liu, T. Prognosis and Immune Infiltration of Chromobox Family Genes in Sarcoma. Front. Oncol. 2021, 11, 1523. [Google Scholar] [CrossRef]
- Li, Q.; Pan, Y.; Cao, Z.; Zhao, S. Comprehensive Analysis of Prognostic Value and Immune Infiltration of Chromobox. Front. Oncol. 2020, 10, 582667. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Shen, H.; Chen, X.; Ding, Y.; Wang, H.; Xu, N.; Teng, L. Expression and Prognostic Value of Chromobox Family Proteins in Esophageal Cancer. Genes 2022, 13, 1582. https://doi.org/10.3390/genes13091582
Liu J, Shen H, Chen X, Ding Y, Wang H, Xu N, Teng L. Expression and Prognostic Value of Chromobox Family Proteins in Esophageal Cancer. Genes. 2022; 13(9):1582. https://doi.org/10.3390/genes13091582
Chicago/Turabian StyleLiu, Jin, Haixiang Shen, Xiangliu Chen, Yongfeng Ding, Haiyong Wang, Nong Xu, and Lisong Teng. 2022. "Expression and Prognostic Value of Chromobox Family Proteins in Esophageal Cancer" Genes 13, no. 9: 1582. https://doi.org/10.3390/genes13091582