
Variations in Fibrinogen-like 1 (FGL1) Gene Locus as a Genetic Marker Related to Fat Deposition Based on Pig Model and Liver RNA-Seq Data



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Methods
2.2. Library Construction, Sequencing, and Aligning Raw Reads to the Pig Transcriptome
2.3. Transcript Variant Identification
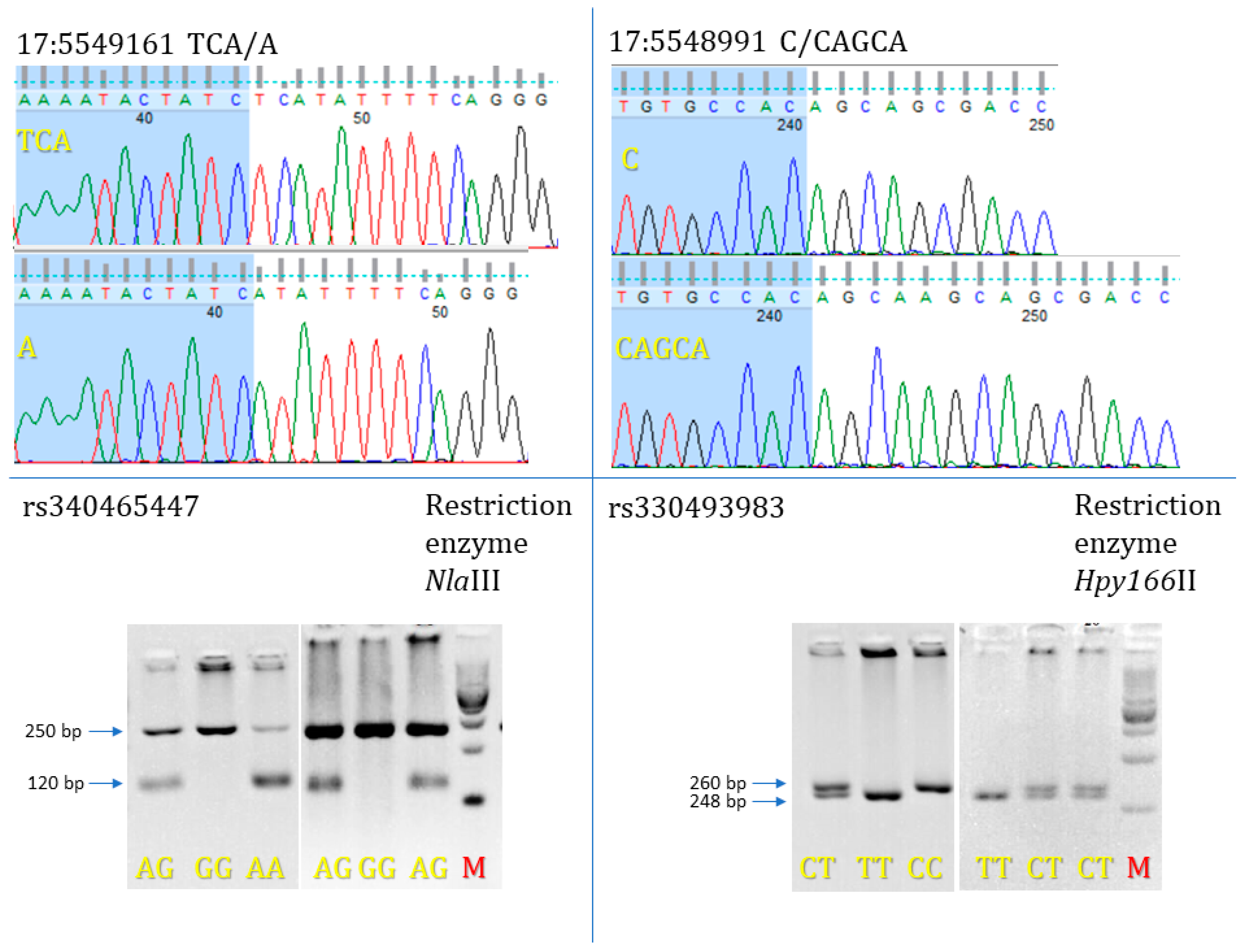
2.4. FGL1 Genotyping, Frequency Estimation, and Statistical Analyses
3. Results
3.1. Animals Used in the RNA-Seq Method
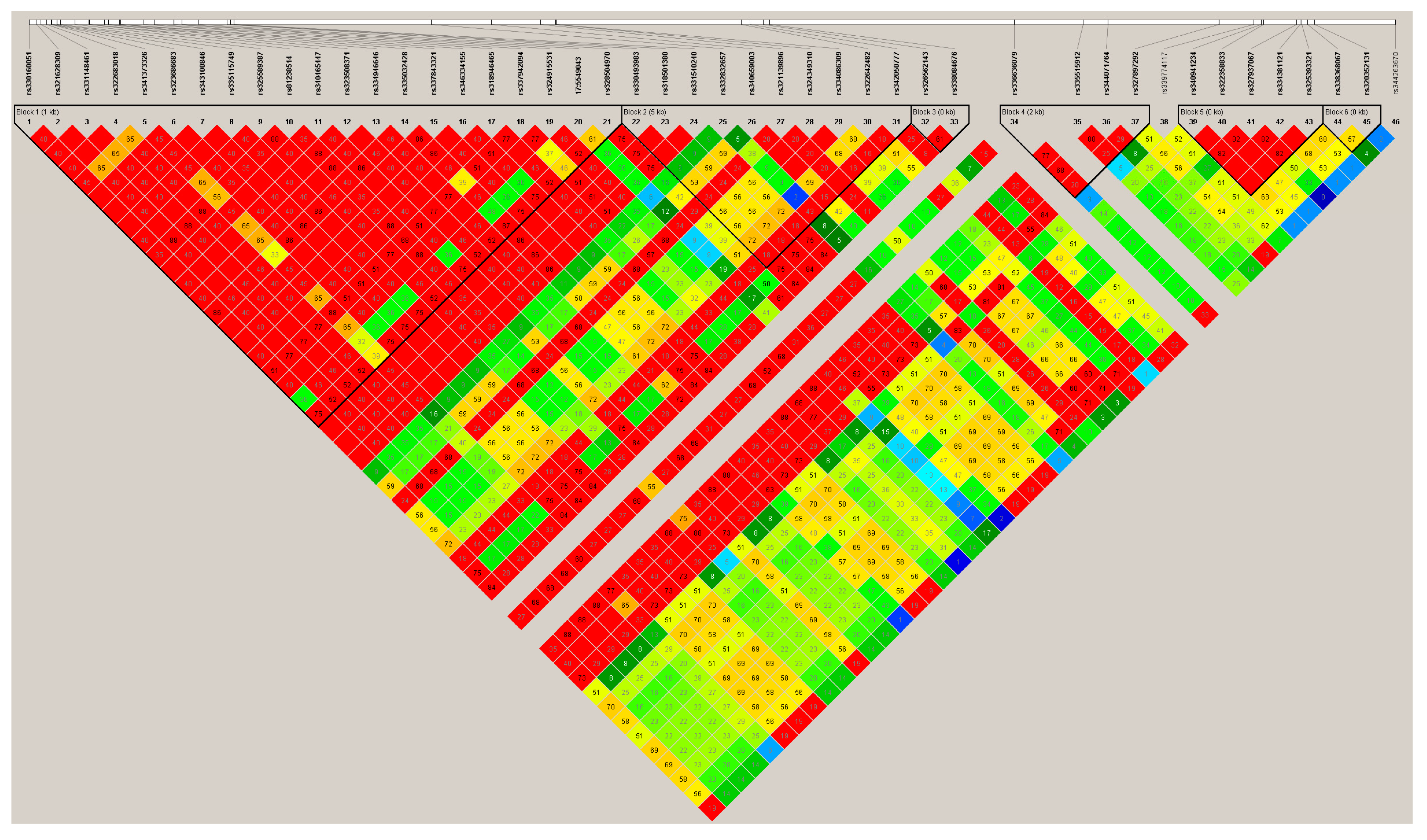
3.2. Variant Calling and χ2 Test Analyses
3.3. FGL1 Mutations Identified Using Variant Calling Analysis for 16 ZW Pigs
3.4. GLM Analysis between Złotnicka White Phenotype and FGL1 Variants
3.5. FGL1 Frequency in Pigs Active in Polish Breeding
4. Discussion
4.1. Pig Potential as an Animal Model in Fat Deposition Research
4.2. FGL1 Variants as a Potential Selective Marker to Improve Fat Content in Pigs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bakhtiarizadeh, M.R.; Salehi, A.; Alamouti, A.A.; Abdollahi-Arpanahi, R.; Salami, S.A. Deep transcriptome analysis using RNA-Seq suggests novel insights into molecular aspects of fat-tail metabolism in sheep. Sci. Rep. 2019, 9, 9203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, K.; Wang, K.; Ao, H.; Chen, S.; Tan, Z.; Wang, Y.; Xitong, Z.; Yang, T.; Zhang, F.; Liu, Y.; et al. Comparative adipose transcriptome analysis digs out genes related to fat deposition in two pig breeds. Sci. Rep. 2019, 9, 12925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rask-Andersen, M.; Karlsson, T.; Ek, W.E.; Johansson, Å. Genome-wide association study of body fat distribution identifies adiposity loci and sex-specific genetic effects. Nat. Commun. 2019, 10, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piórkowska, K.; Żukowski, K.; Ropka-Molik, K.; Tyra, M. Detection of genetic variants between different Polish Landrace and Puławska pigs by means of RNA-seq analysis. Anim. Genet. 2018, 49, 215–225. [Google Scholar] [CrossRef]
- Xing, K.; Zhu, F.; Zhai, L.; Chen, S.; Tan, Z.; Sun, Y.; Hou, Z.; Wang, C. Identification of genes for controlling swine adipose deposition by integrating transcriptome, whole-genome resequencing, and quantitative trait loci data. Sci. Rep. 2016, 6, 23219. [Google Scholar] [CrossRef] [Green Version]
- Piskol, R.; Ramaswami, G.; Li, J.B. Reliable identification of genomic variants from RNA-seq data. Am. J. Hum. Genet. 2013, 93, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Qiao, H.X.; Zhou, Y.T.; Hong, L.; Chen, J.H. Fibrinogen-like-protein 1 promotes the invasion and metastasis of gastric cancer and is associated with poor prognosis. Mol. Med. Rep. 2018, 18, 1465–1472. [Google Scholar] [CrossRef] [Green Version]
- Hara, H.; Yoshimura, H.; Uchida, S.; Toyoda, Y.; Aoki, M.; Sakai, Y.; Morimoto, S.; Shiokawa, K. Molecular cloning and functional expression analysis of a cDNA for human hepassocin, a liver-specific protein with hepatocyte mitogenic activity. Biochim. Biophys. Acta-Gene Struct. Expr. 2001, 1520, 45–53. [Google Scholar] [CrossRef]
- Demchev, V.; Malana, G.; Vangala, D.; Stoll, J.; Desai, A.; Kang, H.W.; Li, Y.; Nayeb-Hashemi, H.; Niepel, M.; Cohen, D.E.; et al. Targeted Deletion of Fibrinogen Like Protein 1 Reveals a Novel Role in Energy Substrate Utilization. PLoS ONE 2013, 8, e58084. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sanmamed, M.F.; Datar, I.; Su, T.T.; Ji, L.; Sun, J.; Chen, L.; Chen, Y.; Zhu, G.; Yin, W.; et al. Fibrinogen-like Protein 1 Is a Major Immune Inhibitory Ligand of LAG-3. Cell 2019, 176, 334–347.e12. [Google Scholar] [CrossRef] [Green Version]
- Sykes, M.; Sachs, D.H. Transplanting organs from pigs to humans. Sci. Immunol. 2019, 4, 41. [Google Scholar] [CrossRef]
- Houpt, K.A.; Houpt, T.R.; Pond, W.G. The pig as a model for the study of obesity and of control of food intake: A review. Yale J. Biol. Med. 1979, 52, 307–329. [Google Scholar]
- Grzeskowiak, E.; Borys, A.; Borzuta, K.; Buczynski, J.T.; Lisiak, D. Slaughter value, meat quality and backfat fatty acid profile in Zlotnicka White and Zlotnicka Spotted fatteners. Anim. Sci. Pap. Rep. 2009, 27, 115–125. [Google Scholar]
- Szyndler-Nędza, M.; Świątkiewicz, M.; Migdał, Ł.; Migdał, W. The Quality and Health-Promoting Value of Meat from Pigs of the Native Breed as the Effect of Extensive Feeding with Acorns. Animals 2021, 11, 789. [Google Scholar] [CrossRef]
- Różycki, M.; Tyra, M. Rules at evaluating the pigs in Pig Slaughter Testing Station. State of pig breeding and pig evaluation results. Inst. Zootech. Kraków 2010, 92–117. [Google Scholar]
- Tyra, M.; Żak, G. Analysis of the possibility of improving the indicators of pork quality through selection with particular consideration of intramuscular fat (imf) conntenttent*. Ann. Anim. Sci. 2013, 13, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Piórkowska, K.; Żukowski, K.; Ropka-Molik, K.; Tyra, M. New long-non coding RNAs related to fat deposition based on pig model. Ann. Anim. Sci. 2022. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Depristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; Del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–501. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Škrlep, M.; Kavar, T.; Čandek-Potokar, M. Comparison of PRKAG3 and RYR1 gene effect on carcass traits and meat quality in Slovenian commercial pigs. Czech J. Anim. Sci. 2010, 55, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Collado-Torres, L.; Nellore, A.; Frazee, A.C.; Wilks, C.; Love, M.I.; Langmead, B.; Irizarry, R.A.; Leek, J.T.; Jaffe, A.E. Flexible expressed region analysis for RNA-seq with derfinder. Nucleic Acids Res. 2017, 45, e9. [Google Scholar] [CrossRef]
- Frayn, K.N.; Arner, P.; Yki-Järvinen, H. Fatty acid metabolism in adipose tissue, muscle and liver in health and disease. Essays Biochem. 2006, 42, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Pillon, N.J.; Loos, R.J.F.; Marshall, S.M.; Zierath, J.R. Metabolic consequences of obesity and type 2 diabetes: Balancing genes and environment for personalized care. Cell 2021, 184, 1530–1544. [Google Scholar] [CrossRef]
- Cohen, J.B. Hypertension in Obesity and the Impact of Weight Loss. Curr. Cardiol. Rep. 2017, 19, 98. [Google Scholar] [CrossRef]
- Arnold, M.; Leitzmann, M.; Freisling, H.; Bray, F.; Romieu, I.; Renehan, A.; Soerjomataram, I. Obesity and cancer: An update of the global impact. Cancer Epidemiol. 2016, 41, 8–15. [Google Scholar] [CrossRef]
- Abdelaal, M.; le Roux, C.W.; Docherty, N.G. Morbidity and mortality associated with obesity. Ann. Transl. Med. 2017, 5, 161. [Google Scholar] [CrossRef] [Green Version]
- Jo, M.G.; Kim, M.W.; Jo, M.H.; bin Abid, N.; Kim, M.O. Adiponectin homolog osmotin, a potential anti-obesity compound, suppresses abdominal fat accumulation in C57BL/6 mice on high-fat diet and in 3T3-L1 adipocytes. Int. J. Obes. 2019, 43, 2422–2433. [Google Scholar] [CrossRef]
- Wang, Y.; Hollis-Hansen, K.; Ren, X.; Qiu, Y.; Qu, W. Do environmental pollutants increase obesity risk in humans? Obes. Rev. 2016, 17, 1179–1197. [Google Scholar] [CrossRef]
- Tuso, P.; Beattie, S. Nutrition reconciliation and nutrition prophylaxis: Toward total health. Perm. J. 2015, 19, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Oster, M.; Reyer, H.; Gerlinger, C.; Trakooljul, N.; Siengdee, P.; Keiler, J.; Ponsuksili, S.; Wolf, P.; Wimmers, K. mRNA Profiles of Porcine Parathyroid Glands Following Variable Phosphorus Supplies throughout Fetal and Postnatal Life. Biomedicines 2021, 9, 454. [Google Scholar] [CrossRef]
- Nielsen, K.L.; Hartvigsen, M.L.; Hedemann, M.S.; Lræke, H.N.; Hermansen, K.; Bach Knudsen, K.E. Similar metabolic responses in pigs and humans to breads with different contents and compositions of dietary fibers: A metabolomics study. Am. J. Clin. Nutr. 2014, 99, 941–949. [Google Scholar] [CrossRef]
- Stachowiak, M.; Szczerbal, I.; Switonski, M. Genetics of Adiposity in Large Animal Models for Human Obesity—Studies on Pigs and Dogs. In Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2016; Volume 140, pp. 233–270. ISBN 9780128046159. [Google Scholar]
- Kim, K.S.; Larsen, N.; Short, T.; Plastow, G.; Rothschild, M.F. A missense variant of the porcine melanocortin-4 receptor (MC4R) gene is associated with fatness, growth, and feed intake traits. Mamm. Genome 2000, 11, 131–135. [Google Scholar] [CrossRef]
- Piórkowska, K.; Tyra, M.; Rogoz, M.; Ropka-Molik, K.; Oczkowicz, M.; Rózycki, M. Association of the melanocortin-4 receptor (MC4R) with feed intake, growth, fatness and carcass composition in pigs raised in Poland. Meat Sci. 2010, 85, 297–301. [Google Scholar] [CrossRef]
- Chagnon, Y.C.; Rankinen, T.; Snyder, E.E.; Weisnagel, S.J.; Pérusse, L.; Bouchard, C. The human obesity gene map: The 2002 update. Obes. Res. 2003, 11, 313–367. [Google Scholar] [CrossRef] [Green Version]
- Sayeed, A.; Dalvano, B.E.; Kaplan, D.E.; Viswanathan, U.; Kulp, J.; Janneh, A.H.; Hwang, L.Y.; Ertel, A.; Doria, C.; Block, T. Research Paper Profiling the circulating mRNA transcriptome in human liver disease. Oncotarget 2020, 11, 2226–2242. [Google Scholar] [CrossRef]
- Li, P.; Chen, X.; Su, L.; Li, C.; Zhi, Q.; Yu, B.; Sheng, H.; Wang, J.; Feng, R.; Cai, Q.; et al. Epigenetic Silencing of miR-338-3p Contributes to Tumorigenicity in Gastric Cancer by Targeting SSX2IP. PLoS ONE 2013, 8, e66782. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhao, L.; Fang, Q.; Sun, J.; Zhang, S.; Zhan, C.; Liu, S.; Zhang, Y. MiR-338-3p inhibits hepatocarcinoma cells and sensitizes these cells to sorafenib by targeting hypoxia-induced factor 1. PLoS ONE 2014, 9, e115565. [Google Scholar] [CrossRef] [Green Version]
- Tyra, M.; Zak, G. Analysis of relationships between fattening and slaughter performance of pigs and the level of intramuscular fat (IMF) in longissimus dorsi muscle. Ann. Anim. Sci. 2012, 12, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Van Laere, A.S.; Nguyen, M.; Braunschweig, M.; Nezer, C.; Collette, C.; Moreau, L.; Archibald, A.L.; Haley, C.S.; Buys, N.; Tally, M.; et al. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature 2003, 425, 832–836. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Traits | HFD (n = 8) | LFD (n = 8) | All Pigs (n = 72) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | p-Value | Group Diff * | Mean | SD | |||
| Daily gain (g) | 641 | a | 34.5 | 718 | b | 84.2 | 0.03 | 11% | 700 | 106 |
| Backfat thickness (cm) | 2.09 | A | 0.31 | 1.53 | B | 0.36 | 0.004 | 27% | 1.90 | 0.39 |
| Peritoneal fat (kg) | 0.72 | A | 0.11 | 0.46 | B | 0.06 | 0.0001 | 36% | 0.61 | 0.16 |
| Backfat thickness in the K1 point (cm) | 2.23 | A | 0.37 | 1.44 | B | 0.20 | 0.0002 | 35% | 2.00 | 0.52 |
| Ham fat mass with skin (kg) | 2.49 | A | 0.21 | 1.80 | B | 0.23 | 1.86 × 10−5 | 28% | 2.15 | 0.37 |
| Loin fat mass with skin (kg) | 2.37 | A | 0.17 | 1.58 | B | 0.28 | 2.06 × 10−5 | 33% | 2.03 | 0.48 |
| Fat over shoulder thickness (cm) | 2.85 | A | 0.27 | 2.11 | B | 0.31 | 0.0002 | 26% | 2.75 | 0.61 |
| Lumbar fat I thickness (cm) | 2.43 | A | 0.26 | 1.44 | B | 0.32 | 1.096 × 10−5 | 41% | 2.16 | 0.55 |
| Lumbar fat II thickness (cm) | 2.20 | A | 0.47 | 1.26 | B | 0.21 | 0.0004 | 43% | 1.95 | 0.54 |
| Lumbar fat III thickness (cm) | 2.74 | A | 0.63 | 1.80 | B | 0.29 | 0.003 | 34% | 2.40 | 0.59 |
| Average backfat thickness (cm) | 2.46 | A | 0.27 | 1.60 | B | 0.17 | 8.63 × 10−6 | 35% | 2.27 | 0.45 |
| FGL1 expression level | 3799.6 | 932.61 | 3731.6 | 1125.4 | 0.45 | |||||
| Traits | FGL1 Genotype | ||
|---|---|---|---|
| GG TT | AG CT | AA CC | |
| Loin weight (kg) | 4.87 ± 0.40 A | 4.38 ± 0.30 B | 4.39 ± 0.30 |
| Ham weight (kg) | 8.06 ± 0.30 a | 7.48 ± 0.30 b | 7.07 ± 0.7 b |
| Feet mass (kg) | 1.02 ± 0.05 Aa | 0.95 ± 0.01 b | 0.89 ± 0.07 Bc |
| Knuckle fat with skin (kg) | 0.21 ± 0.01 A | 0.25 ± 0.02 B | 0.24 ± 0.01 AB |
| Loin eye height (cm) | 6.80 ± 0.16 A | 6.22 ± 0.27 B | 6.45 ± 0.32 AB |
| Loin eye area (cm2) | 47.1 ± 2.75 a | 43.5 ± 3.45 a | 42.4 ± 3.48 ab |
| Meat percentage in primary cuts (kg) | 64.2 ± 1.08 A | 58.9 ± 1.21 Ba | 58.3 ± 2.54 b |
| Meat percentage % | 56.0 ± 1.05 A | 50.5 ± 1.21 Ba | 50.2 ± 2.51 b |
| Peritoneal fat (kg) | 0.46 ± 1.99 A | 0.70 ± 2.88 B | 0.76 ± 4.89 AB |
| Loin fat with skin (kg) | 1.58 ± 0.20 Aa | 2.39 ± 0.20 B | 2.28 ± 0.10 Bb |
| Ham fat with skin (kg) | 1.80 ± 0.30 A | 2.53 ± 0.01 B | 2.38 ± 0.02 B |
| Backfat over shoulder (cm) | 2.11 ± 0.31 A | 2.83 ± 0.10 B | 2.90 ± 0.31 B |
| Backfat over back (cm) | 1.53 ± 0.36 A | 2.10 ± 0.36 B | 2.05 ± 0.05 B |
| Backfat over lumbar I | 1.44 ± 0.32 Aa | 2.43 ± 0.28 B | 2.40 ± 0.20 b |
| Backfat over lumbar II | 1.26 ± 0.21 A | 2.18 ± 0.52 B | 2.25 ± 0.25 B |
| Backfat over lumbar III | 1.80 ± 0.71 Aa | 2.78 ± 0.20 B | 2.60 ± 0.29 b |
| Average backfat thickness (cm) | 1.63 ± 0.18 A | 2.47 ± 0.31 B | 2.44 ± 0.10 B |
| Backfat in the point C1 | 1.44 ± 0.20 Aa | 2.25 ± 0.15 B | 2.15 ± 0.41 Bb |
| Backfat in the point K1 | 1.44 ± 0.20 Aa | 2.25 ± 0.15 B | 2.15 ± 0.41 Bb |
| Traits | FGL1 Genotype | GLM Significance | Additive Effect | Dominance Effect | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AA (n = 26) | AG (n = 16) | GG (n = 30) | p-Value | X2P | FGL1 | A → G | Het → Hom | ||||
| LSM | SE | LSM | SE | LSM | SE | ||||||
| pHham24 | 5.57 AB | 0.03 | 5.64 A | 0.03 | 5.53 B | 0.02 | 0.0080 | x | ** | ns | −0.04 * |
| pHloin24 | 5.59 A | 0.02 | 5.55 AB | 0.03 | 5.46 B | 0.02 | 0.0006 | x | *** | −0.06 *** | ns |
| IMF | 1.64 a | 0.10 | 1.32 b | 0.09 | 1.49 ab | 0.06 | 0.0462 | x | * | ns | +0.13 * |
| Carcass yield (kg) | 74.8 a | 0.18 | 74.6 ab | 0.22 | 74.1 b | 0.16 | 0.0262 | *** | * | ns | ns |
| Average backfat thickness (cm) | 2.61 a | 0.14 | 2.28 ab | 0.13 | 2.12 b | 0.10 | 0.0234 | ns | * | −0.24 ** | ns |
| Loin eye height (cm) | 5.62 A | 0.19 | 6.08 AB | 0.18 | 6.40 B | 0.13 | 0.0046 | ns | ** | +0.41 ** | ns |
| Loin eye area (cm2) | 40.1 a | 1.53 | 44.3 ab | 1.47 | 45.3 b | 1.10 | 0.0236 | ** | * | +2.90 ** | ns |
| Meat percentage (%) | 50.9 | 1.13 | 52.3 | 1.08 | 53.5 | 0.80 | 0.1491 | ns | ns | +1.33 * | ns |
| Primary cut (kg) | 18.8 | 0.43 | 19.3 | 0.41 | 19.8 | 0.31 | 0.1919 | ns | ns | +0.60 * | ns |
| Loin fat with skin (kg) | 2.32 A | 0.13 | 2.17 ABa | 0.13 | 1.76 Bb | 0.09 | AB 0.0033 ab 0.0344 | *** | ** | −0.25 * | ns |
| Backfat over back (cm) | 2.35 | 0.16 | 1.90 | 0.15 | 1.87 | 0.12 | 0.0557 | ns | * | −0.23 * | ns |
| Backfat over lumbar I (cm) | 2.61 A | 0.17 | 2.20 AB | 0.16 | 1.97 B | 0.12 | 0.0087 | ns | ** | −0.32 ** | ns |
| Backfat over lumbar II (cm) | 2.33 a | 0.16 | 1.94 ab | 0.15 | 1.76 b | 0.11 | 0.0136 | ns | ** | −0.28 ** | ns |
| Backfat over lumbar III (cm) | 2.72 | 0.18 | 2.52 | 0.17 | 2.24 | 0.13 | 0.0939 | ns | * | −0.24 * | ns |
| Daily gain (0–180 days) kg | 530 | 16 | 521 | 15 | 493 | 11 | 0.1529 | ns | ns | −18 * | ns |
| Slaughter age (day) | 187 | 6.4 | 192 | 6.2 | 204 | 4.9 | 0.0831 | x | * | +9 * | ns |
| Traits | FGL1 Genotype | GLM Significance | Additive Effect | Dominance Effect | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| TCA/TCA (n = 31) | TCA/A (n = 32) | A/A (n = 9) | p-Value | X2P | FGL1 | Ins → Del | Het → Hom | ||||
| LSM | SE | LSM | SE | LSM | SE | ||||||
| IMF | 1.49 | 0.06 | 1.40 | 0.06 | 1.90 | 0.21 | 0.0760 | x | * | 0.21 * | 0.15 * |
| pHham24 | 5.53 a | 0.02 | 5.62 b | 0.02 | 5.58 ab | 0.06 | 0.0354 | x | ** | ns | ns |
| pHloin24 | 5.47 a | 0.02 | 5.56 b | 0.02 | 5.60 ab | 0.04 | 0.0103 | x | ** | 0.06 * | ns |
| Carcass yield (kg) | 74.08 A | 0.14 | 74.6 B | 0.16 | 75.4 AB | 0.35 | 0.0039 | *** | ** | ns | ns |
| Average backfat thickness (cm) | 2.11 A | 0.10 | 2.56 B | 0.10 | 1.99 AB | 0.24 | 0.0093 | ns | ** | ns | −0.26 ** |
| Loin eye area (cm2) | 45.5 * | 1.10 | 41.6 | 1.20 | 43.7 | 2.70 | 0.0506 | ** | * | ns | ns |
| Loin eye height (cm) | 6.42 a | 0.14 | 5.81 b | 0.15 | 5.82 ab | 0.33 | 0.0107 | ns | ** | −0.32 * | ns |
| Meat percentage (%) | 53.6 a | 0.74 | 50.6 b | 0.80 | 55.8 ab | 1.81 | 0.0205 | ns | ** | ns | +2.04 ** |
| Primary cut (kg) | 19.8 a | 0.28 | 18.6 b | 0.31 | 20.7 ab | 0.69 | 0.0227 | *** | ** | ns | +0.70 * |
| Loin fat with skin (kg) | 1.76 A | 0.09 | 2.33 B | 0.10 | 1.88 AB | 0.22 | 0.0003 | *** | *** | ns | −0.28 ** |
| Loin mass without fat and skin (kg) | 4.62 a | 0.10 | 4.24 b | 0.10 | 4.82 ab | 0.24 | 0.0326 | *** | ** | ns | +0.22 * |
| Ham fat mass (kg) | 1.99 a | 0.08 | 2.30 b | 0.09 | 1.84 ab | 0.19 | 0.0271 | * | ** | ns | −0.21 ** |
| Ham mass (kg) | 7.75 a | 0.14 | 7.31 b | 0.15 | 8.16 ab | 0.26 | 0.0374 | *** | ** | ns | +0.28 * |
| Backfat over back (cm) | 1.85 | 0.11 | 2.23 | 0.12 | 1.72 | 0.3 | 0.0827 | ns | * | ns | −0.22 * |
| Backfat over lumbar I (cm) | 1.97 a | 0.10 | 2.50 b | 0.10 | 2.02 ab | 0.29 | 0.0102 | ns | * | ns | −0.25 ** |
| Backfat over lumbar II (cm) | 1.75 A | 0.11 | 2.25 B | 0.12 | 1.65 AB | 0.26 | 0.0078 | ns | ** | ns | −0.28 ** |
| Backfat over lumbar III (cm) | 2.23 A | 0.12 | 2.78 B | 0.13 | 2.03 AB | 0.29 | 0.0083 | ns | ** | ns | −0.33 ** |
| Backfat in the point C1 | 1.86 a | 0.12 | 2.38 b | 0.13 | 1.55 ab | 0.29 | 0.0130 | ns | ** | ns | −0.34 ** |
| Backfat in the point K1 | 1.85 a | 0.11 | 2.38 b | 0.13 | 1.60 ab | 0.28 | 0.0118 | ns | ** | ns | −0.32 ** |
| Daily gain | 729 | 20 | 662 | 22 | 714 | 50 | 0.0754 | ns | * | ns | +30 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piórkowska, K.; Żukowski, K.; Ropka-Molik, K.; Tyra, M. Variations in Fibrinogen-like 1 (FGL1) Gene Locus as a Genetic Marker Related to Fat Deposition Based on Pig Model and Liver RNA-Seq Data. Genes 2022, 13, 1419. https://doi.org/10.3390/genes13081419
Piórkowska K, Żukowski K, Ropka-Molik K, Tyra M. Variations in Fibrinogen-like 1 (FGL1) Gene Locus as a Genetic Marker Related to Fat Deposition Based on Pig Model and Liver RNA-Seq Data. Genes. 2022; 13(8):1419. https://doi.org/10.3390/genes13081419
Chicago/Turabian StylePiórkowska, Katarzyna, Kacper Żukowski, Katarzyna Ropka-Molik, and Mirosław Tyra. 2022. "Variations in Fibrinogen-like 1 (FGL1) Gene Locus as a Genetic Marker Related to Fat Deposition Based on Pig Model and Liver RNA-Seq Data" Genes 13, no. 8: 1419. https://doi.org/10.3390/genes13081419