
Alternative Splicing Isoforms of Porcine CREB Are Differentially Involved in Transcriptional Transactivation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
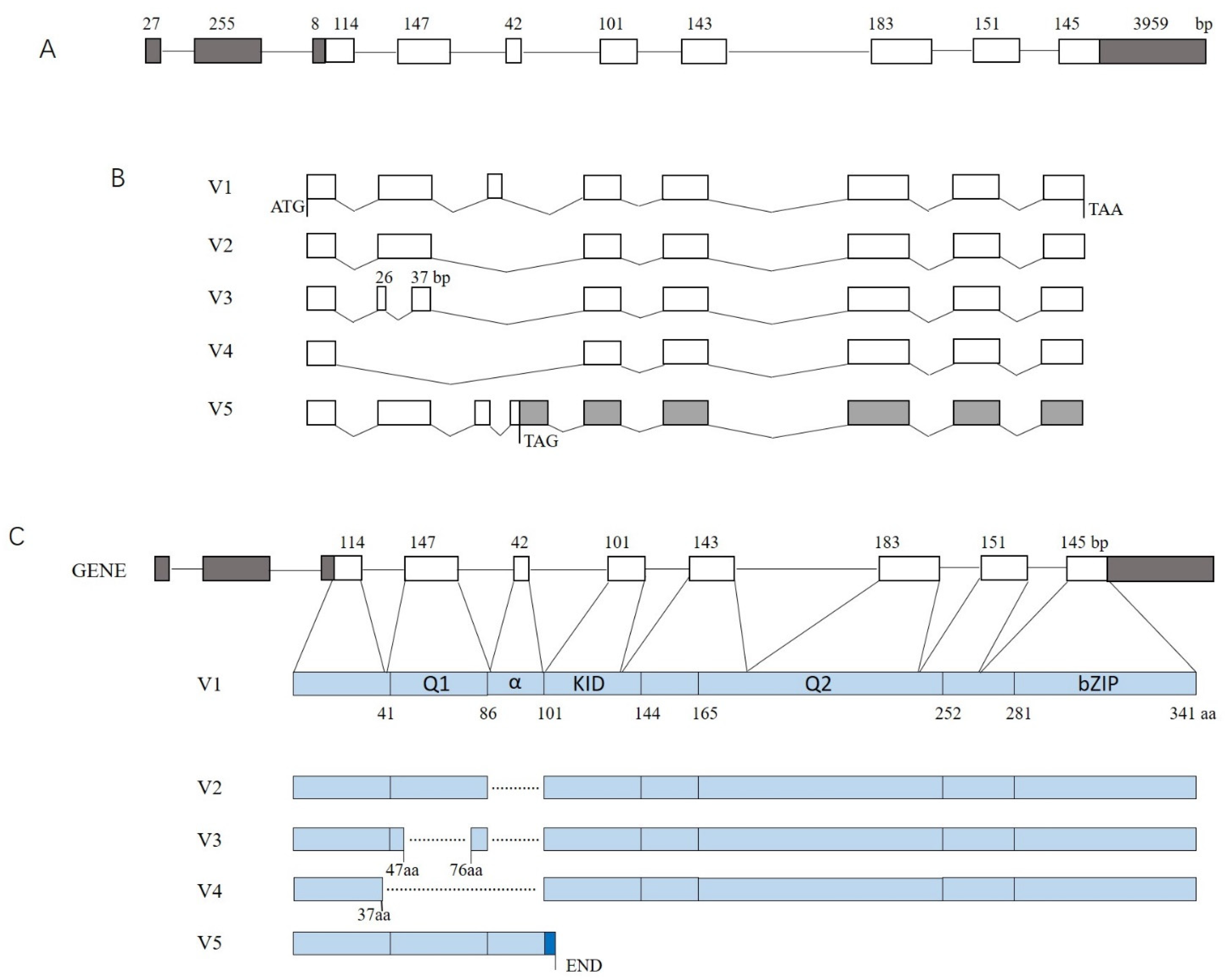
2.1. Identification of Alternative Splicing Transcripts
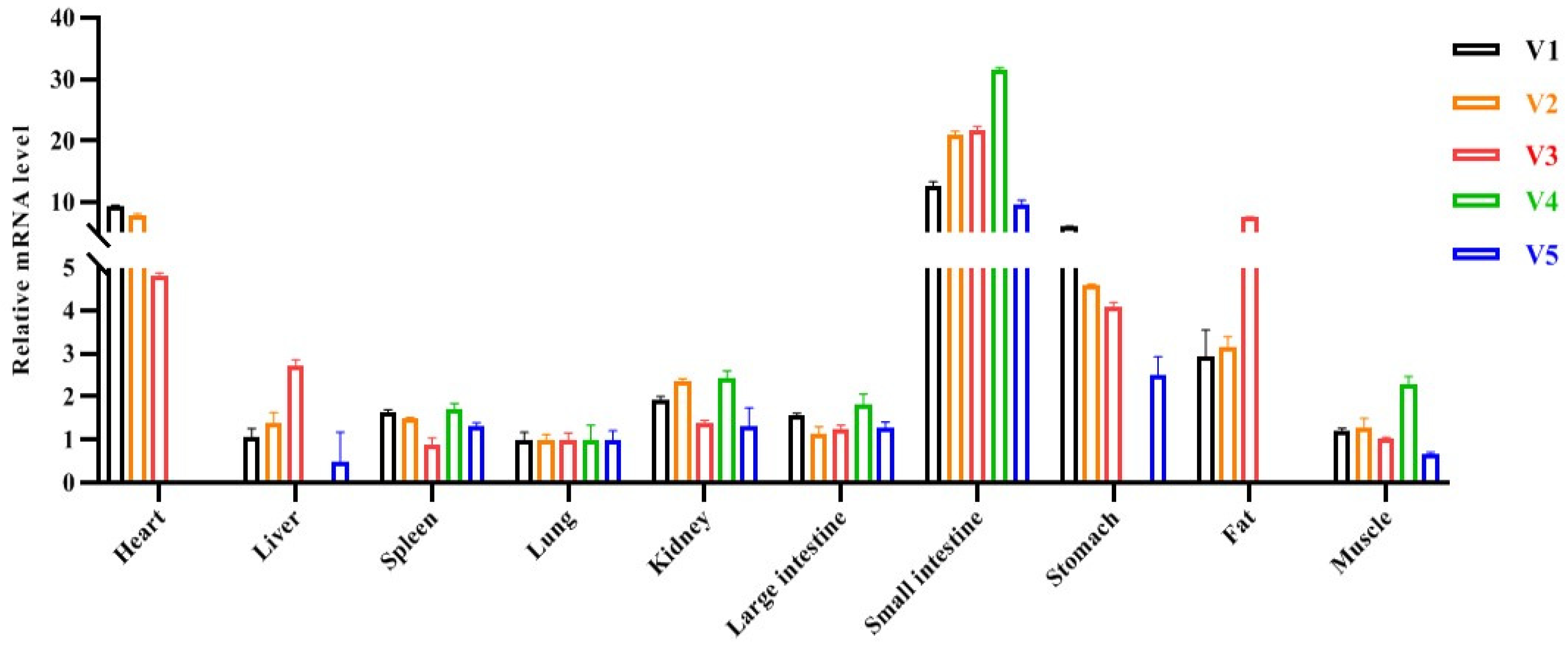
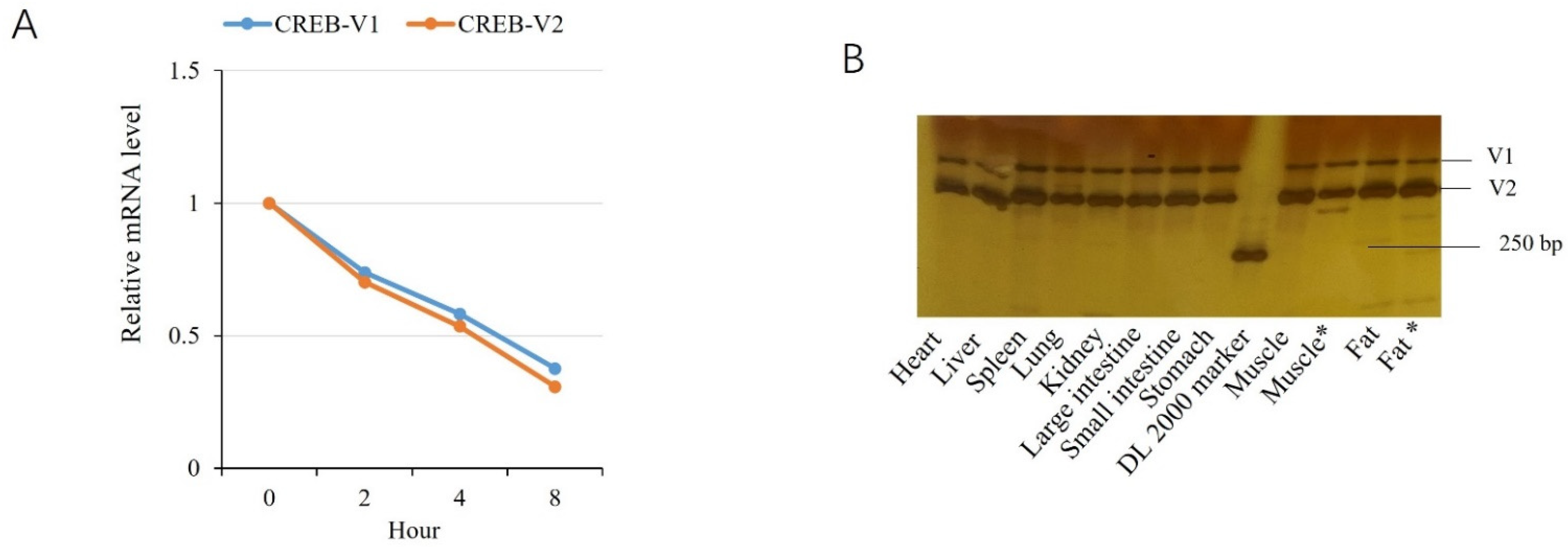
2.2. Tissue Expression Profile
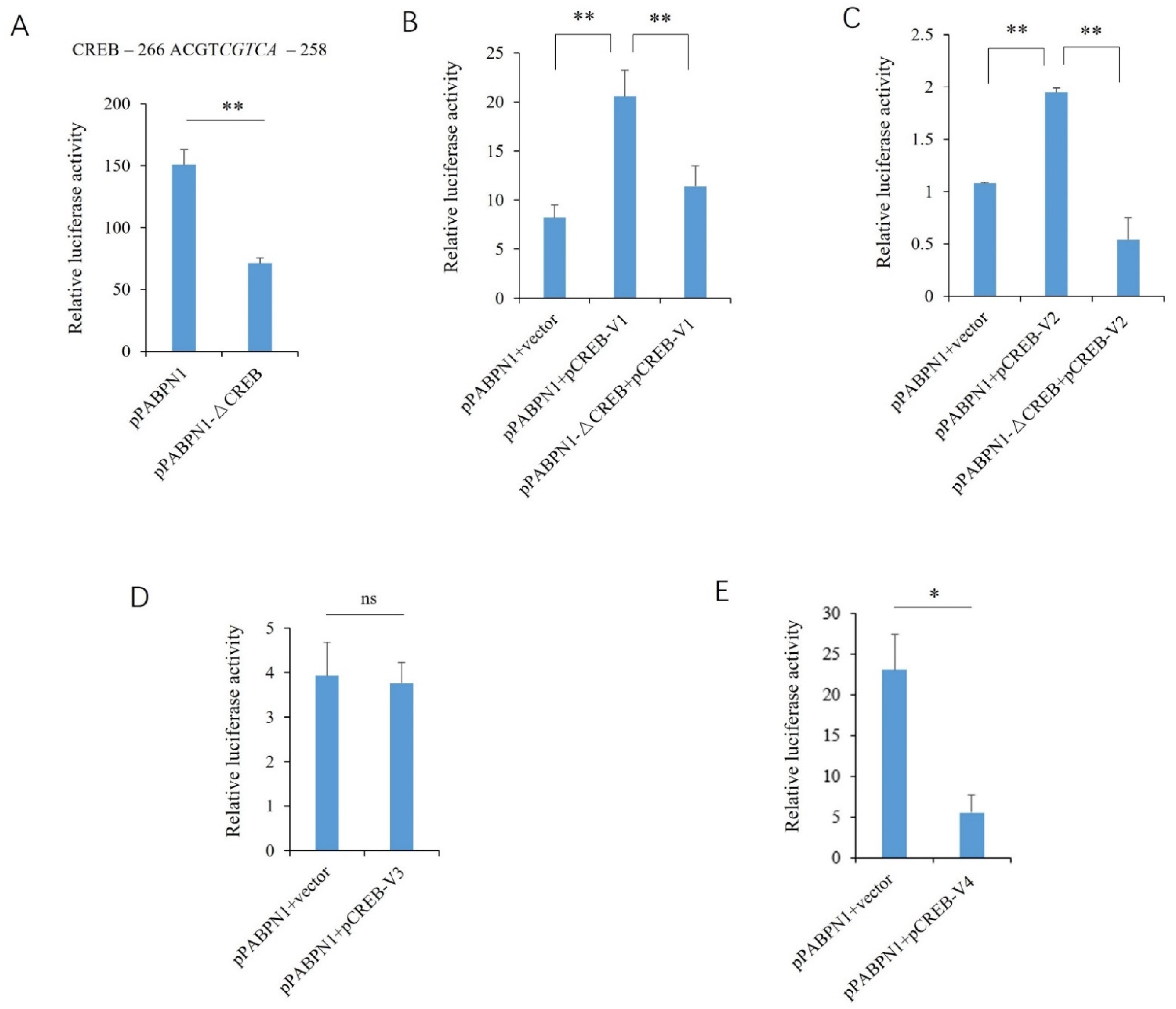
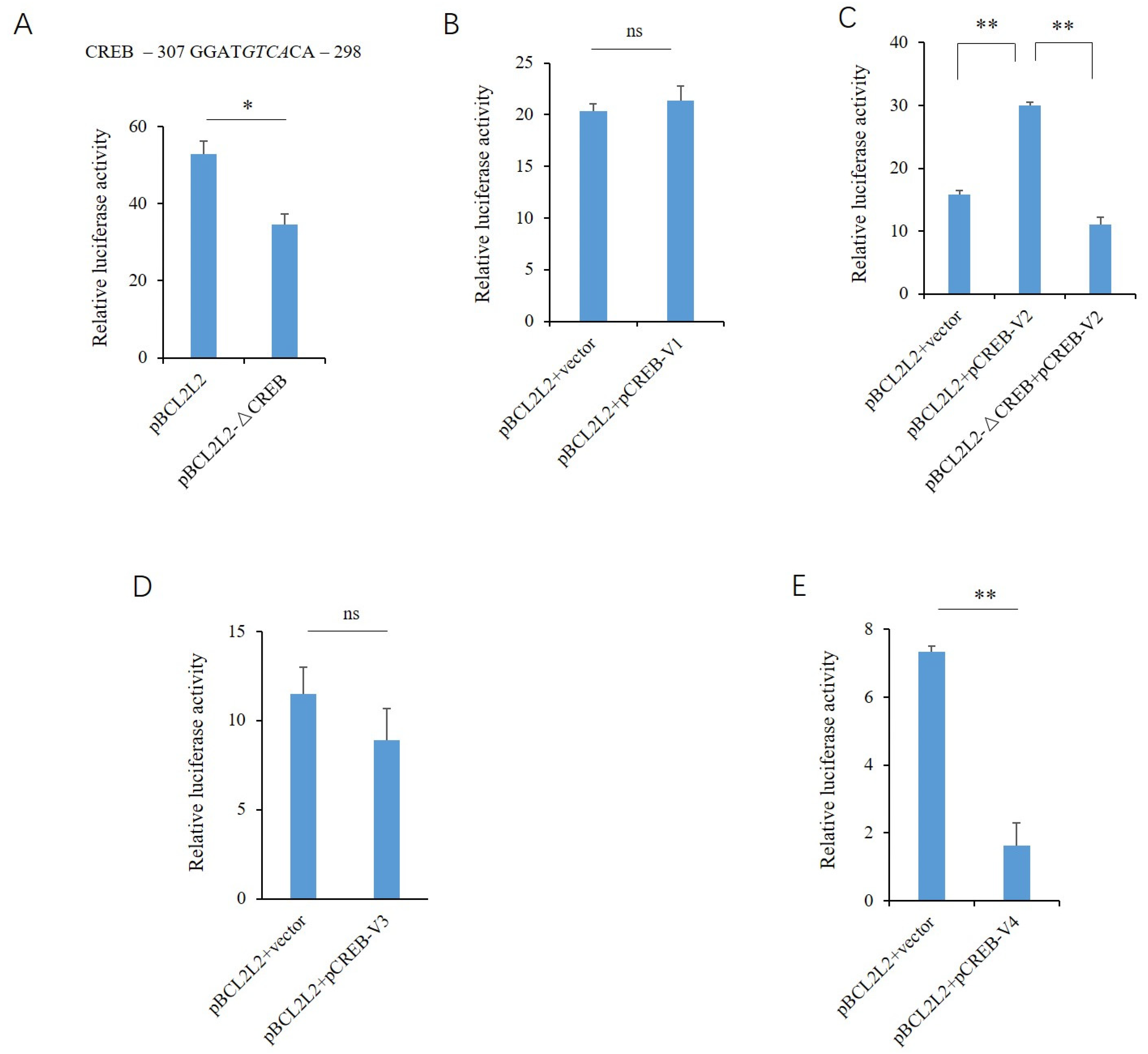
2.3. Transcriptional Regulatory Effects of the CREB Variants
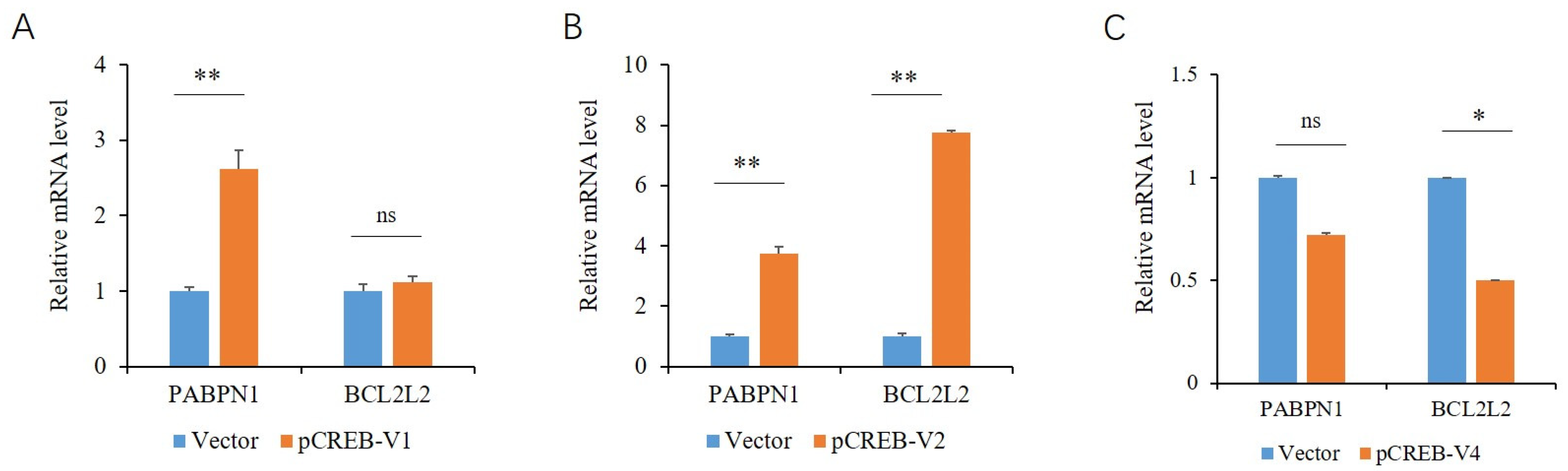
2.4. Effects of the CREB Variants on the Endogenous Expression of PABPN1 and BCL2L2
2.5. Comparison of Variants V1 and V2
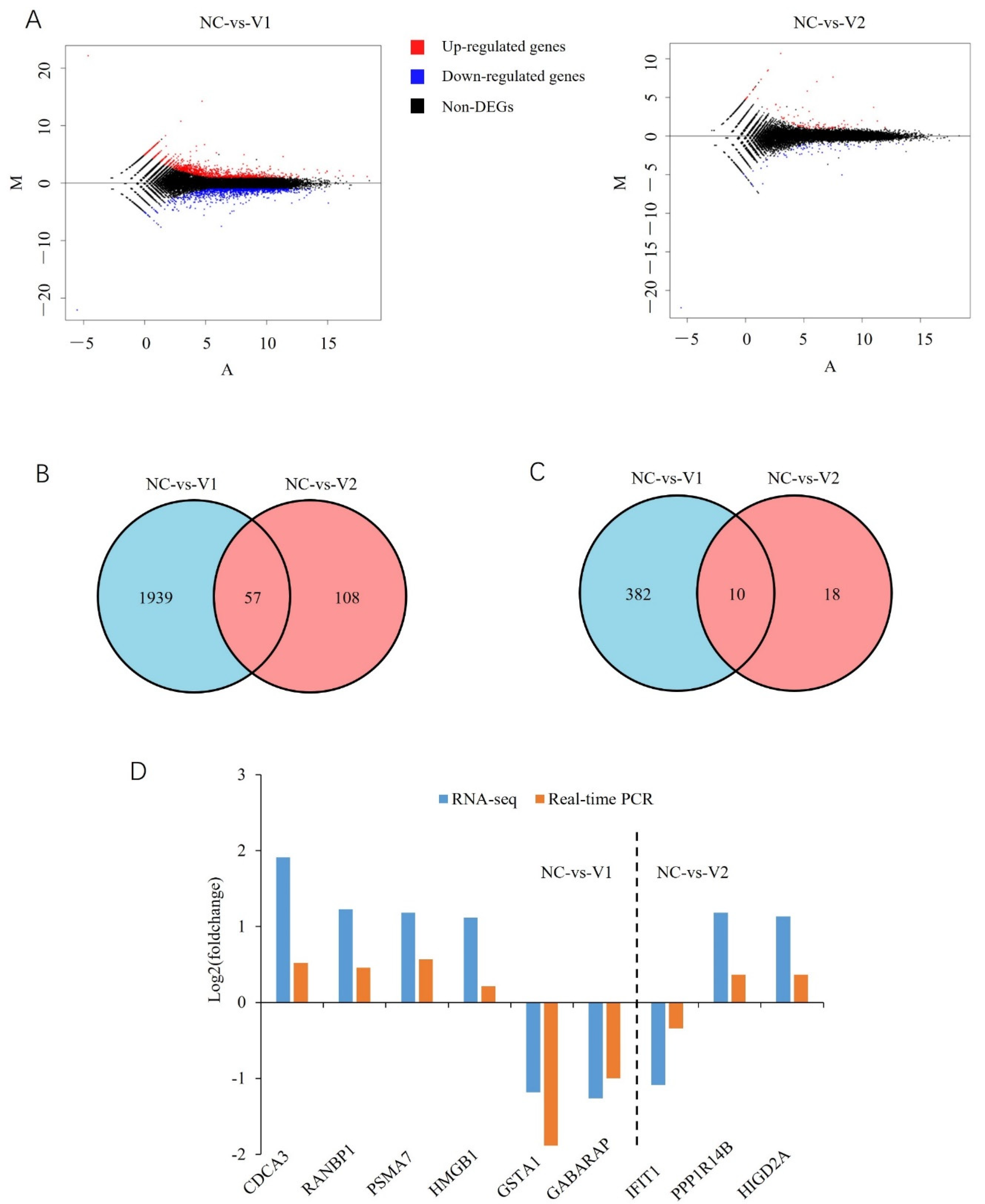
2.6. Overview of the RNA-Seq Data
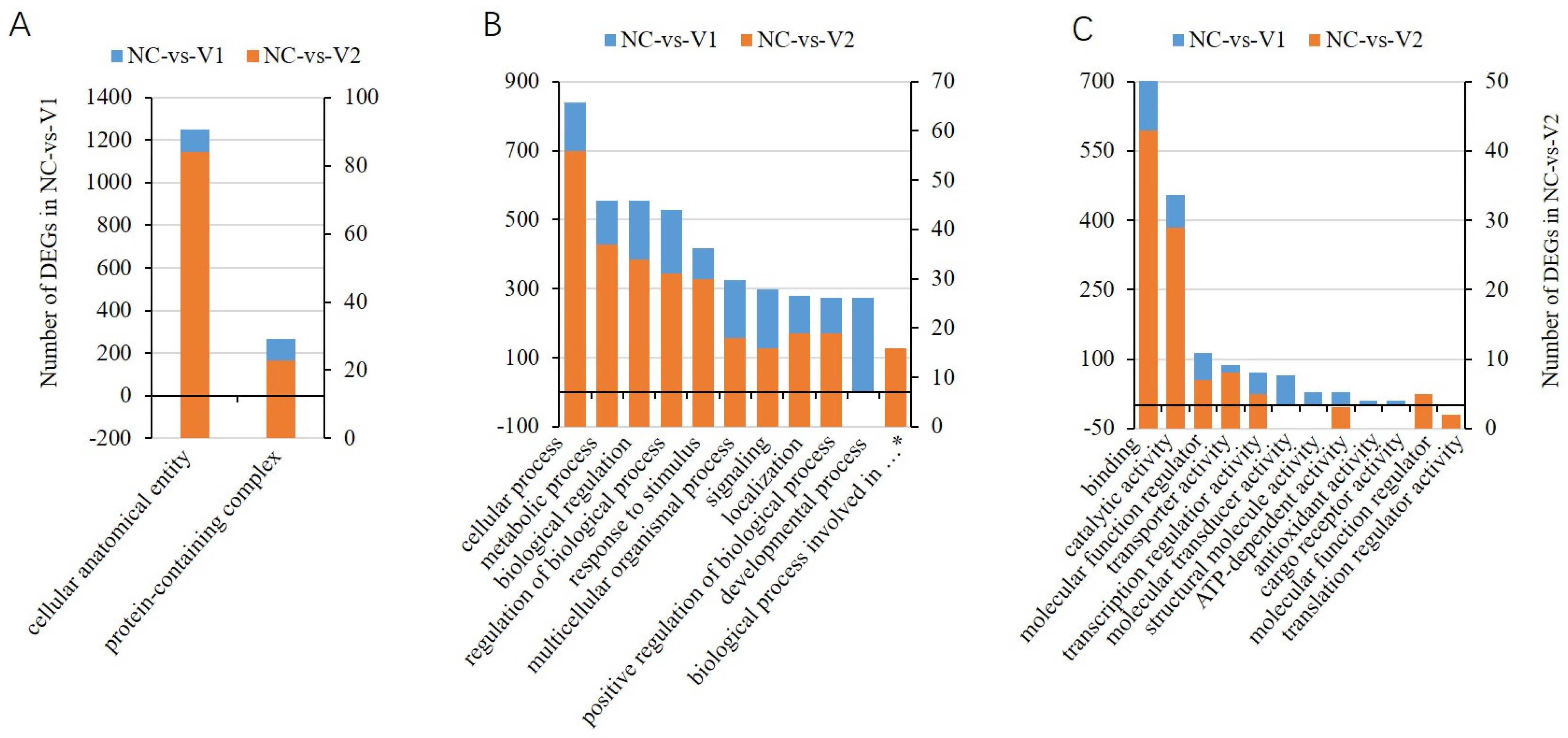
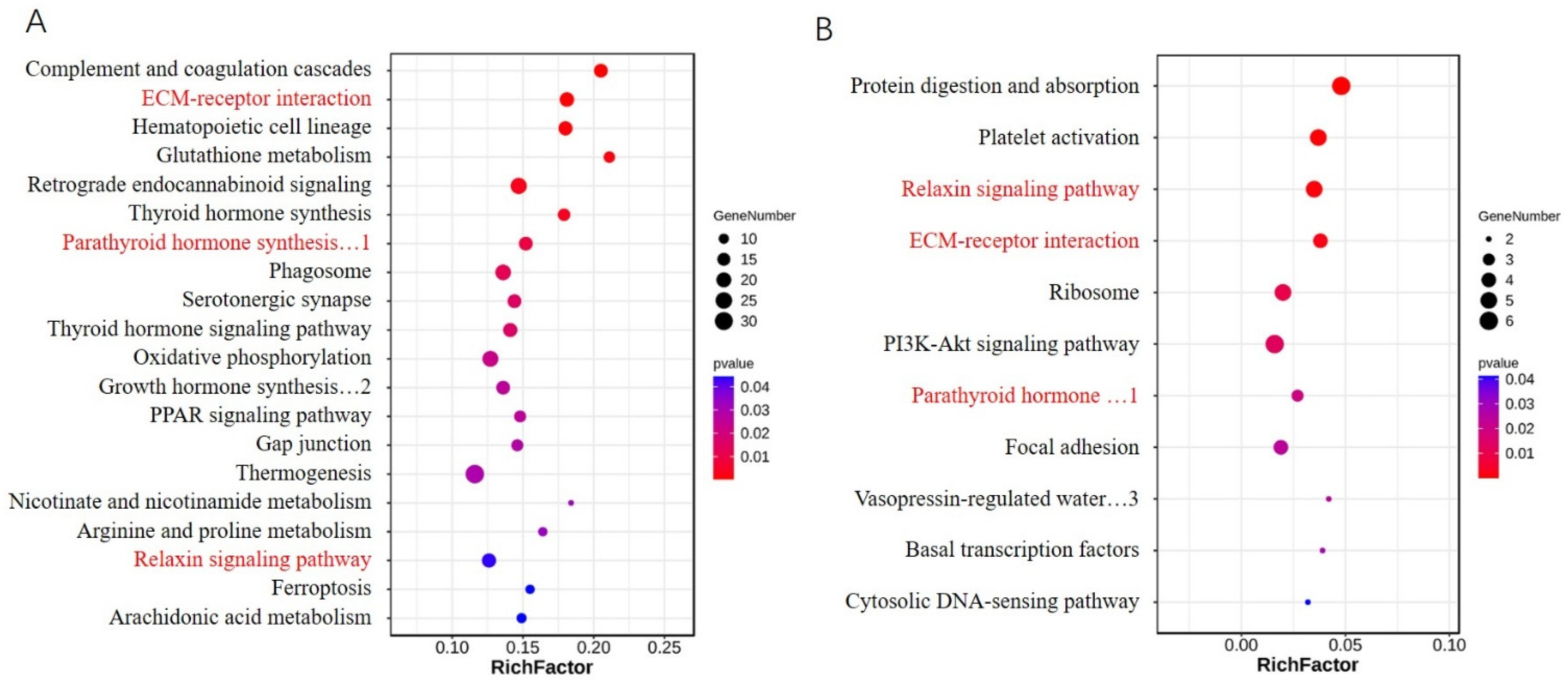
2.7. Functional Characterization of the Differentially Expressed Genes
3. Discussion
4. Materials and Methods
4.1. Animals, Tissues, and cDNA Synthesis
4.2. cDNA Amplification and Sequence Analysis
4.3. Plasmids
4.4. Dual-Luciferase Reporter Assay
4.5. Competitive RT-PCR
4.6. Library Preparation and Sequencing
4.7. RNA-Seq Data Analysis
4.8. Real-Time Quantitative PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; Odom, D.T.; Koo, S.H.; Conkright, M.D.; Canettieri, G.; Best, J.; Chen, H.; Jenner, R.; Herbolsheimer, E.; Jacobsen, E.; et al. Genome-wide analysis of cAMP-response element binding protein occupancy, phosphorylation, and target gene activation in human tissues. Proc. Natl. Acad. Sci. USA 2005, 102, 4459–4464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleckmann, S.C.; Blendy, J.A.; Rudolph, D.; Monaghan, A.P.; Schmid, W.; Schütz, G. Activating transcription factor 1 and CREB are important for cell survival during early mouse development. Mol. Cell Biol. 2002, 22, 1919–1925. [Google Scholar] [CrossRef] [Green Version]
- Mantamadiotis, T.; Lemberger, T.; Bleckmann, S.C.; Kern, H.; Kretz, O.; Martin Villalba, A.; Tronche, F.; Kellendonk, C.; Gau, D.; Kapfhammer, J.; et al. Disruption of CREB function in brain leads to neurodegeneration. Nat. Genet. 2002, 31, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dong, D.L.; Gao, J.H.; Wang, A.K.; Shao, Y.P. β-HB inhibits the apoptosis of high glucose-treated astrocytes via activation of CREB/BDNF axis. Brain Inj. 2021, 35, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Lian, G.; Wang, T.; Chen, W.; Zhuang, W.; Luo, L.; Wang, H.; Xie, L. Zinc-mediated activation of CREB pathway in proliferation of pulmonary artery smooth muscle cells in pulmonary hypertension. Cell Commun. Signal. 2021, 19, 103. [Google Scholar] [CrossRef]
- Dworkin, S.; Mantamadiotis, T. Targeting CREB signalling in neurogenesis. Exper. Opin. Ther. Targets. 2010, 14, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.B.; Xie, J.; Li, Y.T.; Yuan, Y.; Wang, Y.; Li, C.; Shi, Y.F. Knockdown of CERB expression inhibits proliferation and migration of glioma cells line U251. Bratisl. Lek. Listy 2019, 120, 309–315. [Google Scholar] [CrossRef]
- Montminy, M.R.; Sevarino, K.A.; Wagner, J.A.; Mandel, G.; Goodman, R.H. Identification of a cyclic-AMP-responsive element within the rat somatostatin gene. Proc. Natl. Acad. Sci. USA 1986, 83, 6682–6686. [Google Scholar] [CrossRef] [Green Version]
- Nichols, M.; Weih, F.; Schmid, W.; DeVack, C.; Kowenz-Leutz, E.; Luckow, B.; Boshart, M.; Schütz, G. Phosphorylation of CREB affects its binding to high and low affinity sites: Implications for cAMP induced gene transcription. EMBO J. 1992, 11, 3337–3346. [Google Scholar] [CrossRef]
- Craig, J.C.; Schumacher, M.A.; Mansoor, S.E.; Farrens, D.L.; Brennan, R.G.; Goodman, R.H. Consensus and variant cAMP-regulated enhancers have distinct CREB-binding properties. J. Bio. Chem. 2001, 276, 11719–11728. [Google Scholar] [CrossRef] [Green Version]
- Kovacevic-Grujicic, N.; Mojsin, M.; Popovic, J.; Petrovic, I.; Topalovic, V.; Stevanovic, M. Cyclic AMP response element binding (CREB) protein acts as a positive regulator of SOX3 gene expression in NT2/D1 cells. BMB Rep. 2014, 47, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, T.; Morohashi, K.; Takayama, K.; Honda, S.; Wada, T.; Handa, H.; Omura, T. Cooperative transcription activation between Ad1, a CRE-like element, and other elements in the CYP11B gene promoter. J. Biochem. 1992, 112, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Brenig, B.; Duan, Y.; Xing, Y.; Ding, N.; Huang, L.; Schütz, E. Porcine SOX9 Gene Expression Is Influenced by an 18 bp Indel in the 5′-Untranslated Region. PLoS ONE 2015, 10, e0139583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muchardt, C.; Li, C.; Kornuc, M.; Gaynor, R. CREB regulation of cellular cyclic AMP-responsive and adenovirus early promoters. J. Virol. 1990, 64, 4296–4305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fass, D.M.; Craig, J.C.; Impey, S.; Goodman, R.H. Cooperative mechanism of transcriptional activation by a cyclic AMP-response element modulator alpha mutant containing a motif for constitutive binding to CREB-binding protein. J. Biol. Chem. 2001, 276, 2992–2997. [Google Scholar] [CrossRef] [Green Version]
- Merrett, J.E.; Bo, T.; Psaltis, P.J.; Proud, C.G. Identification of DNA response elements regulating expression of CCAAT/enhancer-binding protein (C/EBP) β and δ and MAP kinase-interacting kinases during early adipogenesis. Adipocyte 2020, 9, 427–442. [Google Scholar] [CrossRef]
- Hai, T.; Curran, T. Cross-family dimerization of transcription factors Fos/Jun and ATF/CREB alters DNA binding specificity. Proc. Natl. Acad. Sci. USA 1991, 88, 3720–3724. [Google Scholar] [CrossRef] [Green Version]
- Glick, Y.; Orenstein, Y.; Chen, D.; Avrahami, D.; Zor, T.; Shamir, R.; Gerber, D. Integrated microfluidic approach for quantitative high-throughput measurements of transcription factor binding affinities. Nucleic Acids Res. 2016, 44, e51. [Google Scholar] [CrossRef] [Green Version]
- Ruppert, S.; Cole, T.J.; Boshart, M.; Schmid, E.; Schütz, G. Multiple mRNA isoforms of the transcription activator protein CREB: Generation by alternative splicing and specific expression in primary spermatocytes. EMBO J. 1992, 11, 1503–1512. [Google Scholar] [CrossRef]
- Daniel, P.B.; Habener, J.F. Cyclical alternative exon splicing of transcription factor cyclic adenosine monophosphate response element-binding protein (CREB) messenger ribonucleic acid during rat spermatogenesis. Endocrinology 1998, 139, 3721–3729. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, J.; Lu, L.; Yin, L.; Xu, M.; Wang, Y.; Zhou, Z.; Sha, J. Cloning and expression of a novel CREB mRNA splice variant in human testis. Reproduction 2004, 128, 775–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waeber, G.; Habener, J.F. Novel testis germ cell-specific transcript of the CREB gene contains an alternatively spliced exon with multiple in-frame stop codons. Endocrinology 1992, 131, 2010–2015. [Google Scholar] [CrossRef] [PubMed]
- Saura, C.A.; Cardinaux, J.R. Emerging Roles of CREB-Regulated Transcription Coactivators in Brain Physiology and Pathology. Trends Neurosci. 2017, 40, 720–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amidfar, M.; de Oliveira, J.; Kucharska, E.; Budni, J.; Kim, Y.K. The role of CREB and BDNF in neurobiology and treatment of Alzheimer’s disease. Life Sci. 2020, 257, 118020. [Google Scholar] [CrossRef]
- Berkowitz, L.A.; Gilman, M.Z. Two distinct forms of active transcription factor CREB (cAMP response element binding protein). Proc. Natl. Acad. Sci. USA 1990, 87, 5258–5262. [Google Scholar] [CrossRef] [Green Version]
- Steven, A.; Friedrich, M.; Jank, P.; Heimer, N.; Budczies, J.; Denkert, C.; Seliger, B. What turns CREB on? And off? And why does it matter? Cell Mol. Life Sci. 2020, 77, 4049–4067. [Google Scholar] [CrossRef]
- Walker, W.H.; Girardet, C.; Habener, J.F. Alternative exon splicing controls a translational switch from activator to repressor isoforms of transcription factor CREB during spermatogenesis. J. Biol. Chem. 1996, 271, 20145–21050. [Google Scholar] [CrossRef]
- Walker, W.H.; Daniel, P.B.; Habener, J.F. Inducible cAMP early repressor ICER down-regulation of CREB gene expression in Sertoli cells. Mol. Cell. Endocrinol. 1998, 143, 167–178. [Google Scholar] [CrossRef]
- Zhang, H.; Kong, Q.; Wang, J.; Jiang, Y.; Hua, H. Complex roles of cAMP-PKA-CREB signaling in cancer. Exp. Hematol. Oncol. 2020, 9, 32. [Google Scholar] [CrossRef]
- Quinn, P.G. Distinct activation domains within cAMP response element-binding protein (CREB) mediate basal and cAMP-stimulated transcription. J. Biol. Chem. 1993, 268, 16999–17009. [Google Scholar] [CrossRef]
- Foulkes, N.S.; Sassone-Corsi, P. More is better: Activators and repressors from the same gene. Cell 1992, 68, 411–414. [Google Scholar] [CrossRef]
- Yang, X.Q.; Zhang, C.X.; Wang, J.K.; Wang, L.; Du, X.; Song, Y.F.; Liu, D. Transcriptional regulation of the porcine miR-17-92 cluster. Mol. Genet. Genom. 2019, 294, 1023–1036. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Yang, Z.; Sun, Y.; Li, J.; Zhang, D.; Liu, D.; Yang, X. Characterization of Alternative Splicing Events in Porcine Skeletal Muscles with Different Intramuscular Fat Contents. Biomolecules 2022, 12, 154. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B. Aligning short sequencing reads with Bowtie. Curr. Protoc. Bioinform. 2010, 32, 11.7.1–11.7.14. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome. Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [Green Version]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Zhang, Q.; Wang, L.; Li, J.; Hao, W.; Sun, Y.; Liu, D.; Yang, X. Alternative Splicing Isoforms of Porcine CREB Are Differentially Involved in Transcriptional Transactivation. Genes 2022, 13, 1304. https://doi.org/10.3390/genes13081304
Zhang D, Zhang Q, Wang L, Li J, Hao W, Sun Y, Liu D, Yang X. Alternative Splicing Isoforms of Porcine CREB Are Differentially Involved in Transcriptional Transactivation. Genes. 2022; 13(8):1304. https://doi.org/10.3390/genes13081304
Chicago/Turabian StyleZhang, Dongjie, Qian Zhang, Liang Wang, Jiaxin Li, Wanjun Hao, Yuanlu Sun, Di Liu, and Xiuqin Yang. 2022. "Alternative Splicing Isoforms of Porcine CREB Are Differentially Involved in Transcriptional Transactivation" Genes 13, no. 8: 1304. https://doi.org/10.3390/genes13081304