
Transcriptomic Profile of Genes Regulating the Structural Organization of Porcine Atrial Cardiomyocytes during Primary In Vitro Culture

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Tissues
2.2. Enzymatic Dissociation and Primary Cell Culture
2.3. Morphological Observation of Cells during Long-Term Primary In Vitro Culture
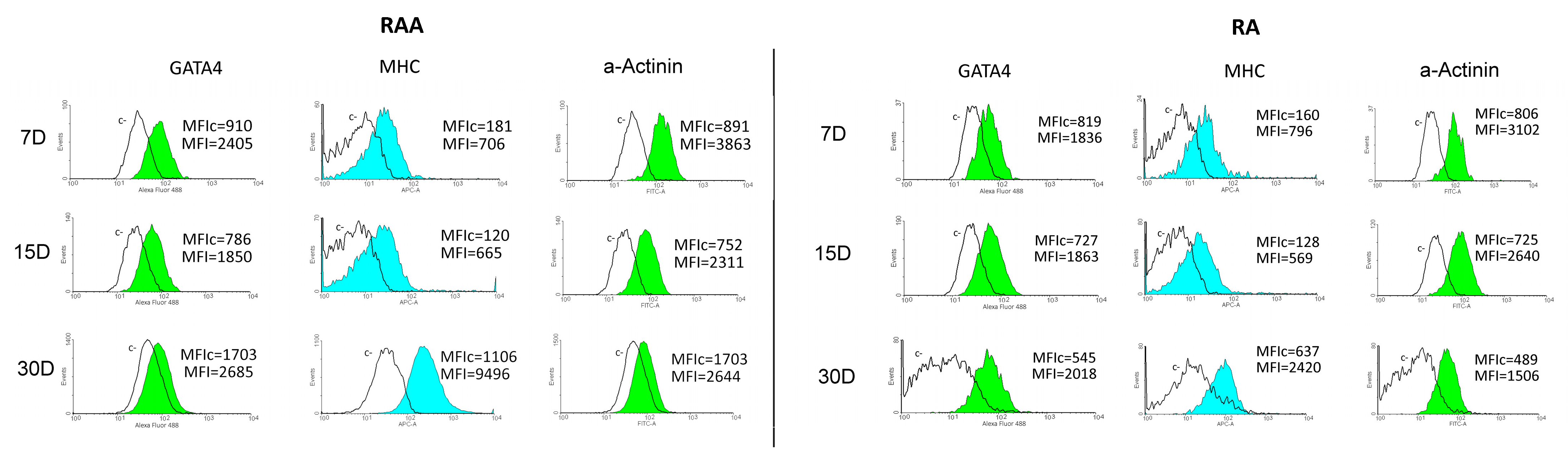
2.4. Flow Cytometry Analysis
2.5. RNA Extraction and Reverse Transcription
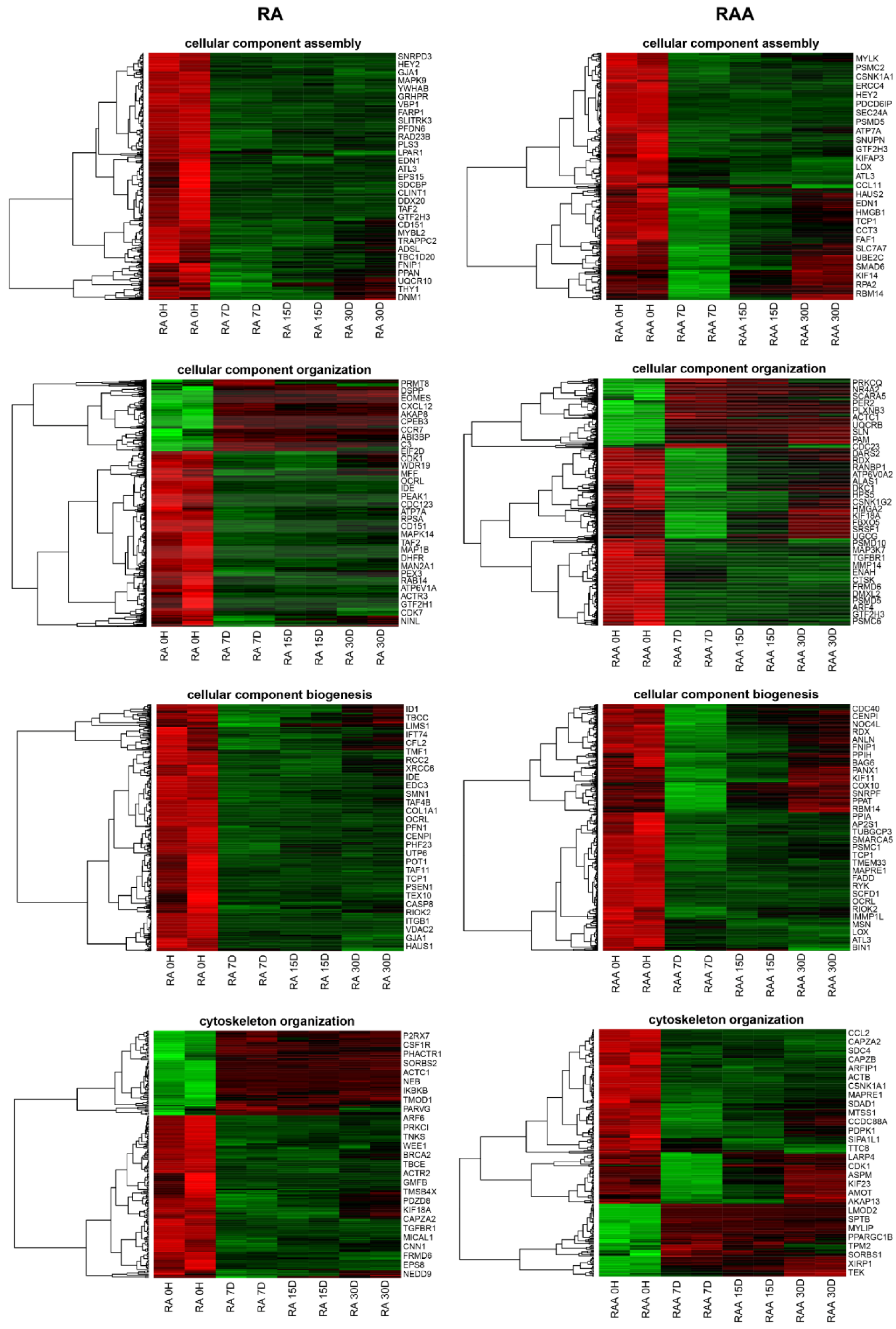
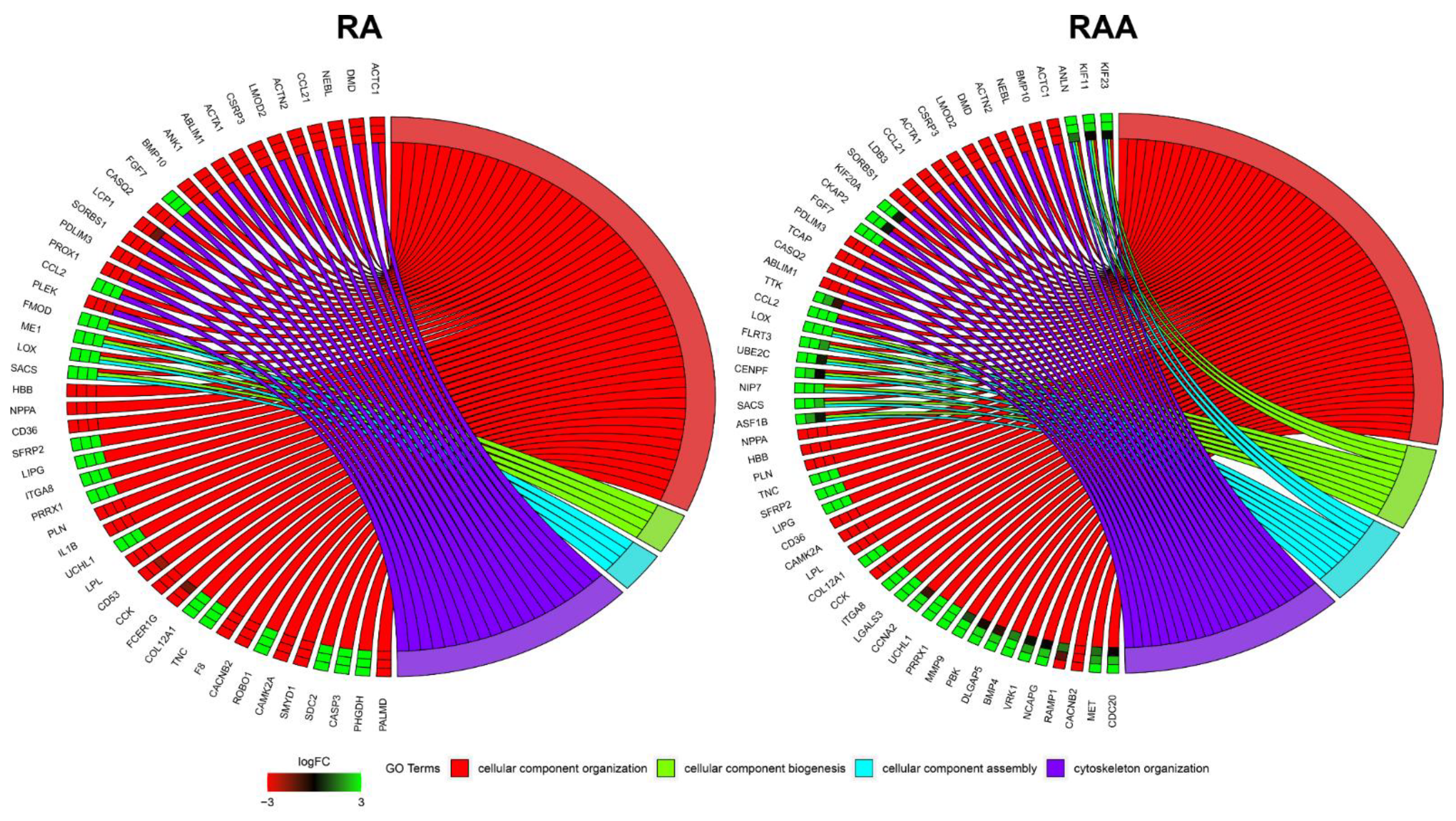
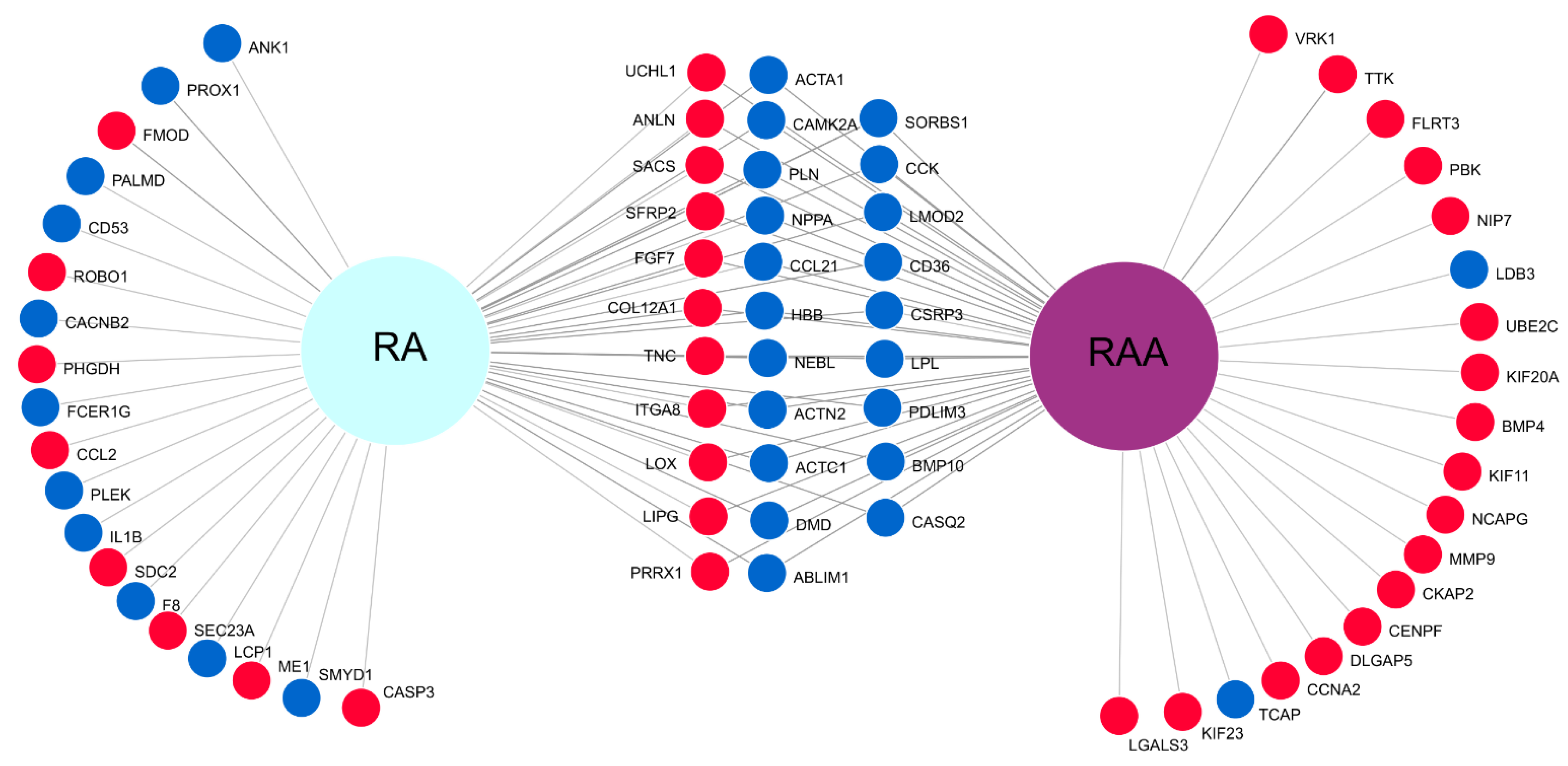
2.6. Microarray Expression Study and Data Analysis
2.7. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
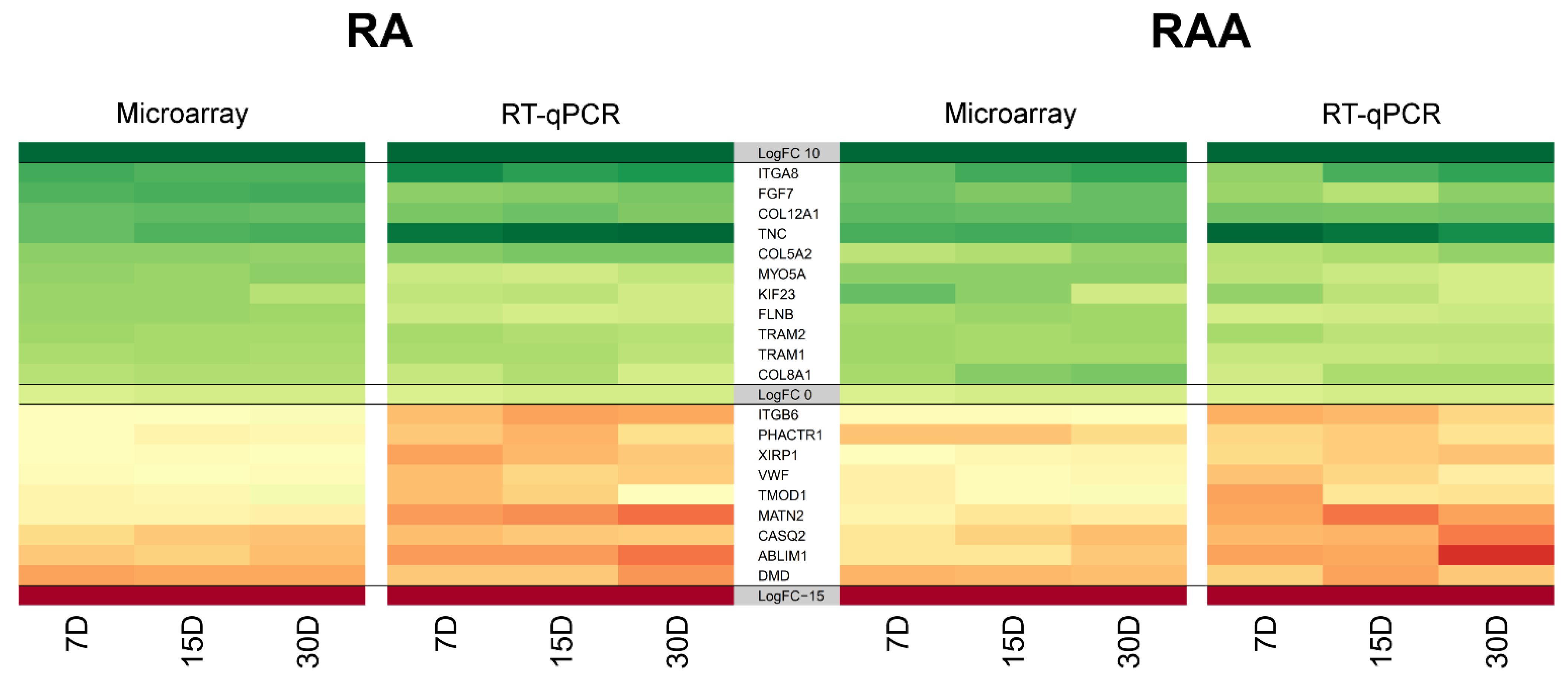
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gummert, J.; Barten, M.; Rahmel, A.; Doll, S.; Krakor, R.; Garbade, J.; Autschbach, R. Therapy for Heart Failure: The Leipzig Approach. Thorac. Cardiovasc. Surg. 2017, 65, S205–S208. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Zuba-Surma, E.K.; Wysoczynski, M.; Wan, W.; Ratajczak, J.; Wojakowski, W.; Kucia, M. Hunt for pluripotent stem cell—Regenerative medicine search for almighty cell. J. Autoimmun. 2008, 30, 151–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, O.; Bhardwaj, R.D.; Bernard, S.; Zdunek, S.; Barnabé-Heider, F.; Walsh, S.; Zupicich, J.; Alkass, K.; Buchholz, B.A.; Druid, H.; et al. Evidence for cardiomyocyte renewal in humans. Science 2009, 324, 98–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, T.L.; Raveendran, G.; Zhang, J.; Garry, D.J. Getting to the heart of myocardial stem cells and cell therapy. Circulation 2011, 123, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Senyo, S.E.; Steinhauser, M.L.; Pizzimenti, C.L.; Yang, V.K.; Cai, L.; Wang, M.; Wu, T.D.; Guerquin-Kern, J.L.; Lechene, C.P.; Lee, R.T. Mammalian heart renewal by pre-existing cardiomyocytes. Nature 2013, 493, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Ellison, G.M.; Vicinanza, C.; Smith, A.J.; Aquila, I.; Leone, A.; Waring, C.D.; Henning, B.J.; Stirparo, G.G.; Papait, R.; Scarfò, M.; et al. Adult c-kitpos Cardiac Stem Cells Are Necessary and Sufficient for Functional Cardiac Regeneration and Repair. Cell 2013, 154, 827–842. [Google Scholar] [CrossRef] [Green Version]
- Hamdani, N.; Kooij, V.; Van Dijk, S.; Merkus, D.; Paulus, W.J.; Dos Remedios, C.; Duncker, D.J.; Stienen, G.J.M.; Van Der Velden, J. Sarcomeric dysfunction in heart failure. Cardiovasc. Res. 2008, 77, 649–658. [Google Scholar] [CrossRef]
- Golob, M.; Moss, R.L.; Chesler, N.C. Cardiac Tissue Structure, Properties, and Performance: A Materials Science Perspective. Ann. Biomed. Eng. 2014, 42, 2003–2013. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsov, A.V.; Javadov, S.; Grimm, M.; Margreiter, R.; Ausserlechner, M.J.; Hagenbuchner, J. Crosstalk between Mitochondria and Cytoskeleton in Cardiac Cells. Cells 2020, 9, 222. [Google Scholar] [CrossRef] [Green Version]
- Song, R.; Zhang, L. Cardiac ECM: Its epigenetic regulation and role in heart development and repair. Int. J. Mol. Sci. 2020, 21, 8610. [Google Scholar] [CrossRef]
- Sreejit, P.; Kumar, S.; Verma, R.S. An improved protocol for primary culture of cardiomyocyte from neonatal mice. Vitr. Cell. Dev. Biol.—Anim. 2008, 44, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Wysoczynski, M.; Guo, Y.; Moore, J.B.; Muthusamy, S.; Li, Q.; Nasr, M.; Li, H.; Nong, Y.; Wu, W.; Tomlin, A.A.; et al. Myocardial Reparative Properties of Cardiac Mesenchymal Cells Isolated on the Basis of Adherence. J. Am. Coll. Cardiol. 2017, 69, 1824–1838. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Kałuzna, S.; Nawrocki, M.J.; Bryl, R.; Stefańska, K.; Jemielity, M.; Mozdziak, P.; Nowicki, M.; Perek, B. Study of the expression of genes associated with post-translational changes in histones in the internal thoracic artery and the saphenous vein grafts used in coronary artery bypass grafting procedure. Med. J. Cell Biol. 2020, 8, 183–189. [Google Scholar] [CrossRef]
- Nawrocki, M.J.; Jopek, K.; Zdun, M.; Mozdziak, P.; Jemielity, M.; Perek, B.; Bukowska, D.; Kempisty, B. Expression profile of genes encoding proteins involved in regulation of vasculature development and heart muscle morphogenesis—A transcriptomic approach based on a porcine model. Int. J. Mol. Sci. 2021, 22, 8794. [Google Scholar] [CrossRef]
- Kałuzna, S.; Nawrocki, M.J.; Jopek, K.; Hutchings, G.; Perek, B.; Jemielity, M.; Kempisty, B.; Malińska, A.; Mozdziak, P.; Nowicki, M. In search of markers useful for evaluation of graft patency-molecular analysis of “muscle system process” for internal thoracic artery and saphenous vein conduits. Med. J. Cell Biol. 2020, 8, 12–23. [Google Scholar] [CrossRef]
- Nawrocki, M.J.; Kałuzna, S.; Jopek, K.; Hutchings, G.; Perek, B.; Jemielity, M.; Malińska, A.; Kempisty, B.; Mozdziak, P.; Nowicki, M. Aortocoronary conduits may show a different inflammatory response-comparative study at transcript level. Med. J. Cell Biol. 2020, 8, 24–34. [Google Scholar] [CrossRef]
- Ichimura, H.; Kadota, S.; Kashihara, T.; Yamada, M.; Ito, K.; Kobayashi, H.; Tanaka, Y.; Shiba, N.; Chuma, S.; Tohyama, S.; et al. Increased predominance of the matured ventricular subtype in embryonic stem cell-derived cardiomyocytes in vivo. Sci. Rep. 2020, 10, 11883. [Google Scholar] [CrossRef]
- Qian, L.; Huang, Y.; Spencer, C.I.; Foley, A.; Vedantham, V.; Liu, L.; Conway, S.J.; Fu, J.; Srivastava, D. In vivo reprogramming of murine cardiac fibroblasts into induced cardiomyocytes. Nature 2012, 485, 593–598. [Google Scholar] [CrossRef]
- Hassan, N.; Tchao, J.; Tobita, K. Concise Review: Skeletal Muscle Stem Cells and Cardiac Lineage: Potential for Heart Repair. Stem Cells Transl. Med. 2014, 3, 183. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, G.; Nawrocki, M.J.; Mozdziak, P.; Kempisty, B. Cardiac Stem Cell Therapy, Resident Progenitor Cells and the role of Cellular Signalling; a Review. Med. J. Cell Biol. 2019, 7, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Nawrocki, M.J.; Bryl, R.; Kałużna, S.; Stefańska, K.; Stelmach, B.; Jemielity, M.; Perek, B.; Bukowska, D.; Mozdziak, P.; Petitte, J.N.; et al. Increased transcript expression levels of DNA methyltransferases type 1 and 3A during cardiac muscle long-term cell culture. Med. J. Cell Biol. 2021, 9, 27–32. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Li, L.; Zhao, Q.; Kong, W. Extracellular matrix remodeling and cardiac fibrosis. Matrix Biol. 2018, 68, 490–506. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The extracellular matrix in ischemic and nonischemic heart failure. Circ. Res. 2019, 125, 117–146. [Google Scholar] [CrossRef]
- Nunes, S.S.; Miklas, J.W.; Liu, J.; Aschar-Sobbi, R.; Xiao, Y.; Zhang, B.; Jiang, J.; Massé, S.; Gagliardi, M.; Hsieh, A.; et al. Biowire: A platform for maturation of human pluripotent stem cell-derived cardiomyocytes. Nat. Methods 2013, 10, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Feaster, T.K.; Cadar, A.G.; Wang, L.; Williams, C.H.; Chun, Y.W.; Hempel, J.E.; Bloodworth, N.; Merryman, W.D.; Lim, C.C.; Wu, J.C.; et al. A method for the generation of single contracting human-induced pluripotent stem cell-derived cardiomyocytes. Circ. Res. 2015, 117, 995–1000. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Shen, B.; Chan, C.W.Y.; Wang, C.; Zhao, Y.; Chan, H.N.; Tian, Q.; Chen, Y.; Yao, C.; et al. Engineering a freestanding biomimetic cardiac patch using biodegradable poly(lactic-co-glycolic acid) (PLGA) and human embryonic stem cell-derived ventricular cardiomyocytes (hESC-VCMs). Macromol. Biosci. 2015, 15, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Herron, T.J.; Da Rocha, A.M.; Campbell, K.F.; Ponce-Balbuena, D.; Willis, B.C.; Guerrero-Serna, G.; Liu, Q.; Klos, M.; Musa, H.; Zarzoso, M.; et al. Extracellular matrix-mediated maturation of human pluripotent stem cell-derived cardiac monolayer structure and electrophysiological function. Circ. Arrhythmia Electrophysiol. 2016, 9, e003638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, A.H.; Romero-López, M.; Heylman, C.M.; Keating, M.; Tran, D.; Sobrino, A.; Tran, A.Q.; Pham, H.H.; Fimbres, C.; Gershon, P.D.; et al. Three-Dimensional Adult Cardiac Extracellular Matrix Promotes Maturation of Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Tissue Eng. 2016, 22, 1016–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garreta, E.; de Oñate, L.; Fernández-Santos, M.E.; Oria, R.; Tarantino, C.; Climent, A.M.; Marco, A.; Samitier, M.; Martínez, E.; Valls-Margarit, M.; et al. Myocardial commitment from human pluripotent stem cells: Rapid production of human heart grafts. Biomaterials 2016, 98, 64–78. [Google Scholar] [CrossRef] [Green Version]
- Guyette, J.P.; Charest, J.M.; Mills, R.W.; Jank, B.J.; Moser, P.T.; Gilpin, S.E.; Gershlak, J.R.; Okamoto, T.; Gonzalez, G.; Milan, D.J.; et al. Bioengineering Human Myocardium on Native Extracellular Matrix. Circ. Res. 2016, 118, 56–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mak, K.M.; Png, C.Y.M.; Lee, D.J. Type V Collagen in Health, Disease, and Fibrosis. Anat. Rec. 2016, 299, 613–629. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.; Takawale, A.; Lee, J.; Kassiri, Z. Cardiac fibroblasts, fibrosis and extracellular matrix remodeling in heart disease. Fibrogenes. Tissue Repair 2012, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Azuaje, F.; Zhang, L.; Jeanty, C.; Puhl, S.L.; Rodius, S.; Wagner, D.R. Analysis of a gene co-expression network establishes robust association between Col5a2 and ischemic heart disease. BMC Med. Genom. 2013, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Keene, D.R.; Jaenisch, R.; Ramirez, F. Targeted mutation in the col5a2 gene reveals a regulatory role for type V collagen during matrix assembly. Nat. Genet. 1995, 9, 31–36. [Google Scholar] [CrossRef]
- Malfait, F.; Wenstrup, R.J.; De Paepe, A. Clinical and genetic aspects of Ehlers-Danlos syndrome, classic type. Genet. Med. 2010, 12, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.B.; Huang, R.; Yang, K.; Xu, M.; Fan, D.; Liu, M.X.; Huang, S.H.; Liu, L.B.; Wu, H.M.; Tang, Q.Z. Identification of differentially expressed genes and preliminary validations in cardiac pathological remodeling induced by transverse aortic constriction. Int. J. Mol. Med. 2019, 44, 1447–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrott, M.E.; Aljrbi, E.; Biederman, D.L.; Montalvo, R.N.; Barth, J.L.; LaVoie, H.A. Feature Article: Maternal cardiac messenger RNA expression of extracellular matrix proteins in mice during pregnancy and the postpartum period. Exp. Biol. Med. 2018, 243, 1220–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skrbic, B.; Engebretsen, K.V.T.; Strand, M.E.; Lunde, I.G.; Herum, K.M.; Marstein, H.S.; Sjaastad, I.; Lunde, P.K.; Carlson, C.R.; Christensen, G.; et al. Lack of collagen VIII reduces fibrosis and promotes early mortality and cardiac dilatation in pressure overload in mice. Cardiovasc. Res. 2015, 106, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Manso, A.M.; Ross, R.S. Talin and Kindlin as Integrin-Activating Proteins: Focus on the Heart. Pediatr. Cardiol. 2019, 40, 1401–1409. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shang, H.; Li, B.; Zhao, L.; Hua, Y.; Wu, K.; Hu, M.; Fan, T. Exploration and validation of hub genes and pathways in the progression of hypoplastic left heart syndrome via weighted gene co-expression network analysis. BMC Cardiovasc. Disord. 2021, 21, 300. [Google Scholar] [CrossRef]
- Chen, Y.M.; Li, H.; Fan, Y.; Zhang, Q.J.; Li, X.; Wu, L.J.; Chen, Z.J.; Zhu, C.; Qian, L.M. Identification of differentially expressed lncRNAs involved in transient regeneration of the neonatal C57BL/6J mouse heart by next-generation high-throughput RNA sequencing. Oncotarget 2017, 8, 28052–28062. [Google Scholar] [CrossRef] [Green Version]
- Imanaka-Yoshida, K. Tenascin-c in heart diseases—The role of inflammation. Int. J. Mol. Sci. 2021, 22, 5828. [Google Scholar] [CrossRef]
- Kanagala, P.; Arnold, J.R.; Khan, J.N.; Singh, A.; Gulsin, G.S.; Chan, D.C.S.; Cheng, A.S.H.; Yang, J.; Li, Z.; Gupta, P.; et al. Plasma Tenascin-C: A prognostic biomarker in heart failure with preserved ejection fraction. Biomarkers 2020, 25, 556–565. [Google Scholar] [CrossRef]
- Imanaka-Yoshida, K.; Tawara, I.; Yoshida, T. Tenascin-C in cardiac disease: A sophisticated controller of inflammation, repair, and fibrosis. Am. J. Physiol.—Cell Physiol. 2020, 319, C781–C796. [Google Scholar] [CrossRef]
- Radosinska, J.; Barancik, M.; Vrbjar, N. Heart failure and role of circulating MMP-2 and MMP-9. Panminerva Med. 2017, 59, 241–253. [Google Scholar] [CrossRef]
- Hughes, B.G.; Schulz, R. Targeting MMP-2 to treat ischemic heart injury. Basic Res. Cardiol. 2014, 109, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Spinale, F.G. Myocardial matrix remodeling and the matrix metalloproteinases: Influence on cardiac form and function. Physiol. Rev. 2007, 87, 1285–1342. [Google Scholar] [CrossRef] [PubMed]
- Kiczak, L.; Tomaszek, A.; Bania, J.; Paslawska, U.; Zacharski, M.; Noszczyk-Nowak, A.; Janiszewski, A.; Skrzypczak, P.; Ardehali, H.; Jankowska, E.A.; et al. Expression and complex formation of MMP9, MMP2, NGAL, and TIMP1 in porcine myocardium but not in skeletal muscles in male pigs with tachycardia-induced systolic heart failure. Biomed. Res. Int. 2013, 2013, 283856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashidani, S.; Tsutsui, H.; Ikeuchi, M.; Shiomi, T.; Matsusaka, H.; Kubota, T.; Imanaka-Yoshida, K.; Itoh, T.; Takeshita, A. Targeted deletion of MMP-2 attenuates early LV rupture and late remodeling after experimental myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H1229–H1235. [Google Scholar] [CrossRef] [Green Version]
- Apple, K.A.; Yarbrough, W.M.; Mukherjee, R.; Deschamps, A.M.; Escobar, P.G.; Mingoia, J.T.; Sample, J.A.; Hendrick, J.W.; Dowdy, K.B.; McLean, J.E.; et al. Selective targeting of matrix metalloproteinase inhibition in post-infarction myocardial remodeling. J. Cardiovasc. Pharmacol. 2006, 47, 228–235. [Google Scholar] [CrossRef]
- Langford, G.M. Myosin-V, a versatile motor for short-range vesicle transport. Traffic 2002, 3, 859–865. [Google Scholar] [CrossRef]
- Schumacher-Bass, S.M.; Vesely, E.D.; Zhang, L.; Ryland, K.E.; McEwen, D.P.; Chan, P.J.; Frasier, C.R.; McIntyre, J.C.; Shaw, R.M.; Martens, J.R. Role for myosin-V motor proteins in the selective delivery of Kv channel isoforms to the membrane surface of cardiac myocytes. Circ. Res. 2014, 114, 982–992. [Google Scholar] [CrossRef] [Green Version]
- Krupp, M.; Weinmann, A.; Galle, P.R.; Teufel, A. Actin binding LIM protein 3 (abLIM3). Int. J. Mol. Med. 2006, 17, 129–133. [Google Scholar] [CrossRef]
- Roof, D.J.; Hayes, A.; Adamian, M.; Chishti, A.H.; Li, T. Molecular characterization of abLIM, a novel actin-binding and double zinc finger protein. J. Cell Biol. 1997, 138, 575–588. [Google Scholar] [CrossRef]
- Kostyukova, A.S.; Rapp, B.A.; Choy, A.; Greenfield, N.J.; Hitchcock-DeGregori, S.E. Structural requirements of tropomodulin for tropomyosin binding and actin filament capping. Biochemistry 2005, 44, 4905–4910. [Google Scholar] [CrossRef]
- Bliss, K.T.; Tsukada, T.; Novak, S.M.; Dorovkov, M.V.; Shah, S.P.; Nworu, C.; Kostyukova, A.S.; Gregorio, C.C. Phosphorylation of tropomodulin1 contributes to the regulation of actin filament architecture in cardiac muscle. FASEB J. 2014, 28, 3987–3995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, J.P.; Raslova, H.; Bryckaert, M. Filamin A: Key actor in platelet biology. Blood 2019, 134, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Kley, R.A.; Maerkens, A.; Leber, Y.; Theis, V.; Schreiner, A.; Van Der Ven, P.F.M.; Uszkoreit, J.; Stephan, C.; Eulitz, S.; Euler, N.; et al. A combined laser microdissection and mass spectrometry approach reveals new disease relevant proteins accumulating in aggregates of filaminopathy patients. Mol. Cell. Proteom. 2013, 12, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Leber, Y.; Ruparelia, A.A.; Kirfel, G.; van der Ven, P.F.M.; Hoffmann, B.; Merkel, R.; Bryson-Richardson, R.J.; Fürst, D.O. Filamin C is a highly dynamic protein associated with fast repair of myofibrillar microdamage. Hum. Mol. Genet. 2016, 25, 2776–2788. [Google Scholar] [CrossRef] [Green Version]
- Allen, P.B.; Greenfield, A.T.; Svenningsson, P.; Haspeslagh, D.C.; Greengard, P. Phactrs 1–4: A family of protein phosphatase 1 and actin regulatory proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 7187–7192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codina-Fauteux, V.A.; Beaudoin, M.; Lalonde, S.; Lo, K.S.; Lettre, G. PHACTR1 splicing isoforms and eQTLs in atherosclerosis-relevant human cells. BMC Med. Genet. 2018, 19, 97. [Google Scholar] [CrossRef]
- Allain, B.; Jarray, R.; Borriello, L.; Leforban, B.; Dufour, S.; Liu, W.Q.; Pamonsinlapatham, P.; Bianco, S.; Larghero, J.Ô.; Hadj-Slimane, R.; et al. Neuropilin-1 regulates a new VEGF-induced gene, Phactr-1, which controls tubulogenesis and modulates lamellipodial dynamics in human endothelial cells. Cell. Signal. 2012, 24, 214–223. [Google Scholar] [CrossRef]
- Jarray, R.; Allain, B.; Borriello, L.; Biard, D.; Loukaci, A.; Larghero, J.; Hadj-Slimane, R.; Garbay, C.; Lepelletier, Y.; Raynaud, F. Depletion of the novel protein PHACTR-1 from human endothelial cells abolishes tube formation and induces cell death receptor apoptosis. Biochimie 2011, 93, 1668–1675. [Google Scholar] [CrossRef]
- Jarray, R.; Pavoni, S.; Borriello, L.; Allain, B.; Lopez, N.; Bianco, S.; Liu, W.Q.; Biard, D.; Demange, L.; Hermine, O.; et al. Disruption of phactr-1 pathway triggers pro-inflammatory and pro-atherogenic factors: New insights in atherosclerosis development. Biochimie 2015, 118, 151–161. [Google Scholar] [CrossRef]
- Reschen, M.E.; Lin, D.; Chalisey, A.; Soilleux, E.J.; O’Callaghan, C.A. Genetic and environmental risk factors for atherosclerosis regulate transcription of phosphatase and actin regulating gene PHACTR1. Atherosclerosis 2016, 250, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Hao, K.; Ermel, R.; Li, L.; Amadori, L.; Koplev, S.; Franzén, O.; D’Escamard, V.; Chandel, N.; Wolhuter, K.; Bryce, N.S.; et al. Integrative Prioritization of Causal Genes for Coronary Artery Disease. Circ. Genom. Precis. Med. 2022, 15, E003365. [Google Scholar] [CrossRef] [PubMed]
- Georges, A.; Yang, M.L.; Berrandou, T.E.; Bakker, M.K.; Dikilitas, O.; Kiando, S.R.; Ma, L.; Satterfield, B.A.; Sengupta, S.; Yu, M.; et al. Genetic investigation of fibromuscular dysplasia identifies risk loci and shared genetics with common cardiovascular diseases. Nat. Commun. 2021, 12, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Saw, J.; Yang, M.L.; Trinder, M.; Tcheandjieu, C.; Xu, C.; Starovoytov, A.; Birt, I.; Mathis, M.R.; Hunker, K.L.; Schmidt, E.M.; et al. Chromosome 1q21.2 and additional loci influence risk of spontaneous coronary artery dissection and myocardial infarction. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- D’Amario, D.; Amodeo, A.; Adorisio, R.; Tiziano, F.D.; Leone, A.M.; Perri, G.; Bruno, P.; Massetti, M.; Ferlini, A.; Pane, M.; et al. A current approach to heart failure in Duchenne muscular dystrophy. Heart 2017, 103, 1770–1779. [Google Scholar] [CrossRef]
- Kamdar, F.; Garry, D.J. Dystrophin-Deficient Cardiomyopathy. J. Am. Coll. Cardiol. 2016, 67, 2533–2546. [Google Scholar] [CrossRef]
- Faggioni, M.; Knollmann, B.C. Calsequestrin 2 and arrhythmias. Am. J. Physiol.—Heart Circ. Physiol. 2012, 302, H1250–H1260. [Google Scholar] [CrossRef] [Green Version]
- Refaat, M.M.; Aouizerat, B.E.; Pullinger, C.R.; Malloy, M.; Kane, J.; Tseng, Z.H. Association of CASQ2 polymorphisms with sudden cardiac arrest and heart failure in patients with coronary artery disease. Heart Rhythm. 2014, 11, 646–652. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Product Size (bp) | |
|---|---|---|---|
| ITGA8 | F | CCAGCAGACCAAAACCCTTC | 164 |
| R | AAGAAGTTGTGCAGCTGTGG | ||
| TNC | F | TTTCAGATGCCACCCCAGAT | 169 |
| R | GTGGCTTCTCTGAGACCTGT | ||
| FGF7 | F | TGGAAATCAGGACAGTGGCT | 192 |
| R | CTCCTCCACTGTGTGTCCAT | ||
| COL12A1 | F | TCCACAGGTTCAAGAGGTCC | 150 |
| R | TTGTTAGCCGGAACCTGGAT | ||
| KIF23 | F | TGCAACAGGAGCTTGAAACC | 243 |
| R | AGGGTCTCTCTGGCTTTTCA | ||
| FLNB | F | AACATCCCGAACAGCCCTTA | 159 |
| R | ACTGACATCACCTTCCCCAG | ||
| COL5A2 | F | TGGTGAAAATGGCCCAACTG | 193 |
| R | TCCTCGACCACCTTTCAGTC | ||
| COL8A1 | F | GGAGAGAAGGGCTTTGGGAT | 249 |
| R | GATCCCATCCTGACCTGGTT | ||
| MYO5A | F | TGAGAAGAAGGTGCCTCTGG | 199 |
| R | TTCCTGACGCTTGAGTGACT | ||
| TRAM1 | F | CCTCGTCAGCTCGTCTACAT | 241 |
| R | AGCCAACAGTGAGTACCGAA | ||
| TRAM2 | F | ACATCTGCCTGTACCTGGTC | 209 |
| R | GGCGAGAGTGAGGATGAAGA | ||
| DMD | F | TCCACTTCTGTCCAAGGTCC | 187 |
| R | GCAGTCTTCGGAGCTTCATG | ||
| ABLIM1 | F | ATGAAGCTCAACTCAGGCCT | 161 |
| R | TAGCCTGGGAGAGATGAGGT | ||
| CASQ2 | F | TCCTTGTCTATGCAACGGGT | 187 |
| R | GCTTTTCCCAGGTGTTGAGG | ||
| TMOD1 | F | ACAGCCGGGTCATAGATCAG | 159 |
| R | GTCAGGGTCCAACTCATCCA | ||
| XIRP1 | F | AGAGCAATGCAGTGAGGACT | 201 |
| R | AGTCCTTCTCGTCCACCAAG | ||
| PHACTR1 | F | TTAACTCGGAAGCTCAGCCA | 174 |
| R | GAGCTCCTTTCGAATGGCAG | ||
| MATN2 | F | ACGACTTGCAGAATCCAGGA | 156 |
| R | TGAGGCACAGTAGTCCACAG | ||
| VWF | F | TGCAACACTTGTGTCTGTCG | 229 |
| R | TGCATTTCAGGGAGGGGTAG | ||
| ITGB6 | F | TGACGACCTCAACACGATCA | 190 |
| R | TCCAAAGGTAGGCAAGCAGA | ||
| ACTB | F | CCCTGGAGAAGAGCTACGAG | 156 |
| R | CGTCGCACTTCATGATGGAG | ||
| HPRT1 | F | CCATCACATCGTAGCCCTCT | 166 |
| R | TATATCGCCCGTTGACTGGT | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawrocki, M.J.; Jopek, K.; Kaczmarek, M.; Zdun, M.; Mozdziak, P.; Jemielity, M.; Perek, B.; Bukowska, D.; Kempisty, B. Transcriptomic Profile of Genes Regulating the Structural Organization of Porcine Atrial Cardiomyocytes during Primary In Vitro Culture. Genes 2022, 13, 1205. https://doi.org/10.3390/genes13071205
Nawrocki MJ, Jopek K, Kaczmarek M, Zdun M, Mozdziak P, Jemielity M, Perek B, Bukowska D, Kempisty B. Transcriptomic Profile of Genes Regulating the Structural Organization of Porcine Atrial Cardiomyocytes during Primary In Vitro Culture. Genes. 2022; 13(7):1205. https://doi.org/10.3390/genes13071205
Chicago/Turabian StyleNawrocki, Mariusz J., Karol Jopek, Mariusz Kaczmarek, Maciej Zdun, Paul Mozdziak, Marek Jemielity, Bartłomiej Perek, Dorota Bukowska, and Bartosz Kempisty. 2022. "Transcriptomic Profile of Genes Regulating the Structural Organization of Porcine Atrial Cardiomyocytes during Primary In Vitro Culture" Genes 13, no. 7: 1205. https://doi.org/10.3390/genes13071205