
Elevated ETV6 Expression in Glioma Promotes an Aggressive In Vitro Phenotype Associated with Shorter Patient Survival
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Background
2. Methods
2.1. Tissue Microarray and Immunohistochemistry
2.2. Database Interrogations
2.3. Flow Cytometry
2.4. Lentiviral Production and Infection
2.5. CCK8 and EdU Assays
2.6. Clonogenic Formation
2.7. Migration and Invasion Assays
2.8. RNA-Seq Library Preparation and Sequencing
2.9. RNA-Seq Read Mapping, Assembly, Differential Expression, and Functional Analysis
2.10. Western Blot
2.11. Statistics
3. Results
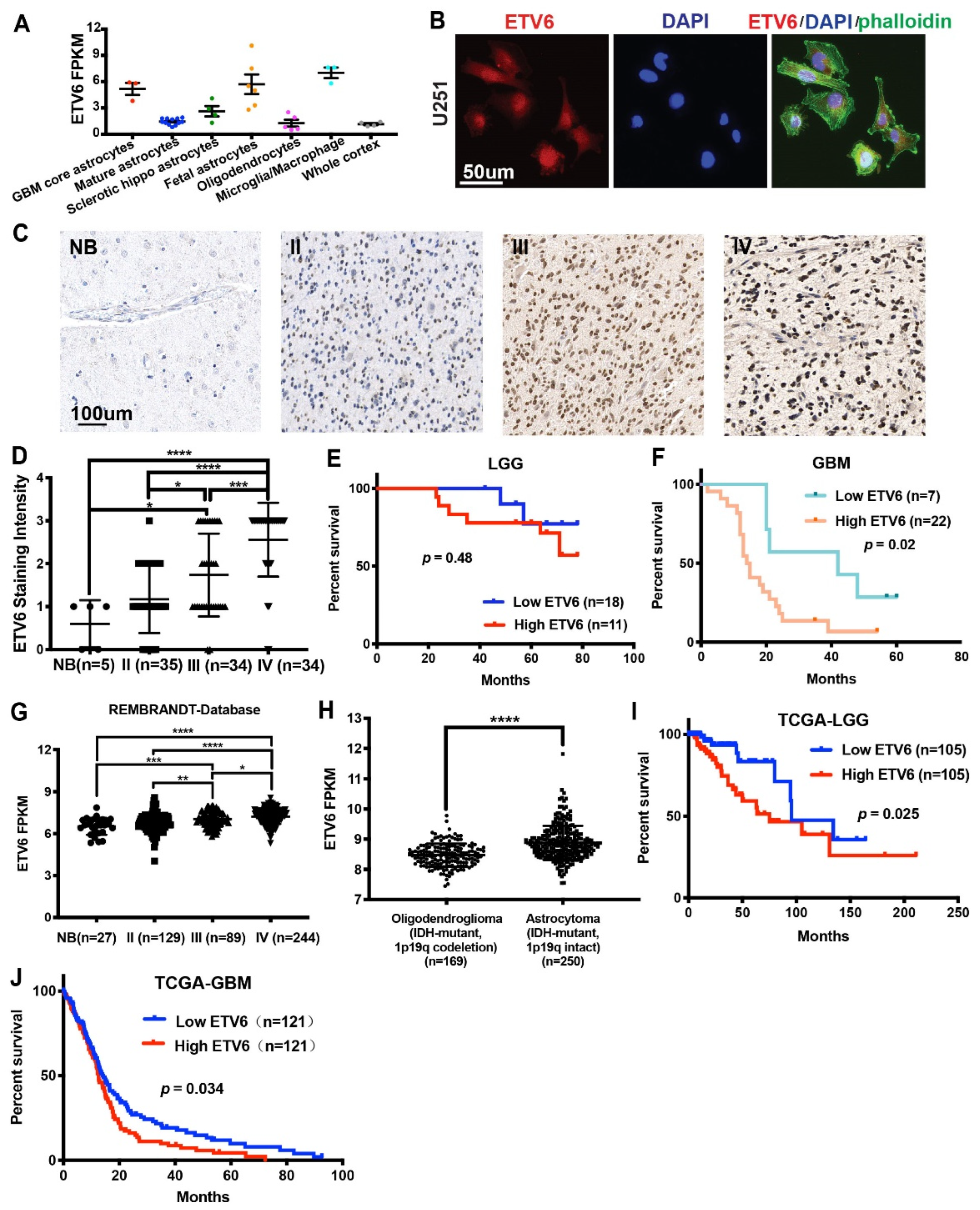
3.1. ETV6 Was Highly Expressed in GBM and Correlated with Poor Prognosis
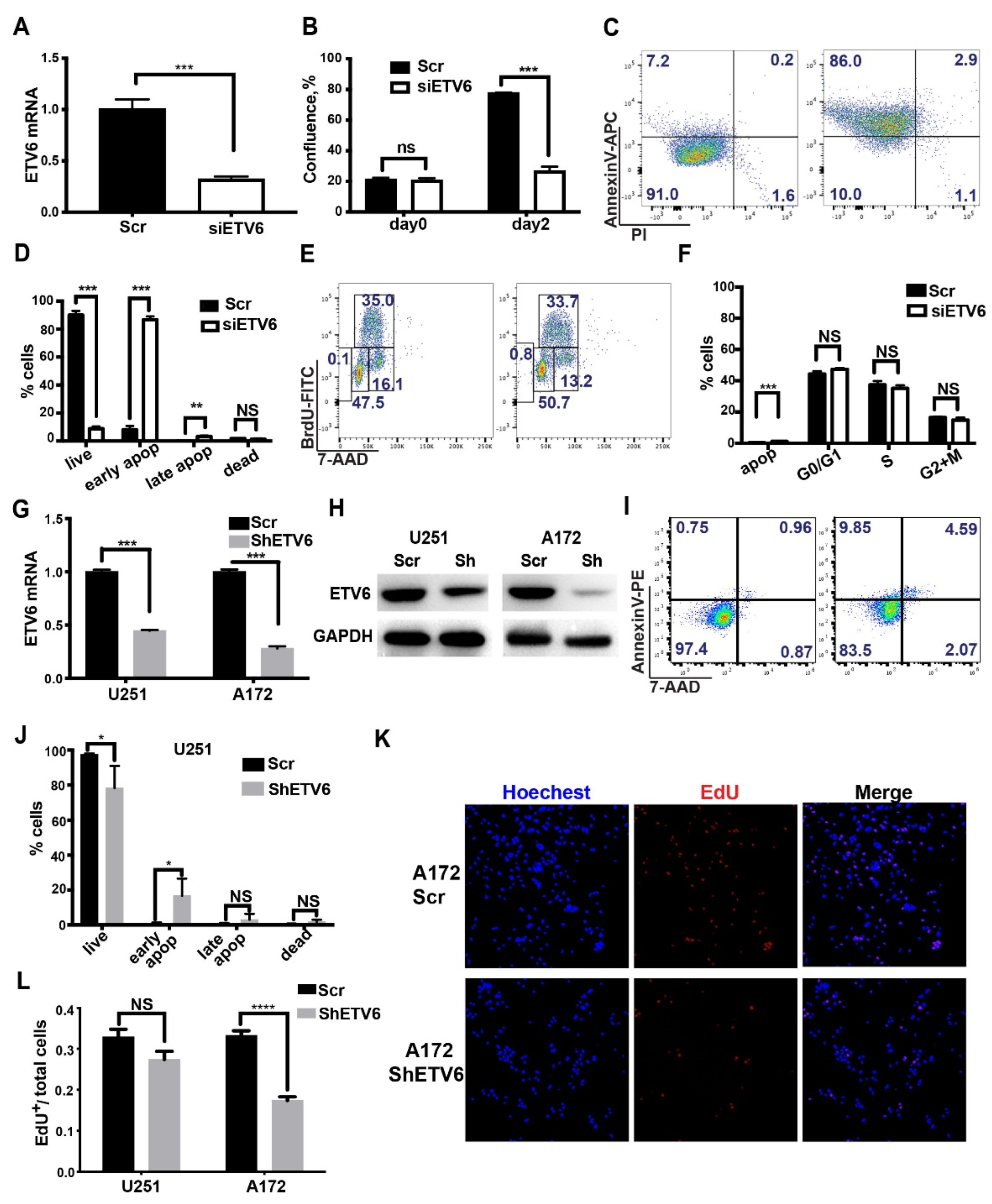
3.2. Knockdown of ETV6 in GBM Cell Lines Induced Apoptosis or Attenuated Proliferation
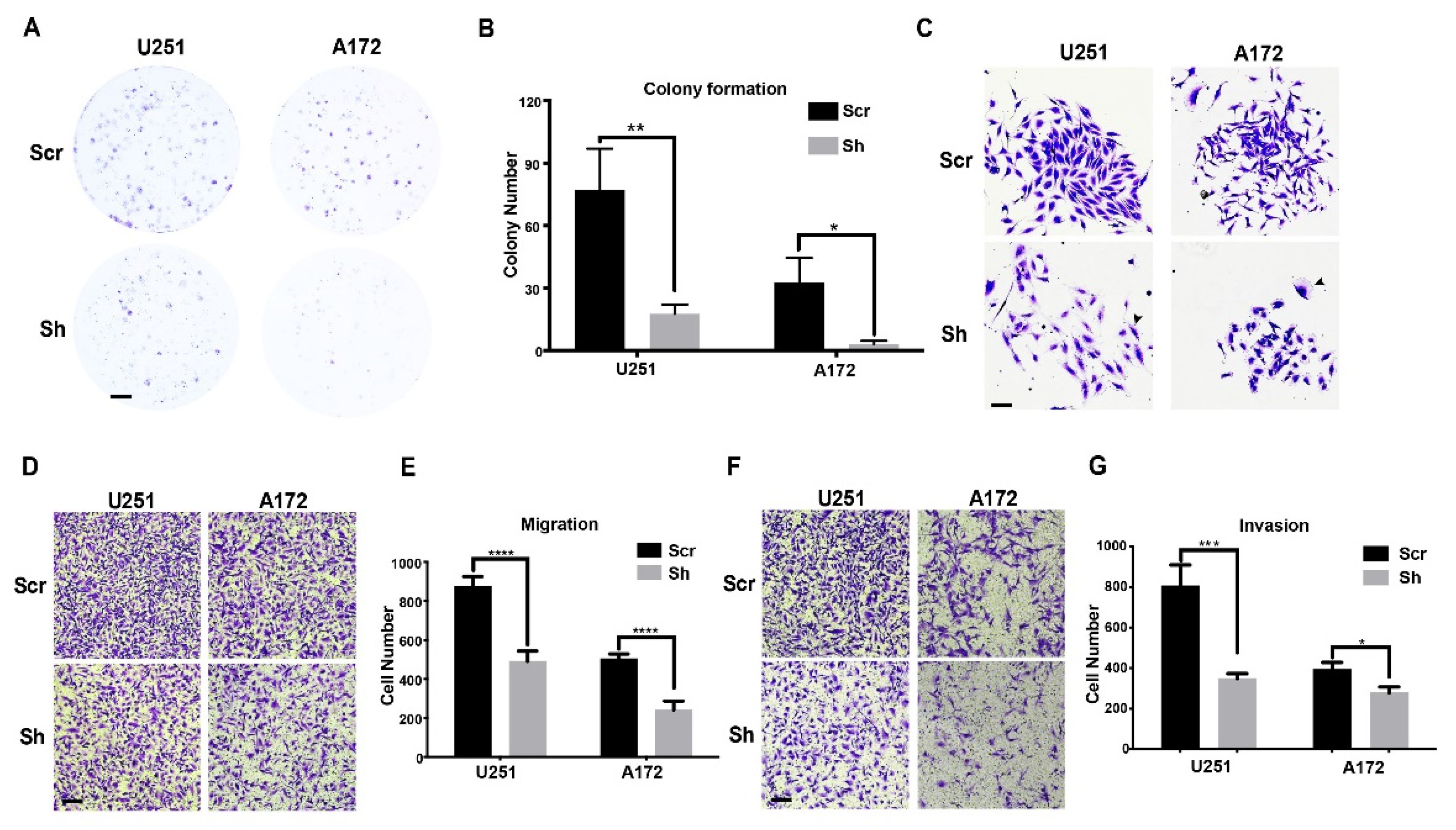
3.3. Knockdown of ETV6 in GBM Cell Lines Decelerated Clonogenic Growth and Inhibited Migration/Invasion
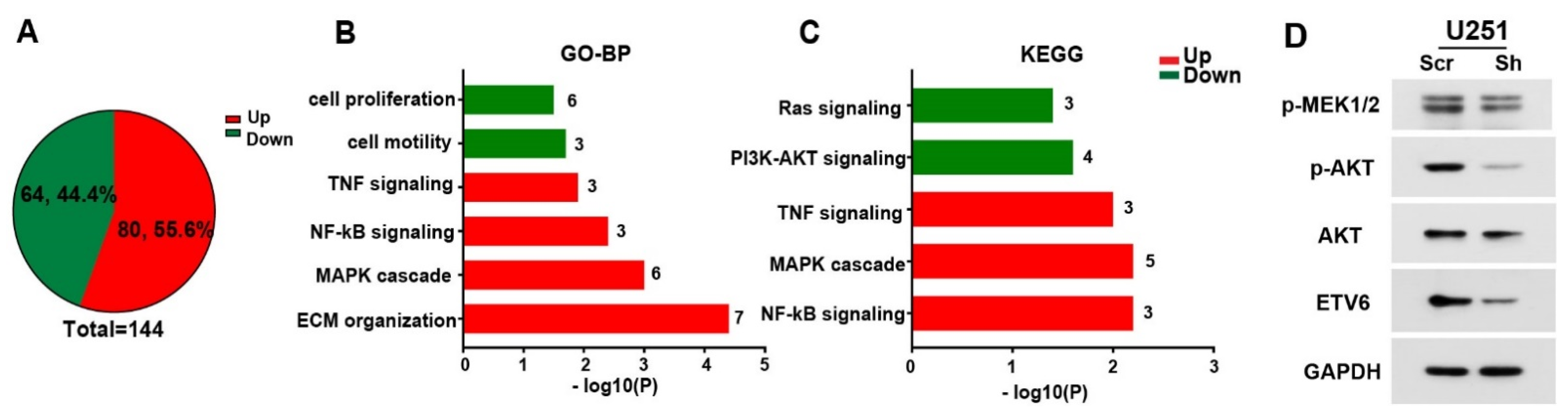
3.4. ETV6-Dependent GBM Phenotype may Involve PI3K-AKT and Ras-MAPK Pathways In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ETV6 | ETS variant 6 |
| GBM | glioblastoma |
| TCGA | The Cancer Genome Atlas |
| REMBRANDT | the REpository for Molecular BRAin Neoplasia DaTa |
| NF-κB | nuclear factor-κB |
| TNF | tumor necrosis factor |
| PI3K | phosphatidylinositol 3-kinase |
| AKT | protein kinase B |
| MAPK | mitogen-activated protein kinase |
| PNT | pointed |
| ERG | ETS-related gene |
| PDGFRβ | platelet derived growth factor receptor β |
| AML1 | acute myeloid leukemia 1 |
| MN1 | meningioma (disrupted in balanced translocation) 1 |
| JAK2 | Janus kinase 2 |
| EVI1 | ecotropic viral integration site 1 |
| CDX2 | caudal type homeobox 2 |
| BTL | breathless |
| NTRK3 | neurotrophic receptor tyrosine kinase |
| 3TMAs | tissue microarrays |
| IDH | isocitrate dehydrogenase |
| MGMT | O6-methylguanine-DNA methyltransferase |
| RT-qPCR | quantitative reverse transcription PCR |
| siETV6 | small interfering ETV6 |
| shETV6 | short hairpin ETV6 |
| CCK8 | cell counting kit-8 |
| GO | gene ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| BrdU | 5-bromo-2’-deoxyuridine |
| FITC | fluorescein isothiocyanate |
| 7-AAD | 7-aminoactinomycin D |
| APC | allophycocyanin |
| PI | propidium iodide |
References
- Zhang, Y.; Sloan, S.A.; Clarke, L.E.; Caneda, C.; Plaza, C.A.; Blumenthal, P.D.; Vogel, H.; Steinberg, G.K.; Edwards, M.S.B.; Li, G.; et al. Purification and Characterization of Progenitor and Mature Human Astrocytes Reveals Transcriptional and Functional Differences with Mouse. Neuron 2016, 89, 37–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molofsk, A.V.; Krenick, R.; Ullian, E.; Tsai, H.H.; Deneen, B.; Richardson, W.D.; Barres, B.A.; Rowitch, D.H. Astrocytes and disease: A neurodevelopmental perspective. Genes Dev. 2012, 26, 891–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowitch, D.H.; Kriegstein, A.R. Developmental genetics of vertebrate glial-cell specification. Nature 2010, 468, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.A.; Mille-Baker, B.; Wainwright, S.M.; Ish-Horowicz, D.; Dibb, N.J. Mae mediates MAP kinase phosphorylation of Ets transcription factors in Drosophila. Nature 2001, 411, 330–334. [Google Scholar] [CrossRef]
- Oikawa, T.; Yamada, T. Molecular biology of the Ets family of transcription factors. Gene 2003, 303, 11–34. [Google Scholar] [CrossRef]
- Seth, A.; Watson, D.K. ETS transcription factors and their emerging roles in human cancer. Eur. J. Cancer 2005, 41, 2462–2478. [Google Scholar] [CrossRef]
- Sementchenko, V.I.; Watson, D.K. Ets target genes: Past, present and future. Oncogene 2000, 19, 6533–6548. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Gu, D.; Du, M.; Xu, Z.; Zhang, S.; Zhu, L.; Lu, J.; Zhang, R.; Xing, J.; Miao, X.; et al. Common genetic variation in ETV6 is associated with colorectal cancer susceptibility. Nat. Commun. 2016, 7, 11478. [Google Scholar] [CrossRef] [Green Version]
- Golub, T.R.; Barker, G.F.; Lovett, M.; Gilliland, D.G. Fusion of PDGF receptor β to a novel ets-like gene, tel, in chronic myelomonocytic leukemia with t(5;12) chromosomal translocation. Cell 1994, 77, 307–316. [Google Scholar] [CrossRef]
- Golub, T.R.; Barker, G.F.; Bohlander, S.K.; Hiebert, S.W.; Ward, D.C.; Bray-Ward, P.; Morgan, E.; Raimondi, S.C.; Rowley, J.D.; Gilliland, D.G. Fusion of the TEL gene on 12p13 to the AML1 gene on 21q22 in acute lymphoblastic leukemia. Proc. Natl. Acad. Sci. USA 1995, 92, 4917–4921. [Google Scholar] [CrossRef]
- Romana, S.P.; Mauchauffe, M.; Le Coniat, M.; Chumakov, I.; Le Paslier, D.; Berger, R.; Bernard, O.A. The t(12;21) of acute lymphoblastic leukemia results in a tel-AML1 gene fusion. Blood 1995, 85, 3662–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, P.; Ridge, S.A.; Boucher, C.A.; Stocking, C.; Wiedemann, L.M. The Novel Activation of ABL by Fusion to an eta-related Gene, TEL. Cancer Res. 1995, 55, 34–38. [Google Scholar] [PubMed]
- Buijs, A.; Sherr, S.; Van Baal, S.; Van Bezouw, S.; Van der Plas, D.; Van Kessel, A.G.; Riegman, P.; Deprez, R.L.; Zwarthoff, E.; Hagemeijer, A.; et al. Translocation (12;22) (p13;q11) in myeloproliferative disorders results in fusion of the ETS-like TEL gene on 12p13 to the MN1 gene on 22q11. Oncogene 1995, 10, 1511–1519. [Google Scholar] [PubMed]
- Lacronique, V.; Boureux, A.; Valle, V.D.; Poirel, H.; Quang, C.T.; Mauchauffé, M.; Berthou, C.; Lessard, M.; Berger, R.; Ghysdael, J.; et al. A TEL-JAK2 fusion protein with constitutive kinase activity in human leukemia. Science 1997, 278, 1309–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeters, P.; Raynaud, S.D.; Cools, J.; Wlodarska, I.; Grosgeorge, J.; Philip, P.; Monpoux, F.; Van Rompaey, L.; Baens, M.; Van Den Berghe, H.; et al. Fusion of TEL, the ETS-variant gene 6 (ETV6), to the receptor- associated kinase JAK2 as a result of t(9;12) in a lymphoid and t(9;15;12) in a myeloid leukemia. Blood 1997, 90, 2535–2540. [Google Scholar] [CrossRef] [Green Version]
- Peeters, P.; Wlodarska, I.; Baens, M.; Criel, A.; Selleslag, D.; Hagemeijer, A.; Van den Berghe, H.; Marynen, P. Fusion of ETV6 to MDS1/EVI1 as a result of t(3;12)(q26;p13) in myeloproliferative disorders. Cancer Res. 1997, 57, 564–569. [Google Scholar]
- Chase, A.; Reiter, A.; Burci, L.; Cazzaniga, G.; Biondi, A.; Pickard, J.; Roberts, I.A.; Goldman, J.M.; Cross, N.C. Fusion of ETV6 to the caudal-related homeobox gene CDX2 in acute myeloid leukemia with the t(12;13)(p13;q12). Blood 1999, 93, 1025–1031. [Google Scholar] [CrossRef]
- Cools, J.; Bilhou-Nabera, C.; Wlodarska, I.; Cabrol, C.; Talmant, P.; Bernard, P.; Hagemeijer, A.; Marynen, P. Fusion of a novel gene, BTL, to ETV6 in acute myeloid leukemias with a t(4;12)(q11-q12;p13). Blood 1999, 94, 1820–1824. [Google Scholar] [CrossRef]
- Knezevich, S.R.; McFadden, D.E.; Tao, W.; Lim, J.F.; Sorensen, P.H. A novel ETV6-NTRK3 gene fusion in congenital fibrosarcoma. Nat. Genet. 1998, 18, 184–187. [Google Scholar] [CrossRef]
- Hunt, J.L. An update on molecular diagnostics of squamous and salivary gland tumors of the head and neck. Arch. Pathol. Lab. Med. 2011, 135, 602–609. [Google Scholar] [CrossRef]
- Tognon, C.; Knezevich, S.R.; Huntsman, D.; Roskelley, C.D.; Melnyk, N.; Mathers, J.A.; Becker, L.; Carneiro, F.; MacPherson, N.; Horsman, D.; et al. Expression of the ETV6-NTRK3 gene fusion as a primary event in human secretory breast carcinoma. Cancer Cell 2002, 2, 367–376. [Google Scholar] [CrossRef]
- Wai, D.H.; Knezevich, S.R.; Lucas, T.; Jansen, B.; Kay, R.J.; Sorensen, P.H. The ETV6-NTRK3 gene fusion encodes a chimeric protein tyrosine kinase that transforms NIH3T3 cells. Oncogene 2000, 19, 906–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tognon, C.; Garnett, M.; Kenward, E.; Kay, R.; Morrison, K.; Sorensen, P.H. The chimeric protein tyrosine kinase ETV6-NTRK3 requires both Ras-Erk1/2 and PI3-kinase-Akt signaling for fibroblast transformation. Cancer Res. 2001, 61, 8909–8916. [Google Scholar] [PubMed]
- Liang, J.Z.; Li, Y.H.; Zhang, Y.; Wu, Q.N.; Wu, Q.L. Expression of ETV6/TEL is associated with prognosis in non-small cell lung cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 2937–2945. [Google Scholar]
- Ke, L.; Zhou, H.; Wang, C.; Xiong, G.; Xiang, Y.; Ling, Y.; Khabir, A.; Tsao, G.S.; Zeng, Y.; Zeng, M.; et al. Nasopharyngeal carcinoma super-enhancer–driven ETV6 correlates with prognosis. Proc. Natl. Acad. Sci. USA 2017, 114, 9683–9688. [Google Scholar] [CrossRef] [Green Version]
- Sizemore, G.M.; Pitarresi, J.R.; Balakrishnan, S.; Ostrowski, M.C. The ETS family of oncogenic transcription factors in solid tumours. Nat. Rev. Cancer 2017, 17, 337–351. [Google Scholar] [CrossRef]
- Randi, A.M.; Sperone, A.; Dryden, N.H.; Birdsey, G.M. Regulation of angiogenesis by ETS transcription factors. Biochem. Soc. Trans. 2009, 37, 1248–1253. [Google Scholar] [CrossRef]
- Oettgen, P. The role of ets factors in tumor angiogenesis. J. Oncol. 2010, 2010, 767384. [Google Scholar] [CrossRef]
- Aldape, K.; Zadeh, G.; Mansouri, S.; Reifenberger, G.; von Deimling, A. Glioblastoma: Pathology, molecular mechanisms and markers. Acta Neuropathol. 2015, 129, 829–848. [Google Scholar] [CrossRef]
- Yan, H.; Parsons, D.W.; Jin, G.; McLendon, R.; Rasheed, B.A.; Yuan, W.; Kos, I.; Batinic-Haberle, I.; Jones, S.; Riggins, G.J.; et al. IDH1 and IDH2 mutations in gliomas. N. Engl. J. Med. 2009, 360, 765–773. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro-Oncology 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/27141961/ (accessed on 27 July 2020).
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.C.; Kuo, F.; Fujiwara, Y.; Gilliland, D.G.; Golub, T.R.; Orkin, S.H. Yolk sac angiogenic defect and intra-embryonic apoptosis in mice lacking the Ets-related factor TEL. EMBO J. 1997, 16, 4374–4383. [Google Scholar] [CrossRef] [Green Version]
- Hock, H.; Meade, E.; Medeiros, S.; Schindler, J.W.; Valk, P.J.M.; Fujiwara, Y.; Orkin, S.H. Tel/Etv6 is an essential and selective regulator of adult hematopoietic stem cell survival. Genes Dev. 2004, 18, 2336–2341. [Google Scholar] [CrossRef]
- De Braekeleer, E.; Douet-Guilbert, N.; Morel, F.; Le Bris, M.J.; Basinko, A.; De Braekeleer, M. ETV6 fusion genes in hematological malignancies: A review. Leuk. Res. 2012, 36, 945–961. [Google Scholar] [CrossRef]
- Brenca, M.; Rossi, S.; Polano, M.; Gasparotto, D.; Zanatta, L.; Racanelli, D.; Valori, L.; Lamon, S.; Dei Tos, A.P.; Maestro, R. Transcriptome sequencing identifies ETV6-NTRK3 as a gene fusion involved in GIST. J. Pathol. 2016, 238, 543–549. [Google Scholar] [CrossRef]
- Kibel, A.S.; Faith, D.A.; Bova, G.S.; Isaacs, W.B. Mutational analysis of ETV6 in prostate carcinoma. Prostate 2002, 52, 305–310. [Google Scholar] [CrossRef]
- Milde-Langosch, K.; Bamberger, A.M.; Rieck, G.; Kelp, B.; Löning, T. Overexpression of the p16 cell cycle inhibitor in breast cancer is associated with a more malignant phenotype. Breast Cancer Res. Treat. 2001, 67, 61–70. [Google Scholar] [CrossRef]
- Unnikrishnan, A.; Guan, Y.F.; Huang, Y.; Beck, D.; Thoms, J.A.I.; Peirs, S.; Knezevic, K.; Ma, S.; De Walle, I.V.; De Jong, I.D.E.; et al. A quantitative proteomics approach identifies ETV6 and IKZF1 as new regulators of an ERG-driven transcriptional network. Nucleic Acids Res. 2016, 44, 10644–10661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putnik, J.; Zhang, C.D.; Archangelo, L.F.; Tizazu, B.; Bartels, S.; Kickstein, M.; Greif, P.A.; Bohlander, S.K. The interaction of ETV6 (TEL) and TIP60 requires a functional histone acetyltransferase domain in TIP60. Biochim. Biophys. Acta—Mol. Basis Dis. 2007, 1772, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Noorani, I.; Bradley, A.; De La Rosa, J. CRISPR and transposon in vivo screens for cancer drivers and therapeutic targets. Genome Biol. 2020, 21, 204. [Google Scholar] [CrossRef] [PubMed]
- Chow, R.D.; Guzman, C.D.; Wang, G.; Schmidt, F.; Youngblood, M.W.; Ye, L.; Errami, Y.; Dong, M.B.; Martinez, M.A.; Zhang, S.; et al. AAV-mediated direct in vivo CRISPR screen identifies functional suppressors in glioblastoma. Nat. Neurosci. 2017, 20, 1329–1341. [Google Scholar] [CrossRef] [Green Version]
- Noorani, I.; De La Rosa, J.; Choi, Y.; Strong, A.; Ponstingl, H.; Vijayabaskar, M.S.; Lee, J.; Lee, E.; Richard-Londt, A.; Friedrich, M.; et al. PiggyBac mutagenesis and exome sequencing identify genetic driver landscapes and potential therapeutic targets of EGFR-mutant gliomas. Genome Biol. 2020, 21, 181. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Chi, H.T.; Ly, B.T.K.; Kano, Y.; Tojo, A.; Watanabe, T.; Sato, Y. ETV6-NTRK3 as a therapeutic target of small molecule inhibitor PKC412. Biochem. Biophys. Res. Commun. 2012, 429, 87–92. [Google Scholar] [CrossRef]
- Roberts, K.G.; Li, Y.; Payne-Turner, D.; Harvey, R.C.; Yang, Y.-L.; Pei, D.; McCastlain, K.; Ding, L.; Lu, C.; Song, G.; et al. Targetable kinase-activating lesions in Ph-like acute lymphoblastic leukemia. N. Engl. J. Med. 2014, 371, 1005–1015. [Google Scholar] [CrossRef] [Green Version]
- Taipale, M.; Krykbaeva, I.; Whitesell, L.; Santagata, S.; Zhang, J.; Liu, Q.; Gray, N.S.; Lindquist, S. Chaperones as thermodynamic sensors of drug-target interactions reveal kinase inhibitor specificities in living cells. Nat. Biotechnol. 2013, 31, 630–637. [Google Scholar] [CrossRef] [Green Version]
- Roberts, K.G.; Janke, L.J.; Zhao, Y.; Seth, A.; Ma, J.; Finkelstein, D.; Smith, S.; Ebata, K.; Tuch, B.B.; Hunger, S.P.; et al. Etv6-ntrk3 induces aggressive acute lymphoblastic leukemia highly sensitive to selective trk inhibition. Blood 2018, 132, 861–865. [Google Scholar] [CrossRef] [Green Version]
- Shukla, N.; Roberts, S.S.; Baki, M.O.; Mushtaq, Q.; Goss, P.E.; Park, B.H.; Gundem, G.; Tian, K.; Geiger, H.; Redfield, K.; et al. Successful Targeted Therapy of Refractory Pediatric ETV6-NTRK3 Fusion-Positive Secretory Breast Carcinoma. JCO Precis. Oncol. 2017, 1, PO.17.00034. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Z.; Wu, S.; Li, F.-j.; Luo, C.; Jin, Q.-y.; Connolly, I.D.; Hayden Gephart, M.; You, L. Elevated ETV6 Expression in Glioma Promotes an Aggressive In Vitro Phenotype Associated with Shorter Patient Survival. Genes 2022, 13, 1882. https://doi.org/10.3390/genes13101882
Xiong Z, Wu S, Li F-j, Luo C, Jin Q-y, Connolly ID, Hayden Gephart M, You L. Elevated ETV6 Expression in Glioma Promotes an Aggressive In Vitro Phenotype Associated with Shorter Patient Survival. Genes. 2022; 13(10):1882. https://doi.org/10.3390/genes13101882
Chicago/Turabian StyleXiong, Zhang, Shuai Wu, Feng-jiao Li, Chen Luo, Qiu-yan Jin, Ian David Connolly, Melanie Hayden Gephart, and Linya You. 2022. "Elevated ETV6 Expression in Glioma Promotes an Aggressive In Vitro Phenotype Associated with Shorter Patient Survival" Genes 13, no. 10: 1882. https://doi.org/10.3390/genes13101882