Identification of Differentially Expressed Genes and Pathways for Abdominal Fat Deposition in Ovariectomized and Sham-Operated Chickens

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Treatments
2.3. RNA Extraction and Identification
2.4. Gene Expression Profiling and Data Analysis
2.5. qRT-PCR Analysis
2.6. Statistical Analysis
3. Results
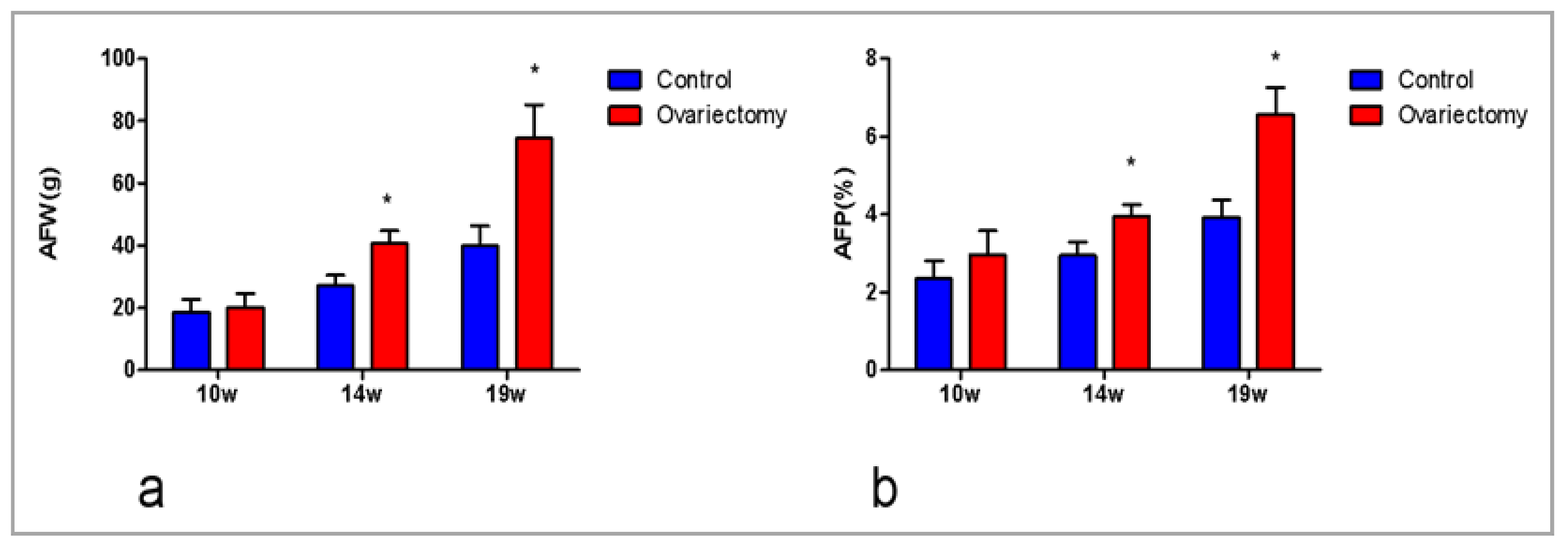
3.1. Ovariectomy Accelerates AF Deposition
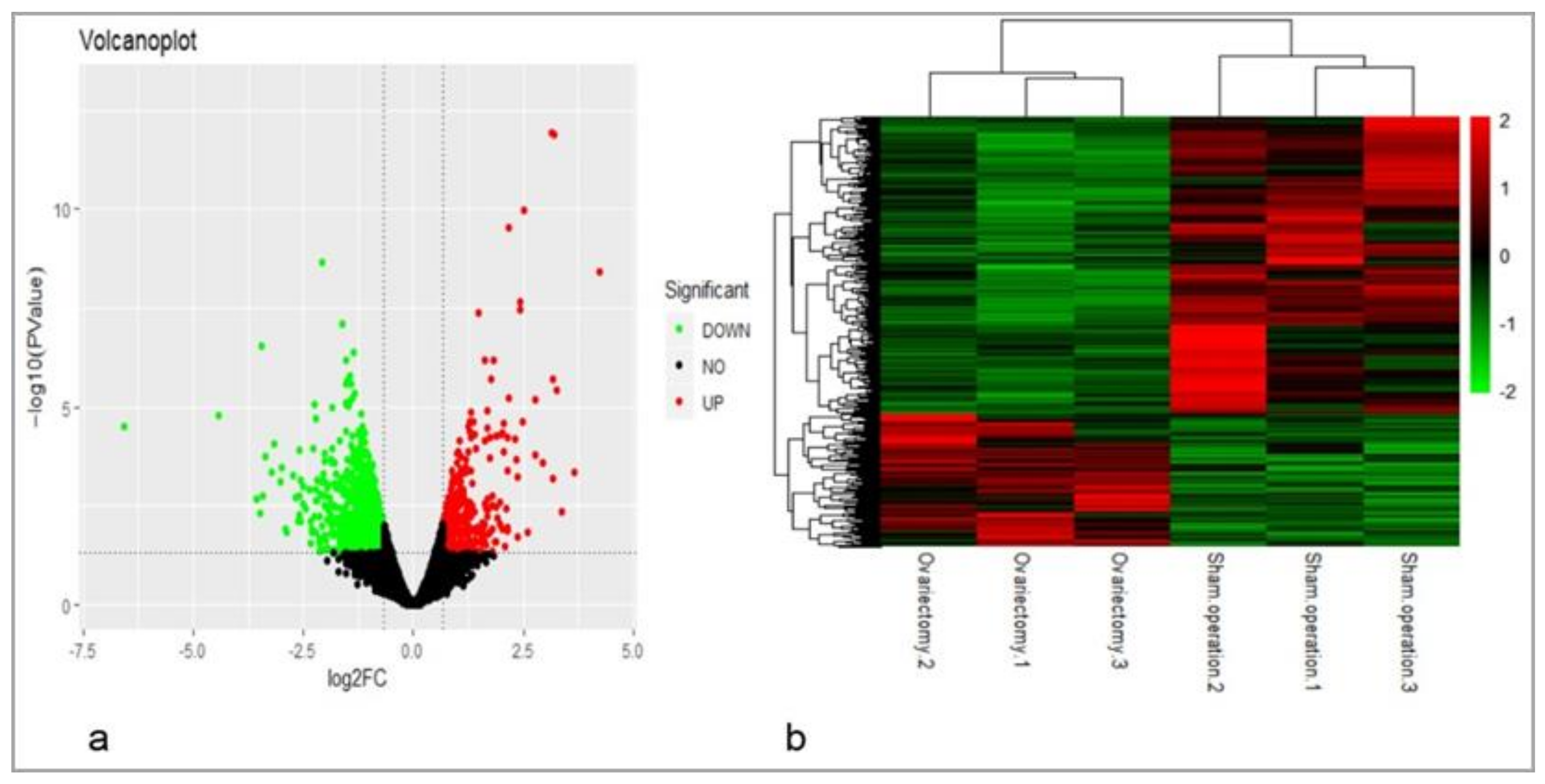
3.2. Identification of DEGs Related to AF Metabolism After Ovariectomy
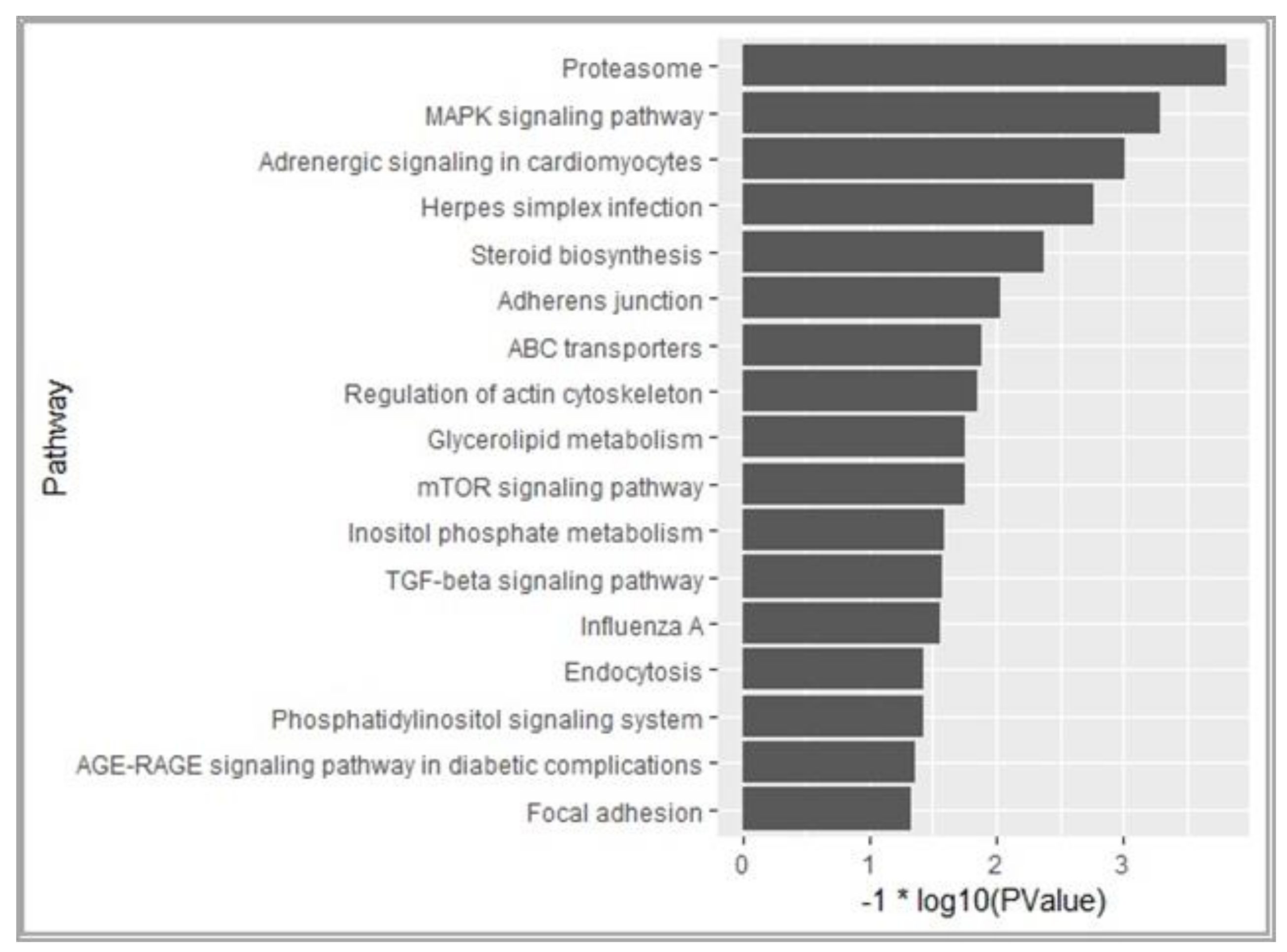
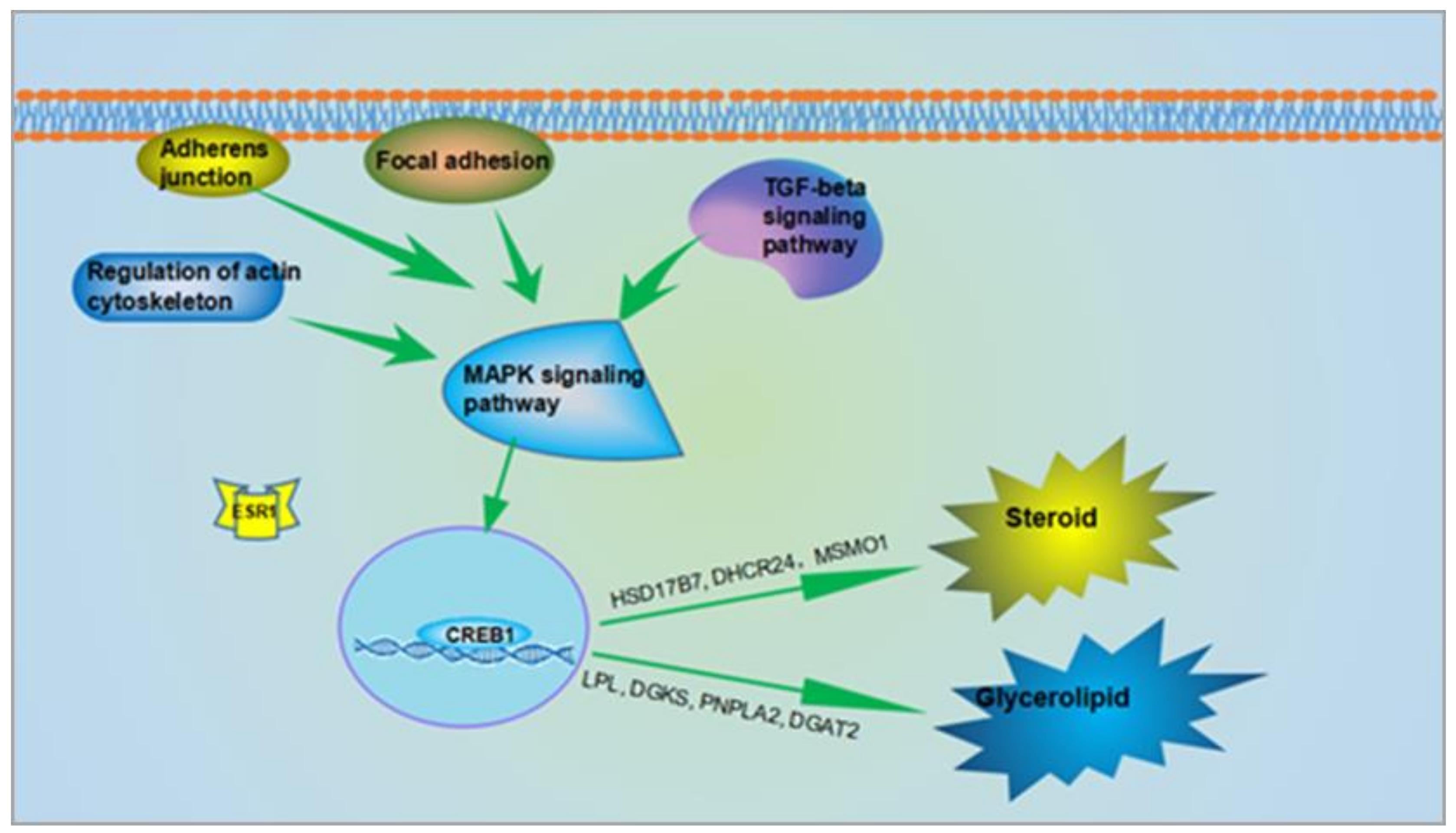
3.3. Pathways and Regulatory Network for AF in Chickens After Ovariectomy
4. Discussion
4.1. Differences in AF Deposition between the Ovariectomized and Sham-Operated Groups
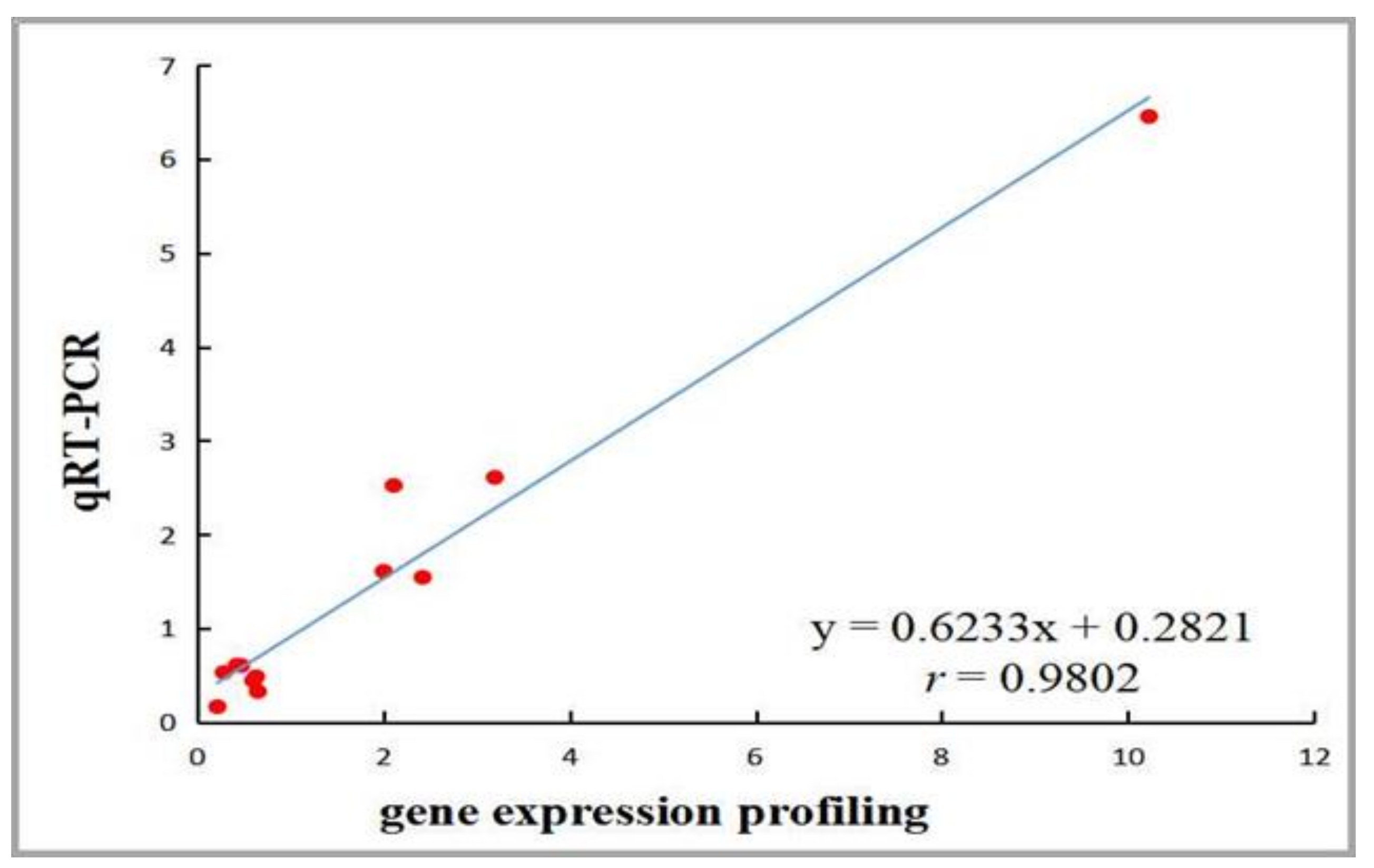
4.2. The Validation of Gene Expression Profiling and Identification of DEGs Related to AF Deposition
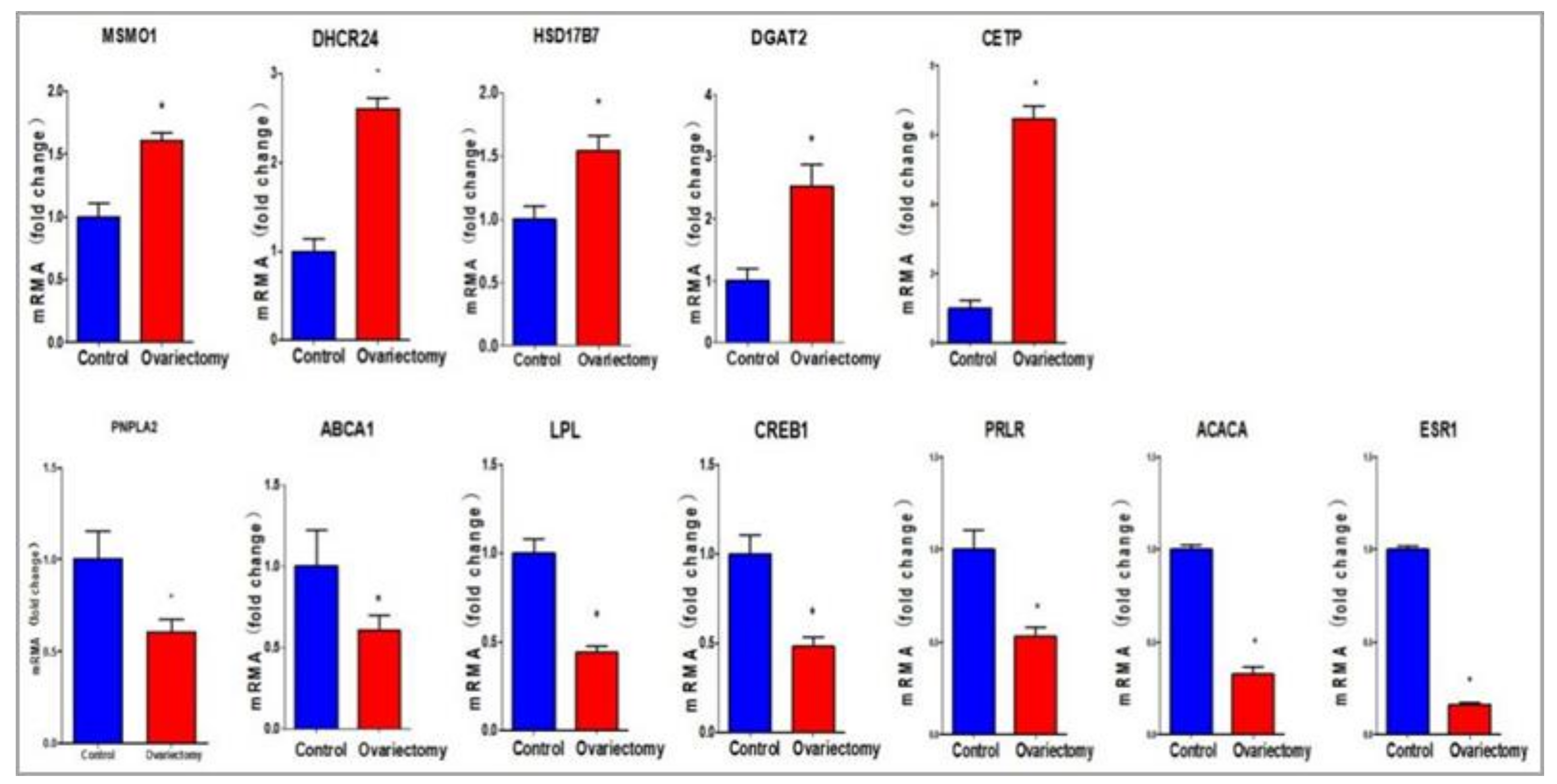
4.3. Key Genes Related to the Metabolism of AF
4.4. Signaling Pathways Related to Lipid Metabolism in AF after Ovariectomy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Amorim, A.; Rodrigues, S.; Pereira, E.; Valentim, R.; Teixeira, A. Effect of caponisation on physicochemical and sensory characteristics of chickens. Animal 2016, 10, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Calik, J.; Połtowicz, K.; Światkiewicz, S.; Krawczyk, J.; Nowak, J. Effect of caponization on meat quality of Greenleg Partridge cockerels. Ann. Anim. Sci. 2015, 15, 541–553. [Google Scholar] [CrossRef]
- Cui, X.; Cui, H.; Liu, L.; Zhao, G.; Liu, R.; Li, Q.; Zheng, M.; Wen, J. Decreased testosterone levels after caponization leads to abdominal fat deposition in chickens. BMC Genom. 2018, 19, 344. [Google Scholar] [CrossRef] [PubMed]
- Becker, W.A.; Spencer, J.V.; Mirosh, L.W.; Verstrate, J.A. Prediction of fat and fat free live weight in broiler chickens using backskin fat, abdominal fat, and live body weight. Poult. Sci. 1979, 58, 835–842. [Google Scholar] [CrossRef]
- Thomas, V.G.; Mainguy, S.K.; Prevett, J.P. Predicting fat content of geese from abdominal fat weight. J. Wildl. Manag. 1983, 47, 1115–1119. [Google Scholar] [CrossRef]
- Butterwith, S.C. Contribution of lipoprotein lipase activity to the differential growth of three adipose tissue depots in young broiler chickens. Br. Poult. Sci. 1989, 30, 927–933. [Google Scholar] [CrossRef]
- Resnyk, C.W.; Carré, W.; Wang, X.; Porter, T.E.; Simon, J.; Bihan-Duval, E.L.; Duclos, M.J.; Aggrey, S.E.; Cogburn, L.A. Transcriptional analysis of abdominal fat in chickens divergently selected on bodyweight at two ages reveals novel mechanisms controlling adiposity: Validating visceral adipose tissue as a dynamic endocrine and metabolic organ. BMC Genom. 2017, 18, 626. [Google Scholar] [CrossRef]
- Leclercq, B. Adipose tissue metabolism and its control in birds. Poult. Sci. 1984, 63, 2044–2054. [Google Scholar] [CrossRef]
- Leveille, G.A. In vitro hepatic lipogenesis in the hen and chick. Comp. Biochem. Physiol. 1969, 28, 431–435. [Google Scholar] [CrossRef]
- Song, Y.; Silversides, F.G. The technique of orthotopic ovarian transplantation in the chicken. Poult. Sci. 2006, 85, 1104–1106. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]
- Wu, J.; Mao, X.; Tao, C.; Luo, J.; Wei, L. KOBAS server: A web-based platform for automated annotation and pathway identification. Nucleic Acids Res. 2006, 34, W720–W724. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 −ΔΔ C T Method. Methods Companion Methods Enzymol. 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, Y.; Fu, M.; Li, G.; An, N.; Li, S.; Zhou, Z. The effects of ovariectomy on meat performance and expression of GH/IGF-I in young goats. Can. J. Anim. Sci. 2014, 94, 619–626. [Google Scholar] [CrossRef]
- Meli, R.; Pacilio, M.; Raso, G.M.; Esposito, E.; Coppola, A.; Nasti, A.; Di Carlo, C.; Nappi, C.; Di Carlo, R. Estrogen and raloxifene modulate leptin and its receptor in hypothalamus and adipose tissue from ovariectomized rats. Endocrinology 2004, 145, 3115–3121. [Google Scholar] [CrossRef]
- Babaei, P.; Dastras, A.; Tehrani, B.S.; Roudbaneh, S.P. The effect of estrogen replacement therapy on visceral fat, serum glucose, lipid profiles and apelin level in ovariectomized rats. J. Menopausal Med. 2017, 23, 182–189. [Google Scholar] [CrossRef]
- Wang, J.F.; Guo, Y.X.; Niu, J.Z.; Liu, J.; Wang, L.Q.; Li, P.H. Effects of Radix Puerariae flavones on liver lipid metabolism in ovariectomized rats. World J. Gastroenterol. 2004, 10, 1967–1970. [Google Scholar] [CrossRef]
- Lee, J.; Park, Y.; Koo, S.I. ATP-binding cassette transporter A1 and HDL metabolism: Effects of fatty acids. J. Nutr. Biochem. 2012, 23, 1–7. [Google Scholar] [CrossRef]
- Liya, Y.; Yanqiao, Z.; F Bradley, H. Sterol regulatory element-binding protein-1 interacts with the nuclear thyroid hormone receptor to enhance acetyl-CoA carboxylase-α transcription in hepatocytes. J. Biol. Chem. 2002, 277, 19554–19565. [Google Scholar]
- Aaltonen, T.; Adelman, J.; Akimoto, T.; Albrow, M.G.; Alvarez, G.L.B.; Amerio, S.; Amidei, D.; Anastassov, A.; Annovi, A.; Antos, J. First measurement of W boson production in association with a single Charm quark in pp collisions at sqrt(s) = 1.96 TeV. Phys. Rev. Lett. 2008, 100, 091803. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, I.J.; Merkel, M. Lipoprotein lipase: Physiology, biochemistry, and molecular biology. Front. Biosci. 2001, 6, D388–D405. [Google Scholar] [PubMed]
- Suzuki, A.; Yamaguchi, S.; Li, M.; Hara, Y.; Miyauchi, H.; Ikeda, Y.; Zhang, B.; Higashi, M.; Ikeda, Y.; Takagi, A.; et al. Tricaprin rescues myocardial abnormality in a mouse model of triglyceride deposit cardiomyovasculopathy. J. Oleo Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D.; Iichiro, S.; Shinji, I.; Yuriy, B.; Hammer, R.E. Overexpression of sterol regulatory element-binding protein-1a in mouse adipose tissue produces adipocyte hypertrophy, increased fatty acid secretion, and fatty liver. J. Biol. Chem. 2003, 278, 36652–36660. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.; Dahlman, I.; Rydén, M.; Nordström, E.A.; Gustafsson, J.A.; Arner, P.; Dahlman-Wright, K. Oestrogen receptor α gene expression levels are reduced in obese compared to normal weight females. Int. J. Obes. 2007, 31, 900. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Bentley, A.; Adeyemo, A.; Shriner, D.; Zhou, J.; Doumatey, A.; Huang, H.; Ramos, E.; Erdos, M.; Gerry, N. Genome-wide association study identifies novel loci association with fasting insulin and insulin resistance in African Americans. Hum. Mol. Genet. 2012, 21, 4530–4536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Dollé, M.E.T.; Imholz, S.; Slot, R.V.T.; Verschuren, W.M.M.; Wijmenga, C.; Feskens, E.J.M.; Boer, J.M.A. Multiple genetic variants along candidate pathways influence plasma high-density lipoprotein cholesterol concentrations. J. Lipid Res. 2008, 49, 2582–2589. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, D.R.; Keeling, J.W.; Evans, M.J.; Kan, A.E.; Bell, J.E.; Porteous, M.E.; Mills, K.; Winter, R.M.; Clayton, P.T. Clinical phenotype of desmosterolosis. Am. J. Med. Genet. Part A 2010, 75, 145–152. [Google Scholar] [CrossRef]
- Laubner, D.; MÖller, G.; Marijanovic, Z.; Adamski, J.; Gege, C.; Husen, B.; Breitling, R. Closing the gap: identification of human 3-ketosteroid reductase, the last unknown enzyme of mammalian cholesterol biosynthesis. Mol. Endocrinol. 2003, 17, 1715–1725. [Google Scholar]
- Cases, S.; Smith, S.J.; Zheng, Y.W.; Myers, H.M.; Lear, S.R.; Sande, E.; Novak, S.; Collins, C.; Welch, C.B.; Lusis, A.J. Identification of a gene encoding an acyl CoA:diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 13018–13023. [Google Scholar] [CrossRef] [Green Version]
- Mun Ock, K.; Ui, L.S.; Hyun-Jun, L.; Kwangman, C.; Hyeongki, K.; Sangku, L.; Soo Jin, O.; Sunhong, K.; Jong Soon, K.; Hyun Sun, L. Identification and validation of a selective small molecule inhibitor targeting the diacylglycerol acyltransferase 2 activity. Biol. Pharm. Bull. 2013, 36, 1167–1173. [Google Scholar]
- Cui, H.-X.; Liu, R.-R.; Zhao, G.-P.; Zheng, M.-Q.; Chen, J.-L.; Wen, J. Identification of differentially expressed genes and pathways for intramuscular fat deposition in pectoralis major tissues of fast-and slow-growing chickens. BMC Genom. 2012, 13, 213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Reue, K. Lipin proteins and glycerolipid metabolism: Roles at the ER membrane and beyond. Biochim. Biophys. Acta (BBA)-Biomembr. 2017, 1859, 1583–1595. [Google Scholar] [CrossRef] [PubMed]
- Chibalin, A.V.; Leng, Y.; Vieira, E.; Krook, A.; Björnholm, M.; Long, Y.C.; Kotova, O.; Zhong, Z.; Sakane, F.; Steiler, T.; et al. Downregulation of diacylglycerol kinase delta contributes to hyperglycemia-induced insulin resistance. Cell 2008, 132, 375–386. [Google Scholar] [CrossRef]
- Shulga, Y.V.; Loukov, D.; Ivanova, P.T.; Milne, S.B.; Myers, D.S.; Hatch, G.M.; Umeh, G.; Jalan, D.; Fullerton, M.D.; Steinberg, G.R. Diacylglycerol kinase delta promotes lipogenesis. Biochemistry 2013, 52, 7766–7776. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.; Guo, B.; Chatterjee, S.; Chen, M.H.; Nelson, D.; Yechoor, V.K.; Ma, K. The adipocyte clock controls brown adipogenesis via TGF-β/BMP signaling pathway. J. Cell Sci. 2015, 128, 1835–1847. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Song, T.J.; Li, X.; Hu, L.; He, Q.; Liu, M.; Lane, M.D.; Tang, Q.Q. BMP signaling pathway is required for commitment of C3H10T1/2 pluripotent stem cells to the adipocyte lineage. Proc. Natl. Acad. Sci. USA 2009, 106, 12670–12675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, T.; Sengupta, S.; Harris, T.; Carmack, A.; Kang, S.; Balderas, E.; Guertin, D.; Madden, K.; Carpenter, A.; Finck, B. mTOR complex 1 regulates lipin 1 localization to control the SREBP pathway. Cell 2011, 146, 408–420. [Google Scholar] [CrossRef]
- Martin, G.R. The roles of FGFs in the early development of vertebrate limbs. Genes Dev. 1998, 12, 1571–1586. [Google Scholar] [CrossRef] [Green Version]
- Chiou, M.J.; Wang, Y.D.; Kuo, C.M.; Chen, J.C.; Chen, J.Y. Functional analysis of mitogen-activated protein kinase-3 (MAPK3) and its regulation of the promoter region in zebrafish. DNA Cell Biol. 2007, 26, 781–790. [Google Scholar] [CrossRef]
- Syed Kashif, Z.; Wen-Jun, S.; Stefanie, B.; Alex, B.; Mclean, M.P.; Jiahuai, H.; Davis, R.J.; Kraemer, F.B.; Salman, A. p38 MAPK regulates steroidogenesis through transcriptional repression of STAR gene. J. Mol. Endocrinol. 2014, 53, 1–16. [Google Scholar] [Green Version]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, X.; Cui, X.; Liu, R.; Li, Q.; Zheng, M.; Zhao, G.; Ge, C.; Wen, J.; Hu, Y.; Cui, H. Identification of Differentially Expressed Genes and Pathways for Abdominal Fat Deposition in Ovariectomized and Sham-Operated Chickens. Genes 2019, 10, 155. https://doi.org/10.3390/genes10020155
Mu X, Cui X, Liu R, Li Q, Zheng M, Zhao G, Ge C, Wen J, Hu Y, Cui H. Identification of Differentially Expressed Genes and Pathways for Abdominal Fat Deposition in Ovariectomized and Sham-Operated Chickens. Genes. 2019; 10(2):155. https://doi.org/10.3390/genes10020155
Chicago/Turabian StyleMu, Xiaopeng, Xiaoyan Cui, Ranran Liu, Qinghe Li, Maiqing Zheng, Guiping Zhao, Changrong Ge, Jie Wen, Yaodong Hu, and Huanxian Cui. 2019. "Identification of Differentially Expressed Genes and Pathways for Abdominal Fat Deposition in Ovariectomized and Sham-Operated Chickens" Genes 10, no. 2: 155. https://doi.org/10.3390/genes10020155