Targeting the JAK/STAT Signaling Pathway Using Phytocompounds for Cancer Prevention and Therapy

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Literature Search Methodology
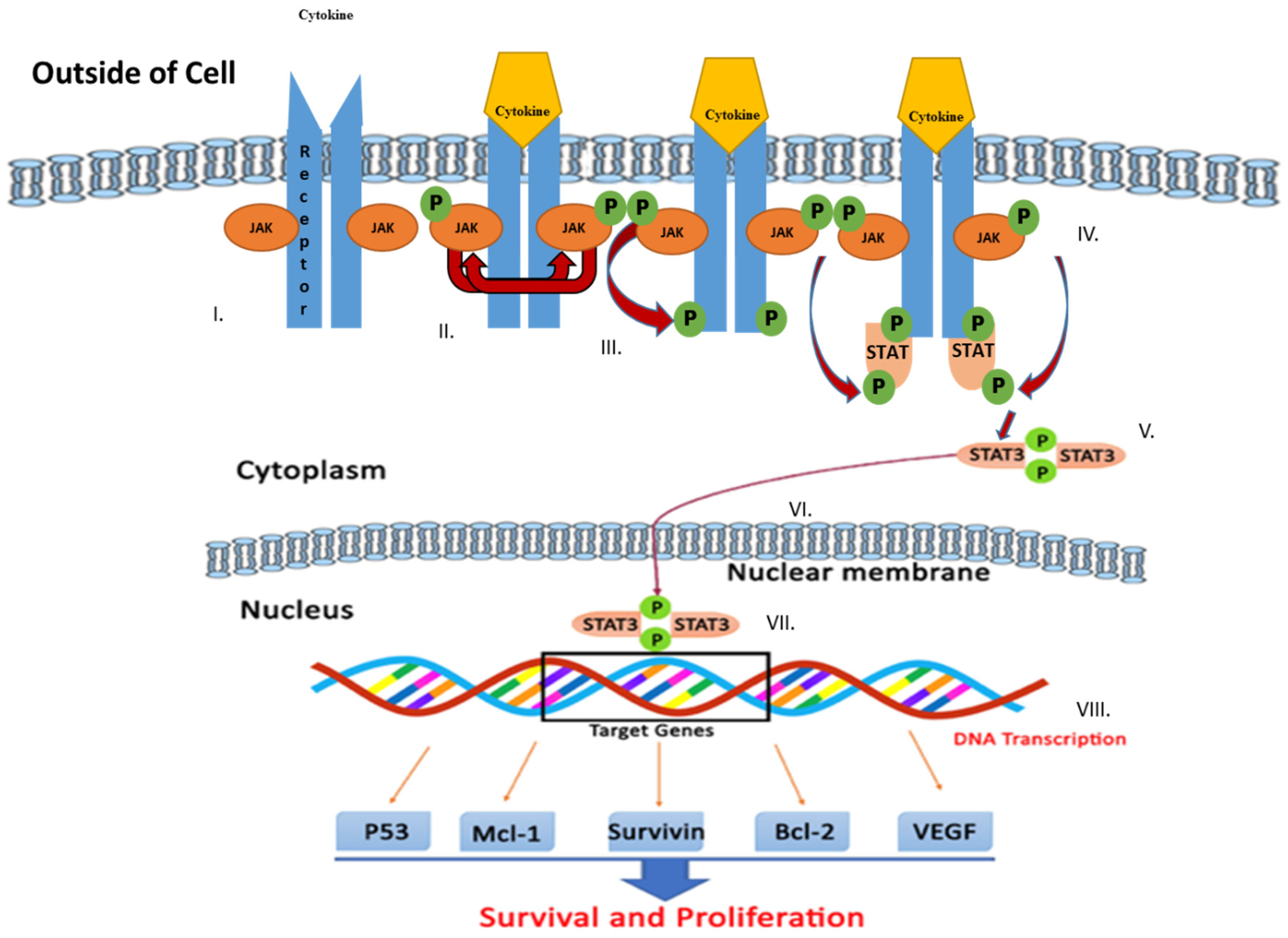
3. JAK/STAT Signaling Pathway
4. JAK/STAT Inhibitors
4.1. JAK Inhibitors
4.2. STAT Inhibitors
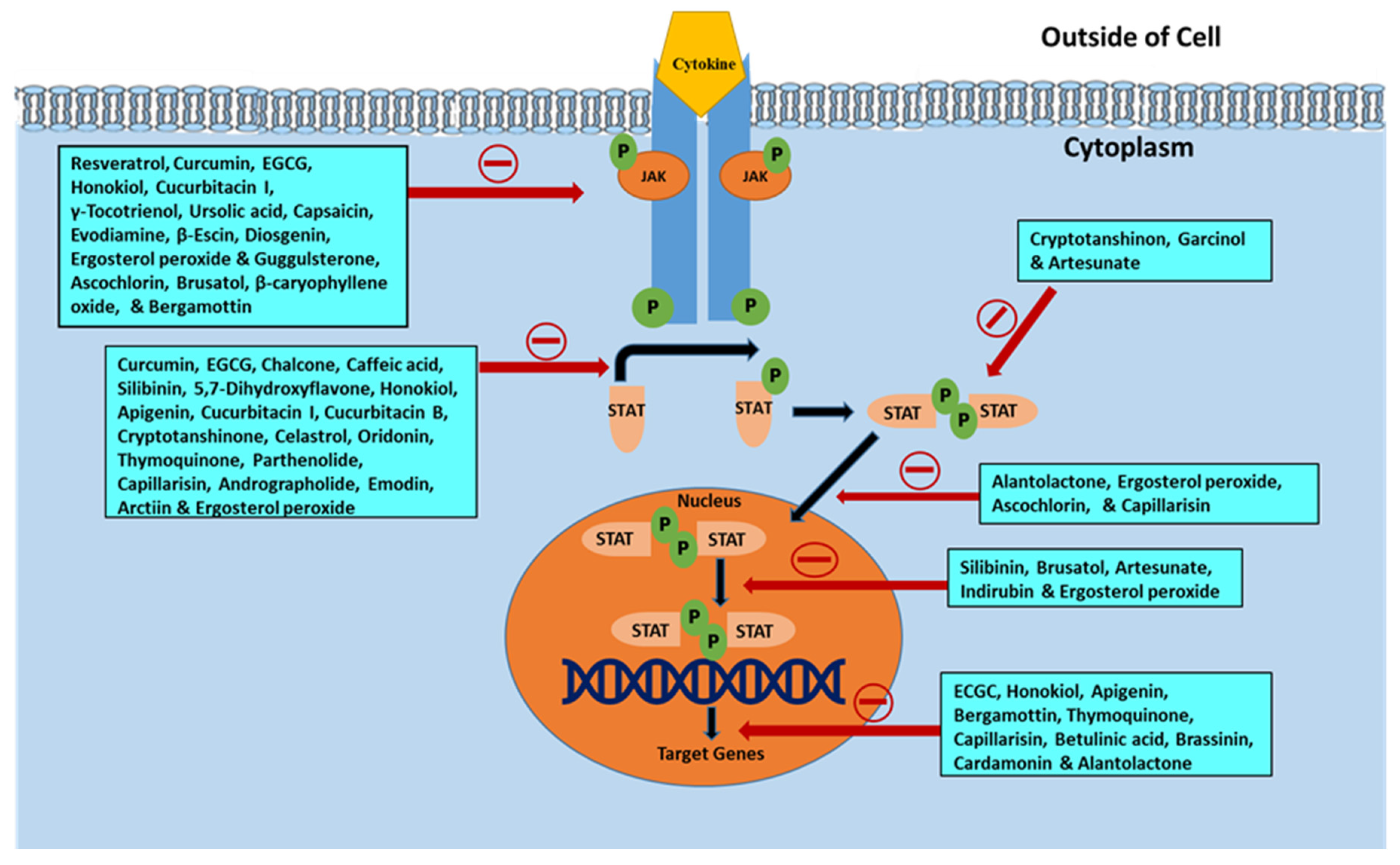
5. Phytochemicals Inhibiting the JAK/STAT Signaling Pathway
5.1. Phenolics and Polyphenols
5.1.1. Resveratrol
5.1.2. Curcumin
5.1.3. Ascochlorin
5.1.4. Bergamottin
5.1.5. Capillarisin
5.1.6. Bavachin
5.1.7. Epigallocatechin Gallate (EGCG)
5.1.8. Emodin
5.1.9. Chalcones
5.1.10. Formononetin
5.1.11. Garcinol
5.1.12. Cardamonin
5.1.13. Caffeic Acid
5.1.14. Silibinin
5.1.15. Butein
5.1.16. 5,7-Dihydroxyflavone
5.1.17. Honokiol
5.1.18. Casticin
5.1.19. Apigenin
5.1.20. Wedelolactone
5.2. Terpenoids
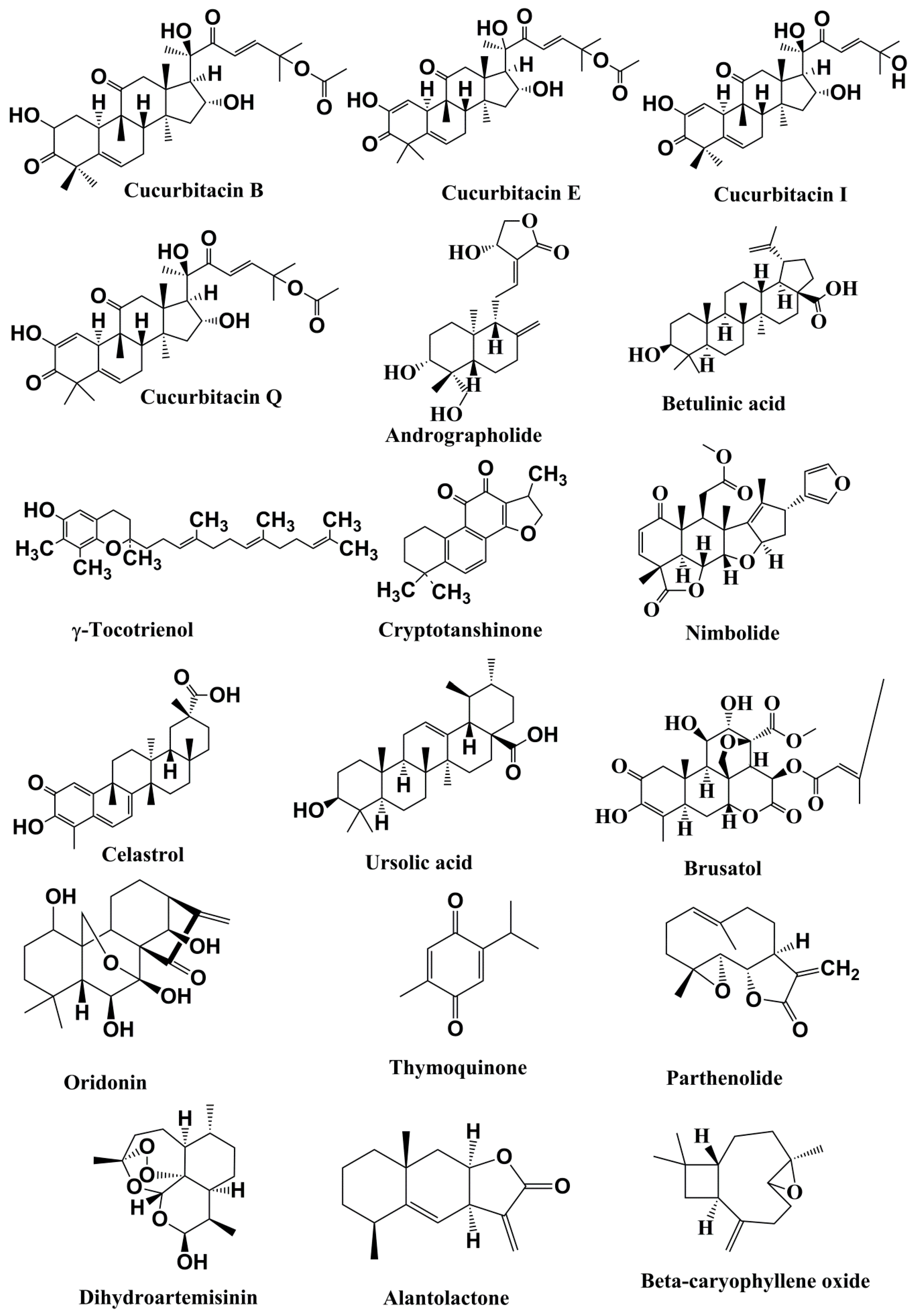
5.2.1. Cucurbitacins
5.2.2. Andrographolide
5.2.3. Betulinic Acid
5.2.4. γ-Tocotrienol
5.2.5. Cryptotanshinone
5.2.6. Nimbolide
5.2.7. Celastrol
5.2.8. Ursolic Acid
5.2.9. Brusatol
5.2.10. Oridonin
5.2.11. Thymoquinone
5.2.12. Parthenolide
5.2.13. Dihydroartemisinin
5.2.14. Alantolacton
5.2.15. β-Caryophyllene oxide
5.3. Alkaloids
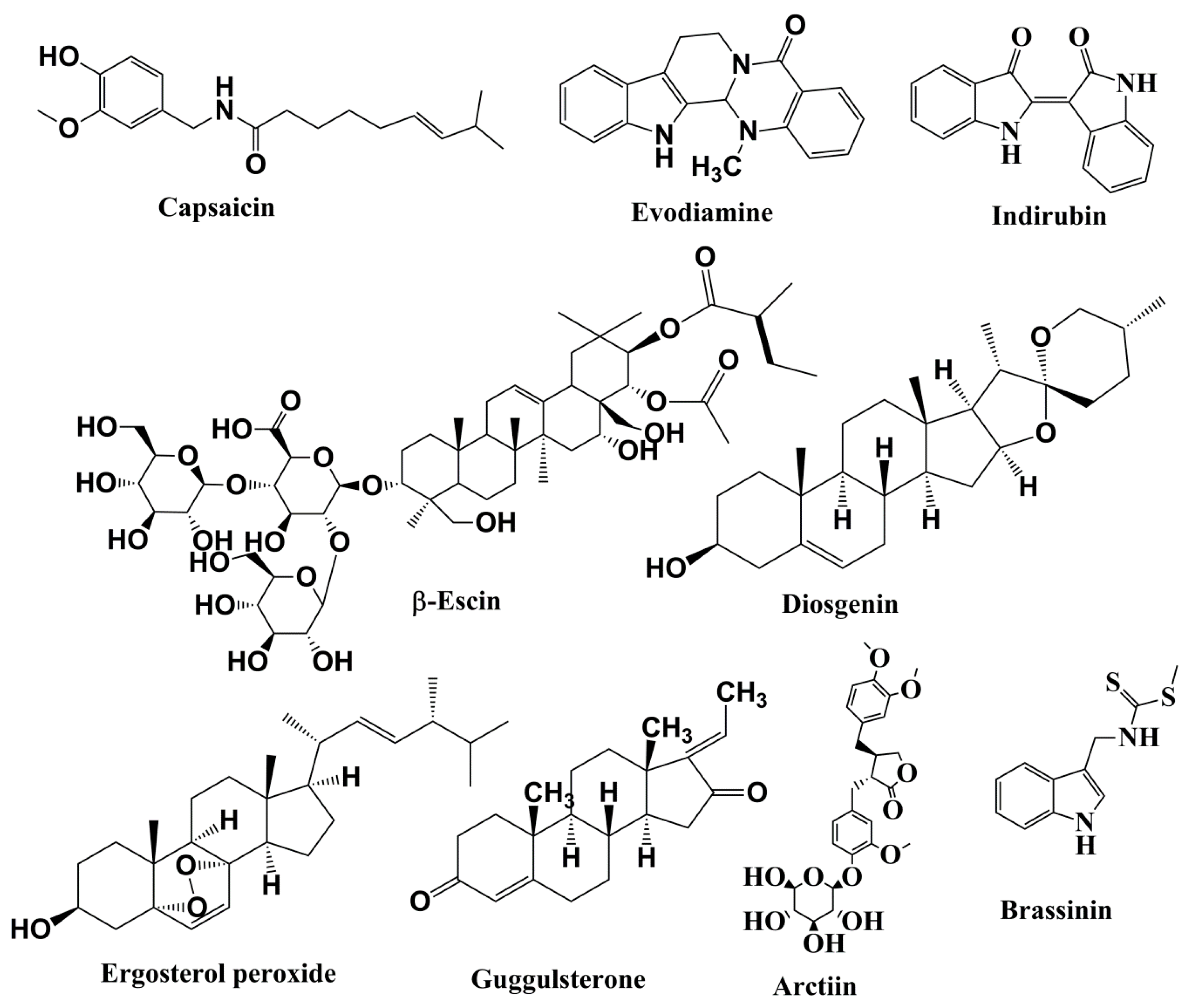
5.3.1. Capsaicin
5.3.2. Evodiamine
5.3.3. Indirubin
5.4. Saponins
β-Escin
5.5. Steroids
5.5.1. Diosgenin
5.5.2. Ergosterol Peroxide
5.5.3. Guggulsterone
5.6. Lignan
Arctiin
5.7. Phytoalexin
Brassinin
6. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Murray, T. Cancer statistics. CA Cancer J. Clin. 2008, 58, 71–96. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer statistics. CA Cancer J. Clin. 2010, 60, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Wiart, C. Goniothalamus species: A source of drugs for the treatment of cancers and bacterial infections. Evid. Based Complement. Alternat. Med. 2007, 4, 299–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Glade, M.J. Food, nutrition, and the prevention of cancer: A global perspective. American Institute for Cancer Research/World Cancer Research Fund, American Institute for Cancer Research. Nutrition 1999, 15, 523–526. [Google Scholar]
- Sporn, M.B.; Liby, K.T. NRF2 and cancer: The good, the bad and the importance of context. Nat. Rev. Cancer 2012, 12, 564–571. [Google Scholar] [CrossRef]
- Manson, M.M. Cancer prevention-the potential for diet to modulate molecular signalling. Trends Mol. Med. 2003, 9, 11–18. [Google Scholar] [CrossRef]
- Gullett, N.P.; Ruhul Amin, A.R.; Bayraktar, S.; Pezzuto, J.M.; Shin, D.M.; Khuri, F.R.; Aggarwal, B.B.; Surh, Y.J.; Kucuk, O. Cancer prevention with natural compounds. Semin. Oncol. 2010, 37, 258–281. [Google Scholar] [CrossRef]
- Block, K.I.; Gyllenhaal, C.; Lowe, L.; Amedei, A.; Amin, A.R.M.R.; Amin, A.; Aquilano, K.; Arbiser, J.; Arreola, A.; Arzumanyan, A.; et al. Designing a broad-spectrum integrative approach for cancer prevention and treatment. Semin. Cancer Biol. 2015, 35, 276–304. [Google Scholar] [CrossRef]
- Bishayee, A.; Sethi, G. Bioactive natural products in cancer prevention and therapy: Progress and promise. Semin. Cancer Biol. 2016, 40–41, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Kotecha, R.; Takami, A.; Espinoza, J.L. Dietary phytochemicals and cancer chemoprevention: A review of the clinical evidence. Oncotarget 2016, 7, 52517–52529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christen, P.; Cuendet, M. Plants as a source of therapeutic and health products. Chimia 2012, 66, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Geldenhuys, W.J.; Bishayee, A.; Darvesh, A.S.; Carroll, R.T. Natural products of dietary origin as lead compounds in virtual screening and drug design. Curr. Pharm. Biotechnol. 2012, 13, 117–124. [Google Scholar] [CrossRef]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef]
- McCullough, M.L.; Giovannucci, E.L. Diet and cancer prevention. Oncogene 2004, 23, 6349–6364. [Google Scholar] [CrossRef] [Green Version]
- Russo, G.L. Ins and outs of dietary phytochemicals in cancer chemoprevention. Biochem. Pharmacol. 2007, 74, 533–544. [Google Scholar] [CrossRef]
- Amin, A.R.; Kucuk, O.; Khuri, F.R.; Shin, D.M. Perspectives for cancer prevention with natural compounds. J. Clin. Oncol. 2009, 27, 2712–2725. [Google Scholar] [CrossRef] [Green Version]
- Meadows, G.G. Diet, nutrients, phytochemicals, and cancer metastasis suppressor genes. Cancer Metastasis Rev. 2012, 31, 441–454. [Google Scholar] [CrossRef] [Green Version]
- Fernando, W.; Rupasinghe, H.P.V. Anticancer properties of phytochemicals present in medicinal plants of North America. In Using Old Solutions to New Problems—Natural Drug Discovery in the 21st Century, 1st ed.; Kulka, M., Ed.; InTech: London, UK, 2013. [Google Scholar]
- Patra, J.K.; Das, G.; Bose, S.; Banerjee, S.; Vishnuprasad, C.N.; del Pilar Rodriguez-Torres, M.; Shin, H.S. Star anise (Illicium verum): Chemical compounds, antiviral properties, and clinical relevance. Phytother. Res. 2020, 2020, 1–20. [Google Scholar] [CrossRef]
- Bowman, T.; Garcia, R.; Turkson, J.; Jove, R. STATs in oncogenesis. Oncogene 2000, 19, 2474–2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grivennikov, S.I.; Karin, M. Dangerous liaisons: STAT3 and NFkappaB collaboration and crosstalk in cancer. Cytokine Growth Factor Rev. 2010, 21, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch-Barrera, J.; Queralt, B.; Menendez, J.A. Targeting STAT3 with silibinin to improve cancer therapeutics. Cancer Treat. Rev. 2017, 58, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Ozbey, U.; Attar, R.; Romero, M.A.; Alhewairini, S.S.; Afshar, B.; Sabitaliyevich, U.Y.; Hanna-Wakim, L.; Ozcelik, B.; Farooqi, A.A. Apigenin as an effective anticancer natural product: Spotlight on TRAIL, WNT/-catenin, JAK-STAT pathways, and microRNAs. J. Cell Biochem. 2018, 120, 1060–1067. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Wang, L.; Goh, B.C.; Ahn, K.S.; Sethi, G. Modulation of diverse oncogenic transcription factors by thymoquinone, an essential oil compound isolated from the seeds of Nigella sativa Linn. Pharmacol. Res. 2018, 129, 357–364. [Google Scholar] [CrossRef]
- Siveen, K.S.; Sikka, S.; Surana, R.; Dai, X.; Zhang, J.; Kumar, A.P.; Tan, B.K.; Sethi, G.; Bishayee, A. Targeting the STAT3 signaling pathway in cancer: Role of synthetic and natural inhibitors. Biochim. Biophys. Acta 2014, 1845, 136–154. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.N.; Singh, H.B.; Singh, A.; Naqvi, A.H.; Singh, B.R. Dietary phytochemicals alter epigenetic events and signaling pathways for inhibition of metastasis cascade. Cancer Metastasis Rev. 2014, 33, 41–85. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.P.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Mustafa, M.R. Cancer prevention and therapy through the modulation of transcription factors by bioactive natural compounds. Semin. Cancer Biol. 2016, 40, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, R.H. Potential mechanisms of action of dietary phytochemicals for cancer prevention by targeting cellular signaling transduction pathways. J. Agric. Food Chem. 2018, 66, 3260–3276. [Google Scholar] [CrossRef]
- Rajagopal, C.; Lankadasari, M.B.; Aranjani, J.M.; Harikumar, K.B. Targeting oncogenic transcription factors by polyphenols: A novel approach for cancer therapy. Pharmacol. Res. 2018, 130, 273–291. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. PLoS Med. 2009, 6, e1000100. [Google Scholar] [CrossRef] [PubMed]
- Shuai, K.; Stark, G.R.; Kerr, I.M.; Darnell, J.E. A single phosphotyrosine residue of STAT91 required for gene activation by interferon-gamma. Science 1993, 261, 1744–1746. [Google Scholar] [CrossRef] [PubMed]
- Darnell, J.E.; Kerr, I.M.; Stark, G.R. JAK-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 1994, 264, 1415–1421. [Google Scholar] [CrossRef] [Green Version]
- Yeh, T.C.; Pellegrini, S. The Janus kinase family of protein tyrosine kinases and their role in signaling. Cell. Mol. Life Sci. 1999, 55, 1523–1534. [Google Scholar] [CrossRef]
- Yamaoka, K.; Saharinen, P.; Pesu, M.; Holt, V.E.; Silvennoinen, O.; O’Shea, J.J. The Janus kinases (Jaks). Genome Biol. 2004, 5, 253. [Google Scholar] [CrossRef] [Green Version]
- Shuai, K.; Horvath, C.M.; Huang, L.H.; Qureshi, S.A.; Cowburn, D.; Darnell, J.E. Interferon activation of the transcription factor STAT91 involves dimerization through SH2-phosphotyrosyl peptide interactions. Cell 1994, 76, 821–828. [Google Scholar] [CrossRef]
- Sadowski, H.B.; Shuai, K.; Darnell, J.E.; Gilman, M.Z. A common nuclear signal transduction pathway activated by growth factor and cytokine receptors. Science 1993, 261, 1739–1744. [Google Scholar] [CrossRef] [PubMed]
- Darnell, J.E. Stats and gene regulation. Science 1997, 277, 1630–1635. [Google Scholar] [CrossRef]
- Buettner, R.; Mora, L.B.; Jove, R. Activated STAT signaling in human tumors provides novel molecular targets for therapeutic intervention. Clin. Cancer Res. 2002, 8, 945–954. [Google Scholar]
- Imada, K.; Leonard, W.J. The JAK-STAT pathway. Mol. Immunol. 2000, 37, 1–11. [Google Scholar] [CrossRef]
- Dearolf, C.R. JAKs and STATs in invertebrate model organisms. Cell. Mol. Life Sci. 1999, 55, 1578–1584. [Google Scholar] [CrossRef]
- Ihle, J.N. Cytokine receptor signalling. Nature 1995, 377, 591–594. [Google Scholar] [CrossRef] [PubMed]
- Schindler, C.; Darnell, J.E. Transcriptional responses to polypeptide ligands: The JAK-STAT pathway. Annu. Rev. Biochem. 1995, 64, 621–651. [Google Scholar] [CrossRef] [PubMed]
- Kotenko, S.V.; Pestka, S. JAK-STAT signal transduction pathway through the eyes of cytokine class II receptor complexes. Oncogene 2000, 19, 2557–2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, C.Y.; Arya, A.; Naema, A.F.; Wong, W.F.; Sethi, G.; Looi, C.Y. Signal Transducer and Activator of Transcription (STATs) Proteins in Cancer and Inflammation: Functions and Therapeutic Implication. Front. Oncol. 2019, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meydan, N.; Grunberger, T.; Dadi, H. Inhibition of acute lymphoblastic leukaemia by a Jak-2 inhibitor. Nature 1996, 379, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Samadi, A.K.; Bilsland, A.; Georgakilas, A.G.; Amedei, A.; Amin, A.; Bishayee, A.; Azmi, A.S.; Lokeshwar, B.L.; Grue, B.; Panis, C.; et al. A multi-targeted approach to suppress tumor-promoting inflammation. Semin. Cancer Biol. 2015, 35, 151–184. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Kunnumakkara, A.B.; Harikumar, K.B. Signal transducer and activator of transcription-3, inflammation, and cancer: How intimate is the relationship? Ann. N. Y. Acad. Sci. 2009, 1171, 59–76. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, S.; Drewry, J.A.; Shahani, V.M. Molecular disruption of oncogenic signal transducer and activator of transcription 3 (STAT3) protein. Biochem. Cell Biol. 2009, 87, 825–833. [Google Scholar] [CrossRef]
- Wernig, G.; Kharas, M.G.; Okabe, R. Efficacy of TG101348, a selective JAK2 inhibitor, in treatment of a murine model of JAK2V617F-induced polycythemia vera. Cancer Cell 2008, 13, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Koppikar, P.; Abdel-Wahab, O.; Hedvat, C. Efficacy of the JAK2 inhibitor INCB16562 in a murine model of MPLW515L-induced thrombocytosis and myelofibrosis. Blood 2010, 115, 2919–2927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinta´s-Cardama, A.; Vaddi, K.; Liu, P. Preclinical characterization of the selective JAK1/2 inhibitor INCB018424: Therapeutic implications for the treatment of myeloproliferative neoplasms. Blood 2010, 115, 3109–3117. [Google Scholar] [CrossRef] [PubMed]
- Sansone, P.; Bromberg, J. Targeting the interleukin-6/Jak/stat pathway in human malignancies. J. Clin. Oncol. 2012, 30, 1005–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedvat, M.; Huszar, D.; Herrmann, A. The JAK2 inhibitor AZD1480 potently blocks Stat3 signaling and oncogenesis in solid tumors. Cancer Cell. 2009, 16, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Xi, S.; Gooding, W.E.; Grandis, J.R. In vivo antitumor efficacy of STAT3 blockade using a transcription factor decoy approach: Implications for cancer therapy. Oncogene 2005, 24, 970–979. [Google Scholar] [CrossRef] [Green Version]
- Lui, V.W.; Boehm, A.L.; Koppikar, P. Antiproliferative mechanisms of a transcription factor decoy targeting signal transducer and activator of transcription (STAT) 3: The role of STAT1. Mol. Pharmacol. 2007, 71, 1435–1443. [Google Scholar] [CrossRef] [Green Version]
- Boehm, A.L.; Sen, M.; Seethala, R. Combined targeting of epidermal growth factor receptor, signal transducer and activator of transcription-3, and Bcl-X(L) enhances antitumor effects in squamous cell carcinoma of the head and neck. Mol. Pharmacol. 2008, 73, 1632–1642. [Google Scholar] [CrossRef] [Green Version]
- Sen, M.; Tosca, P.J.; Zwayer, C. Lack of toxicity of a STAT3 decoy oligonucleotide. Cancer Chemother. Pharmacol. 2009, 63, 983–995. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Jaganathan, S.; Turkson, J. A cell permeable STAT3 SH2 domain mimetic inhibits STAT3 activation and induces antitumor cell effects in vitro. J. Biol. Chem. 2010, 285, 35855–35865. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yue, P.; Fletcher, S. A novel small molecule disrupts Stat3 SH2 domain-phosphotyrosine interactions and Stat3-dependent tumor processes. Biochem. Pharmacol. 2010, 79, 1398–1409. [Google Scholar] [CrossRef] [Green Version]
- Redell, M.S.; Ruiz, M.J.; Alonzo, T.A. Stat3 signaling in acute myeloid leukemia: Ligand dependent and -independent activation and induction of apoptosis by a novel small-molecule Stat3 inhibitor. Blood 2011, 117, 5701–5709. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, S.; Singh, J.; Zhang, X.; Yue, P.; Page, B.D.; Sharmeen, S.; Shahani, V.M.; Zhao, W.; Schimmer, A.D.; Turkson, J.; et al. Disruption of transcriptionally active Stat3 dimers with non-phosphorylated, salicylic acid-based small molecules: Potent in vitro and tumor cell activities. Chembiochem 2009, 10, 1959–1964. [Google Scholar] [CrossRef] [PubMed]
- Madoux, F.; Koenig, M.; Sessions, H. Modulators of STAT transcription factors for the targeted therapy of cancer (STAT3 inhibitors). In Probe Reports from the NIH Molecular Libraries Program; National Center for Biotechnology Information: Bethesda, MD, USA, 2011; pp. 1–20. [Google Scholar]
- Nelson, E.A.; Sharma, S.V.; Settleman, J. A chemical biology approach to developing STAT inhibitors: Molecular strategies for accelerating clinical translation. Oncotarget 2011, 2, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.A.; Walker, S.R.; Weisberg, E. The STAT5 inhibitor pimozide decreases survival of chronic myelogenous leukemia cells resistant to kinase inhibitors. Blood 2011, 117, 3421–3429. [Google Scholar] [CrossRef]
- Nelson, E.A.; Walker, S.R.; Kepich, A. Nifuroxazide inhibits survival of multiple myeloma cells by directly inhibiting STAT3. Blood 2008, 112, 5095–5102. [Google Scholar] [CrossRef] [Green Version]
- Li, W.X. Canonical and non-canonical JAK/STAT signaling. Trends Cell. Biol. 2008, 18, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Regis, G.; Pensa, S.; Boselli, D. Ups and downs: The STAT1:STAT3 seesaw of interferon and gp130 receptor signalling. Semin. Cell Dev. Biol. 2008, 19, 351–359. [Google Scholar] [CrossRef]
- Kumar, A.; Commane, M.; Flickinger, T.W. Defective TNF-alpha-induced apoptosis in STAT1-null cells due to low constitutive levels of caspases. Science 1997, 278, 1630–1632. [Google Scholar] [CrossRef]
- Yang, J.; Chatterjee, K.M.; Staugaitis, S.M. Novel roles of unphosphorylated STAT3 in oncogenesis and transcriptional regulation. Cancer Res. 2005, 65, 939–947. [Google Scholar]
- Zhang, Q.; Wang, H.Y.; Marzec, M. STAT3- and DNA methyltransferase 1-mediated epigenetic silencing of SHP-1 tyrosine phosphatase tumor suppressor gene in malignant T lymphocytes. Proc. Natl. Acad. Sci. USA. 2005, 102, 6948–6953. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, H.Y.; Liu, X. STAT5A is epigenetically silenced by the tyrosine kinase NPM1-ALK and acts as a tumor suppressor by reciprocally inhibiting NPM1-ALK expression. Nat. Med. 2007, 13, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Gough, D.J.; Corlett, A.; Schlessinger, K. Mitochondrial STAT3 supports Ras-dependent oncogenic transformation. Science 2009, 324, 1713–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, N.C. STAT3 revs up the powerhouse. Sci. Signal. 2009, 2, 61–62. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.R.; Chaudhury, M.; Nelson, E.A. Microtubule-targeted chemotherapeutic agents inhibit signal transducer and activator of transcription 3 (STAT3) signaling. Mol. Pharmacol. 2010, 78, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Hazan-Halevy, I.; Harris, D.; Liu, Z. STAT3 is constitutively phosphorylated on serine 727 residues, binds DNA, and activates transcription in CLL cells. Blood 2010, 115, 2852–2863. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.Y.; Park, E.J.; Joe, E.H.; Jou, I. Curcumin suppresses Janus kinase-STAT inflammatory signaling through activation of Src homology 2 domain-containing tyrosine phosphatase 2 in brain microglia. J. Immunol. 2003, 171, 6072–6079. [Google Scholar] [CrossRef] [Green Version]
- Calvisi, D.F.; Ladu, S.; Gorden, A.; Farina, M.; Conner, E.A.; Lee, J.S.; Factor, V.M.; Thorgeirsson, S.S. Ubiquitous activation of Ras and Jak/Stat pathways in human HCC. Gastroenterology 2006, 130, 1117–1128. [Google Scholar] [CrossRef]
- Rhee, Y.H.; Jeong, S.J.; Lee, H.J. Inhibition of STAT3 signaling and induction of SHP1 mediate antiangiogenic and antitumor activities of ergosterol peroxide in U266 multiple myeloma cells. BMC Cancer 2012, 12, 28. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Cai, X.; Lu, W.; Hu, C.; Xu, X.; Yu, Q.; Cao, P. Evodiamine inhibits STAT3 signaling by inducing phosphatase shatterproof 1 in hepatocellular carcinoma cells. Cancer Lett. 2013, 328, 243–251. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Bhardwaj, A.; Aggarwal, R.S.; Seeram, N.P.; Shishodia, S.; Takada, Y. Role of resveratrol in prevention and therapy of cancer: Preclinical and clinical studies. Anticancer Res. 2004, 24, 2783–2840. [Google Scholar]
- Bishayee, A. Cancer prevention and treatment with resveratrol: From rodent studies to clinical trials. Cancer Prev. Res. 2009, 2, 409–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.H.; Sethi, G.; Um, J.Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The role of resveratrol in cancer therapy. Int. J. Mol. Sci. 2017, 18, 2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauf, A.; Imran, M.; Butt, M.S.; Nadeem, M.; Peters, D.G.; Mubarak, M.S. Resveratrol as an anti-cancer agent: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1428–1447. [Google Scholar] [CrossRef] [PubMed]
- Madan, E.; Prasad, S.; Roy, P.; George, J.; Shukla, Y. Regulation of apoptosis by resveratrol through JAK/STAT and mitochondria mediated pathway in human epidermoid carcinoma A431 cells. Biochem. Biophys. Res. Commun. 2008, 377, 1232–1237. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.; Sethi, G.; Vadhan-Raj, S. Resveratrol inhibits proliferation, induces apoptosis, and overcomes chemoresistance through down-regulation of STAT3 and nuclear factor-kappaB-regulated antiapoptotic and cell survival gene products in human multiple myeloma cells. Blood 2007, 109, 2293–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotha, A.; Sekharam, M.; Cilenti, L. Resveratrol inhibits Src and Stat3 signaling and induces the apoptosis of malignant cells containing activated Stat3 protein. Mol. Cancer Ther. 2006, 5, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Molavi, O.; Haddadi, A.; Lai, R.; Gossage, R.A.; Lavasanifar, A. Resveratrol analog trans 3, 4, 5, 4′-tetramethoxystilbene (DMU-212) mediates anti-tumor effects via mechanism different from that of resveratrol. Cancer Chemother. Pharmacol. 2008, 63, 27–35. [Google Scholar] [CrossRef]
- Nwachukwu, J.C.; Srinivasan, S.; Bruno, N.E. Resveratrol modulates the inflammatory response via an estrogen receptor-signal integration network. Elife 2014, 3, 02057. [Google Scholar] [CrossRef]
- Anand, P.; Sundaram, C.; Jhurani, S.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin and cancer: An “old-age” disease with an “age-old” solution. Cancer Lett. 2008, 267, 133–164. [Google Scholar] [CrossRef]
- Darvesh, A.S.; Aggarwal, B.B.; Bishayee, A. Curcumin and liver cancer: A review. Curr. Pharm. Biotechnol. 2012, 13, 218–228. [Google Scholar] [CrossRef]
- Sinha, D.; Biswas, J.; Sung, B.; Aggarwal, B.B.; Bishayee, A. Chemopreventive and chemotherapeutic potential of curcumin in breast cancer. Curr. Drug Targets 2012, 13, 1799–1819. [Google Scholar] [CrossRef] [PubMed]
- Kasi, P.D.; Tamilselvam, R.; Skalicka-Woźniak, K.; Nabavi, S.F.; Daglia, M.; Bishayee, A.; Nabavi, S.M. Molecular targets of curcumin for cancer therapy: An updated review. Tumor Biol. 2016, 37, 13017–13028. [Google Scholar] [CrossRef] [PubMed]
- Bharti, A.C.; Donato, N.; Aggarwal, B.B. Curcumin (diferuloylmethane) inhibits constitutive and IL-6-inducible STAT3 phosphorylation in human multiple myeloma cells. J. Immunol. 2003, 171, 3863–3871. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.L.; Liu, Y.Y.; Ma, Y.G. Curcumin blocks small cell lung cancer cells migration, invasion, angiogenesis, cell cycle and neoplasia through Janus kinase–STAT3 signalling pathway. PLoS ONE 2012, 7, e37960. [Google Scholar] [CrossRef]
- Rajasingh, J.; Raikwar, H.P.; Muthian, G.; Johnson, C.; Bright, J.J. Curcumin induces growth-arrest and apoptosis in association with the inhibition of constitutively active JAK–STAT pathway in T cell leukemia. Biochem. Biophys. Res. Commun. 2006, 340, 359–368. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Bordoloi, D.; Harsha, C.; Banik, K.; Gupta, S.C.; Aggarwal, B.B. Curcumin mediates anticancer effects by modulating multiple cell signaling pathways. Clin. Sci. 2017, 131, 1781–1799. [Google Scholar] [CrossRef]
- Min-Wen, J.C.; Yan-Jiang, B.C.; Mishra, S.; Dai, X.; Magae, J.; Shyh-Chang, N.; Kumar, A.P.; Sethi, G. Molecular Targets of Ascochlorin and Its Derivatives for Cancer Therapy. Adv. Protein Chem. Struct. Biol. 2017, 108, 199–225. [Google Scholar] [CrossRef]
- Cho, H.J.; Park, J.H.; Nam, J.H.; Chang, Y.C.; Park, B.; Hoe, H.S. Ascochlorin suppresses MMP-2-mediated migration and invasion by targeting FAK and JAK-STAT signaling cascades. J. Cell. Biochem. 2018, 119, 300–313. [Google Scholar] [CrossRef]
- Ko, J.H.; Arfuso, F.; Sethi, G.; Ahn, K.S. Pharmacological Utilization of Bergamottin, Derived from Grapefruits, in Cancer Prevention and Therapy. Int. J. Mol. Sci. 2018, 19, 4048. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M.; Lee, J.H.; Sethi, G.; Kim, C.; Baek, S.H.; Nam, D.; Ahn, K.S. Bergamottin, a natural furanocoumarin obtained from grapefruit juice induces chemosensitization and apoptosis through the inhibition of STAT3 signaling pathway in tumor cells. Cancer Lett. 2014, 354, 153–163. [Google Scholar] [CrossRef]
- Lee, J.H.; Chiang, S.Y.; Nam, D.; Chung, W.S.; Lee, J.; Na, Y.S.; Ahn, K.S. Capillarisin inhibits constitutive and inducible STAT3 activation through induction of SHP-1 and SHP-2 tyrosine phosphatases. Cancer Lett. 2014, 345, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.; Tsubaki, M.; Tomonari, Y.; Kawashima, K.; Itoh, T.; Imano, M.; Nishida, S. Bavachin induces the apoptosis of multiple myeloma cell lines by inhibiting the activation of nuclear factor kappa B and signal transducer and activator of transcription 3. Biomed. Pharmacother. 2018, 100, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darvesh, A.S.; Bishayee, A. Chemopreventive and therapeutic potential of tea polyphenols in hepatocellular cancer. Nutr. Cancer 2013, 65, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, H. Cancer preventive activities of tea catechins. Molecules 2016, 21, 1679. [Google Scholar] [CrossRef]
- Sinha, D.; Biswas, J.; Nabavi, S.M.; Bishayee, A. Tea phytochemicals for breast cancer prevention and intervention: From bench to bedside and beyond. Semin. Cancer Biol. 2017, 46, 33–54. [Google Scholar] [CrossRef]
- Tang, S.N.; Fu, J.; Shankar, S.; Srivastava, R.K. EGCG enhances the therapeutic potential of gemcitabine and CP690550 by inhibiting STAT3 signaling pathway in human pancreatic cancer. PLoS ONE 2012, 7, 31067. [Google Scholar] [CrossRef]
- Shimizu, M.; Shirakami, Y.; Sakai, H. EGCG inhibits activation of the insulin-like growth factor (IGF)/IGF-1 receptor axis in human hepatocellular carcinoma cells. Cancer Lett. 2008, 262, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Masuda, M.; Suzui, M.; Weinstein, I.B. Effects of epigallocatechin-3-gallate on growth, epidermal growth factor receptor signaling pathways, gene expression, and chemosensitivity in human head and neck squamous cell carcinoma cell lines. Clin. Cancer Res. 2001, 7, 4220–4229. [Google Scholar]
- Masuda, M.; Suzui, M.; Lim, J.T.E.; Weinstein, I.B. Epigallocatechin-3-gallate inhibits activation of HER-2/neu and downstream signaling pathways in human head and neck and breast carcinoma cells. Clin. Cancer Res. 2003, 9, 3486–3491. [Google Scholar]
- Xiao, X.; Jiang, K.; Xu, Y.; Peng, H.; Wang, Z.; Liu, S.; Zhang, G. (-)-Epigallocatechin-3-gallate induces cell apoptosis in chronic myeloid leukemia by regulating Bcr/Abl-mediated p38-MAPK/JNK and JAK2/STAT3/AKT signaling pathways. Clin. Exp. Pharmacol. Physiol. 2019, 46, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Harlev, E.; Nevo, E.; Lansky, E.P.; Ofir, R.; Bishayee, A. Anticancer potential of aloes: Antioxidant, antiproliferative, and immunostimulatory attributes. Planta Med. 2012, 78, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.T.; Lin, S.Z.; Liu, D.L.; Wang, Z.H. The distinct mechanisms of the antitumor activity of emodin in different types of cancer (Review). Oncol. Rep. 2013, 30, 2555–2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, B.; Ray, A.M.; Goldberg, S.; Clark, T.; McDaniel, H.R.; Atlas, S.E.; Farooqi, A.; Konefal, J.; Lages, L.C.; Lopez, J.; et al. Anti-cancer effects of aloe-emodin: A systematic review. J. Clin. Transl. Res. 2017, 3, 283–296. [Google Scholar] [PubMed]
- He, Y.; Huang, J.; Wang, P.; Shen, X.; Li, S.; Yang, L.; Wang, F. Emodin potentiates the antiproliferative effect of interferon α/β by activation of JAK/STAT pathway signaling through inhibition of the 26S proteasome. Oncotarget 2016, 7, 4664. [Google Scholar] [CrossRef] [Green Version]
- Mahapatra, D.K.; Bharti, S.K.; Asati, V. Anti-cancer chalcones: Structural and molecular target perspectives. Eur. J. Med. Chem. 2015, 98, 69–114. [Google Scholar] [CrossRef]
- León-González, A.J.; Acero, N.; Muñoz-Mingarro, D.; Navarro, I.; Martín-Cordero, C. Chalcones as Promising Lead Compounds on Cancer Therapy. Curr. Med. Chem. 2015, 22, 3407–3425. [Google Scholar] [CrossRef]
- Liu, Y.C.; Hsieh, C.W.; Wu, C.C.; Wung, B.S. Chalcone inhibits the activation of NF-kappaB and STAT3 in endothelial cells via endogenous electrophile. Life Sci. 2007, 80, 1420–1430. [Google Scholar] [CrossRef]
- Ong, S.K.L.; Shanmugam, M.K.; Fan, L.; Fraser, S.E.; Arfuso, F.; Ahn, K.S.; Sethi, G.; Bishayee, A. Focus on Formononetin: Anticancer Potential and Molecular Targets. Cancers 2019, 11, 611. [Google Scholar] [CrossRef] [Green Version]
- Tay, K.C.; Tan, L.T.; Chan, C.K.; Hong, S.L.; Chan, K.G.; Yap, W.H.; Pusparajah, P.; Lee, L.H.; Goh, B.H. Formononetin: A Review of Its Anticancer Potentials and Mechanisms. Front. Pharmacol. 2019, 10, 820. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.L.; Li, Y.; Zhao, Q.; Fan, L.Q. Formononetin inhibits colon carcinoma cell growth and invasion by microRNA-149-mediated EphB3 downregulation and inhibition of PI3K/AKT and STAT3 signaling pathways. Mol. Med. Rep. 2018, 17, 7721–7729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Ho, P.C.; Wong, F.C.; Sethi, G.; Wang, L.Z.; Goh, B.C. Garcinol: Current status of its anti-oxidative, anti-inflammatory and anti-cancer effects. Cancer Lett. 2015, 362, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Behera, A.K.; Swamy, M.M.; Natesh, N.; Kundu, T.K. Garcinol and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 928, 435–452. [Google Scholar] [PubMed]
- Sethi, G.; Chatterjee, S.; Rajendran, P.; Li, F.; Shanmugam, M.K.; Wong, K.F.; Basha, J. Inhibition of STAT3 dimerization and acetylation by garcinol suppresses the growth of human hepatocellular carcinoma In Vitro and In Vivo. Mol. Cancer 2014, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Chua, L.K.; Lim, C.L.; Ling, A.P.K.; Chye, S.M.; Koh, R.Y. Anticancer Potential of Syzygium Species: A Review. Plant Foods Hum. Nutr. 2019, 74, 18–27. [Google Scholar] [CrossRef]
- Wu, N.; Liu, J.; Zhao, X.; Yan, Z.; Jiang, B.; Wang, L.; Cao, S.; Shi, D.; Lin, X. Cardamonin induces apoptosis by suppressing STAT3 signaling pathway in glioblastoma stem cells. Tumor Biol. 2015, 36, 9667–9676. [Google Scholar] [CrossRef]
- Murtaza, G.; Sajjad, A.; Mehmood, Z.; Shah, S.H.; Siddiqi, A.R. Possible molecular targets for therapeutic applications of caffeic acid phenethyl ester in inflammation and cancer. J. Food Drug Anal. 2015, 23, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Damasceno, S.S.; Dantas, B.B.; Ribeiro-Filho, J.; Antônio, M.; Araújo, D.; Galberto, M.; da Costa, J. Chemical Properties of Caffeic and Ferulic Acids in Biological System: Implications in Cancer Therapy. A Review. Curr. Pharm. Des. 2017, 23, 3015–3023. [Google Scholar] [CrossRef]
- Jung, J.E.; Kim, H.S.; Lee, C.S. Caffeic acid and its synthetic derivative CADPE suppress tumor angiogenesis by blocking STAT3-mediated VEGF expression in human renal carcinoma cells. Carcinogenesis 2007, 28, 1780–1787. [Google Scholar] [CrossRef]
- Deep, G.; Agarwal, R. Targeting tumor microenvironment with silibinin: Promise and potential for a translational cancer chemopreventive strategy. Curr. Cancer Drug Target. 2013, 13, 486–499. [Google Scholar] [CrossRef] [Green Version]
- Mastron, J.K.; Siveen, K.S.; Sethi, G.; Bishayee, A. Silymarin and hepatocellular carcinoma: A systematic, comprehensive, and critical review. Anti-Cancer Drugs 2015, 26, 475–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.X.; Ding, Y.H.; Wu, Y.; Qian, L.Y.; Zou, H.; He, Q. Silibinin: A potential old drug for cancer therapy. Expert Rev. Clin. Pharmacol. 2016, 9, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, C.; Tyagi, A.; Kaur, M.; Agarwal, R. Silibinin inhibits constitutive activation of Stat3 and causes caspase activation and apoptotic death of human prostate carcinoma DU145 cells. Carcinogenesis 2007, 28, 1463–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, A.; Singh, R.P.; Ramasamy, K. Growth inhibition and regression of lung tumors by silibinin: Modulation of angiogenesis by macrophage-associated cytokines and nuclear factorkappaB and signal transducers and activators of transcription 3. Cancer Prev. Res. 2009, 2, 74–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmavathi, G.; Roy, N.K.; Bordoloi, D.; Arfuso, F.; Mishra, S.; Sethi, G.; Bishayee, A.; Kunnumakkara, A.B. Butein in health and disease: A comprehensive review. Phytomedicine 2017, 25, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Jayasooriya, R.G.P.T.; Molagoda, I.M.N.; Park, C.; Jeong, J.W.; Choi, Y.H.; Moon, D.O.; Kim, M.O.; Kim, G.Y. Molecular chemotherapeutic potential of butein: A concise review. Food Chem. Toxicol. 2018, 112, 1–10. [Google Scholar] [CrossRef]
- Bhutani, M.; Pathak, A.K.; Nair, A.S. Capsaicin is a novel blocker of constitutive and interleukin-6-inducible STAT3 activation. Clin. Cancer Res. 2007, 13, 3024–3032. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, P.; Ong, T.H.; Chen, L. Suppression of signal transducer and activator of transcription 3 activation by butein inhibits growth of human hepatocellular carcinoma in vivo. Clin. Cancer Res. 2011, 17, 1425–1439. [Google Scholar] [CrossRef] [Green Version]
- Kasala, E.R.; Bodduluru, L.N.; Madana, R.M.; V, A.K.; Gogoi, R.; Barua, C.C. Chemopreventive and therapeutic potential of chrysin in cancer: Mechanistic perspectives. Toxicol. Lett. 2015, 233, 214–225. [Google Scholar] [CrossRef]
- Khoo, B.Y.; Chua, S.L.; Balaram, P. Apoptotic effects of chrysin in human cancer cell lines. Int. J. Mol. Sci. 2010, 11, 2188–2199. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Ye, T.; Cai, X.; Yang, J.; Lu, W.; Hu, C. 5, 7-Dihydroxyflavone enhances the apoptosis-inducing potential of Trail in human tumor cells via regulation of apoptosis-related proteins. Evid. Based Complement. Alternat. Med. 2013, 2013, 434709. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.R.; Lu, R.; Dan, H.X.; Liao, G.; Zhou, M.; Li, X.Y. Honokiol: A promising small molecular weight natural agent for the growth inhibition of oral squamous cell carcinoma cells. Int. J. Oral Sci. 2011, 3, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Dikalov, S.; Losik, T.; Arbiser, J.L. Honokiol is a potent scavenger of superoxide and peroxyl radicals. Biochem. Pharmacol. 2008, 76, 589–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Averett, C.; Arora, S.; Zubair, H.; Singh, S.; Bhardwaj, A.; Singh, A.P. Molecular Targets of Honokiol: A Promising Phytochemical for Effective Cancer Management. Enzymes 2014, 36, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Katiyar, S.K. Honokiol, an Active Compound of Magnolia Plant, Inhibits Growth, and Progression of Cancers of Different Organs. Adv. Exp. Med. Biol. 2016, 928, 245–265. [Google Scholar] [PubMed]
- Banik, K.; Ranaware, A.M.; Deshpande, V.; Nalawade, S.P.; Padmavathi, G.; Bordoloi, D.; Sailo, B.L.; Shanmugam, M.K.; Fan, L.; Arfuso, F.; et al. Honokiol for cancer therapeutics: A traditional medicine that can modulate multiple oncogenic targets. Pharmacol. Res. 2019, 144, 192–209. [Google Scholar] [CrossRef]
- Bi, L.; Yu, Z.; Wu, J.; Yu, K.; Hong, G.; Lu, Z.; Gao, S. Honokiol inhibits constitutive and inducible STAT3 signaling via PU. 1-induced SHP1 expression in acute myeloid leukemia cells. Tohoku J. Exp. Med. 2015, 237, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Rasul, A.; Zhao, B.J.; Liu, J.; Liu, B.; Sun, J.X.; Li, J.; Li, X.M. Molecular mechanisms of casticin action: An update on its antitumor functions. Asian Pac. J. Cancer Prev. 2014, 15, 9049–9058. [Google Scholar] [CrossRef] [Green Version]
- Chan, E.W.C.; Wong, S.K.; Chan, H.T. Casticin from Vitex species: A short review on its anticancer and anti-inflammatory properties. J. Integr. Med. 2018, 16, 147–152. [Google Scholar] [CrossRef]
- Chen, D.; Cao, J.; Tian, L.; Liu, F.; Sheng, X. Induction of apoptosis by casticin in cervical cancer cells through reactive oxygen species mediated mitochondrial signaling pathways. Oncol. Rep. 2011, 26, 1287–1294. [Google Scholar] [CrossRef]
- Zeng, F.; Tian, L.; Liu, F.; Cao, J.; Quan, M.; Sheng, X. Induction of apoptosis by casticin in cervical cancer cells: Reactive oxygen species-dependent sustained activation of Jun N-terminal kinase. Acta Biochim. Biophys. Sin. 2012, 44, 442–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Kim, C.; Ko, J.H.; Jung, Y.Y.; Jung, S.H.; Kim, E.; Sethi, G. Casticin inhibits growth and enhances ionizing radiation–induced apoptosis through the suppression of STAT3 signaling cascade. J. Cell Biochem. 2019, 120, 9787–9798. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, A.; Bodalska, A.; Miranowicz, M.; Karłowicz-Bodalska, K. Insights into novel anticancer applications for apigenin. Adv. Clin. Exp. Med. 2017, 26, 1143–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madunić, J.; Madunić, I.V.; Gajski, G.; Popić, J.; Garaj-Vrhovac, V. Apigenin: A dietary flavonoid with diverse anticancer properties. Cancer Lett. 2018, 413, 11–22. [Google Scholar] [CrossRef]
- Maeda, Y.; Takahashi, H.; Nakai, N.; Yanagita, T.; Ando, N.; Okubo, T.; Saito, K.; Shiga, K.; Hirokawa, T.; Hara, M.; et al. Apigenin induces apoptosis by suppressing Bcl-xl and Mcl-1 simultaneously via signal transducer and activator of transcription 3 signaling in colon cancer. Int. J. Oncol. 2018, 52, 1661–1673. [Google Scholar] [CrossRef]
- Pinto, D.C.G.A.; Silva, A.M.S. Anticancer Natural Coumarins as Lead Compounds for the Discovery of New Drugs. Curr. Top. Med. Chem. 2017, 17, 3190–3198. [Google Scholar] [CrossRef]
- Benes, P.; Knopfova, L.; Trcka, F.; Nemajerova, A.; Pinheiro, D.; Soucek, K. Inhibition of topoisomerase IIalpha: Novel function of wedelolactone. Cancer Lett. 2011, 303, 29–38. [Google Scholar] [CrossRef]
- Benes, P.; Alexova, P.; Knopfova, L.; Spanova, A.; Smarda, J. Redox state alters anti-cancer effects of wedelolactone. Environ. Mol. Mutagen. 2012, 53, 515–524. [Google Scholar] [CrossRef]
- Chen, Z.; Sun, X.; Shen, S.; Zhang, H.; Ma, X.; Liu, J.; Kuang, S.; Yu, Q. Wedelolactone, a naturally occurring coumestan, enhances interferon-γ signaling through inhibiting STAT1 protein dephosphorylation. J. Biol. Chem. 2013, 288, 14417–14427. [Google Scholar] [CrossRef] [Green Version]
- Rabi, T.; Bishayee, A. Terpenoids and breast cancer chemoprevention. Breast Cancer Res. Treat. 2009, 115, 223–239. [Google Scholar] [CrossRef]
- Thoppil, R.J.; Bishayee, A. Terpenoids as potential chemopreventive and therapeutic agents in liver cancer. World J. Hepatol. 2011, 3, 228–249. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Dai, X.; Kumar, A.P.; Tan, B.K.; Sethi, G.; Bishayee, A. Oleanolic acid and its synthetic derivatives for the prevention and therapy of cancer: Preclinical and clinical evidence. Cancer Lett. 2014, 346, 206–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.C.; Chiu, M.H.; Nie, R.L.; Cordell, G.A.; Qiu, S.X. Cucurbitacins and cucurbitane glycosides: Structures and biological activities. Nat. Prod. Rep. 2005, 22, 386–399. [Google Scholar] [CrossRef]
- Lee, D.H.; Iwanski, G.B.; Thoennissen, N.H. Cucurbitacin: Ancient compound shedding new light on cancer treatment. Sci. World J. 2010, 10, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Fang, X.; He, C.; Li, P.; Xiao, F.; Wang, Y.; Chen, M. Cucurbitacins: A Systematic Review of the Phytochemistry and Anticancer Activity. Am. J. Chin. Med. 2015, 43, 1331–1350. [Google Scholar] [CrossRef] [PubMed]
- Thoennissen, N.H.; Iwanski, G.B.; Doan, N.B. Cucurbitacin B induces apoptosis by inhibition of the JAK/STAT pathway and potentiates antiproliferative effects of gemcitabine on pancreatic cancer cells. Cancer Res. 2009, 69, 5876–5884. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Liu, M.; Chen, Y.; Xu, S.; Guo, Y.; Zhao, L. Cucurbitacin B suppresses proliferation of pancreatic cancer cells by ceRNA: Effect of miR-146b-5p and lncRNA-AFAP1-AS1. J. Cell Physiol. 2019, 234, 4655–4667. [Google Scholar] [CrossRef]
- Chan, K.T.; Li, K.; Liu, S.L.; Chu, K.H.; Toh, M.; Xie, W.D. Cucurbitacin B inhibits STAT3 and the Raf/MEK/ERK pathway in leukemia cell line K562. Cancer Lett. 2010, 289, 46–52. [Google Scholar] [CrossRef]
- Huang, W.W.; Yang, J.S.; Lin, M.W. Cucurbitacin E Induces G2/M phase arrest through STAT3/p53/p21 signaling and provokes apoptosis via Fas/CD95 and mitochondria dependent pathways in human bladder cancer T24 cells. Evid. Based Complement. Altern. Med. 2012, 2012, 952762. [Google Scholar] [CrossRef] [Green Version]
- Blaskovich, M.A.; Sun, J.; Cantor, A.; Turkson, J.; Jove, R.; Sebti, S.M. Discovery of JSI-124 (cucurbitacin I), a selective Janus kinase/signal transducer and activator of transcription 3 signaling pathway inhibitor with potent antitumor activity against human and murine cancer cells in mice. Cancer Res. 2003, 63, 1270–1279. [Google Scholar]
- Su, Y.; Li, G.; Zhang, X. JSI-124 inhibits glioblastoma multiforme cell proliferation through G2/M cell cycle arrest and apoptosis augmentation. Cancer Biol. Ther. 2014, 7, 1243–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kester, M.S.; Out-Luiting, J.J.; von dem Borne, P.A.; Willemze, R.; Tensen, C.P.; Vermeer, M.H. Cucurbitacin I inhibits Stat3 and induces apoptosis in Sézary cells. J. Investig. Dermatol. 2008, 128, 1691–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Franko, B.; Frantz, C.; Amin, H.M.; Lai, R. JSI-124 (cucurbitacin I) inhibits Janus kinase-3/signal transducer and activator of transcription-3 signalling, downregulates nucleophosmin-anaplastic lymphoma kinase (ALK), and induces apoptosis in ALK-positive anaplastic large cell lymphoma cells. Br. J. Haematol. 2006, 135, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Blaskovich, M.A.; Jove, R.; Livingston, S.K.; Coppola, D.; Sebti, S.M. Cucurbitacin Q: A selective STAT3 activation inhibitor with potent antitumor activity. Oncogene 2005, 24, 3236–3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.K.; Tripathi, S.; Shukla, A.; Oh, S.H.; Kim, H.M. Andrographolide and analogues in cancer prevention. Front. Biosci. 2015, 7, 255–266. [Google Scholar] [CrossRef]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Islam, M.A.; Shaw, S.; Khan, I.N.; Saravi, S.S.S.; Ahmad, S.; Rehman, S.; Gupta, V.K.; et al. Andrographolide, a diterpene lactone from Andrographis paniculate and its therapeutic promises in cancer. Cancer Lett. 2018, 420, 129–145. [Google Scholar] [CrossRef]
- Zhou, J.; Ong, C.N.; Hur, G.M.; Shen, H.M. Inhibition of the JAK-STAT3 pathway by andrographolide enhances chemosensitivity of cancer cells to doxorubicin. Biochem. Pharmacol. 2010, 79, 1242–1250. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, J.; Chen, Y. Betulinic acid and the pharmacological effects of tumor suppression (Review). Mol. Med. Rep. 2016, 14, 4489–4495. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Bhadauria, A.S.; Singh, A.K.; Saha, S. Betulinic acid as apoptosis activator: Molecular mechanisms, mathematical modeling and chemical modifications. Life Sci. 2018, 209, 24–33. [Google Scholar] [CrossRef]
- Pandey, M.K.; Sung, B.; Aggarwal, B.B. Betulinic acid suppresses STAT3 activation pathway through induction of protein tyrosine phosphatase SHP-1 in human multiple myeloma cells. Int. J. Cancer 2010, 127, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, V.; Kashyap, D.; Sak, K.; Tuli, H.S.; Jain, A.; Chaudhary, A.; Garg, V.K.; Sethi, G.; Yerer, M.B. Molecular Mechanisms of Action of Tocotrienols in Cancer: Recent Trends and Advancements. Int. J. Mol. Sci. 2019, 20, 656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagnani Marelli, M.; Marzagalli, M.; Fontana, F.; Raimondi, M.; Moretti, R.M.; Limonta, P. Anticancer properties of tocotrienols: A review of cellular mechanisms and molecular targets. J. Cell. Physiol. 2019, 234, 1147–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajendran, P.; Li, F.; Manu, K.A. γ-Tocotrienol is a novel inhibitor of constitutive and inducible STAT3 signalling pathway in human hepatocellular carcinoma: Potential role as an antiproliferative, pro-apoptotic and chemosensitizing agent. Br. J. Pharmacol. 2011, 163, 283–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, S.; Bose, S.; Mandal, S.C.; Dawn, S.; Sahoo, U.; Ramadan, M.A.M.; Mandal, S. Pharmacological Property of Pentacyclic Triterpenoids. Egypt. J. Chem. 2019, 62, 13–35. [Google Scholar] [CrossRef]
- Chen, W.; Lu, Y.; Chen, G.; Huang, S. Molecular evidence of cryptotanshinone for treatment and prevention of human cancer. Anticancer Agents Med. Chem. 2013, 13, 979–987. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.S.; Kim, H.N.; Shin, K.D. Cryptotanshinone inhibits constitutive signal transducer and activator of transcription 3 function through blocking the dimerization in DU145 prostate cancer cells. Cancer Res. 2009, 69, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.M.; Nagulapalli Venkata, K.C.; Bhattacharyya, P.; Sethi, G.; Bishayee, A. Potential of neem (Azadirachta indica L.) for prevention and treatment of oncologic diseases. Semin. Cancer Biol. 2016, 40–41, 100–115. [Google Scholar] [CrossRef]
- Wang, L.; Phan, D.D.; Zhang, J.; Ong, P.S.; Thuya, W.L.; Soo, R.; Wong, A.L.; Yong, W.P.; Lee, S.C.; Ho, P.C.; et al. Anticancer properties of nimbolide and pharmacokinetic considerations to accelerate its development. Oncotarget. 2016, 7, 44790–44802. [Google Scholar] [CrossRef] [Green Version]
- Karkare, S.; Chhipa, R.R.; Anderson, J.; Liu, X.; Henry, H.; Gasilina, A.; Pauletti, G.M. Direct inhibition of retinoblastoma phosphorylation by nimbolide causes cell-cycle arrest and suppresses glioblastoma growth. Clin. Cancer Res. 2014, 20, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ahn, K.S.; Kim, C.; Shanmugam, M.K.; Siveen, K.S.; Arfuso, F.; Goh, B.C. Nimbolide-induced oxidative stress abrogates STAT3 signaling cascade and inhibits tumor growth in transgenic adenocarcinoma of mouse prostate model. Antioxid. Redox Signal. 2016, 24, 575–589. [Google Scholar] [CrossRef]
- Kannaiyan, R.; Shanmugam, M.K.; Sethi, G. Molecular targets of celastrol derived from Thunder of God Vine: Potential role in the treatment of inflammatory disorders and cancer. Cancer Lett. 2011, 303, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Sharma, A.; Tuli, H.S.; Sak, K.; Mukherjee, T.; Bishayee, A. Molecular targets of celastrol in cancer: Recent trends and advancements. Crit. Rev. Oncol. Hematol. 2018, 128, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Kannaiyan, R.; Hay, H.S.; Rajendran, P. Celastrol inhibits proliferation and induces chemosensitization through downregulation of NF-κB and STAT3 regulated gene products in multiple myeloma cells. Br. J. Pharmacol. 2011, 164, 1506–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugam, M.K.; Dai, X.; Kumar, A.P.; Tan, B.K.; Sethi, G.; Bishayee, A. Ursolic acid in cancer prevention and treatment: Molecular targets, pharmacokinetics and clinical studies. Biochem. Pharmacol. 2013, 85, 1579–1587. [Google Scholar] [CrossRef] [Green Version]
- Chan, E.W.C.; Soon, C.Y.; Tan, J.B.L.; Wong, S.K.; Hui, Y.W. Ursolic acid: An overview on its cytotoxic activities against breast and colorectal cancer cells. J. Integr. Med. 2019, 17, 155–160. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Rajendran, P.; Li, F. Ursolic acid inhibits multiple cell survival pathways leading to suppression of growth of prostate cancer xenograft in nude mice. J. Mol. Med. 2011, 89, 713–727. [Google Scholar] [CrossRef]
- Lee, J.H.; Rangappa, S.; Mohan, C.D.; Sethi, G.; Lin, Z.X.; Rangappa, K.S.; Ahn, K.S. Brusatol, a Nrf2 Inhibitor Targets STAT3 Signaling Cascade in Head and Neck Squamous Cell Carcinoma. Biomolecules 2019, 9, 550. [Google Scholar] [CrossRef] [Green Version]
- Owona, B.A.; Schluesener, H.J. Molecular Insight in the Multifunctional Effects of Oridonin. Drugs RD 2015, 15, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wold, E.A.; Ding, Y.; Shen, Q.; Zhou, J. Therapeutic Potential of Oridonin and Its Analogs: From Anticancer and Antiinflammation to Neuroprotection. Molecules 2018, 23, 474. [Google Scholar] [CrossRef] [Green Version]
- Bu, H.Q.; Luo, J.; Chen, H.; Zhang, J.H.; Li, H.H.; Guo, H.C. Oridonin enhance es antitumor activity of gemcitabine in pancreatic cancer through MAPK-p38 signaling pathway. Int. J. Oncol. 2012, 41, 949–958. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.H.; Wang, S.B.; Li, E.M.; Chen, L.M.; Yuan, S.J.; Wang, R.L. Inhibitory effect of Oridonin injection on heterotransplanted gastric adenocarcinoma in nude mice and its mechanism. Zhonghua Zhong Liu Za Zhi. 2008, 30, 89–92. [Google Scholar] [PubMed]
- Chen, G.; Wang, K.; Yang, B.Y.; Tang, B.; Chen, J.X.; Hua, Z.C. Synergistic antitumor activity of oridonin and arsenic trioxide on hepatocellular carcinoma cells. Int. J. Oncol. 2012, 40, 139–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.Y.; Xu, B.; Chen, S.F.; Chen, A.S.; Zhang, T.; Ren, J.; Xu, J. Effect of oridonin-mediated hallmark changes on inflammatory pathways in human pancreatic cancer (BxPC-3) cells. World J. Gastroenterol. 2014, 20, 14895–14903. [Google Scholar] [CrossRef] [PubMed]
- Vitale, G.; Zappavigna, S.; Marra, M.; Dicitore, A.; Meschini, S.; Condello, M.; Arancia, G.; Castiglioni, S.; Maroni, P.; Bendinelli, P.; et al. The PPAR-γ agonist troglitazone antagonizes survival pathways induced by STAT-3 in recombinant interferon-β treated pancreatic cancer cells. Biotechnol. Adv. 2012, 30, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rauf, A.; Khan, I.A.; Shahbaz, M.; Qaisrani, T.B.; Fatmawati, S.; Gondal, T.A. Thymoquinone: A novel strategy to combat cancer: A review. Biomed. Pharmacother. 2018, 106, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.Q.; Wang, J.; Guo, X.F.; Liu, Z.; Dong, W.G. Thymoquinone inhibits proliferation in gastric cancer via the STAT3 pathway In Vivo and In Vitro. World J. Gastroenterol. 2016, 22, 4149. [Google Scholar] [CrossRef]
- Li, F.; Rajendran, P.; Sethi, G. Thymoquinone inhibits proliferation, induces apoptosis and chemosensitizes human multiple myeloma cells through suppression of signal transducer and activator of transcription 3 activation pathway. Br. J. Pharmacol. 2010, 161, 541–554. [Google Scholar] [CrossRef] [Green Version]
- Ghantous, A.; Sinjab, A.; Herceg, Z.; Darwiche, N. Parthenolide: From plant shoots to cancer roots. Drug Discov. Today 2013, 18, 894–905. [Google Scholar] [CrossRef]
- Cheng, G.; Xie, L. Parthenolide induces apoptosis and cell cycle arrest of human 5637 bladder cancer cells in vitro. Molecules 2011, 16, 6758–6768. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.W.; Cai, M.X.; Xin, Y.; Wu, Q.S.; Ma, J.; Yang, P. Parthenolide induces proliferation inhibition and apoptosis of pancreatic cancer cells in vitro. J. Exp. Clin. Cancer Res. 2010, 29, 108. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, R.; Kusumanchi, P.; Appaiah, H.; Cheng, L.; Crooks, P.; Neelakantan, S. A water soluble parthenolide analog suppresses In Vivo tumor growth of two tobacco-associated cancers, lung and bladder cancer, by targeting NF-kappaB and generating reactive oxygen species. Int. J. Cancer 2011, 128, 2481–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Xiao, C.; Sun, M.; Tan, M.; Hu, L.; Yu, Q. Parthenolide inhibits STAT3 signaling by covalently targeting Janus kinases. Molecules 2018, 23, 1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efferth, T. From ancient herb to modern drug: Artemisia annua and artemisinin for cancer therapy. Semin. Cancer Biol. 2017, 46, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Slezakova, S.; Ruda-Kucerova, J. Anticancer Activity of Artemisinin and its Derivatives. Anticancer Res. 2017, 37, 5995–6003. [Google Scholar]
- Jia, L.; Song, Q.; Zhou, C.; Li, X.; Pi, L.; Ma, X. Dihydroartemisinin as a putative STAT3 inhibitor, suppresses the growth of head and neck squamous cell carcinoma by targeting Jak2/STAT3 signaling. PLoS ONE 2016, 11, 0147157. [Google Scholar] [CrossRef]
- Wang, D.; Zhong, B.; Li, Y.; Liu, X. Dihydroartemisinin increases apoptosis of colon cancer cells through targeting Janus kinase 2/signal transducer and activator of transcription 3 signaling. Oncol. Lett. 2017, 15, 1949–1954. [Google Scholar] [CrossRef] [Green Version]
- Rasul, A.; Khan, M.; Ali, M.; Li, J.; Li, X. Targeting apoptosis pathways in cancer with alantolactone and isoalantolactone. Sci. World J. 2013, 2013, 248532. [Google Scholar] [CrossRef] [Green Version]
- Pal, H.C.; Sehar, I.; Bhushan, S.; Gupta, B.D.; Saxena, A.K. Activation of caspases and poly (ADP-ribose) polymerase cleavage to induce apoptosis in leukemia HL-60 cells by Inula racemosa. Toxicol. In Vitro 2010, 24, 1599–1609. [Google Scholar] [CrossRef]
- Khan, M.; Yi, F.; Rasul, A.; Li, T.; Wang, N.; Gao, H. Alantolactone induces apoptosis in glioblastoma cells via GSH depletion, ROS generation, and mitochondrial dysfunction. IUBMB Life 2012, 64, 783–794. [Google Scholar] [CrossRef]
- Chun, J.; Li, R.J.; Cheng, M.S.; Kim, Y.S. Alantolactone selectively suppresses STAT3 activation and exhibits potent anticancer activity in MDA-MB-231 cells. Cancer Lett. 2015, 357, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-caryophyllene and β-caryophyllene oxide-natural compounds of anticancer and analgesic properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef]
- Kim, C.; Cho, S.K.; Kapoor, S.; Kumar, A.; Vali, S.; Abbasi, T.; Ahn, K.S. β-caryophyllene oxide inhibits constitutive and inducible STAT3 signaling pathway through induction of the SHP-1 protein tyrosine phosphatase. Mol. Carcinog. 2014, 53, 793–806. [Google Scholar] [CrossRef] [PubMed]
- Mondal, A.; Gandhi, A.; Fimognari, C.; Atanasov, A.G.; Bishayee, A. Alkaloids for cancer prevention and therapy: Current progress and future perspectives. Eur. J. Pharmacol. 2019, 858, 172472. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Oesch, F. Repurposing of plant alkaloids for cancer therapy: Pharmacology and toxicology. Semin. Cancer Biol. 2019, in press. [Google Scholar] [CrossRef]
- Clark, R.; Lee, S.H. Anticancer Properties of Capsaicin Against Human Cancer. Anticancer Res. 2016, 36, 837–843. [Google Scholar] [PubMed]
- Chapa-Oliver, A.M.; Mejía-Teniente, L. Capsaicin: From plants to a cancer-suppressing agent. Molecules 2016, 21, 931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Wang, D.; Huang, J.; Hu, Y.; Xu, Y. Application of capsaicin as a potential new therapeutic drug in human cancers. J. Clin. Pharm. Ther. 2020, 45, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Li, D.; Chu, C.; Li, X.; Wang, X.; Jia, Y.; Hua, H.; Xu, F. Antiproliferative Effects of Alkaloid Evodiamine and Its Derivatives. Int. J. Mol. Sci. 2018, 19, 3403. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.X.; Li, X.P.; Zhou, P.; Li, D.Y.; Lyu, X.T.; Chen, Y.; Lyu, Y.-W.; Tian, K.; Yuan, D.-Z.; Ran, J.-H.; et al. Evodiamine induces apoptosis in SMMC-7721 and HepG2 cells by suppressing NOD1 signal pathway. Int. J. Mol. Sci. 2018, 19, 3419. [Google Scholar] [CrossRef] [Green Version]
- Blažević, T.; Heiss, E.H.; Atanasov, A.G.; Breuss, J.M.; Dirsch, V.M.; Uhrin, P. Indirubin and Indirubin Derivatives for Counteracting Proliferative Diseases. Evid. Based Complement. Alternat. Med. 2015, 2015, 654098. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Merz, K.H. The Role of Indirubins in Inflammation and Associated Tumorigenesis. Adv. Exp. Med. Biol. 2016, 929, 269–290. [Google Scholar] [PubMed]
- Chen, L.; Wang, J.; Wu, J.; Zheng, Q.; Hu, J. Indirubin suppresses ovarian cancer cell viabilities through the STAT3 signaling pathway. Drug. Des. Devel. Ther. 2018, 12, 3335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.H.; Li, T.; Fong, M.; Chen, X.; Chen, X.J.; Wang, Y.T.; Huang, M.Q.; Lu, J.J. Saponins from Chinese Medicines as Anticancer Agents. Molecules 2016, 21, 1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koczurkiewicz, P.; Klaś, K.; Grabowska, K.; Piska, K.; Rogowska, K.; Wójcik-Pszczoła, K.; Podolak, I.; Galanty, A.; Michalik, M.; Pękala, E. Saponins as chemosensitizing substances that improve effectiveness and selectivity of anticancer drug-Minireview of in vitro studies. Phytother. Res. 2019, 33, 2141–2151. [Google Scholar] [CrossRef]
- Cheong, D.H.J.; Arfuso, F.; Sethi, G.; Wang, L.; Hui, K.M.; Kumar, A.P.; Tran, T. Molecular targets and anti-cancer potential of escin. Cancer Lett. 2018, 422, 1–8. [Google Scholar] [CrossRef]
- Tan, S.M.L.; Li, F.; Rajendran, P.; Kumar, A.P.; Hui, K.M.; Sethi, G. Identification of beta-escin as a novel inhibitor of signal transducer and activator of transcription 3/Janus-activated kinase 2 signaling pathway that suppresses proliferation and induces apoptosis in human hepatocellular carcinoma cells. J. Pharmacol. Exp. Ther. 2010, 334, 285–293. [Google Scholar] [CrossRef]
- Blanco-Vaca, F.; Cedo, L.; Julve, J. Phytosterols in cancer: From molecular mechanisms to preventive and therapeutic potentials. Curr. Med. Chem. 2019, 26, 6735–6749. [Google Scholar] [CrossRef]
- Jiang, L.; Zhao, X.; Xu, J.; Li, C.; Yu, Y.; Wang, W.; Zhu, L. The Protective Effect of Dietary Phytosterols on Cancer Risk: A Systematic Meta-Analysis. J. Oncol. 2019, 2019, 7479518. [Google Scholar] [CrossRef] [Green Version]
- Raju, J.; Mehta, R. Cancer chemopreventive and therapeutic effects of diosgenin, a food saponin. Nutr. Cancer 2009, 61, 27–35. [Google Scholar] [CrossRef]
- Nagulapalli Venkata, K.C.; Swaroop, A.; Bagchi, D.; Bishayee, A. A small plant with big benefits: Fenugreek (Trigonellafoenum graecum L.) for disease prevention and health promotion. Mol. Nutr. Food Res. 2017, 61, 1600950. [Google Scholar] [CrossRef]
- Sethi, G.; Shanmugam, M.K.; Warrier, S.; Merarchi, M.; Arfuso, F.; Kumar, A.P.; Bishayee, A. Pro-apoptotic and anti-cancer properties of diosgenin: A comprehensive and critical review. Nutrients 2018, 10, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Fernandez, P.P.; Rajendran, P.; Hui, K.M.; Sethi, G. Diosgenin, a steroidal saponin, inhibits STAT3 signaling pathway leading to suppression of proliferation and chemosensitization of human hepatocellular carcinoma cells. Cancer Lett. 2010, 292, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Bian, D.; Li, Z.; Ma, H.; Mu, S.; Ma, C.; Cui, B.; Gao, L.; Zhao, J. Effects of diosgenin on cell proliferation induced by IGF-1 in primary human thyrocytes. Arch. Pharmacal. Res. 2011, 34, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Merdivan, S.; Lindequist, U. Ergosterol Peroxide: A Mushroom-Derived Compound with Promising Biological Activities—A Review. Int. J. Med. Mushrooms 2017, 19, 93–105. [Google Scholar] [CrossRef]
- Shishodia, S.; Azu, N.; Rosenzweig, J.A.; Jackson, D.A. Guggulsterone for Chemoprevention of Cancer. Curr. Pharm Des. 2016, 22, 294–306. [Google Scholar] [CrossRef]
- Bhat, A.A.; Prabhu, K.S.; Kuttikrishnan, S.; Krishnankutty, R.; Babu, J.; Mohammad, R.M.; Uddin, S. Potential therapeutic targets of Guggulsterone in cancer. Nutr. Metab. 2017, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Kunnumakkara, A.B.; Banik, K.; Bordoloi, D.; Harsha, C.; Sailo, B.L.; Padmavathi, G.; Roy, N.K.; Gupta, S.C.; Aggarwal, B.B. Googling the Guggul (Commiphora and Boswellia) for Prevention of Chronic Diseases. Front. Pharmacol. 2018, 9, 686. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.S.; Sethi, G.; Sung, B.; Goel, A.; Ralhan, R.; Aggarwal, B.B. Guggulsterone, a farnesoid X receptor antagonist, inhibits constitutive and inducible STAT3 activation through induction of a protein tyrosine phosphatase SHP-1. Cancer Res. 2008, 68, 4406–4415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Morales, L.D.; Jang, I.S.; Cho, Y.Y.; Kim, D.J. Protein tyrosine phosphatases as potential regulators of STAT3 signaling. Int. J. Mol. Sci. 2018, 19, 2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touré, A.; Xueming, X. Flaxseed lignans: Source, biosynthesis, metabolism, antioxidant activity, bio-active components, and health benefits. Compr. Rev. Food Sci. Food Saf. 2010, 9, 261–269. [Google Scholar] [CrossRef]
- Gao, Q.; Yang, M.; Zuo, Z. Overview of the anti-inflammatory effects, pharmacokinetic properties and clinical efficacies of arctigenin and arctiin from Arctium lappa L. Acta Pharmacol Sin. 2018, 39, 787–801. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, C.; Lee, J.; Um, J.Y.; Sethi, G.; Ahn, K.S. Arctiin is a pharmacological inhibitor of STAT3 phosphorylation at tyrosine 705 residue and potentiates bortezomib-induced apoptotic and anti-angiogenic effects in human multiple myeloma cells. Phytomedicine 2019, 55, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Romagnolo, D.F.; Davis, C.D.; Milner, J.A. Phytoalexins in cancer prevention. Front. Biosci. 2012, 17, 2035–2058. [Google Scholar] [CrossRef]
- Chripkova, M.; Zigo, F.; Mojzis, J. Antiproliferative Effect of Indole Phytoalexins. Molecules 2016, 21, 1626. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kim, C.; Sethi, G.; Ahn, K.S. Brassinin inhibits STAT3 signaling pathway through modulation of PIAS-3 and SOCS-3 expression and sensitizes human lung cancer xenograft in nude mice to paclitaxel. Oncotarget 2015, 6, 6386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bousoik, E.; Aliabadi, H.M. “Do we know Jack” about JAK? A closer look at JAK/STAT signaling pathway. Front. Oncol. 2018, 8, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemical Name | Sources | Anticancer Effect | Mechanism of Action | EC50/IC50 | References |
|---|---|---|---|---|---|
| Resveratrol | Vitis vinifera L., Vaccinium macrocarpon Aiton and Arachis hypogaea L. | Inhibited proliferation in human epidermoid carcinoma (A431) cells | ┴ Phosphorylation of JAK that prevented STAT1 phosphorylation | 40 µM | Madan et al., 2008 [86] |
| Inhibited proliferation in human multiple myeloma (U266 and RPMI 8226) cells | ┴ Constitutive and inducible STAT3 activation | 50 and 50 µM | Bhardwaj et al., 2007 [87] | ||
| Inhibited tumor growth, induction of cytotoxicity, cell cycle arrest of v-Src-transformed mouse fibroblasts (NIH3T3/v-Src) at G0-G1 phase. Showed cytotoxicity in human breast cancer (MDA-MB-231), pancreatic carcinoma (Panc-1), and prostate carcinoma (DU145) cells | ┴ Src tyrosine kinase activity; ┴ constitutive STAT3 activation | 40, 70. and 25 µM | Kotha et al., 2006 [88] | ||
| Curcumin | Curcuma longa L. | Induction of cytotoxicity in human multiple myeloma (U266, RPMI 8226, and MM.1S) cells | ┴ Constitutive and IL-6-inducible STAT3 phosphorylation; ┴ IFN-inducible STAT1 phosphorylation | 13.8, 12.7 and 10 µM | Bharti et al., 2003 [95] |
| Inhibited proliferation in human small cell lung cancer (NCI-H446 and NCI-1688) cells | ┴ STAT3 phosphorylation; ↓ STAT3 downstream gene expression | 15 and 15 µM | Yang et al., 2012 [96] | ||
| Antitumor activity in T-cell leukemia, treatment causes growth arrest with increase in apoptosis | ┴ Activation of the JAK/STAT pathway; ↓ JAK and STAT phosphorylation | Not specified | Rajasingh et al., 2006 [97]; Kunnumakkara et al., 2017 [98] | ||
| Ascochlorin | Ascochyta viciae | Inhibited cell migration and invasion in U373MG and A172 cancer cells | ┴ MMP-9 expression, ┴ MMP-2 gelatinolytic action and expression; ┴ JAK2/STAT3 phosphorylation, cancer cell migration, and nuclear translocation of STAT3 | 10 μM | Cho et al., 2018 [100] |
| Bergamottin | Grape juice, lime, lemon, and bergamot oils | Antitumor activity in U266 cells, induction of substantial apoptosis at sub-G1 stage | ┴ Constitutive STAT3 activation; ┴ Phosphorylation of JAK 1/2 and c-Src | 100 μM | Kim et al., 2014 [102] |
| Capillarisin | Artemisia capillaries Thunb. | Negative regulator of growth and metastasis in human multiple myeloma cells; induces apoptosis; downregulated the expression level of various STAT3-regulated proteins | ┴ Constitutive and inducible STAT3 activation at tyrosine 705; ┴ STAT3 phosphorylation; ┴ Activation of JAK1, JAK2, and c-Src kinases | Not mentioned | Lee et al., 2014 [103] |
| Bavachin | Psoralea corylifolia Linn. [Cullen corylifolium (L.) Medik.] | Induced cytotoxicity in multiple myeloma cell lines; induced apoptosis by activation of caspase-3 and caspase-9; inhibited activation of NF-κB, expression levels of Bcl-2, X-linked inhibitor of apoptosis protein (XIAP), survivin, and B cell lymphoma-extra large (Bcl-xL) | ┴ Activation of STAT3; ┴ Phosphorylation of STAT3 | 10 μM | Takeda et al., 2018 [104] |
| Epigallocatechin-3-gallate (EGCG) | Camellia sinensis (L.) Kuntze | Anticancer activity in human pancreatic (AsPC-1 and PANC-1), breast (T47D), head and neck cancer (YCU-H861) cells | ┴ Phosphorylation and expression of both JAK3 and STAT3 proteins | 40, 40, 14.17 µM, 1 µg/mL | Tang et al., 2012 [109] |
| Emodin | Rheum palmatum L. | Stimulated the antiproliferation activity of interferon α/β in cervical carcinoma cell line (HeLa) and antitumor activity in Huh7 (hepatocellular cancer cell)-bearing mice in vivo | ┴ STAT3 activation, ┴ 26S proteasome activity ↑ STAT1 phosphorylation, ┴ STAT3 phosphorylation | Not specified, 1.22 μM | He et al., 2016 [117] |
| Chalcone | Malus domestica (Suckow) Borkh. and Solanum lycopersicum L. | Induced cytotoxicity in Bovine aortic endothelial cells (BAEC) | ┴ IL-6-induced and LPS-induced STAT3 phosphorylation | 0.0069 µM | Liu et al., 2007 [120] |
| Formononetin | Astragalus mongholicus Bunge | Inhibited proliferation and invasion of colon carcinoma cell lines HCT116 and SW1116, cell cycle arrest at the G0/G1 stage | ┴ STAT3 signaling pathway; ┴ STAT3 phosphorylation | Not specified | Wang et al., 2018 [123] |
| Garcinol | Garcinia indica (Thouars.) Choisy | Induced cytotoxicity in hepatocellular carcinoma cells, cell cycle arrest at G0/G1 phase | ┴ Constitutive and IL-6 inducible STAT3 activation | Not specified | Sethi et al., 2014 [126] |
| Cardamonin | Alpinia conchigera Griff., and Alpinia hainanensis K. Schum. | Antitumor activity in U87 cells (in vitro) and CD133+ GSCs (in vitro and in vivo); induced apoptosis | ┴ STAT3 signaling pathway; ┴ STAT3 activation; ┴ downstream STAT3 gene expressions (i.e., Bcl-xL, Bcl-2, Mcl-1, survivin, and VEGF) | Not specified | Wu et al., 2015 [128] |
| Caffeic acid or its derivative 3-(3,4-dihydroxy-phenyl)-acrylic acid 2-(3,4-dihydroxy-phenyl)-ethyl ester (CADPE) | Camellia sinensis L. Kuntze., and Coffea arabica L. | Induction of cytotoxicity in human renal carcinoma (Caki-1) cells | ┴ STAT3 phosphorylation; ┴ Src tyrosine kinase. | 30 µM | Jung et al., 2007 [131] |
| Silibinin | Silybum marianum (L.) Gaertn. | Induction of cytotoxicity in human prostate cancer (DU145) cells | ┴ Constitutively active STAT3, ↑ apoptosis and ┴ constitutive STAT3–DNA binding | 50 µM | Agarwal et al., 2007 [135] |
| Suppressed transcriptional function in urethane-induced lung tumors in A/J mice | ┴ STAT3 phosphorylation | Not specified | Tyagi et al., 2009 [136] | ||
| Butein | Toxicodendron vernicifluum (Stokes) F.A. Barkley and Butea monosperma (Lam.) Kuntze | Exhibited antitumor activity in human hepatocellular carcinoma (HepG2 and SNU-387) | ┴ Constitutive and IL-6- induced STAT3 activation by inactivating JAK1 and c-Src. | 50 and 50 µM | Bhutani et al., 2007 [139]; Rajendran et al., 2011 [140] |
| 5,7-Dihydroxyflavone | Passiflora incarnate L., Passiflora caerulea L., and Oroxylum indicum (L.) Kurz | Inhibited proliferation HepG2 tumor xenografts in vivo | ↓ Phosphorylation of STAT3 | 20 µM | Zhang et al., 2013 [143] |
| Honokiol | Magnolia officinalis Rehder & E. H. Wilson and Magnolia grandiflora L. | Inhibited proliferation, induced apoptosis in human leukemic cell lines (HEL and THP1), multiple myeloma cells (U266) and murine myeloid cell (32D) | ┴ Constitutive and inducible STAT3 activation; ┴ mRNA levels of STAT3 target genes in a concentration-dependent manner; ↓ Nuclear translocation of STAT3 | 40 µM | Bi et al., 2015 [149] |
| Casticin | Achillea millefolium L., Artemisia abrotanum L., Vitex trifolia subsp. Litoralis Steenis, Camellia sinensis (L.) Kuntz, Centipeda minima (L.) A.Braun & Asch, Clausena excavate Burm.f., Crataegus pinnatifida Bunge, Croton betulaster Müll Arg., Daphne genkwa Siebold & Zucc., Ficus microcarpa L.f., Nelsonia canescens (Lam.) Spreng., Pavetta crassipes K. Schum., Vitex trifolia subsp. Litoralis steenis, Vitex agnus-castus L., Vitex negundo L. and Vitex trifolia L. | Inhibited proliferation, induced apoptosis, cell cycle arrest at G2/M phase in colon (Panc-1), breast (MCF-7), lung (A549), gastric (SGC-7901), ovarian (SKOV3), liver (HepG2), leukemia (K562) cancer cells | ┴ Constitutively active STAT3 and modulates STAT3 activation by modifying upstream STAT3 regulator activity. | 10, 8.5, 14.3, 1, 2.18, 30, 5.95 µM | Chen et al., 2011 [152]; Zeng et al., 2012 [153] |
| Induced apoptosis in 786-O, YD-8, and HN-9 cancer cells | ┴ constitutively activation of STAT3; Modulated STAT3 activation by altering the activity of upstream STAT3 regulators, and abrogated IL-6-induced STAT3 activation; ┴ JAK/STAT pathway | 5 μM | Lee et al., 2019 [154] | ||
| Apigenin | Petroselinum crispum (Mill.) Fuss., Apium graveolens L., and Matricaria chamomilla L. | Anticancer and antitumor activity in colon cancer (HCT-116) cells | ┴ Phosphorylation of STAT3 and consequently downregulated the antiapoptotic proteins Bcl-xL and Mcl-1 | 47.33 µM | Ozbey et al., 2018 [25]; Maeda et al., 2018 [157] |
| Anticancer activity in BT-474 (breast cancer) cells | ┴ JAK/STAT pathway ┴ STAT3 nuclear accumulation ┴ Phosphorylation of JAK1/2, and STAT3 | Not specified | Ozbey et al., 2018 [25] | ||
| Wedelolactone | Eclipta prostrate (L.) L. and Sphagneticola calendulacea (L.) Pruski | Inhibited proliferation, induced apoptosis, causes cell cycle arrest at S and G2/M phases in breast (MDA-MB-231) and HepG2 cancer cells | ┴ STAT1 dephosphorylation and prolonging STAT1 activation, ┴ T-cell protein tyrosine phosphatase | Not specified in MDA-MB-231; 20 µM (EC50) in HepG2 cells | Benes et al., 2011 [159]; Benes et al., 2012 [160]; Chen et al., 2013 [161] |
| Phytochemical Name | Sources | Anticancer Effect | Mechanism of Action | EC50 /IC50 | References |
|---|---|---|---|---|---|
| Cucurbitacin B | Hemsleya endecaphylla C.Y. Wu | Induced cytotoxicity in human pancreatic cancer (MiaPaCa-2, AsPC-1) cells | ┴ JAK2, ┴ STAT3, and ┴ STAT5 activation | 0.278, 0.017 μM | Thoennissen et al., 2009 [168]; Zhou et al., 2017 [169] |
| Induced cytotoxicity in leukemia K562 cells | ┴ STAT3 activation | 50 µM | Chan et al., 2010 [170] | ||
| Cucurbitacin E | Wilbrandia ebracteate Cogn. | Induced cytotoxicity in human bladder cancer (T24) cells | ↓ Levels of phosphorylated STAT3 (p-STAT3) | 1012 nM | Huang et al., 2012 [171] |
| Cucurbitacin I | Cucumis melo L. | Induced cytotoxicity in human lung adenocarcinoma (A549) cells | ↓ Phosphotyrosine STAT3, ↓ JAK levels. ┴ STAT3-DNA binding; ┴ STAT3-mediated gene transcription | 500 nM | Blaskovich et al., 2003 [172] |
| Induced cytotoxicity in glioblastoma multiforme cells, G2/M cell cycle arrest by downregulating cyclin B1 and cdc2 expression | ┴ JAK/STAT pathway; ↓ Phosphorylated STAT3 levels | Not specified | Su et al., 2014 [173] | ||
| Inhibitory activity in Sézary (Sz) syndrome and anaplastic large cell lymphoma | ┴ JAK/STAT pathway; ↓ Phosphorylated STAT3 levels | 30 μM | van Kester et al., 2008 [174]; Shi et al., 2006 [175] | ||
| Cucurbitacin Q | Picrorhiza kurrooa Royle ex Benth. | Antiproliferative effect in human non-small-cell lung carcinoma (A549) cells | ↓ STAT3 but not JAK2 activation | 3.7 µM | Sun et al., 2005 [176] |
| Andrographolide | Andrographis paniculata (Burm.f.) Nees. | Enhanced anticancer activity of doxorubicin | ┴ STAT3 signaling pathway; ┴ Constitutively actuated and IL-6-induced phosphorylation of STAT3 and subsequent nuclear translocation | Not specified | Zhou et al., 2010 [179] |
| Betulinic acid | White-barked birch plants | Inhibited proliferation and induced apoptosis in human multiple myeloma (U266) cells; Cell cycle arrest at sub-G1 stage; downregulated the expression level of STAT3-regulated gene products | ┴ STAT3 signaling, ┴ STAT3-directed gene expression; ┴ Constitutive activation of STAT3, Src kinase, and JAK1/2 | Not specified | Pandey et al., 2010 [182] |
| γ-Tocotrienol | Elaeis guineensis Jacq. | Anticancer effect in human hepatocellular carcinoma (HepG2, C3A, Hep3B, SNU-387, and PLC/PRF5) cells | ┴ Both constitutive and inducible activation of STAT3; ┴ Phosphorylation of JAK1, JAK2, and c-Src | Not specified | Rajendran et al., 2011 [185]; Banerjee et al., 2019 [186] |
| Cryptotanshinone | Salvia miltiorrhiza Bunge | Induced cytotoxicity in human prostate cancer (DU145) cells | ┴ Phosphorylation of STAT3 through an independent mechanism of JAK2 phosphorylation | 7.59 µM | Shin et al., 2009 [188] |
| Nimbolide | Azadirachta indica A. Juss. | Induced cytotoxicity and cell cycle arrest at G1–S stage in glioblastoma multiforme cells. It downregulates Bcl2 | ┴ STAT3 pathway, ┴ STAT3 phosphorylation | Not specified | Karkare et al., 2014 [191]; Zhang et al., 2016 [192] |
| Celastrol | Tripterygium wilfordii Hook.f. | Induced cytotoxicity in human multiple myeloma (U266, RPMI 8226 and RPMI-8226-LR-5) cells | ┴ Phosphorylation of STAT3 as well as STAT3-mediated IL-17 expression | Not specified | Kannaiyan et al., 2011 [193] |
| Inhibited differentiation and cell proliferation in multiple myeloma cells | ┴ STAT3 phosphorylation; ┴ STAT3-mediated IL-17 expression ┴ T-helper 17 (Th17) | Not specified | Banerjee et al., 2019 [186] | ||
| Ursolic acid | Mirabilis jalapa L. | Induced cytotoxicity in human prostate cancer (DU145 and LNCaP) cells | ┴ Activation of constitutive and inducible STAT3; ↓ phosphorylation of Src and JAK2 | 80 and 47 µM | Shanmugam et al., 2011b [198] |
| Inhibited tumor growth in prostate xenograft tumor in TRAMPmice in vivo | ┴ JAK/STAT signaling; ┴ activation of STAT3 | Not specified | Shanmugam et al., 2011 [198] | ||
| Brusatol | Brucea javanica (L.) Merr. | Induced cytotoxicity in head and neck squamous cell carcinoma | ┴ STAT3 signaling pathway; ┴ Activation of STAT3 as well as JAK1/2, and Src | Not specified | Lee et al., 2019 [199] |
| Oridonin | Isodon rubescens (Hemsl.) H. Hara | Inhibited proliferation and induced apoptosis and cell cycle arrest at G2/M phase in breast (MCF-7), leukemia (K562), lung (A549), prostate (PC-3), liver (Bel7402), gastric (BGC823), and uterine cervix cancer (HeLa) cells The viability of BxPC-3 (human pancreatic cancer) cells is reduced on treatment | ┴ STAT3 signaling pathway; ↓ IL-6, ↓ STAT3, and ↓ phospho-STAT3 expression level, ↓ p21, ↓ cyclin A, ↓ cyclin B1, ↓ cyclin D1, ↓ VEGF, and ↓ MMP-2 | 18.4, 4.3, 18.6, 15.2, 7.6, 13.7 μg/mL 19.32 μg/mL | Bu et al., 2012 [202]; Chen et al., 2008 [203]; Chen et al., 2012 [204]; Chen et al., 2014 [205] |
| Thymoquinone | Nigella sativa L. | Induced cytotoxicity in multiple myeloma (U266 and RPMI 8226) cells. | ┴ STAT3 phosphorylation | 15 and 15 µM | Zhu et al., 2016 [208]; Li et al., 2010 [209] |
| Parthenolide | Tanacetum parthenium (L.) Sch. Bip., Tanacetum vulgare L., Centaurea ainetensis Boiss., Tanacetum larvatum (Pant.) Hayek., and Helianthus annuus L. | ┴ Proliferation, ↑ apoptosis, cell cycle arrest at G2/M phase in breast (MCF-7), skin (MDMB-231), melanoma (LCC9), malignant glioma (ABCB5+), epidermal tumorigenesis (A375), liver (1205Lu), gastric cancer (WM793) cells | ┴ IL-6-induced STAT3 phosphorylation; ┴ JAK2 kinase activity | 9.54, 10 µM, 600 nM, 12, 2.9, 6, 12 µM | Cheng et al., 2011 [211]; Liu &Xie, 2010 [212]; Shanmugam et al., 2011 [213] |
| Inhibited JAK2 kinase activity in MDA-MB-231 cells | ┴ IL-6-induced STAT3 phosphorylation | Not specified | Liu et al., 2018 [214] | ||
| Dihydroartemisinin | Artemisia annua L. | Inhibited tumor cell growth in human head and neck cancer (FaDu), liver cancer (Hep-G2), colon cancer (HCT-116), and tongue cancer (Cal-27) cells | ┴ JAK2/STAT3 signaling activation; ↓ targeted proteins | 160, 80, 25, 80 µM | Jia et al., 2016 [217]; Wang et al., 2017 [218] |
| Alantolactone | Aucklandia costus Falc., Inula helenium L., Inula japonica Thunb., and Inula racemose Hook. f. | Antiproliferative effect in glioblastoma (U87), colon (HCT-8), leukemia (HL-60), liver (HepG2), lung cancer (A549) cells | ┴ Both constitutive and inducible STAT3 activation at tyrosine 705; ┴ STAT3 translocation to the nucleus; ┴ DNA-binding, and ┴ STAT3 target gene expression | 135.27 µM, 5 µg/mL, 1.1, 40, 8.39 µM | Pal et al., 2010 [220]; Khan et al., 2012 [221]; Chun et al., 2015 [222] |
| β-Caryophyllene oxide | Cannabis sativa L., Humulus lupulus L., Origanum vulgare L., Psidium guajava L., Salvia Rosmarinus Spenn., and Syzygium aromaticum (L.) Merr. & L. M. Perry | Inhibited proliferation in multiple myeloma, prostate, and breast cancer cell lines | ┴ STAT3 pathway; ┴ STAT3 activation as well as JAK 1/2 and c-Src | Not specified | Kim et al., 2014 [224] |
| Phytochemical Class | Phytochemical Name | Sources | Anticancer Effect | Mechanism of Action | EC50/IC50 | References |
|---|---|---|---|---|---|---|
| Alkaloids | Capsaicin | Capsicum frutescens L. | Induction of cytotoxicity in human multiple myeloma (U266 and MM.1S) cells | ┴ Constitutive and IL-6-induced activation of STAT3; ┴ JAK1 and c-Src activation | 50 and 50 µM | Bhutani et al., 2007 [139] |
| Evodiamine | Tetradium ruticarpum (A. Juss.) T. G. Hartley | Inhibited proliferation, and induced apoptosis; Cell cycle arrest at G2/M phase in murine Lewis lung (LLC), hepatocellular (HepG2), leukemia (K562), gastric (SGC-7901), colon (COLO-205) cancer cells. | ↓ Constitutive and IL-6-induced activation of STAT3 tyrosine 705 (Tyr705); ↓ JAK2, Src and ERK1/2; ┴ STAT3–DNA binding activity | 113, 8.516, 5, 10, 27.15 µM | Yang et al., 2013 [81] | |
| Indirubin | Angelica sinensis (Oliv.) Diels | Reduced cell viability in human prostate and breast cancer cells; induced apoptosis; Cell cycle arrest at G1/S or G2/M | ┴ STAT3 signaling; ┴ STAT3 phosphorylation | ~4 µM for each cell line | Chen et al., 2018 [234] | |
| Saponins | β-Escin | Aesculus hippocastanum L. | Induction of cytotoxicity in human hepatocellular carcinoma (HepG2, PLC/PRF5, and HUH-7) cells | ┴ Activation of STAT3 and induced expression of SHP-1; ┴ Phosphorylation of JAK1, JAK2, and c-Src | Not specified | Tan et al., 2010 [238] |
| Steroid | Diosgenin | Trigonella foenum-graecum L. | Induced cytotoxicity in human hepatocellular carcinoma (C3A, HUH-7, and HepG2) cells | ┴ Constitutive and inducible activation of STAT3 | 100, 100 and 50 µM | Li et al., 2010 [244] |
| Ergosterol peroxide | Agaricus bisporus | Induced cytotoxicity in human multiple myeloma (U266) cells | ┴ Phosphorylation of JAK2; ┴ phosphorylation; ┴ DNA binding activity; ┴ Nuclear translocation of STAT3 | Not specified | Rhee et al., 2012 [80] | |
| Guggulsterone | Commiphora mukul (Hook. ex Stocks) Engl. | Induced cytotoxicity in human multiple myeloma (U266) cells | ┴ Constitutive andIL-6-induced STAT3; ┴ Phosphorylation of JAK2 and Src | 25 µM | Ahn et al., 2008 [250] | |
| Lignan | Arctiin | Centaurea imperialis Hausskn. ex Bornm., Arctium lappa L., Forsythia viridissima Lindl., Himalaiella heteromalla (D. Don) Raab-Straube., and Trachelospermumas iaticum (Siebold & Zucc.) Nakai | Inhibited proliferation, cell cycle arrest at G2/M stage in human multiple myeloma cells | ┴ STAT3 phosphorylation in tyrosine 705; ┴ Constitutive enactment of Src phosphorylation and JAKs 1/2; ↑ mRNA, ↑ protein levels of protein tyrosine phosphatase ε; ┴ STAT3 regulated gene products | Not specified | Lee et al., 2019b [254] |
| Phytoalexin | Brassinin | Brassica rapa L. | Induced apoptosis in lung cancer cells (A549) in an in vivo mouse model | ┴ STAT3 activation; ┴ Both constitutive and IL-6-inducible STAT3 activation; ┴ Phospho-STAT3, ┴ Ki-67 and ┴ CD31 | Not specified | Lee et al., 2015 [257] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bose, S.; Banerjee, S.; Mondal, A.; Chakraborty, U.; Pumarol, J.; Croley, C.R.; Bishayee, A. Targeting the JAK/STAT Signaling Pathway Using Phytocompounds for Cancer Prevention and Therapy. Cells 2020, 9, 1451. https://doi.org/10.3390/cells9061451
Bose S, Banerjee S, Mondal A, Chakraborty U, Pumarol J, Croley CR, Bishayee A. Targeting the JAK/STAT Signaling Pathway Using Phytocompounds for Cancer Prevention and Therapy. Cells. 2020; 9(6):1451. https://doi.org/10.3390/cells9061451
Chicago/Turabian StyleBose, Sankhadip, Sabyasachi Banerjee, Arijit Mondal, Utsab Chakraborty, Joshua Pumarol, Courtney R. Croley, and Anupam Bishayee. 2020. "Targeting the JAK/STAT Signaling Pathway Using Phytocompounds for Cancer Prevention and Therapy" Cells 9, no. 6: 1451. https://doi.org/10.3390/cells9061451