Rac1 S71 Mediates the Interaction between Rac1 and 14-3-3 Proteins


Abstract
:
{kind=link}
{kind=link}
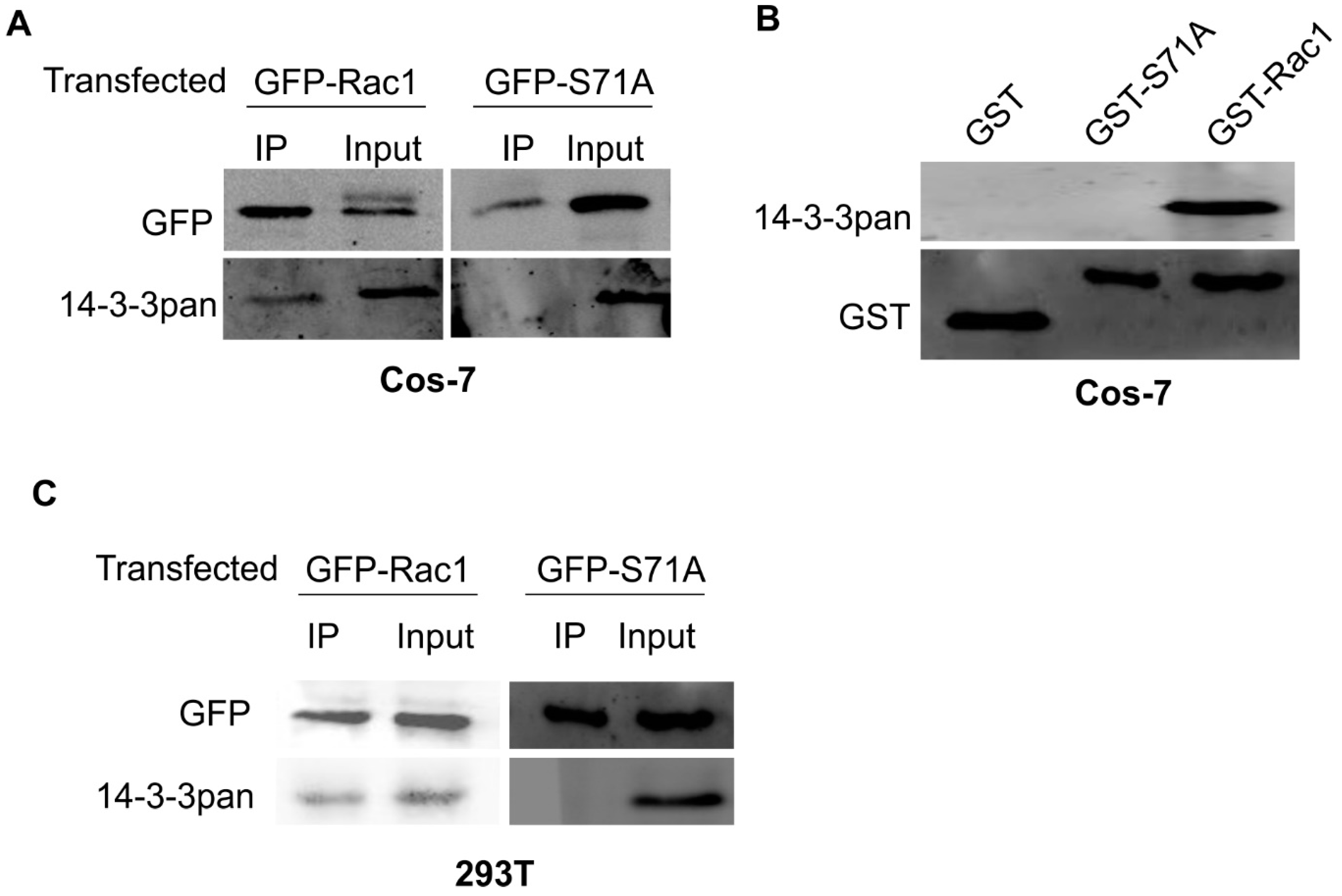{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatment
2.2. Transient Transfection
2.3. Antibodies and Chemicals
2.4. Plasmids
2.5. Expression and Purification of GST-Fusion Proteins
2.6. GST Pull-Down Assay
2.7. Rac1 Activity Assay
2.8. Immunoprecipitation
2.9. Immunoblotting
2.10. Fluorescence Microscopy
2.11. Artificial Network Analysis
2.12. Quantification and Statistical analysis
3. Results
3.1. Association between Rac1 and 14-3-3s in Response to EGF-Induced Akt Phosphorylation
3.2. Rac1 and 14-3-3s Interaction is Mediated by Rac1 S71
3.3. The Relationship between Rac1 Activity and Its Interaction with 14-3-3s
3.4. The Interaction between Rac1 and Various 14-3-3 Isoforms
3.5. The Effects of Rac1 and 14-3-3 Protein Interaction on the Subcellular Localization of Rac1 and 14-3-3s
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Matos, P.; Skaug, J.; Marques, B.; Beck, S.; Verissimo, F.; Gespach, C.; Boavida, M.G.; Scherer, S.W.; Jordan, P. Small GTPase Rac1: Structure, localization, and expression of the human gene. Biochem. Biophys. Res. Commun. 2000, 277, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Brandwein, D.; Wang, Z. Interaction between Rho GTPases and 14-3-3 proteins. Int. J. Mol. Sci. 2017, 18, 2148. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J. Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking. Trends Cell Biol. 2006, 16, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Pertz, O. Spatio-temporal Rho GTPase signaling—Where are we now? J. Cell Sci. 2010, 123, 1841–1850. [Google Scholar] [CrossRef] [PubMed]
- Abdrabou, A.; Wang, Z. Post-translational modification and subcellular distribution of Rac1: An update. Cells 2018, 7, 263. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M. Rho signalling at a glance. J. Cell Sci. 2004, 117, 5457–5458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchsbaum, R.J. Rho activation at a glance. J. Cell Sci. 2007, 120, 1149–1152. [Google Scholar] [CrossRef] [Green Version]
- Cherfils, J.; Zeghouf, M. Regulation of small GTPases by GEFs, GAPs, and GDIs. Physiol. Rev. 2013, 93, 269–309. [Google Scholar] [CrossRef]
- Symons, M.; Settleman, J. Rho family GTPases: More than simple switches. Trends Cell Biol. 2000, 10, 415–419. [Google Scholar] [CrossRef]
- Moon, S.; Zheng, Y. Rho GTPase-activating proteins in cell regulation. Trends Cell Biol. 2003, 13, 13–22. [Google Scholar] [CrossRef]
- Goncalves, V.; Henriques, A.F.; Pereira, J.F.; Neves Costa, A.; Moyer, M.P.; Moita, L.F.; Gama-Carvalho, M.; Matos, P.; Jordan, P. Phosphorylation of SRSF1 by SRPK1 regulates alternative splicing of tumor-related Rac1b in colorectal cells. RNA 2014, 20, 474–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Fu, X.; Chen, P.; Wu, P.; Fan, X.; Li, N.; Zhu, H.; Jia, T.T.; Ji, H.; Wang, Z.; et al. SPSB1-mediated HnRNP A1 ubiquitylation regulates alternative splicing and cell migration in EGF signaling. Cell Res. 2017, 27, 540–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Bi, F.; Zhou, X.; Zheng, Y. Rho GTPase regulation by miRNAs and covalent modifications. Trends Cell Biol. 2012, 22, 365–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worby, C.A.; Mattoo, S.; Kruger, R.P.; Corbeil, L.B.; Koller, A.; Mendez, J.C.; Zekarias, B.; Lazar, C.; Dixon, J.E. The fic domain: Regulation of cell signaling by adenylylation. Mol. Cell 2009, 34, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.F. Rho GTPases, their post-translational modifications, disease-associated mutations and pharmacological inhibitors. Small GTPases 2018, 9, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Loirand, G.; Guilluy, C.; Pacaud, P. Regulation of Rho proteins by phosphorylation in the cardiovascular system. Trends Cariovasc. Med. 2006, 16, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.; Kwon, D.; Chun, J.; Kim, J.; Kang, S. Akt protein kinase inhibits Rac1-GTP binding through phosphorylation at serine 71 of Rac1. J. Biol. Chem. 2000, 275, 423–428. [Google Scholar] [CrossRef]
- Schoentaube, J.; Olling, A.; Tatge, H.; Just, I.; Gerhard, R. Serine-71 phosphorylation of Rac1/Cdc42 diminishes the pathogenic effect of Clostridium difficile toxin A. Cell. Microbiol. 2009, 11, 1816–1826. [Google Scholar] [CrossRef]
- Schwarz, J.; Proff, J.; Havemeier, A.; Ladwein, M.; Rottner, K.; Barlag, B.; Pich, A.; Tatge, H.; Just, I.; Gerhard, R. Serine-71 phosphorylation of Rac1 modulates downstream signaling. PLoS ONE 2012, 7, e44358. [Google Scholar] [CrossRef]
- Chang, F.; Lemmon, C.; Lietha, D.; Eck, M.; Romer, L. Tyrosine phosphorylation of Rac1: A role in regulation of cell spreading. PLoS ONE 2011, 6, e28587. [Google Scholar] [CrossRef]
- Tong, J.; Li, L.; Ballermann, B.; Wang, Z. Phosphorylation of Rac1 T108 by extracellular signal-regulated kinase in response to epidermal growth factor: A novel mechanism to regulate Rac1 function. Mol. Cell. Biol. 2013, 33, 4538–4551. [Google Scholar] [CrossRef] [PubMed]
- Mhawech, P. 14-3-3 proteins—An update. Cell Res. 2005, 15, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Tseng, H.C.; Yang, C.M. Bradykinin-mediated cell proliferation depends on transactivation of EGF receptor in corneal fibroblasts. J. Cell. Physiol. 2012, 227, 1367–1381. [Google Scholar] [CrossRef] [PubMed]
- Rittinger, K.; Budman, J.; Xu, J.; Volinia, S.; Cantley, L.C.; Smerdon, S.J.; Gamblin, S.J.; Yaffe, M.B. Structural analysis of 14-3-3 phosphopeptide complexes identifies a dual role for the nuclear export signal of 14-3-3 in ligand binding. Mol. Cell 1999, 4, 153–166. [Google Scholar] [CrossRef]
- Yaffe, M.B.; Rittinger, K.; Volinia, S.; Caron, P.R.; Aitken, A.; Leffers, H.; Gamblin, S.J.; Smerdon, S.J.; Cantley, L.C. The structural basis for 14-3-3:phosphopeptide binding specificity. Cell 1997, 91, 961–971. [Google Scholar] [CrossRef]
- Coblitz, B.; Wu, M.; Shikano, S.; Li, M. C-terminal binding: An expanded repertoire and function of 14-3-3 proteins. FEBS Lett. 2006, 580, 1531–1535. [Google Scholar] [CrossRef] [Green Version]
- Shikano, S.; Coblitz, B.; Sun, H.; Li, M. Genetic isolation of transport signals directing cell surface expression. Nat. Cell Biol. 2005, 7, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Sluchanko, N.N.; Gusev, N.B. Moonlighting chaperone-like activity of the universal regulatory 14-3-3 proteins. FEBS J. 2017, 284, 1279–1295. [Google Scholar] [CrossRef]
- Thomas, D.; Guthridge, M.; Woodcock, J.; Lopez, A. 14-3-3 protein signaling in development and growth factor responses. Curr. Top. Dev. Biol. 2005, 67, 285–303. [Google Scholar]
- Gardino, A.K.; Yaffe, M.B. 14-3-3 proteins as signaling integration points for cell cycle control and apoptosis. Semin. Cell Dev. Biol. 2011, 22, 688–695. [Google Scholar] [CrossRef]
- Freeman, A.K.; Morrison, D.K. 14-3-3 proteins: Diverse functions in cell proliferation and cancer progression. Semin. Cell Dev. Biol. 2011, 22, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Fan, Z.; Ding, M.; Zhang, H.; Mu, L.; Ding, Y.; Zhang, Y.; Jia, B.; Chen, L.; Chang, Z.; et al. An EGFR/PI3K/Akt axis promotes accumulation of the Rac1-GEF Tiam1 that is critical in EGFR-driven tumorigenesis. Oncogene 2015, 34, 5971–5982. [Google Scholar] [CrossRef] [PubMed]
- Moores, S.L.; Selfors, L.M.; Fredericks, J.; Breit, T.; Fujikawa, K.; Alt, F.W.; Brugge, J.S.; Swat, W. Vav family proteins couple to diverse cell surface receptors. Mol. Cell Biol. 2000, 20, 6364–6373. [Google Scholar] [CrossRef] [PubMed]
- Aghazadeh, Y.; Papadopoulos, V. The role of the 14-3-3 protein family in health, disease, and drug development. Drug Discov. Today 2016, 21, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.D.; Zhang, X.; Zhang, J.N.; Yang, Z.J.; Zhen, H.N.; Cheng, G.; Li, B.; Gao, D. Immunocytochemical detection of 14-3-3 in primary nervous system tumors. J. Neurooncol. 2006, 77, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Marino-Enriquez, A.; Ou, W.; Zhu, M.; Ali, R.H.; Chiang, S.; Amant, F.; Gilks, C.B.; van de Rijn, M.; Oliva, E.; et al. The clinicopathologic features of YWHAE-FAM22 endometrial stromal sarcomas: A histologically high-grade and clinically aggressive tumor. Am. J. Surg. Pathol. 2012, 36, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Masters, S.C.; Subramanian, R.R.; Truong, A.; Yang, H.; Fujii, K.; Zhang, H.; Fu, H. Survival-promoting functions of 14-3-3 proteins. Biochem. Soc. Trans. 2002, 30, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K.; Hashizume, S.; Kato, M.; Honjoh, T.; Setoguchi, Y.; Yasumoto, K. Elevated expression levels of the 14-3-3 family of proteins in lung cancer tissues. Hum. Antib. 1997, 8, 189–194. [Google Scholar] [CrossRef]
- Radhakrishnan, V.M.; Martinez, J.D. 14-3-3gamma induces oncogenic transformation by stimulating MAP kinase and PI3K signaling. PLoS ONE 2010, 5, e11433. [Google Scholar] [CrossRef]
- Wang, Z.; Nesland, J.M.; Suo, Z.; Trope, C.G.; Holm, R. The prognostic value of 14-3-3 isoforms in vulvar squamous cell carcinoma cases: 14-3-3beta and epsilon are independent prognostic factors for these tumors. PLoS ONE 2011, 6, e24843. [Google Scholar] [CrossRef]
- Li, S.; Wang, Q.; Wang, Y.; Chen, X.; Wang, Z. PLC-gamma1 and Rac1 coregulate EGF-induced cytoskeleton remodeling and cell migration. Mol. Endocrinol. 2009, 23, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Mancini, M.; Corradi, V.; Petta, S.; Barbieri, E.; Manetti, F.; Botta, M.; Santucci, M.A. A new nonpeptidic inhibitor of 14-3-3 induces apoptotic cell death in chronic myeloid leukemia sensitive or resistant to imatinib. J. Pharmacol. Exp. Ther. 2011, 336, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Abdrabou, A.B.D.; Liu, A.; Wang, Z. Differential subcellular distribution and translocation of seven 14-3-3 isoforms in response to EGF and during cell cycle. Cancers 2019. under revision. [Google Scholar]
- Seimiya, H.; Sawada, H.; Muramatsu, Y.; Shimizu, M.; Ohko, K.; Yamane, K.; Tsuruo, T. Involvement of 14-3-3 proteins in nuclear localization of telomerase. EMBO J. 2000, 19, 2652–2661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, J.; Kwon, T.; Lee, E.J.; Kim, C.H.; Han, Y.S.; Hong, S.K.; Hyun, S.; Kang, S.S. 14-3-3 protein mediates phosphorylation of microtubule-associated protein tau by serum- and glucocorticoid-induced protein kinase 1. Mol. Cells 2004, 18, 360–368. [Google Scholar] [PubMed]
- Sumioka, A.; Nagaishi, S.; Yoshida, T.; Lin, A.; Miura, M.; Suzuki, T. Role of 14-3-3gamma in FE65-dependent gene transactivation mediated by the amyloid beta-protein precursor cytoplasmic fragment. J. Biol. Chem. 2005, 280, 42364–42374. [Google Scholar] [CrossRef] [PubMed]
- Satoh, J.; Onoue, H.; Arima, K.; Yamamura, T. The 14-3-3 protein forms a molecular complex with heat shock protein Hsp60 and cellular prion protein. J. Neuropathol. Exp. Neurol. 2005, 64, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Cummings, N.; Batchelor, M.; Hill, P.J.; Dubois, T.; Mellits, K.H.; Frankel, G.; Connerton, I. Host protein interactions with enteropathogenic Escherichia coli (EPEC): 14-3-3tau binds Tir and has a role in EPEC-induced actin polymerization. Cell. Microbiol. 2006, 8, 55–71. [Google Scholar] [CrossRef]
- Ottmann, C.; Yasmin, L.; Weyand, M.; Veesenmeyer, J.L.; Diaz, M.H.; Palmer, R.H.; Francis, M.S.; Hauser, A.R.; Wittinghofer, A.; Hallberg, B. Phosphorylation-independent interaction between 14-3-3 and exoenzyme S: From structure to pathogenesis. EMBO J. 2007, 26, 902–913. [Google Scholar] [CrossRef]
- Ito, T.; Nakata, M.; Fukazawa, J.; Ishida, S.; Takahashi, Y. Phosphorylation-independent binding of 14-3-3 to NtCDPK1 by a new mode. Plant Signal. Behav. 2014, 9, e977721. [Google Scholar] [CrossRef]
- West-Foyle, H.; Kothari, P.; Osborne, J.; Robinson, D.N. 14-3-3 proteins tune non-muscle myosin II assembly. J. Biol. Chem. 2018, 293, 6751–6761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, S.; Zhou, R.; Zhu, Y. Structure of the 14-3-3zeta-LKB1 fusion protein provides insight into a novel ligand-binding mode of 14-3-3. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2015, 71, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Ding, S.; Zhou, R.; Wu, J. Structure of the complex of phosphorylated liver kinase B1 and 14-3-3zeta. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2017, 73, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Gerber, K.J.; Squires, K.E.; Hepler, J.R. 14-3-3gamma binds regulator of G protein signaling 14 (RGS14) at distinct sites to inhibit the RGS14: Galphai-AlF4(-) signaling complex and RGS14 nuclear localization. J. Biol. Chem. 2018, 293, 14616–14631. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Li, L.; Ballermann, B.; Wang, Z. Phosphorylation and activation of RhoA by ERK in response to epidermal growth factor stimulation. PLoS ONE 2016, 11, e0147103. [Google Scholar] [CrossRef] [PubMed]
- Payapilly, A.; Malliri, A. Compartmentalisation of Rac1 signalling. Curr. Opin. Cell Biol. 2018, 54, 50–56. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdrabou, A.; Brandwein, D.; Liu, C.; Wang, Z. Rac1 S71 Mediates the Interaction between Rac1 and 14-3-3 Proteins. Cells 2019, 8, 1006. https://doi.org/10.3390/cells8091006
Abdrabou A, Brandwein D, Liu C, Wang Z. Rac1 S71 Mediates the Interaction between Rac1 and 14-3-3 Proteins. Cells. 2019; 8(9):1006. https://doi.org/10.3390/cells8091006
Chicago/Turabian StyleAbdrabou, Abdalla, Daniel Brandwein, Changyu Liu, and Zhixiang Wang. 2019. "Rac1 S71 Mediates the Interaction between Rac1 and 14-3-3 Proteins" Cells 8, no. 9: 1006. https://doi.org/10.3390/cells8091006