
Cholangiocarcinoma Malignant Traits Are Promoted by Schwann Cells through TGFβ Signaling in a Model of Perineural Invasion

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Histomorphology and Immunohistochemistry on Human CCA Samples
2.3. Histopathological Assessment of Human CCA Samples
2.4. Cell Cultures and Conditioned Media Preparation
2.5. Nerve Explants and 3D Perineural Invasion (PNI) Assay
2.6. Migration and Invasion Assays
2.7. Cell Viability Assay
2.8. Clonogenic Assay
2.9. Proliferation Assay (BrdU Incorporation)
2.10. Western Blotting and Differential Proteome
2.10.1. Protein Extraction
2.10.2. Western Blotting
2.10.3. Differential Proteome
2.11. Immunocytochemistry
2.12. Statistical Analyses
3. Results
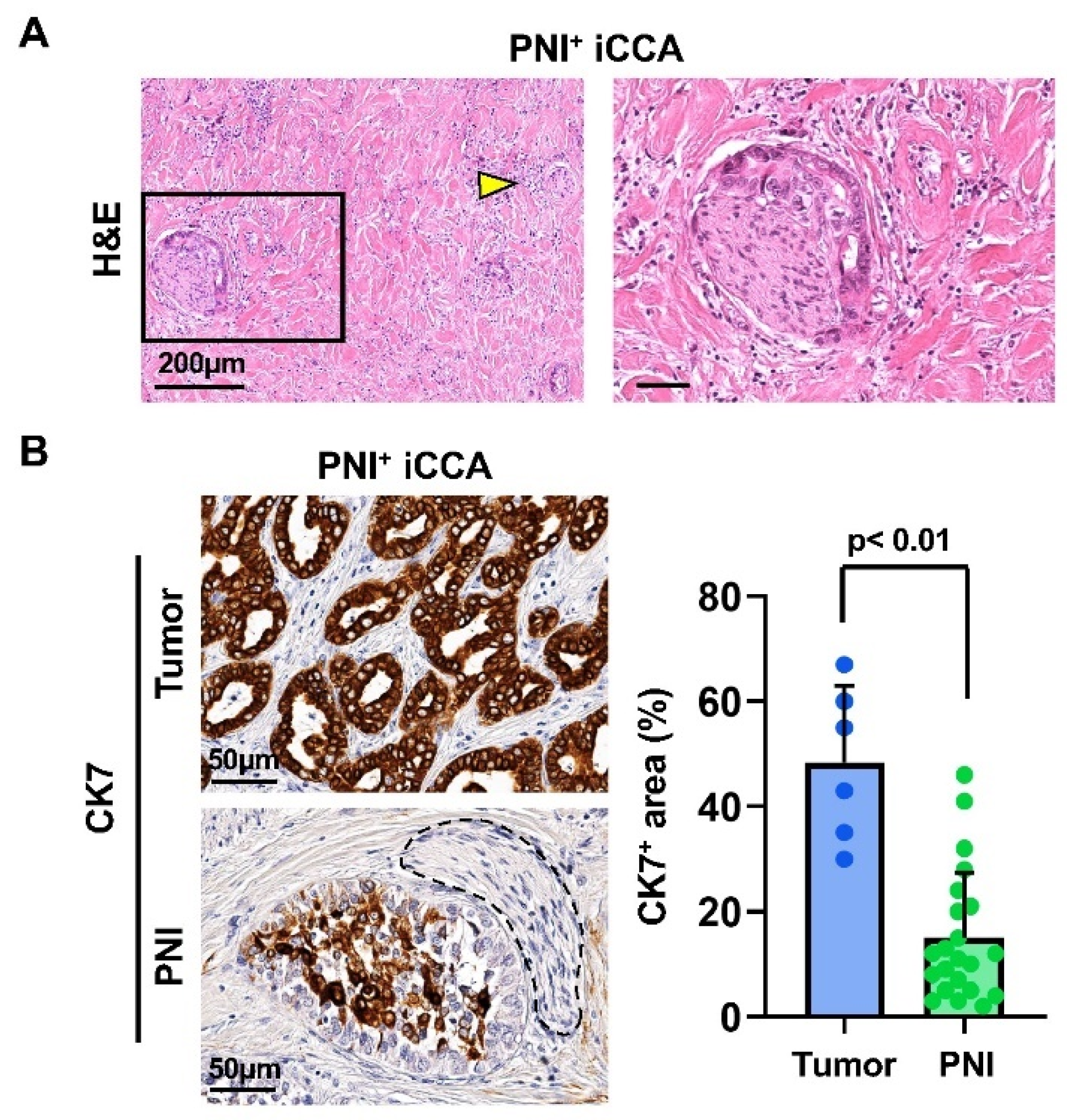
3.1. Nerve-Infiltrating CCA Cells Show Lower Cytokeratin 7 Positivity Compared to the Tumor Mass in Human Intrahepatic CCA Samples
3.1.1. Patient Characteristics and Sample Selection
3.1.2. In Situ Phenotype of Tumor Cells Invading Nerves
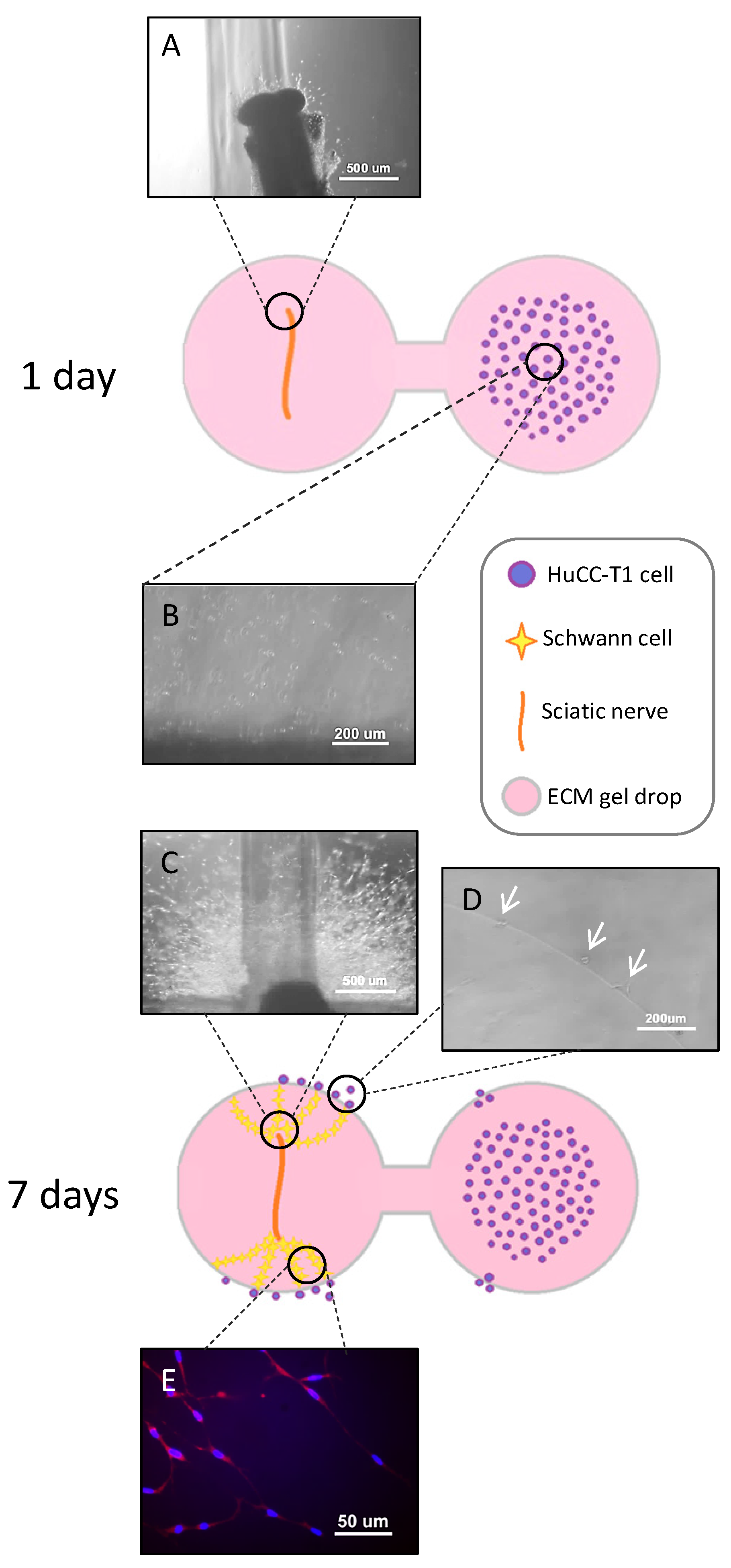
3.2. HuCC-T1 Cells Migrate towards Sciatic Nerve Explants through an Extracellular Matrix Scaffold in a 3D Model of Perineural Invasion
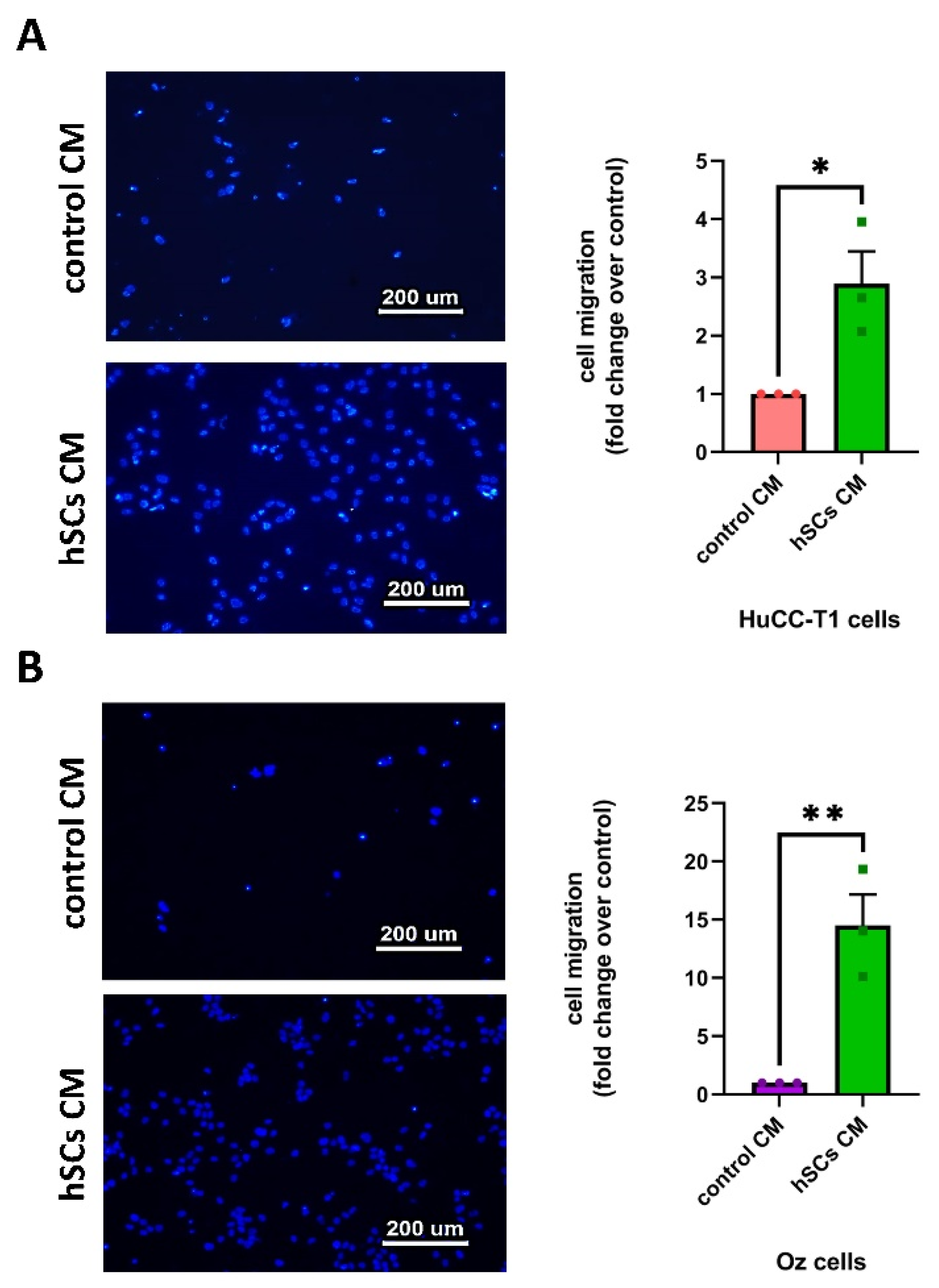
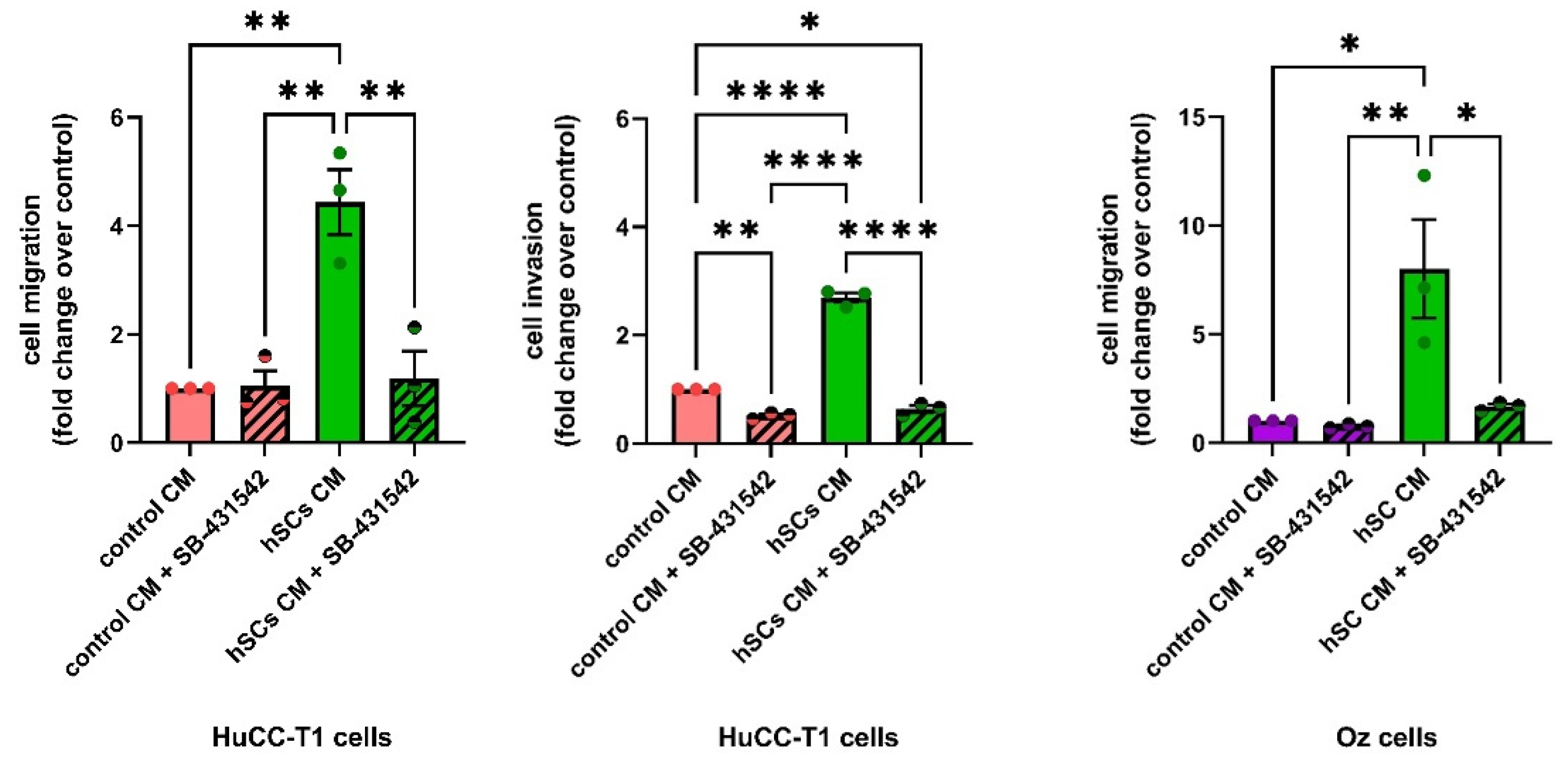
3.3. Human SC-Conditioned Medium Increases the Migration of iCCA-Derived Cells
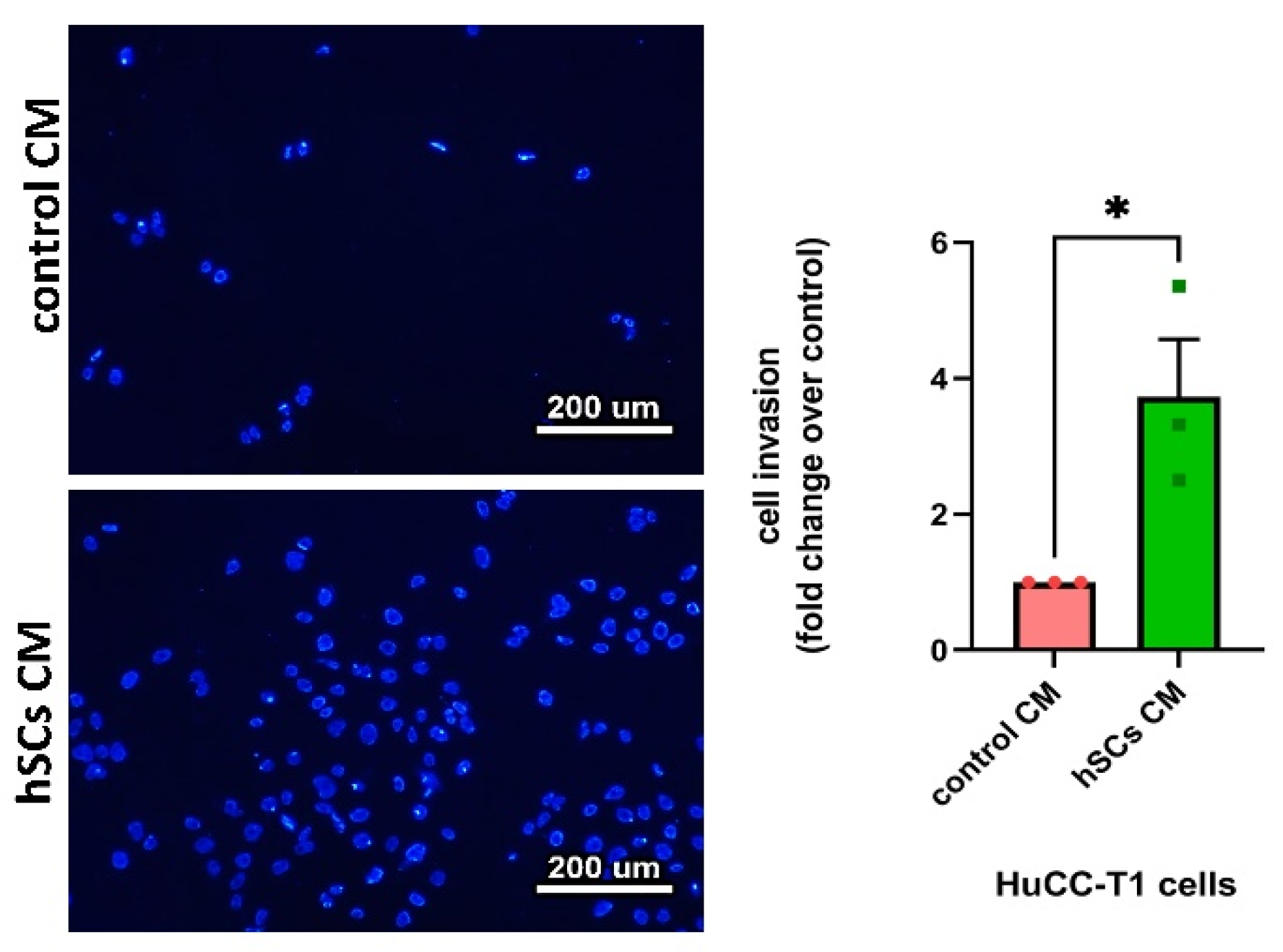
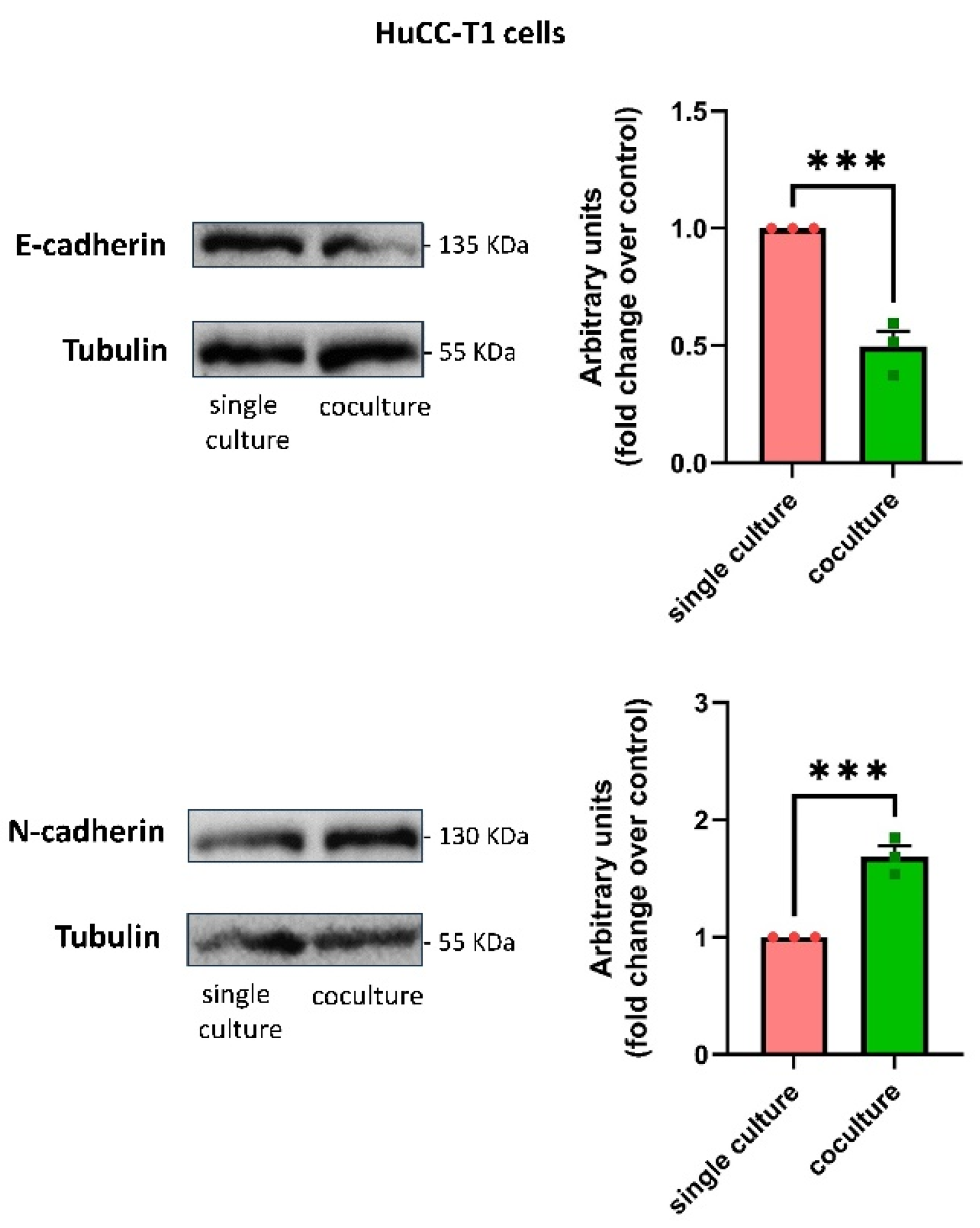
3.4. Human SC-Conditioned Medium Increases Invasion and Induces Cadherin Switch in HuCC-T1 Cells
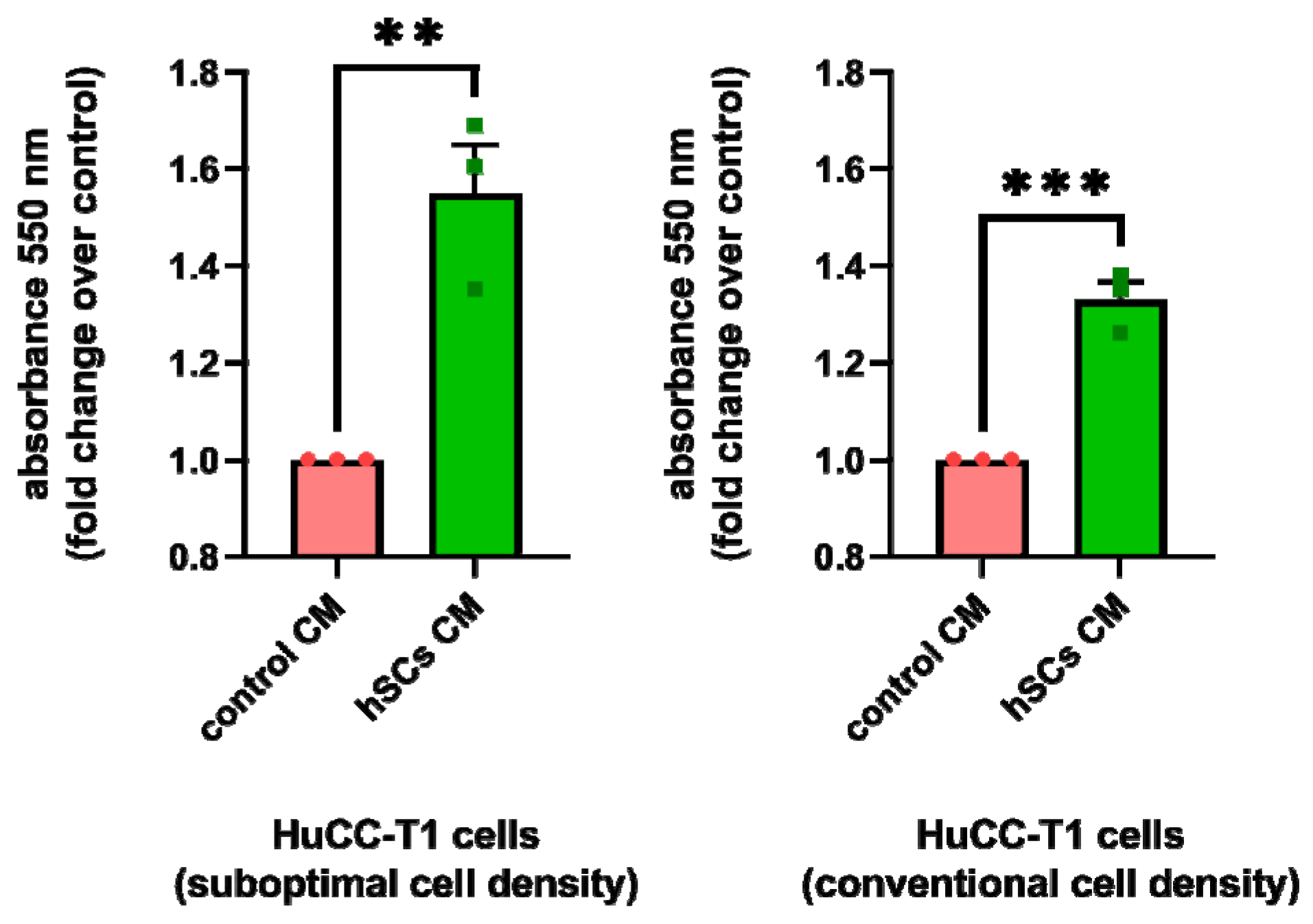
3.5. Human SC-Conditioned Medium Stimulates Cell Viability and Proliferation of iCCA Cells
3.5.1. Human SC-Conditioned Medium Increases HuCC-T1 Cell Viability
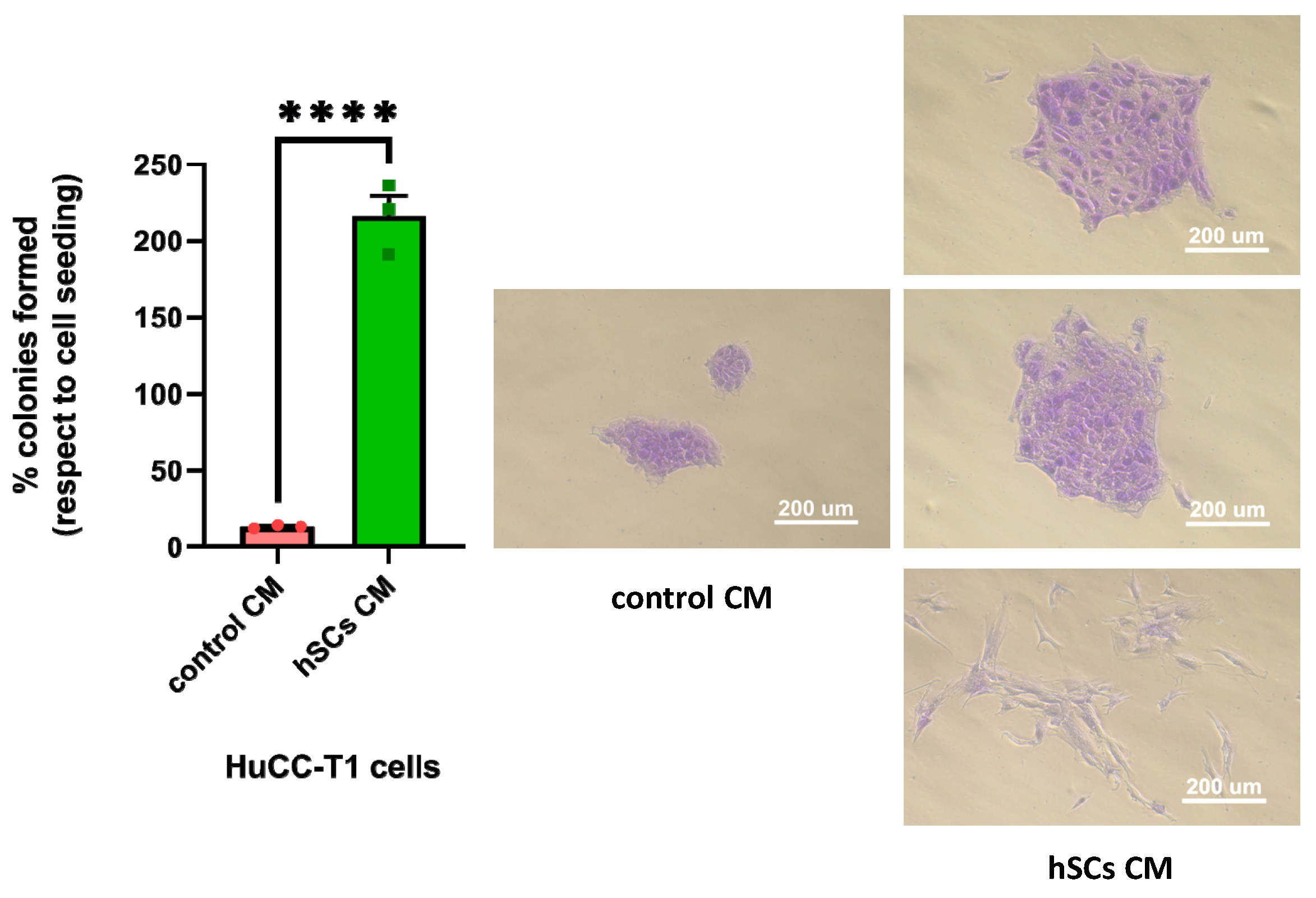
3.5.2. Human SC CM Increased Clonogenicity of HuCC-T1 Cells
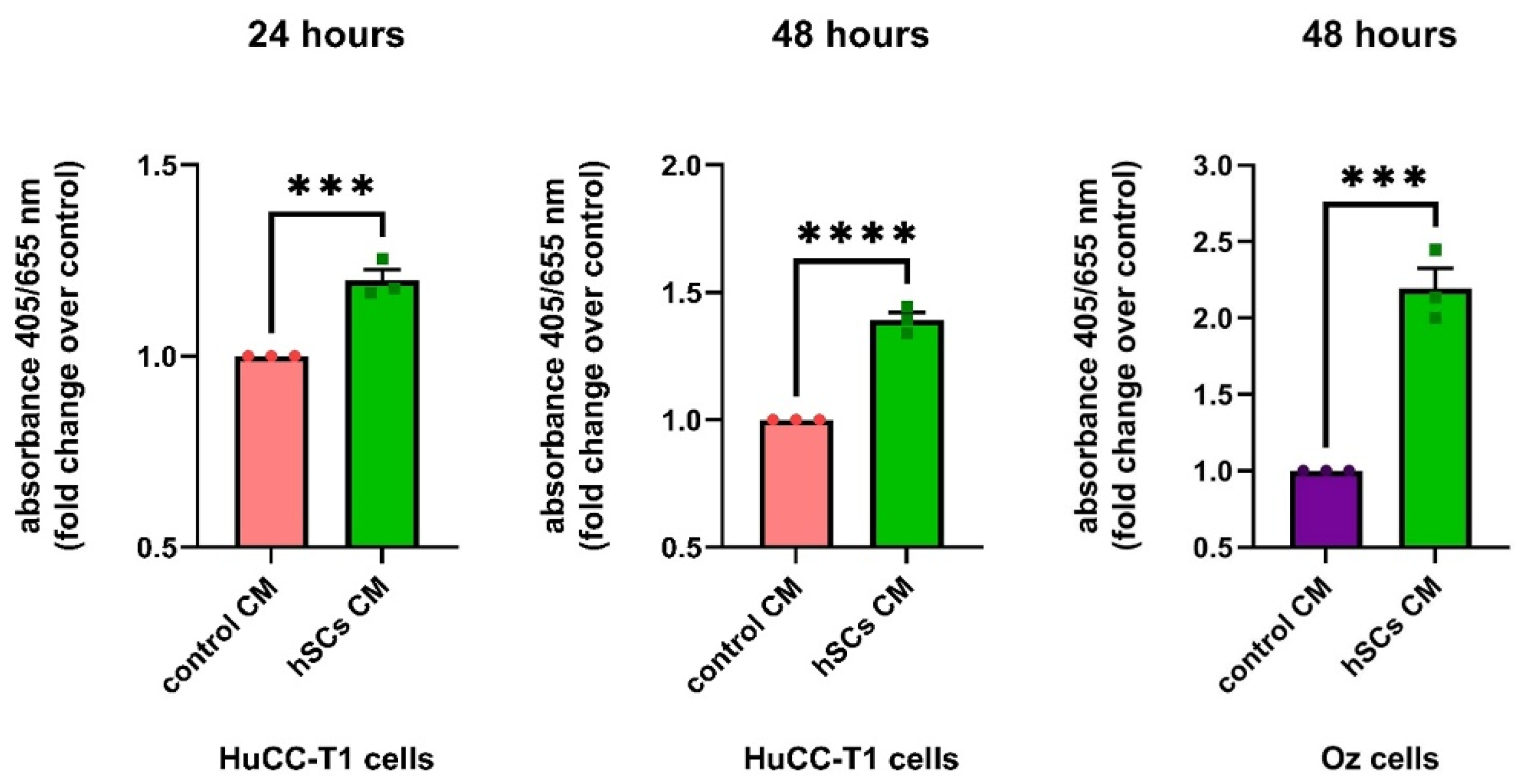
3.5.3. Human SC CM Increases HuCC-T1 Proliferation
3.6. Differential Proteome Analysis of HuCC-T1 Cells Cocultured with Human SCs
3.7. The Effect of hSC CM on the Migration and Invasion of iCCA Cells Is Blocked by an Antagonist of TGFBR1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Massa, A.; Varamo, C.; Vita, F.; Tavolari, S.; Peraldo-Neia, C.; Brandi, G.; Rizzo, A.; Cavalloni, G.; Aglietta, M. Evolution of the Experimental Models of Cholangiocarcinoma. Cancers 2020, 12, 2308. [Google Scholar] [CrossRef] [PubMed]
- Montal, R.; Sia, D.; Montironi, C.; Leow, W.Q.; Esteban-Fabró, R.; Pinyol, R.; Torres-Martin, M.; Bassaganyas, L.; Moeini, A.; Peix, J.; et al. Molecular classification and therapeutic targets in extrahepatic cholangiocarcinoma. J. Hepatol. 2020, 73, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Sivakumar, S.; Bednarsch, J.; Wiltberger, G.; Kather, J.N.; Niehues, J.; de Vos-Geelen, J.; Valkenburg-van Iersel, L.; Kintsler, S.; Roeth, A.; et al. Nerve fibers in the tumor microenvironment in neurotropic cancer-pancreatic cancer and cholangiocarcinoma. Oncogene 2021, 40, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Winkler, F.; Venkatesh, H.S.; Amit, M.; Batchelor, T.; Demir, I.E.; Deneen, B.; Gutmann, D.H.; Hervey-Jumper, S.; Kuner, T.; Mabbott, D.; et al. Cancer neuroscience: State of the field, emerging directions. Cell 2023, 186, 1689–1707. [Google Scholar] [CrossRef] [PubMed]
- Stierli, S.; Imperatore, V.; Lloyd, A.C. Schwann cell plasticity-roles in tissue homeostasis, regeneration, and disease. Glia 2019, 67, 2203–2215. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Chen, S.; Chen, M. Schwann Cells in the Tumor Microenvironment: Need More Attention. J. Oncol. 2022, 2022, 1058667. [Google Scholar] [CrossRef]
- Demir, I.E.; Boldis, A.; Pfitzinger, P.L.; Teller, S.; Brunner, E.; Klose, N.; Kehl, T.; Maak, M.; Lesina, M.; Laschinger, M.; et al. Investigation of Schwann cells at neoplastic cell sites before the onset of cancer invasion. J. Natl. Cancer Inst. 2014, 106, dju184. [Google Scholar] [CrossRef]
- Shurin, G.V.; Kruglov, O.; Ding, F.; Lin, Y.; Hao, X.; Keskinov, A.A.; You, Z.; Lokshin, A.E.; LaFramboise, W.A.; Falo, L.D., Jr.; et al. Melanoma-Induced Reprogramming of Schwann Cell Signaling Aids Tumor Growth. Cancer Res. 2019, 79, 2736–2747. [Google Scholar] [CrossRef]
- Zhou, Y.; Shurin, G.V.; Zhong, H.; Bunimovich, Y.L.; Han, B.; Shurin, M.R. Schwann Cells Augment Cell Spreading and Metastasis of Lung Cancer. Cancer Res. 2018, 78, 5927–5939. [Google Scholar] [CrossRef]
- Zaccari, P.; Cardinale, V.; Severi, C.; Pedica, F.; Carpino, G.; Gaudio, E.; Doglioni, C.; Petrone, M.C.; Alvaro, D.; Arcidiacono, P.G.; et al. Common features between neoplastic and preneoplastic lesions of the biliary tract and the pancreas. World J. Gastroenterol. 2019, 25, 4343–4359. [Google Scholar] [CrossRef] [PubMed]
- Deborde, S.; Omelchenko, T.; Lyubchik, A.; Zhou, Y.; He, S.; McNamara, W.F.; Chernichenko, N.; Lee, S.Y.; Barajas, F.; Chen, C.H.; et al. Schwann cells induce cancer cell dispersion and invasion. J. Clin. Investig. 2016, 126, 1538–1554. [Google Scholar] [CrossRef]
- Deborde, S.; Gusain, L.; Powers, A.; Marcadis, A.; Yu, Y.; Chen, C.H.; Frants, A.; Kao, E.; Tang, L.H.; Vakiani, E.; et al. Reprogrammed Schwann Cells Organize into Dynamic Tracks that Promote Pancreatic Cancer Invasion. Cancer Discov. 2022, 12, 2454–2473. [Google Scholar] [CrossRef]
- Su, D.; Guo, X.; Huang, L.; Ye, H.; Li, Z.; Lin, L.; Chen, R.; Zhou, Q. Tumor-neuroglia interaction promotes pancreatic cancer metastasis. Theranostics 2020, 10, 5029–5047. [Google Scholar] [CrossRef]
- Gundlach, J.P.; Kerber, J.; Hendricks, A.; Bernsmeier, A.; Halske, C.; Röder, C.; Becker, T.; Röcken, C.; Braun, F.; Sebens, S.; et al. Paracrine Interaction of Cholangiocellular Carcinoma with Cancer-Associated Fibroblasts and Schwann Cells Impact Cell Migration. J. Clin. Med. 2022, 11, 2785. [Google Scholar] [CrossRef]
- Yamazaki, S.; Ema, H.; Karlsson, G.; Yamaguchi, T.; Miyoshi, H.; Shioda, S.; Taketo, M.M.; Karlsson, S.; Iwama, A.; Nakauchi, H. Nonmyelinating Schwann cells maintain hematopoietic stem cell hibernation in the bone marrow niche. Cell 2011, 147, 1146–1158. [Google Scholar] [CrossRef]
- Yamazaki, S.; Nakauchi, H. Bone marrow Schwann cells induce hematopoietic stem cell hibernation. Int. J. Hematol. 2014, 99, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Brockes, J.P. Nerve dependence in tissue, organ, and appendage regeneration. Trends Neurosci. 2012, 35, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Roger, E.; Martel, S.; Bertrand-Chapel, A.; Depollier, A.; Chuvin, N.; Pommier, R.M.; Yacoub, K.; Caligaris, C.; Cardot-Ruffino, V.; Chauvet, V.; et al. Correction: Schwann cells support oncogenic potential of pancreatic cancer cells through TGFβ signaling. Cell Death Dis. 2020, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Baker, D.; Ten Dijke, P. TGF-β-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis. Int. J. Mol. Sci. 2019, 20, 2767. [Google Scholar] [CrossRef]
- Petrungaro, S.; de Franchis, V.; Filippini, A.; Facchiano, A.; Gaudio, E.; Giampietri, C. Autophagy impairment in human bile duct carcinoma cells. Front. Physiol. 2023, 14, 1249264. [Google Scholar] [CrossRef] [PubMed]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Yi, S.Q.; Dai, Y.; Kurosawa, K.; Miwa, Y.; Sato, I. Clinical anatomy of the anterior and posterior hepatic plexuses, including relations with the pancreatic plexus: A cadaver study. Clin. Anat. 2020, 33, 630–636. [Google Scholar] [CrossRef]
- Mavros, M.N.; Economopoulos, K.P.; Alexiou, V.G.; Pawlik, T.M. Treatment and Prognosis for Patients With Intrahepatic Cholangiocarcinoma: Systematic Review and Meta-analysis. JAMA Surg. 2014, 149, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Abuetabh, Y.; Persad, S.; Nagamori, S.; Huggins, J.; Al-Bahrani, R.; Sergi, C. Expression of E-cadherin and β-catenin in two cholangiocarcinoma cell lines (OZ and HuCCT1) with different degree of invasiveness of the primary tumor. Ann. Clin. Lab. Sci. 2011, 41, 217–223. [Google Scholar]
- Ryynänen, J.; Sollberg, S.; Olsen, D.R.; Uitto, J. Transforming growth factor-beta up-regulates type VII collagen gene expression in normal and transformed epidermal keratinocytes in culture. Biochem. Biophys. Res. Commun. 1991, 180, 673–680. [Google Scholar] [CrossRef]
- Calonge, M.J.; Seoane, J.; Massagué, J. Opposite Smad and chicken ovalbumin upstream promoter transcription factor inputs in the regulation of the collagen VII gene promoter by transforming growth factor-beta. J. Biol. Chem. 2004, 279, 23759–23765. [Google Scholar] [CrossRef]
- Buchholz, M.; Biebl, A.; Neesse, A.; Wagner, M.; Iwamura, T.; Leder, G.; Adler, G.; Gress, T.M. SERPINE2 (protease nexin I) promotes extracellular matrix production and local invasion of pancreatic tumors in vivo. Cancer Res. 2003, 63, 4945–4951. [Google Scholar]
- Seshacharyulu, P.; Pandey, P.; Datta, K.; Batra, S.K. Phosphatase: PP2A structural importance, regulation and its aberrant expression in cancer. Cancer Lett. 2013, 335, 9–18. [Google Scholar] [CrossRef]
- Xu, L.; Chen, X.; Jiang, H.; Xu, J.; Wang, L.; Sun, Y. NDUFC1 Is Upregulated in Gastric Cancer and Regulates Cell Proliferation, Apoptosis, Cycle and Migration. Front. Oncol. 2021, 11, 709044. [Google Scholar] [CrossRef]
- Lu, Z.; Wang, J.; Zheng, T.; Liang, Y.; Yin, D.; Song, R.; Pei, T.; Pan, S.; Jiang, H.; Liu, L. FTY720 inhibits proliferation and epithelial-mesenchymal transition in cholangiocarcinoma by inactivating STAT3 signaling. BMC Cancer 2014, 14, 783. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Guo, D.; Zhang, X.; Zhu, Y.; Wang, Z.; Jin, Y.; Guo, W.; Zhang, S. ENO3 Inhibits Growth and Metastasis of Hepatocellular Carcinoma via Wnt/β-Catenin Signaling Pathway. Front. Cell Dev. Biol. 2021, 9, 797102. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Kong, Y.; Xu, M.; Zhang, H. Notch3 functions as a tumor suppressor by controlling cellular senescence. Cancer Res. 2013, 73, 3451–3459. [Google Scholar] [CrossRef]
- Araki, K.; Shimura, T.; Suzuki, H.; Tsutsumi, S.; Wada, W.; Yajima, T.; Kobayahi, T.; Kubo, N.; Kuwano, H. E/N-cadherin switch mediates cancer progression via TGF-β-induced epithelial-to-mesenchymal transition in extrahepatic cholangiocarcinoma. Br. J. Cancer 2011, 105, 1885–1893. [Google Scholar] [CrossRef]
- Cristóbal, I.; Lamarca, A. Role of the PP2A Pathway in Cholangiocarcinoma: State of the Art and Future Perspectives. Cancers 2022, 14, 5422. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Gudey, S.K.; Landström, M. Non-Smad signaling pathways. Cell Tissue Res. 2012, 347, 11–20. [Google Scholar] [CrossRef]
- Yasuoka, C.; Ihara, Y.; Ikeda, S.; Miyahara, Y.; Kondo, T.; Kohno, S. Antiapoptotic activity of Akt is down-regulated by Ca2+ in myocardiac H9c2 cells. Evidence of Ca2+-dependent regulation of protein phosphatase 2Ac. J. Biol. Chem. 2004, 279, 51182–51192. [Google Scholar] [CrossRef]
- Lonning, S.; Mannick, J.; McPherson, J.M. Antibody targeting of TGF-β in cancer patients. Curr. Pharm. Biotechnol. 2011, 12, 2176–2189. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| iCCA with PNI | |
|---|---|
| Age (years) | 70 [64–73] |
| Gender | F: 3 |
| M: 3 | |
| Growth pattern | MF: 5 |
| PI: 1 | |
| IG: 0 | |
| TNM | |
| T | T1: 0 |
| T2: 3 | |
| T3: 3 | |
| T4: 0 | |
| N | N0: 5 |
| N1: 1 | |
| M | M0: 6 |
| M1: 0 | |
| Stage | I: 0 |
| II: 3 | |
| III: 3 | |
| IV: 0 |
| Protein Name | Involvement in Migration, Invasion, EMT (TGFβ Pathway) | Involvement in Proliferation, Survival | Regulation |
|---|---|---|---|
| Collagen alpha-1 (VII) chain [26,27] | X | ↑ | |
| Protease Nexin 1 (PN1) [28,29] | X | ↑ | |
| NADH dehydrogenase [ubiquinone] 1 alpha subcomplex subunit 1 (NDUFC1) [30] | X | X | ↑ |
| Cytoplasmic dynein 1 intermediate chain 1 | X | ↑ | |
| Protein 4.1 | X | ↑ | |
| Serine/threonine-protein phosphatase 2A catalytic subunit alpha isoform (PP2Ac) [29,31] | X | X | ↓ |
| Beta-Enolase 3 [32] | X | X | ↓ |
| Neurogenic locus notch homolog protein 3 (NOTCH3) [33] | X | X | ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Franchis, V.; Petrungaro, S.; Pizzichini, E.; Camerini, S.; Casella, M.; Somma, F.; Mandolini, E.; Carpino, G.; Overi, D.; Cardinale, V.; et al. Cholangiocarcinoma Malignant Traits Are Promoted by Schwann Cells through TGFβ Signaling in a Model of Perineural Invasion. Cells 2024, 13, 366. https://doi.org/10.3390/cells13050366
de Franchis V, Petrungaro S, Pizzichini E, Camerini S, Casella M, Somma F, Mandolini E, Carpino G, Overi D, Cardinale V, et al. Cholangiocarcinoma Malignant Traits Are Promoted by Schwann Cells through TGFβ Signaling in a Model of Perineural Invasion. Cells. 2024; 13(5):366. https://doi.org/10.3390/cells13050366
Chicago/Turabian Stylede Franchis, Valerio, Simonetta Petrungaro, Elisa Pizzichini, Serena Camerini, Marialuisa Casella, Francesca Somma, Enrico Mandolini, Guido Carpino, Diletta Overi, Vincenzo Cardinale, and et al. 2024. "Cholangiocarcinoma Malignant Traits Are Promoted by Schwann Cells through TGFβ Signaling in a Model of Perineural Invasion" Cells 13, no. 5: 366. https://doi.org/10.3390/cells13050366