
The Wnt Co-Receptor PTK7/Otk and Its Homolog Otk-2 in Neurogenesis and Patterning

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Crosses and Expression of UAS Construct
2.2. Fly Lines and Crosses
2.2.1. Fly Lines Used
- mataTub-Gal4VP16 15 (D. St. Johnston)
- daughterless-Gal4 BDSC 55850
- -> Recombined to make mat, da Gal4 (this study)
- UAS-Otk-1 pUASg 3XHA [11]
- UAS-Wnt-4 RNAi BDSC Stock #29442 [50]
- UAS–Otk-1 RNAi BDSC Stock #25790
- UAS–Otk-2 RNAi BDSC Stock #38973
- UAS–Otk-1 RNAi BDSC Stock #55869
- UAS–Otk-2 RNAi BDSC Stock #55892
- UAS–Otk-2 RNAi BDSC Stock #57040
- UAS-Frankenbody HA nanobody-GFP; daughterless-GAL4 [51]
- Mi[Trojan-GAL4]otk BDSC Stock #76759
- UAS-wg BDSC 5918 [52]
- UAS-Td-Tomato BDSC Stock #36328 (Joost Schulte and Katharine Sepp)
- elavGal4, UAS-GFP BDSC Stock #5146
2.2.2. Fly Crosses Performed
- mat, da GAL4 × UAS-Otk-1
- mat, da GAL4 × UAS-Wg
- mat, da GAL4 × UAS-Otk-1, UAS-Wnt-4
- mat, da GAL4 × UAS-Otk-1 RNAi
- mat, da GAL4 × UAS-Otk-2 RNAi
- mat, da GAL4 × UAS-Otk-1 RNAi, UAS-Wnt-4 RNAi
- mat, da GAL4 × UAS-Otk-2 RNAi, UAS-Wnt-4 RNAi
- mat, da GAL4 × UAS-Wnt-4
- mat, da GAL4 × UAS-Wnt-4 RNAi
- mat, da GAL4 × UAS-Otk-1 RNAi, UAS-Otk-2 RNAi
- da GAL4 × UAS-Otk1, UAS-Franken HA-GFP
- Mi[Trojan-GAL4]otk × UAS-Td-Tomato
- elavGal4, UAS-GFP × UAS-Otk-1 RNAi, UAS-Otk-2 RNAi
2.3. Imaging
2.3.1. Light Sheet Imaging
2.3.2. Confocal Imaging
2.4. Transcriptomic Analysis
3. Results
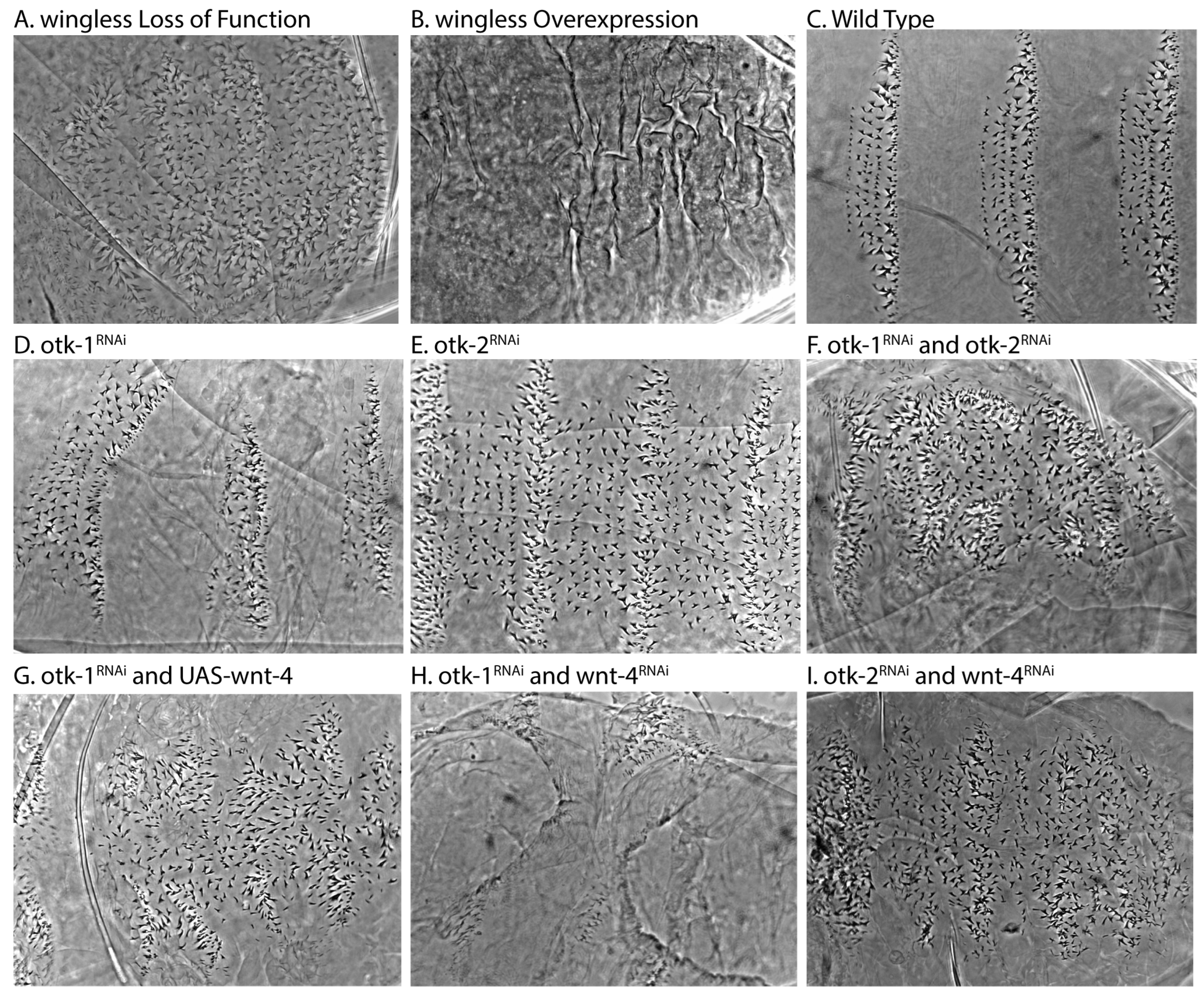
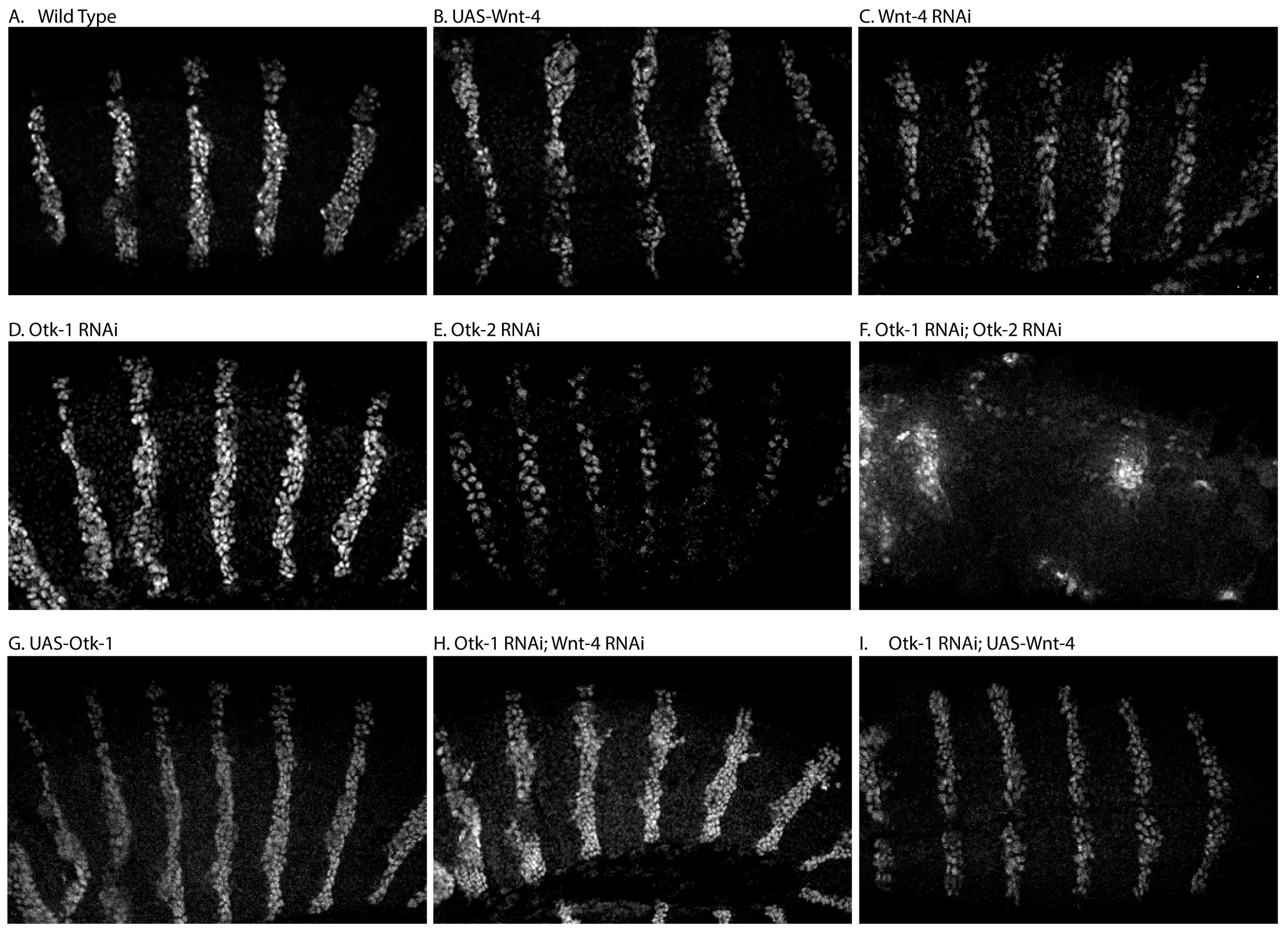
3.1. Patterning
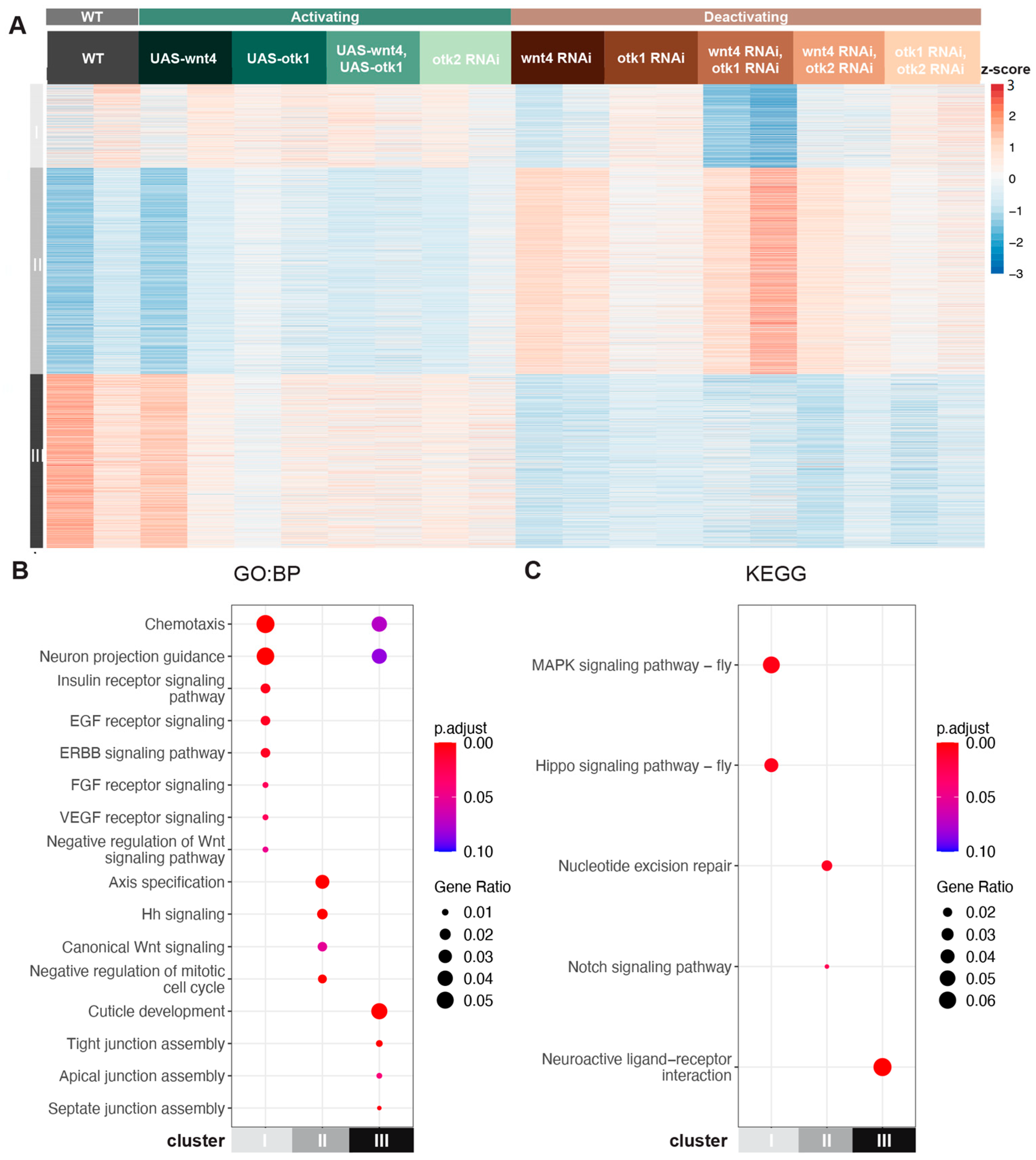
3.2. Transcriptomic Analysis Identifies Wnt-4-Dependent and Wnt-4-Independent Clusters
3.2.1. Wnt-4 Activates Otk-1 to Oppose Canonical Wnt Signaling
3.2.2. Wnt-4 Activates Otk-1 to Affect Tight Junction Assembly
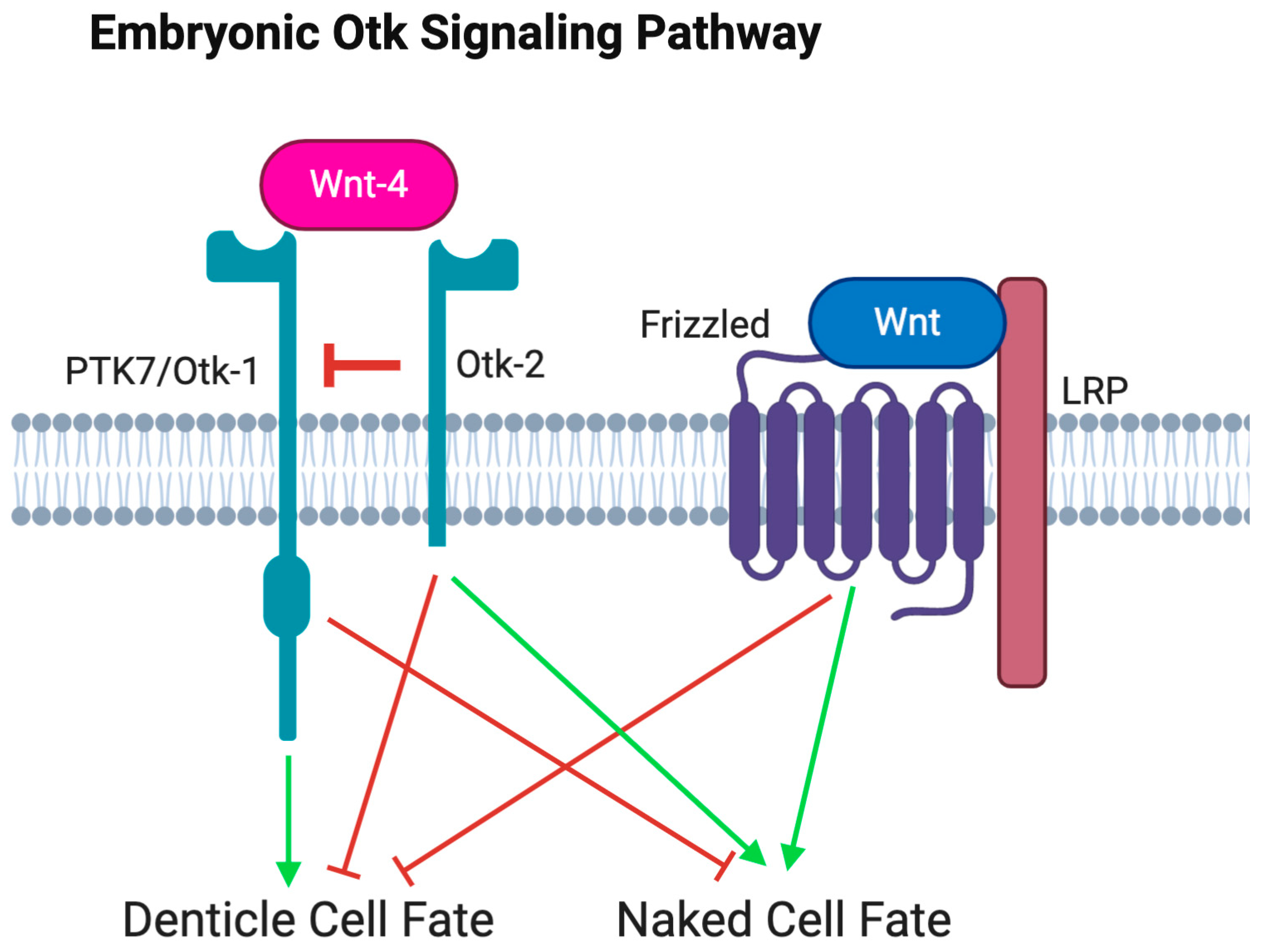
3.2.3. Otk-2 Is an Inhibitor of Otk-1
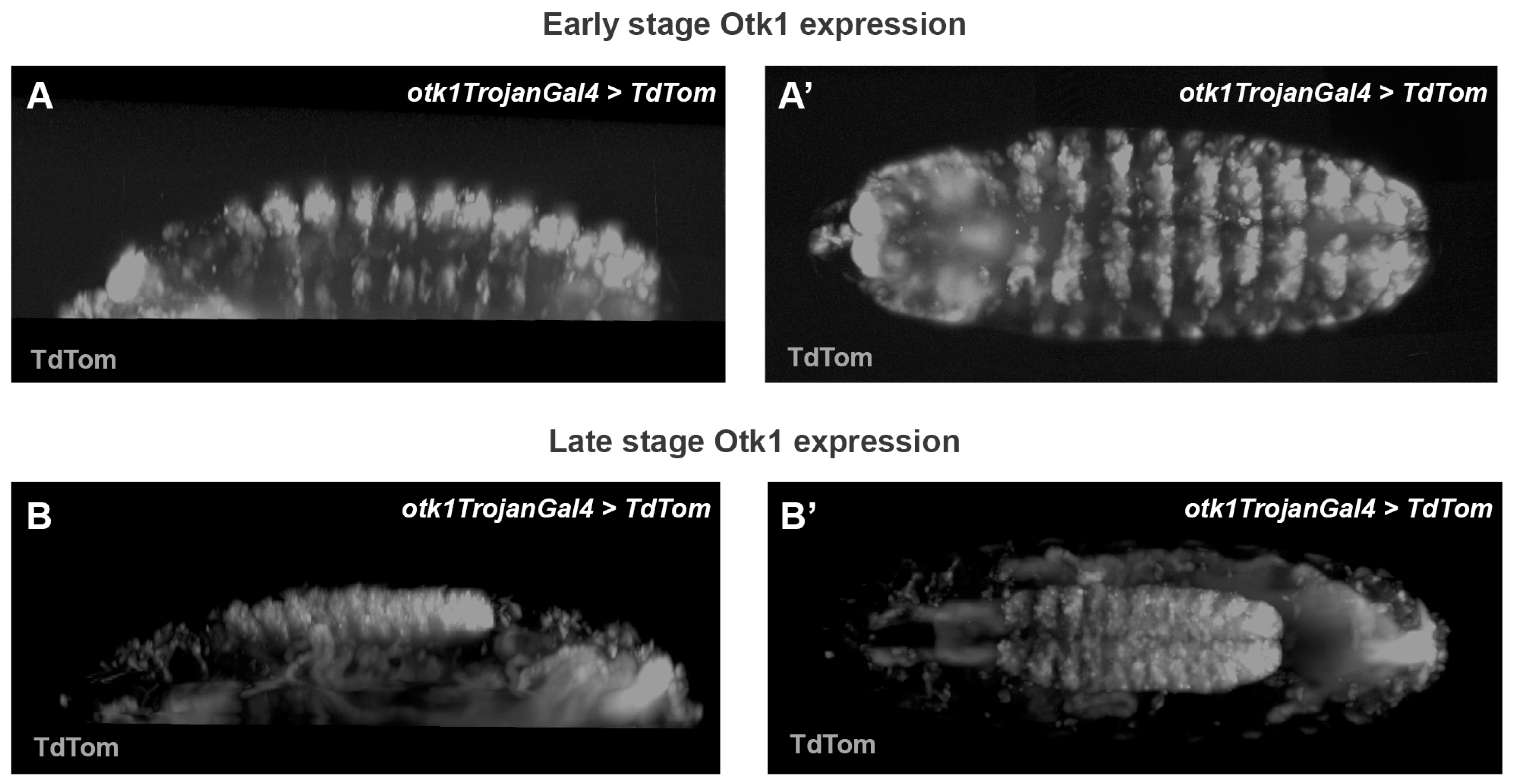
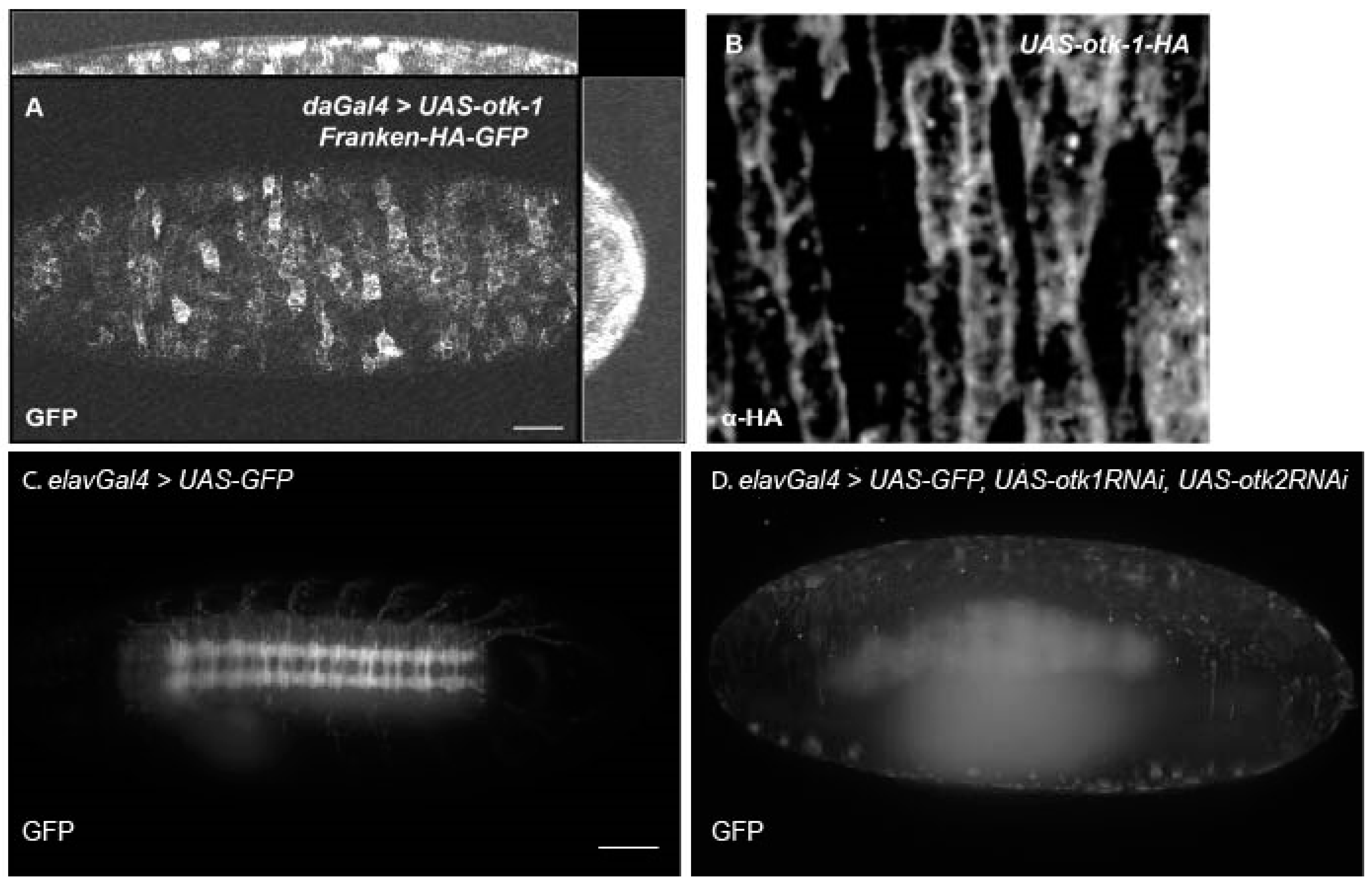
3.3. Tracking Otk Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanson, B. Generating patterns from fields of cells. Examples from drosophila segmentation. EMBO Rep. 2001, 2, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Cadigan, K.M.; Nusse, R. Wnt signaling: A common theme in animal development. Genes. Dev. 1997, 11, 3286–3305. [Google Scholar] [CrossRef] [PubMed]
- van Amerongen, R.; Mikels, A.; Nusse, R. Alternative wnt signaling is initiated by distinct receptors. Sci. Signal. 2008, 1, re9. [Google Scholar] [CrossRef] [PubMed]
- Angers, S.; Moon, R.T. Proximal events in wnt signal transduction. Nat. Rev. Mol. Cell Biol. 2009, 10, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Chen, Y.G. Dishevelled: The hub of wnt signaling. Cell. Signal. 2010, 22, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.F.; Kaur, P.; Bunnag, N.; Suresh, J.; Sung, I.C.H.; Tan, Q.H.; Gruber, J.; Tolwinski, N.S. Wnt signaling in disease. Cells 2019, 8, 826. [Google Scholar] [CrossRef] [PubMed]
- Reya, T.; Clevers, H. Wnt signalling in stem cells and cancer. Nature 2005, 434, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Orte, E.; Saenz-Narciso, B.; Moreno, S.; Cabello, J. Multiple functions of the noncanonical wnt pathway. Trends Genet. 2013, 29, 545–553. [Google Scholar] [CrossRef]
- Schlessinger, K.; Hall, A.; Tolwinski, N. Wnt signaling pathways meet Rho GTPases. Genes Dev. 2009, 23, 265–277. [Google Scholar] [CrossRef]
- Shnitsar, I.; Borchers, A. Ptk7 recruits dsh to regulate neural crest migration. Development 2008, 135, 4015–4024. [Google Scholar] [CrossRef]
- Peradziryi, H.; Kaplan, N.A.; Podleschny, M.; Liu, X.; Wehner, P.; Borchers, A.; Tolwinski, N.S. Ptk7/otk interacts with wnts and inhibits canonical wnt signalling. EMBO J. 2011, 30, 3729–3740. [Google Scholar] [CrossRef]
- Davey, C.F.; Moens, C.B. Planar cell polarity in moving cells: Think globally, act locally. Development 2017, 144, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Borchers, A.G.; Jolicoeur, C.; Rayburn, H.; Baker, J.C.; Tessier-Lavigne, M. Ptk7/cck-4 is a novel regulator of planar cell polarity in vertebrates. Nature 2004, 430, 93–98. [Google Scholar] [CrossRef]
- Yen, W.W.; Williams, M.; Periasamy, A.; Conaway, M.; Burdsal, C.; Keller, R.; Lu, X.; Sutherland, A. Ptk7 is essential for polarized cell motility and convergent extension during mouse gastrulation. Development 2009, 136, 2039–2048. [Google Scholar] [CrossRef]
- De Calisto, J.; Araya, C.; Marchant, L.; Riaz, C.F.; Mayor, R. Essential role of non-canonical wnt signalling in neural crest migration. Development 2005, 132, 2587–2597. [Google Scholar] [CrossRef]
- Linnemannstons, K.; Ripp, C.; Honemann-Capito, M.; Brechtel-Curth, K.; Hedderich, M.; Wodarz, A. The ptk7-related transmembrane proteins off-track and off-track 2 are co-receptors for drosophila wnt2 required for male fertility. PLoS Genet. 2014, 10, e1004443. [Google Scholar] [CrossRef]
- Dunn, N.R.; Tolwinski, N.S. Ptk7 and mcc, unfancied components in non-canonical wnt signaling and cancer. Cancers 2016, 8, 68. [Google Scholar] [CrossRef]
- Berger, H.; Wodarz, A.; Borchers, A. Ptk7 faces the wnt in development and disease. Front. Cell Dev. Biol. 2017, 5, 31. [Google Scholar] [CrossRef]
- Lhoumeau, A.C.; Puppo, F.; Prebet, T.; Kodjabachian, L.; Borg, J.P. Ptk7: A cell polarity receptor with multiple facets. Cell Cycle 2011, 10, 1233–1236. [Google Scholar] [CrossRef]
- Peradziryi, H.; Tolwinski, N.S.; Borchers, A. The many roles of ptk7: A versatile regulator of cell-cell communication. Arch. Biochem. Biophys. 2012, 524, 71–76. [Google Scholar] [CrossRef]
- Kroiher, M.; Miller, M.A.; Steele, R.E. Deceiving appearances: Signaling by “dead” and “fractured” receptor protein-tyrosine kinases. Bioessays 2001, 23, 69–76. [Google Scholar] [CrossRef]
- Na, H.W.; Shin, W.S.; Ludwig, A.; Lee, S.T. The cytosolic domain of protein-tyrosine kinase 7 (ptk7), generated from sequential cleavage by a disintegrin and metalloprotease 17 (adam17) and gamma-secretase, enhances cell proliferation and migration in colon cancer cells. J. Biol. Chem. 2012, 287, 25001–25009. [Google Scholar] [CrossRef]
- Golubkov, V.S.; Chekanov, A.V.; Cieplak, P.; Aleshin, A.E.; Chernov, A.V.; Zhu, W.; Radichev, I.A.; Zhang, D.; Dong, P.D.; Strongin, A.Y. The wnt/planar cell polarity protein-tyrosine kinase-7 (ptk7) is a highly efficient proteolytic target of membrane type-1 matrix metalloproteinase: Implications in cancer and embryogenesis. J. Biol. Chem. 2010, 285, 35740–35749. [Google Scholar] [CrossRef]
- Golubkov, V.S.; Prigozhina, N.L.; Zhang, Y.; Stoletov, K.; Lewis, J.D.; Schwartz, P.E.; Hoffman, R.M.; Strongin, A.Y. Protein-tyrosine pseudokinase 7 (ptk7) directs cancer cell motility and metastasis. J. Biol. Chem. 2014, 289, 24238–24249. [Google Scholar] [CrossRef]
- Hu, D.J.; Yun, J.; Elstrott, J.; Jasper, H. Non-canonical wnt signaling promotes directed migration of intestinal stem cells to sites of injury. Nat. Commun. 2021, 12, 7150. [Google Scholar] [CrossRef]
- Winberg, M.L.; Tamagnone, L.; Bai, J.; Comoglio, P.M.; Montell, D.; Goodman, C.S. The transmembrane protein off-track associates with plexins and functions downstream of semaphorin signaling during axon guidance. Neuron 2001, 32, 53–62. [Google Scholar] [CrossRef]
- Pulido, D.; Campuzano, S.; Koda, T.; Modolell, J.; Barbacid, M. Dtrk, a drosophila gene related to the trk family of neurotrophin receptors, encodes a novel class of neural cell adhesion molecule. EMBO J. 1992, 11, 391–404. [Google Scholar] [CrossRef]
- Bin-Nun, N.; Lichtig, H.; Malyarova, A.; Levy, M.; Elias, S.; Frank, D. Ptk7 modulates wnt signaling activity via lrp6. Development 2014, 141, 410–421. [Google Scholar] [CrossRef]
- Wehner, P.; Shnitsar, I.; Urlaub, H.; Borchers, A. Rack1 is a novel interaction partner of ptk7 that is required for neural tube closure. Development 2011, 138, 1321–1327. [Google Scholar] [CrossRef]
- Andreeva, A.; Lee, J.; Lohia, M.; Wu, X.; Macara, I.G.; Lu, X. Ptk7-src signaling at epithelial cell contacts mediates spatial organization of actomyosin and planar cell polarity. Dev. Cell 2014, 29, 20–33. [Google Scholar] [CrossRef]
- Llimargas, M.; Lawrence, P.A. Seven wnt homologues in drosophila: A case study of the developing tracheae. Proc. Natl. Acad. Sci. USA 2001, 98, 14487–14492. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.J.S.; Maugarny-Calès, A.; Pelletier, S.; Alexandre, C.; Bellaiche, Y.; Vincent, J.P.; McGough, I.J. Frizzled-dependent planar cell polarity without secreted wnt ligands. Dev. Cell 2020, 54, 583–592.e585. [Google Scholar] [CrossRef]
- Cohen, E.D.; Mariol, M.C.; Wallace, R.M.; Weyers, J.; Kamberov, Y.G.; Pradel, J.; Wilder, E.L. Dwnt4 regulates cell movement and focal adhesion kinase during drosophila ovarian morphogenesis. Dev. Cell 2002, 2, 437–448. [Google Scholar] [CrossRef]
- Gieseler, K.; Graba, Y.; Mariol, M.C.; Wilder, E.L.; Martinez-Arias, A.; Lemaire, P.; Pradel, J. Antagonist activity of dwnt-4 and wingless in the drosophila embryonic ventral ectoderm and in heterologous xenopus assays. Mech. Dev. 1999, 85, 123–131. [Google Scholar] [CrossRef]
- Gieseler, K.; Mariol, M.C.; Sagnier, T.; Graba, Y.; Pradel, J. Wingless and dwnt4, 2 drosophila wnt genes, have related expression, regulation and function during the embryonic development. Comptes Rendus Acad. Sci. III 1995, 318, 1101–1110. [Google Scholar]
- Inaki, M.; Yoshikawa, S.; Thomas, J.B.; Aburatani, H.; Nose, A. Wnt4 is a local repulsive cue that determines synaptic target specificity. Curr. Biol. 2007, 17, 1574–1579. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Nguyen, V.M.; Jeong, S. Regulation of off-track bidirectional signaling by semaphorin-1a and wnt signaling in the drosophila motor axon guidance. Insect Biochem. Mol. Biol. 2022, 150, 103857. [Google Scholar] [CrossRef]
- Ewen-Campen, B.; Comyn, T.; Vogt, E.; Perrimon, N. No evidence that wnt ligands are required for planar cell polarity in drosophila. Cell Rep. 2020, 32, 108121. [Google Scholar] [CrossRef]
- Wu, C.H.; Nusse, R. Ligand receptor interactions in the wnt signaling pathway in drosophila. J. Biol. Chem. 2002, 277, 41762–41769. [Google Scholar] [CrossRef]
- Yoshikawa, S.; McKinnon, R.D.; Kokel, M.; Thomas, J.B. Wnt-mediated axon guidance via the drosophila derailed receptor. Nature 2003, 422, 583–588. [Google Scholar] [CrossRef]
- Lim, C.H.; Kaur, P.; Teo, E.; Lam, V.Y.M.; Zhu, F.; Kibat, C.; Gruber, J.; Mathuru, A.S.; Tolwinski, N.S. Application of optogenetic amyloid-beta distinguishes between metabolic and physical damages in neurodegeneration. eLife 2020, 9, e52589. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Saunders, T.E.; Tolwinski, N.S. Coupling optogenetics and light-sheet microscopy, a method to study wnt signaling during embryogenesis. Sci. Rep. 2017, 7, 16636. [Google Scholar] [CrossRef] [PubMed]
- Gramates, L.S.; Agapite, J.; Attrill, H.; Calvi, B.R.; Crosby, M.A.; dos Santos, G.; Goodman, J.L.; Goutte-Gattat, D.; Jenkins, V.K.; Kaufman, T.; et al. Flybase: A guided tour of highlighted features. Genetics 2022, 220, iyac035. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, V.K.; Larkin, A.; Thurmond, J. Using flybase: A database of drosophila genes and genetics. Methods Mol. Biol. 2022, 2540, 1–34. [Google Scholar]
- Nagarkar-Jaiswal, S.; Lee, P.T.; Campbell, M.E.; Chen, K.; Anguiano-Zarate, S.; Gutierrez, M.C.; Busby, T.; Lin, W.W.; He, Y.; Schulze, K.L.; et al. A library of mimics allows tagging of genes and reversible, spatial and temporal knockdown of proteins in drosophila. eLife 2015, 4, e05338. [Google Scholar] [CrossRef]
- Lee, P.T.; Zirin, J.; Kanca, O.; Lin, W.W.; Schulze, K.L.; Li-Kroeger, D.; Tao, R.; Devereaux, C.; Hu, Y.; Chung, V.; et al. A gene-specific t2a-gal4 library for drosophila. eLife 2018, 7, e35574. [Google Scholar] [CrossRef]
- Li-Kroeger, D.; Kanca, O.; Lee, P.T.; Cowan, S.; Lee, M.T.; Jaiswal, M.; Salazar, J.L.; He, Y.; Zuo, Z.; Bellen, H.J. An expanded toolkit for gene tagging based on mimic and scarless crispr tagging in drosophila. eLife 2018, 7, e38709. [Google Scholar] [CrossRef]
- Khmelinskii, A.; Keller, P.J.; Bartosik, A.; Meurer, M.; Barry, J.D.; Mardin, B.R.; Kaufmann, A.; Trautmann, S.; Wachsmuth, M.; Pereira, G.; et al. Tandem fluorescent protein timers for in vivo analysis of protein dynamics. Nat. Biotechnol. 2012, 30, 708–714. [Google Scholar] [CrossRef]
- Khmelinskii, A.; Knop, M. Analysis of protein dynamics with tandem fluorescent protein timers. Methods Mol. Biol. 2014, 1174, 195–210. [Google Scholar]
- Perkins, L.A.; Holderbaum, L.; Tao, R.; Hu, Y.; Sopko, R.; McCall, K.; Yang-Zhou, D.; Flockhart, I.; Binari, R.; Shim, H.S.; et al. The transgenic rnai project at harvard medical school: Resources and validation. Genetics 2015, 201, 843–852. [Google Scholar] [CrossRef]
- Murakawa, T.; Nakamura, T.; Kawaguchi, K.; Murayama, F.; Zhao, N.; Stasevich, T.J.; Kimura, H.; Fujita, N. A drosophila toolkit for ha-tagged proteins unveils a block in autophagy flux in the last instar larval fat body. Development 2022, 149, dev200243. [Google Scholar] [CrossRef]
- Hays, R.; Gibori, G.B.; Bejsovec, A. Wingless signaling generates pattern through two distinct mechanisms. Development 1997, 124, 3727–3736. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Kibat, C.; Teo, E.; Gruber, J.; Mathuru, A.; Tolwinski, A.N.S. Use of optogenetic amyloid-β to monitor protein aggregation in drosophila melanogaster, danio rerio and caenorhabditis elegans. Bio Protoc. 2020, 10, e3856. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Bioinformatics, Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. Star: Ultrafast universal rna-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. Rsem: Accurate transcript quantification from rna-seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Kaller, M. Multiqc: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for rna-seq data with deseq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Zhu, A.; Ibrahim, J.G.; Love, M.I. Heavy-tailed prior distributions for sequence count data: Removing the noise and preserving large differences. Bioinformatics 2019, 35, 2084–2092. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. Clusterprofiler: An r package for comparing biological themes among gene clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Nusslein-Volhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef]
- DiNardo, S.; Heemskerk, J.; Dougan, S.; O’Farrell, P.H. The making of a maggot: Patterning the drosophila embryonic epidermis. Curr. Opin. Genet. Dev. 1994, 4, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Hatini, V.; DiNardo, S. Divide and conquer: Pattern formation in drosophila embryonic epidermis. Trends Genet. 2001, 17, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Gubb, D.; Garcia-Bellido, A. A genetic analysis of the determination of cuticular polarity during development in drosophila melanogaster. J. Embryol. Exp. Morphol. 1982, 68, 37–57. [Google Scholar]
- Schulte, J.; Charish, K.; Que, J.; Ravn, S.; MacKinnon, C.; Auld, V.J. Gliotactin and discs large form a protein complex at the tricellular junction of polarized epithelial cells in drosophila. J. Cell Sci. 2006, 119, 4391–4401. [Google Scholar] [CrossRef] [PubMed]
- Izumi, Y.; Yanagihashi, Y.; Furuse, M. A novel protein complex, mesh–ssk, is required for septate junction formation in the drosophila midgut. J. Cell Sci. 2012, 125, 4923–4933. [Google Scholar] [PubMed]
- Cafferty, P.; Yu, L.; Rao, Y. The receptor tyrosine kinase off-track is required for layer-specific neuronal connectivity in drosophila. Development 2004, 131, 5287–5295. [Google Scholar] [CrossRef] [PubMed]
- Diao, F.; Ironfield, H.; Luan, H.; Diao, F.; Shropshire, W.C.; Ewer, J.; Marr, E.; Potter, C.J.; Landgraf, M.; White, B.H. Plug-and-play genetic access to drosophila cell types using exchangeable exon cassettes. Cell Rep. 2015, 10, 1410–1421. [Google Scholar] [CrossRef]
- Durrieu, L.; Kirrmaier, D.; Schneidt, T.; Kats, I.; Raghavan, S.; Hufnagel, L.; Saunders, T.E.; Knop, M. Bicoid gradient formation mechanism and dynamics revealed by protein lifetime analysis. Mol. Syst. Biol. 2018, 14, e8355. [Google Scholar] [CrossRef]
- Hartenstein, V. Atlas of Drosophila Development; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1993. [Google Scholar]
- Zhao, N.; Kamijo, K.; Fox, P.D.; Oda, H.; Morisaki, T.; Sato, Y.; Kimura, H.; Stasevich, T.J. A genetically encoded probe for imaging nascent and mature ha-tagged proteins in vivo. Nat. Commun. 2019, 10, 2947. [Google Scholar] [CrossRef]
- Campos, A.R.; Rosen, D.R.; Robinow, S.N.; White, K. Molecular analysis of the locus elav in drosophila melanogaster: A gene whose embryonic expression is neural specific. EMBO J. 1987, 6, 425–431. [Google Scholar] [CrossRef]
- Lin, D.M.; Goodman, C.S. Ectopic and increased expression of fasciclin ii alters motoneuron growth cone guidance. Neuron 1994, 13, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Hansen, S.; Morris, O.; Madden, D.T.; Libeu, C.P.; Kumar, A.J.; Wehrfritz, C.; Nile, A.H.; Zhang, Y.; Zhou, L.; et al. Senescent cells perturb intestinal stem cell differentiation through ptk7 induced noncanonical wnt and yap signaling. Nat. Commun. 2023, 14, 156. [Google Scholar] [CrossRef] [PubMed]
- Martinez, S.; Scerbo, P.; Giordano, M.; Daulat, A.M.; Lhoumeau, A.C.; Thome, V.; Kodjabachian, L.; Borg, J.P. The ptk7 and ror2 protein receptors interact in the vertebrate wnt/planar cell polarity (pcp) pathway. J. Biol. Chem. 2015, 290, 30562–30572. [Google Scholar] [CrossRef] [PubMed]
- Oishi, I.; Suzuki, H.; Onishi, N.; Takada, R.; Kani, S.; Ohkawara, B.; Koshida, I.; Suzuki, K.; Yamada, G.; Schwabe, G.C. The receptor tyrosine kinase ror2 is involved in non-canonical wnt5a/jnk signalling pathway. Genes Cells 2003, 8, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Podleschny, M.; Grund, A.; Berger, H.; Rollwitz, E.; Borchers, A. A ptk7/ror2 co-receptor complex affects xenopus neural crest migration. PLoS ONE 2015, 10, e0145169. [Google Scholar] [CrossRef] [PubMed]
- Ripp, C.; Loth, J.; Petrova, I.; Linnemannstons, K.; Ulepic, M.; Fradkin, L.; Noordermeer, J.; Wodarz, A. Drosophila ror is a nervous system-specific co-receptor for wnt ligands. Biol. Open 2018, 7, bio033001. [Google Scholar]
- Tolwinski, N.S.; Wieschaus, E. A nuclear escort for beta-catenin. Nat. Cell Biol. 2004, 6, 579–580. [Google Scholar] [CrossRef] [PubMed]
- Tolwinski, N.S.; Wieschaus, E. A nuclear function for armadillo/beta-catenin. PLoS Biol. 2004, 2, E95. [Google Scholar] [CrossRef]
- Roegiers, F.; Kavaler, J.; Tolwinski, N.; Chou, Y.T.; Duan, H.; Bejarano, F.; Zitserman, D.; Lai, E.C. Frequent unanticipated alleles of lethal giant larvae in drosophila second chromosome stocks. Genetics 2009, 182, 407–410. [Google Scholar] [CrossRef]
- Puppo, F.; Thome, V.; Lhoumeau, A.C.; Cibois, M.; Gangar, A.; Lembo, F.; Belotti, E.; Marchetto, S.; Lecine, P.; Prebet, T.; et al. Protein tyrosine kinase 7 has a conserved role in wnt/beta-catenin canonical signalling. EMBO Rep. 2011, 12, 43–49. [Google Scholar] [CrossRef]
- Kani, S.; Oishi, I.; Yamamoto, H.; Yoda, A.; Suzuki, H.; Nomachi, A.; Iozumi, K.; Nishita, M.; Kikuchi, A.; Takumi, T.; et al. The receptor tyrosine kinase ror2 associates with and is activated by casein kinase iepsilon. J. Biol. Chem. 2004, 279, 50102–50109. [Google Scholar] [CrossRef] [PubMed]
- Brugge, J.S.; Erikson, R.L. Identification of a transformation-specific antigen induced by an avian sarcoma virus. Nature 1977, 269, 346–348. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Lhoumeau, A.C.; Arcangeli, M.L.; De Grandis, M.; Giordano, M.; Orsoni, J.C.; Lembo, F.; Bardin, F.; Marchetto, S.; Aurrand-Lions, M.; Borg, J.P. Ptk7-deficient mice have decreased hematopoietic stem cell pools as a result of deregulated proliferation and migration. J. Immunol. 2016, 196, 4367–4377. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, Q.H.; Otgonbaatar, A.; Kaur, P.; Ga, A.F.; Harmston, N.P.; Tolwinski, N.S. The Wnt Co-Receptor PTK7/Otk and Its Homolog Otk-2 in Neurogenesis and Patterning. Cells 2024, 13, 365. https://doi.org/10.3390/cells13050365
Tan QH, Otgonbaatar A, Kaur P, Ga AF, Harmston NP, Tolwinski NS. The Wnt Co-Receptor PTK7/Otk and Its Homolog Otk-2 in Neurogenesis and Patterning. Cells. 2024; 13(5):365. https://doi.org/10.3390/cells13050365
Chicago/Turabian StyleTan, Qian Hui, Agimaa Otgonbaatar, Prameet Kaur, Angelica Faye Ga, Nathan P. Harmston, and Nicholas S. Tolwinski. 2024. "The Wnt Co-Receptor PTK7/Otk and Its Homolog Otk-2 in Neurogenesis and Patterning" Cells 13, no. 5: 365. https://doi.org/10.3390/cells13050365