

Effects of Simulated Microgravity In Vitro on Human Metaphase II Oocytes: An Electron Microscopy-Based Study
 , , , ,
, , , ,  , and
, and
Abstract
:Highlights
- For the first time, the in vitro effects of simulated microgravity using a Random Positioning Machine on the ultrastructural features of human metaphase II oocytes were investigated by transmission electron microscopy.
- Microgravity might compromise oocyte quality affecting the ultrastructural morphology of mitochondria, endoplasmic reticulum, and cortical granules due to a possible alteration of the cytoskeleton.
- Microgravity-induced organelle modifications potentially affect the oocyte’s ability to fully mature and fertilize and develop into a viable embryo.
Abstract
1. Introduction
2. Materials and Methods
2.1. Oocyte Recovery
2.2. Clinorotation to Simulate Microgravity
2.3. Microgravity Exposure and Cell Treatments
2.4. Light/Electron Microscopy
2.5. Statistical Analysis
3. Results
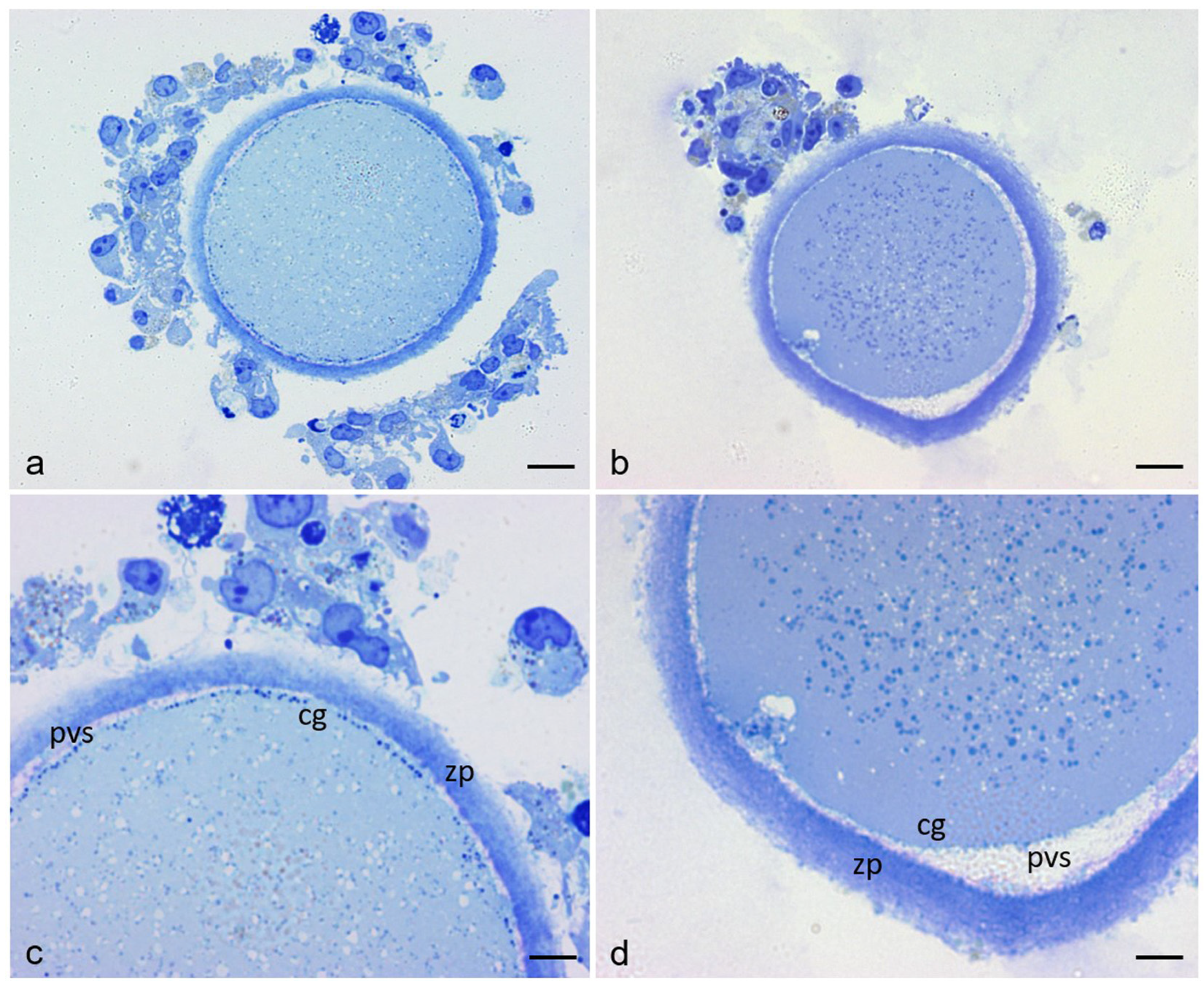
3.1. Sample Characteristics by Light Microscope
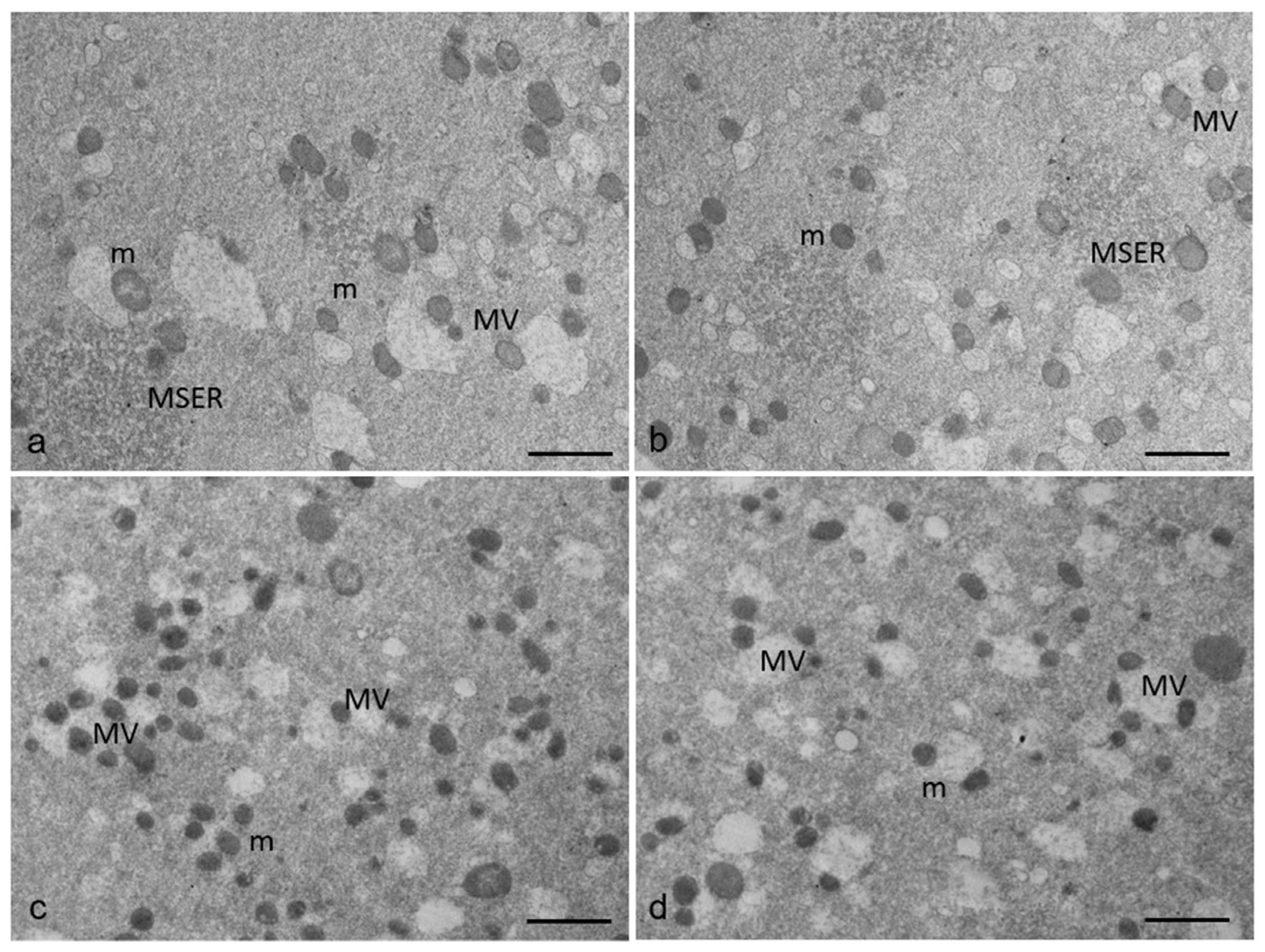
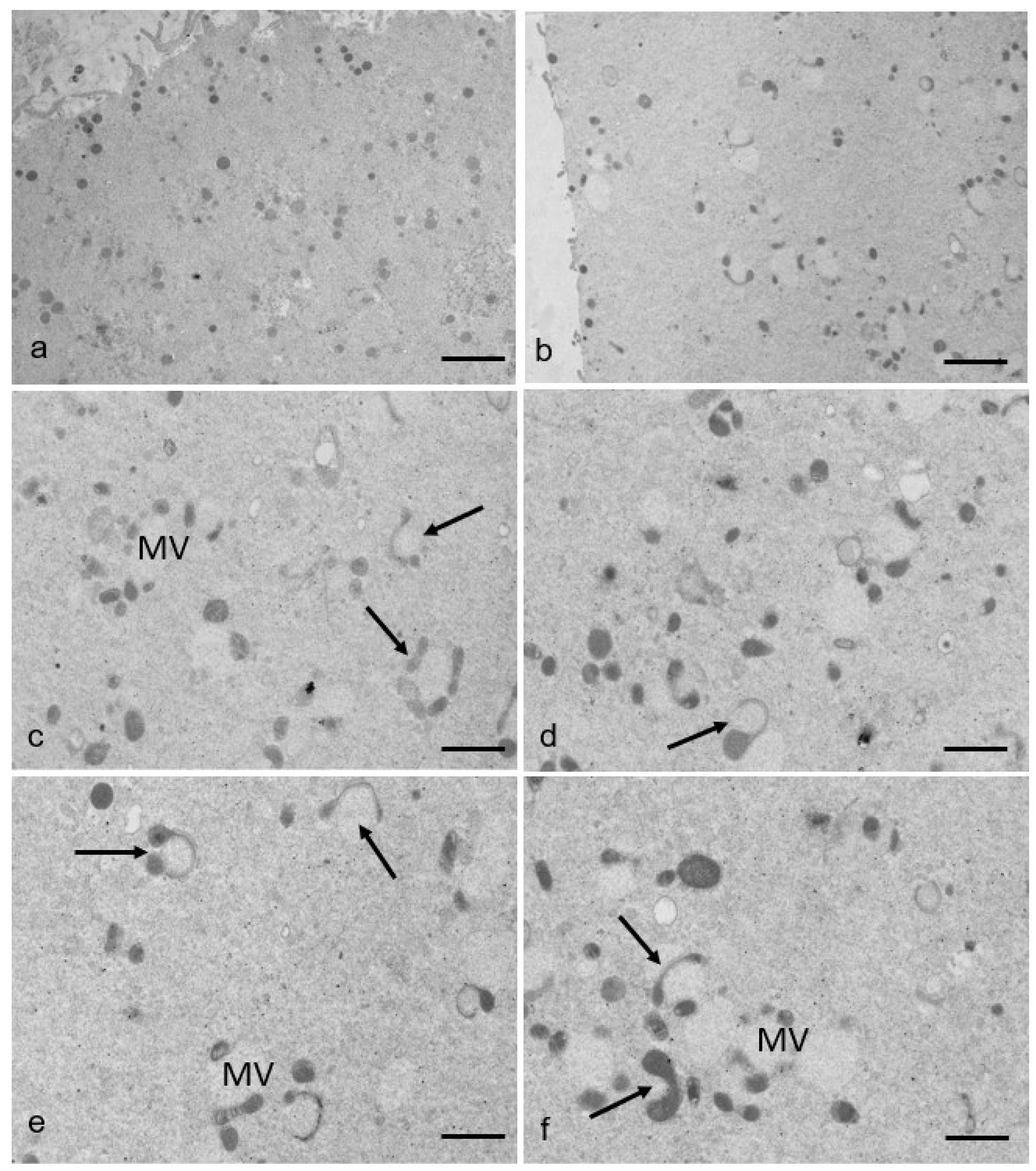
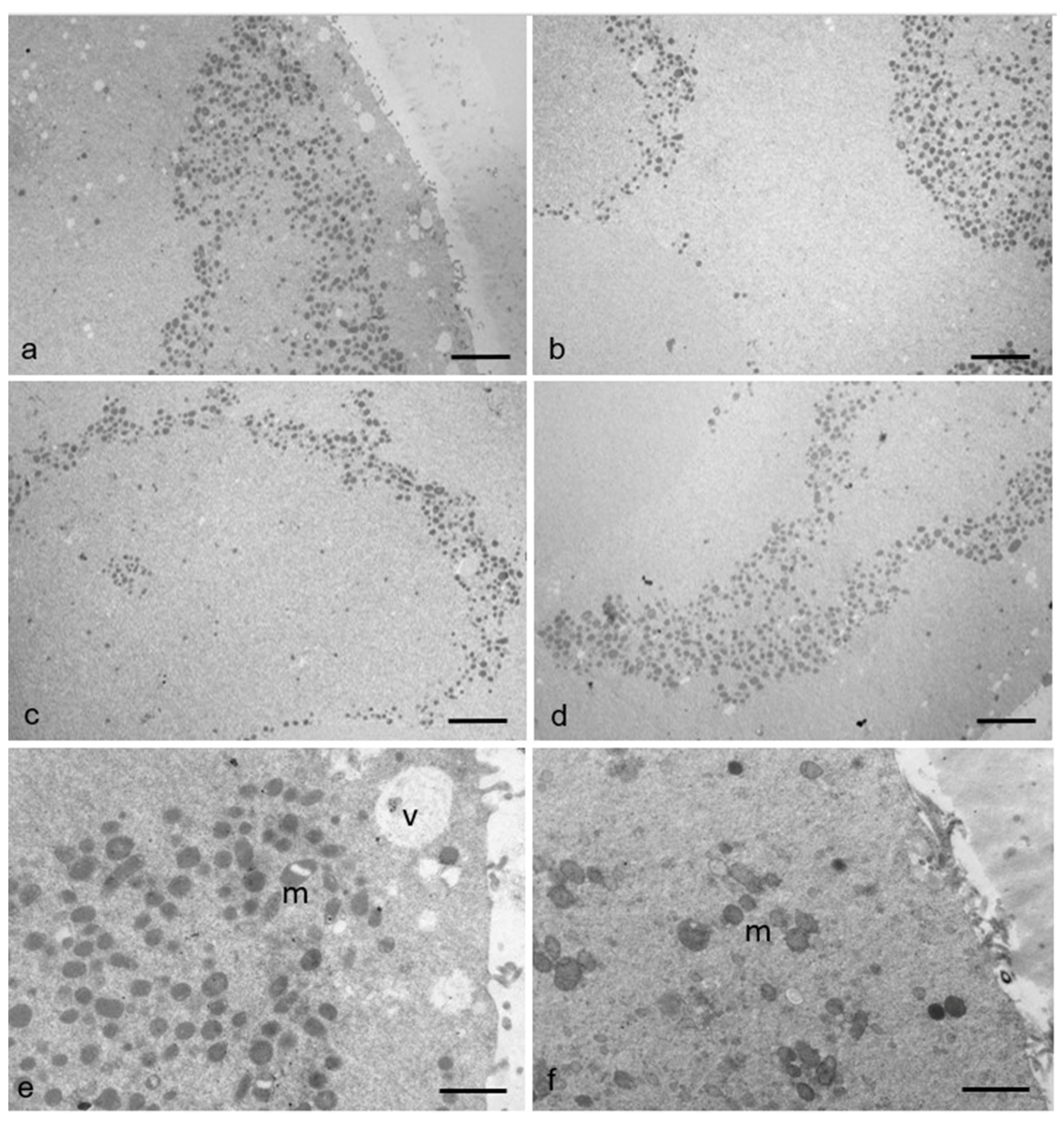
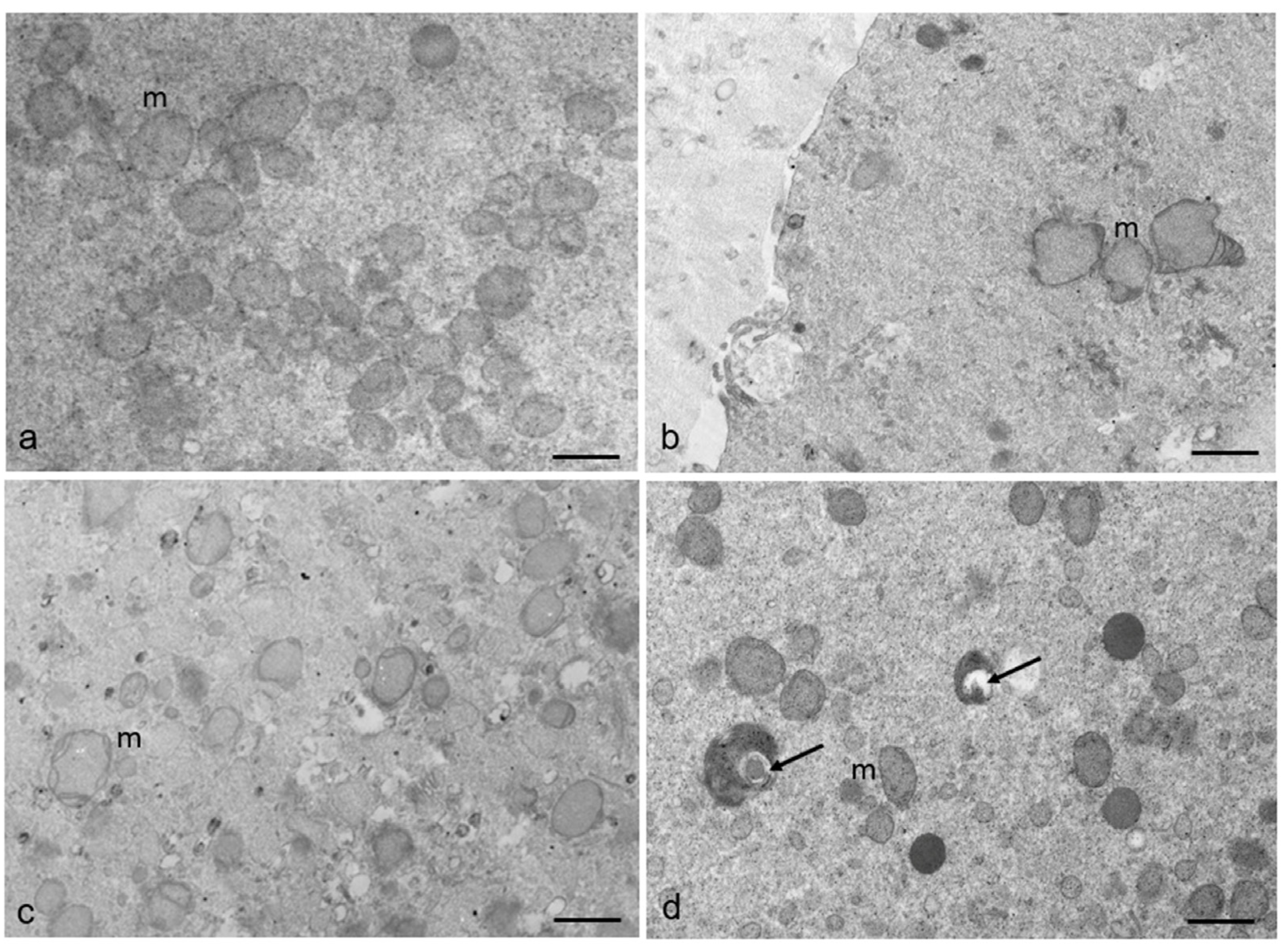
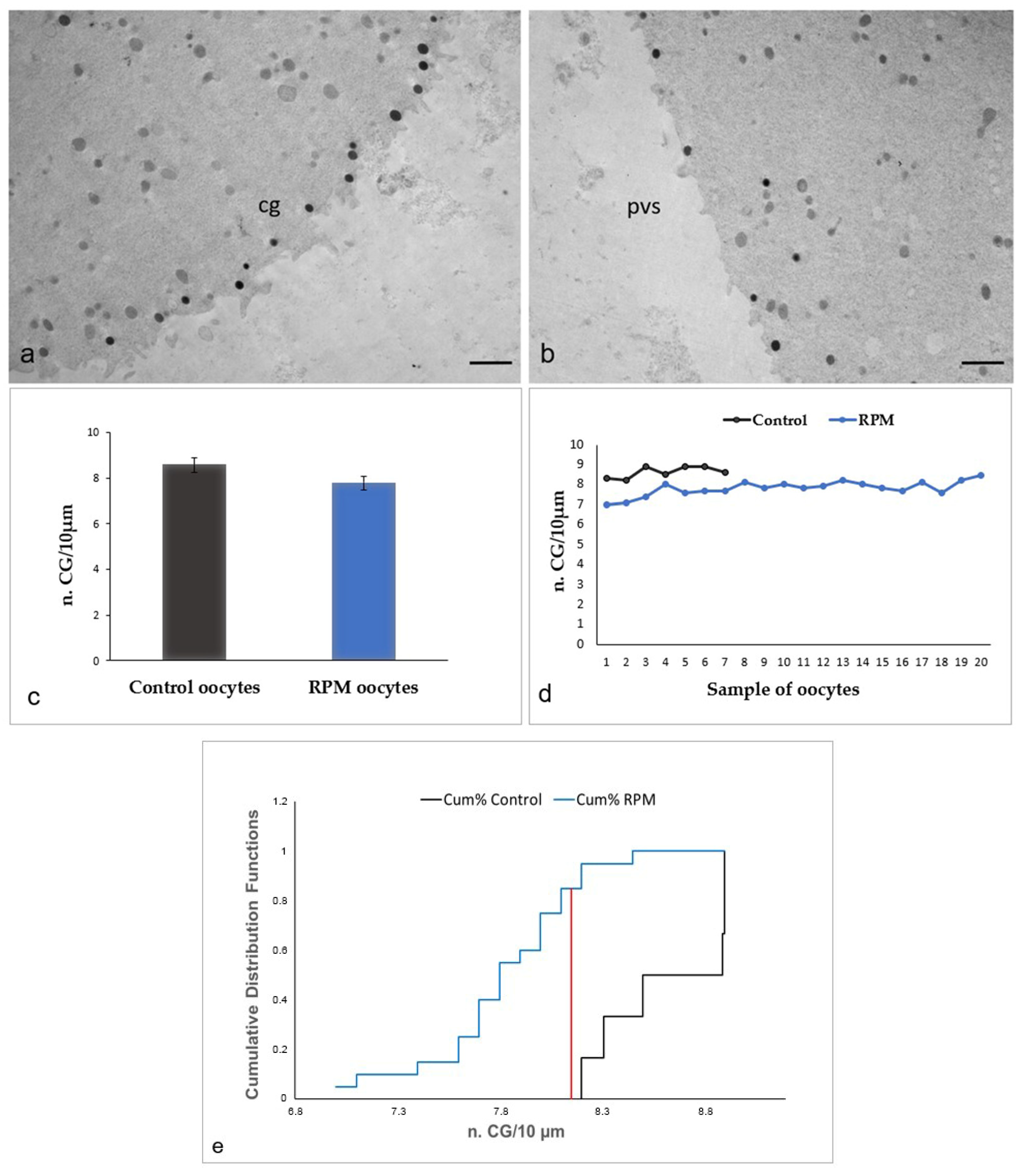
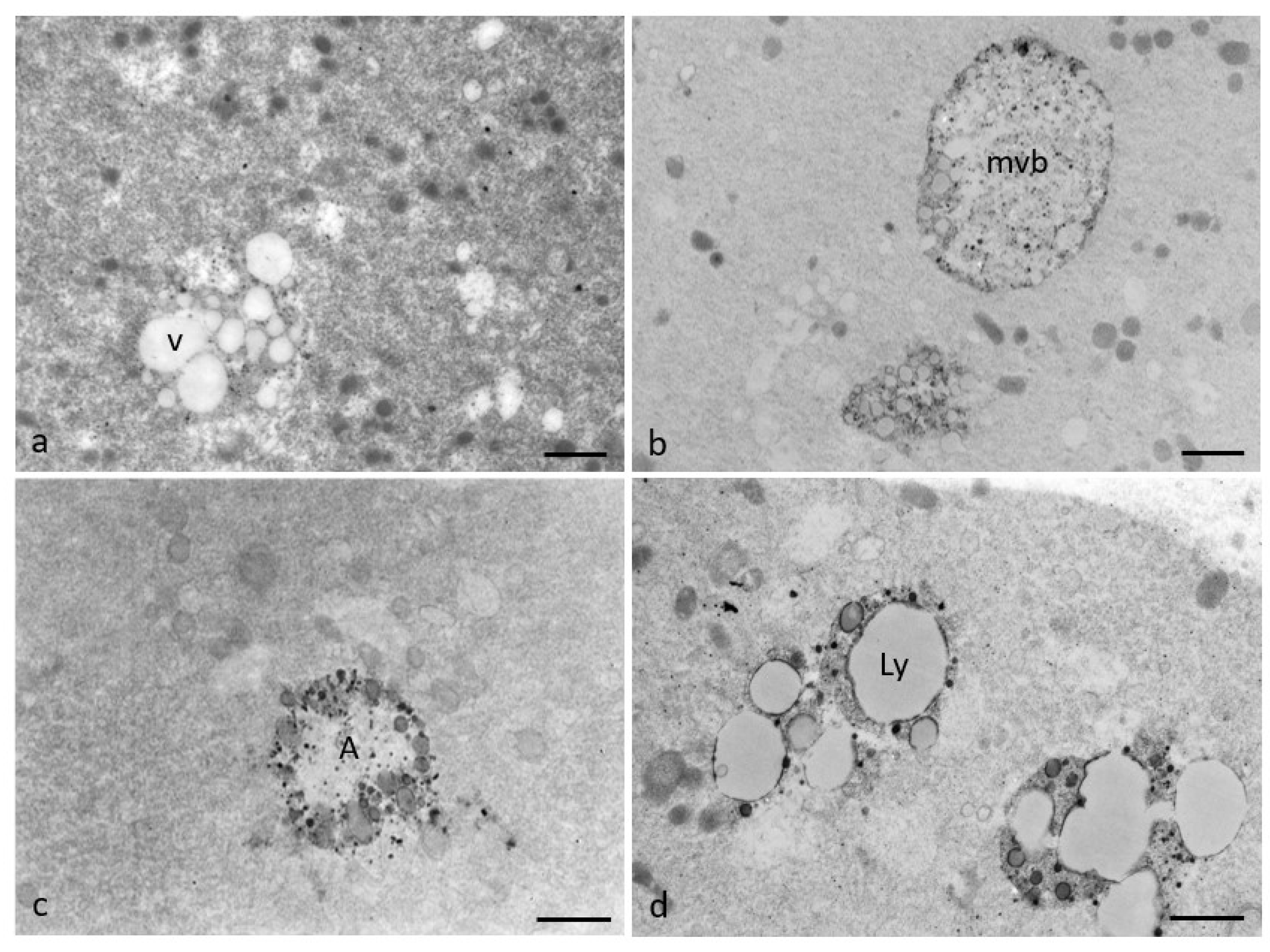
3.2. Ultrastructural Features in the Ooplasmic Compartment
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanaka, K.; Nishimura, N.; Kawai, Y. Adaptation to microgravity, deconditioning, and countermeasures. J. Physiol. Sci. 2017, 67, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.; Wu, H.; Choi, J.U.; Rowan, A.E.; Zhang, H.; Poole, K.; Lauko, J.; Chou, J. Modeling the Impact of Microgravity at the Cellular Level: Implications for Human Disease. Front. Cell Dev. Biol. 2020, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Zhang, K.; Wei, D.; Tian, Y.; Gao, Y.; Chen, Z.; Qian, A. The Impact of Spaceflight and Simulated Microgravity on Cell Adhesion. Int. J. Mol. Sci. 2020, 21, 3031. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Vaquer, S.; Mazzolai, L.; Roberts, L.N.; Pavela, J.; Watanabe, M.; Weerts, G.; Green, D.A. The effect of microgravity on the human venous system and blood coagulation: A systematic review. Exp. Physiol. 2021, 106, 1149–1158. [Google Scholar] [CrossRef]
- Ferranti, F.; Caruso, M.; Cammarota, M.; Masiello, M.G.; Corano Scheri, K.; Fabrizi, C.; Fumagalli, L.; Schiraldi, C.; Cucina, A.; Catizone, A.; et al. Cytoskeleton Modifications and Autophagy Induction in TCam-2 Seminoma Cells Exposed to Simulated Microgravity. Biomed. Res. Int. 2014, 2014, 904396. [Google Scholar] [CrossRef]
- Ingber, D. How cells (might) sense microgravity. Faseb J. 1999, 13 (Suppl. S3–S15), 1169–1183. [Google Scholar] [CrossRef]
- Crawford-Young, S.J. Effects of microgravity on cell cytoskeleton and embryogenesis. Int. J. Dev. Biol. 2006, 50, 183–191. [Google Scholar] [CrossRef]
- Schatten, H.; Lewis, M.L.; Chakrabari, A. Spaceflight and clinorotation cause cytoskeleton and mitochondria changes and increases in apoptosis in cultured cells. Acta Astronaut. 2001, 49, 399–418. [Google Scholar] [CrossRef]
- Michaletti, A.; Gioia, M.; Tarantino, U.; Zolla, L. Effects of microgravity on osteoblast mitochondria: A proteomic and metabolomics profile. Sci. Rep. 2017, 7, 15376. [Google Scholar] [CrossRef]
- Zhang, R.; Ran, H.-H.; Cai, L.-L.; Zhu, L.; Sun, J.-F.; Peng, L.; Liu, X.-J.; Zhang, L.-N.; Fang, Z.; Fan, Y.-Y.; et al. Simulated microgravity-induced mitochondrial dysfunction in rat cerebral arteries. FASEB J. 2014, 28, 2715–2724. [Google Scholar] [CrossRef]
- Nguyen, H.P.; Tran, H.P.; Kim, K.-S.; Yang, S.-G. The effects of real and simulated microgravity on cellular mitochondrial function. NPJ Microgravity 2021, 7, 44. [Google Scholar] [CrossRef]
- Kast, D.J.; Dominguez, R. The Cytoskeleton-Autophagy Connection. Curr. Biol. 2017, 27, R318–R326. [Google Scholar] [CrossRef] [PubMed]
- Tou, J.; Ronca, A.; Grindeland, R.; Wade, C. Models to Study Gravitational Biology of Mammalian Reproduction. Biol. Reprod. 2002, 67, 1681–1687. [Google Scholar] [CrossRef]
- Ruden, D.; Bolnick, A.; Awonugar, A.; Abdulhasan, M.; Perez, G.; Puscheck, E.E.; Rappolee, D.A. Effects of gravity, microgravity or microgravity simulation on early mammalian development. Stem. Cells Dev. 2018, 1230–1236. [Google Scholar] [CrossRef]
- Liu, S.; Li, Y.; Feng, H.L.; Yan, J.H.; Li, M.; Ma, S.Y.; Chen, Z.J. Dynamic modulation of cytoskeleton during in vitro maturation in human oocytes. Am. J. Obs. Gynecol. 2010, 203, e1–e7. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, D.; Wu, Y.; Lin, W.; Chen, Z.; Meng, L.; Liu, J.; Zhou, Y. Simulated Microgravity Using a Rotary Culture System Compromises the In Vitro Development of Mouse Preantral Follicles. PLoS ONE 2016, 11, e0151062. [Google Scholar] [CrossRef] [PubMed]
- Dal Canto, M.; Guglielmo, M.C.I.; Renzini, M.M.; Fadini, R.; Moutier, C.; Merola, M.; De Ponti, E.; Coticchio, G. Dysmorphic patterns are associated with cytoskeletal alterations in human oocytes. Hum. Reprod. 2017, 32, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Goswami, N.; White, O.; Blaber, A.; Evans, J.; van Loon, J.J.W.A.; Clement, G. Human physiology adaptation to altered gravity environments. Acta Astronaut. 2021, 189, 216–221. [Google Scholar] [CrossRef]
- Borst, A.G.; van Loon, J.J.W.A. Technology and Developments for the Random Positioning Machine, RPM. Microgravity Sci. Technol. 2009, 21, 287–292. [Google Scholar] [CrossRef]
- Masiello, M.G.; Cucina, A.; Proietti, S.; Palombo, A.; Coluccia, P.; D’Anselmi, F.; Dinicola, S.; Pasqualato, A.; Morini, V.; Bizzarri, M. Phenotypic Switch Induced by Simulated Microgravity on MDA-MB-231 Breast Cancer Cells. BioMed. Res. Int. 2014, 2014, 652434. [Google Scholar] [CrossRef]
- Coticchio, G.; Dal Canto, M.; Fadini, R.; Renzini, M.M.; Guglielmo, M.C.; Miglietta, S.; Palmerini, M.G.; Macchiarelli, G.; Nottola, S.A. Ultrastructure of human oocytes after in vitro maturation. Mol. Hum. Reprod. 2016, 22, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Luderer, U. Reproductive hazards of space travel in women and men. Nat. Rev. Endocrinol. 2019, 15, 713–730. [Google Scholar] [CrossRef] [PubMed]
- Drago-Ferrante, R.; Di Fiore, R.; Karouia, F.; Subbannayya, Y.; Das, S.; Aydogan Mathyk, B.; Arif, S.; Guevara-Cerdán, A.P.; Seylani, A.; Galsinh, A.S.; et al. Extraterrestrial Gynecology: Could Spaceflight Increase the Risk of Developing Cancer in Female Astronauts? An Updated Review. Int. J. Mol. Sci. 2022, 23, 7465. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Guo, X.; Wang, F.; Li, X.; Tian, X.C.; Li, L.; Wu, Z.; Zhang, S. Simulated Microgravity Compromises Mouse Oocyte Maturation by Disrupting Meiotic Spindle Organization and Inducing Cytoplasmic Blebbing. PLoS ONE 2011, 6, e22214. [Google Scholar] [CrossRef]
- Cheng, K.; Feng, X.; Yang, C.; Ma, C.; Niu, S.; Jia, L.; Yang, X.; Liang, J.; Bo, Y.; Geng, K.; et al. Simulated microgravity reduces quality of ovarian follicles and oocytes by disrupting communications of follicle cells. NPJ Microgravity 2023, 9, 7. [Google Scholar] [CrossRef]
- Nottola, S.A.; Macchiarelli, G.; Coticchio, G.; Bianchi, S.; Cecconi, S.; De Santis, L.; Scaravelli, G.; Flamigni, C.; Borini, A. Ultrastructure of human mature oocytes after slow cooling cryopreservation using different sucrose concentrations. Hum. Reprod. 2007, 22, 1123–1133. [Google Scholar] [CrossRef]
- Coticchio, G.; Dal Canto, M.; Mignini Renzini, M.; Guglielmo, M.C.; Brambillasca, F.; Turchi, D.; Novara, P.V.; Fadini, R. Oocyte maturation: Gamete-somatic cells interactions, meiotic resumption, cytoskeletal dynamics and cytoplasmic reorganization. Hum. Reprod. Update 2015, 21, 427–454. [Google Scholar] [CrossRef]
- Nottola, S.A.; Coticchio, G.; De Santis, L.; Macchiarelli, G.; Maione, M.; Bianchi, S.; Iaccarino, M.; Flamigni, C.; Borini, A. Ultrastructure of human mature oocytes after slow cooling cryopreservation with ethylene glycol. Reprod. BioMed. Online 2008, 17, 368–377. [Google Scholar] [CrossRef]
- Nottola, S.A.; Coticchio, G.; Sciajno, R.; Gambardella, A.; Maione, M.; Scaravelli, G.; Bianchi, S.; Macchiarelli, G.; Borini, A. Ultrastructural markers of quality in human mature oocytes vitrified using cryoleaf and cryoloop. Reprod. BioMed. 2009, 19 (Suppl. S3), 17–27. [Google Scholar] [CrossRef]
- Coticchio, G.; Borini, A.; Distratis, V.; Maione, M.; Scaravelli, G.; Bianchi, V.; Macchiarelli, G.; Nottola, S. Qualitative and morphometric analysis of the ultrastructure of human oocytes cryopreserved by two alternative slow cooling protocols. J. Assist. Reprod. Genet. 2010, 27, 131–140. [Google Scholar] [CrossRef]
- Bianchi, V.; Macchiarelli, G.; Borini, A.; Lappi, M.; Cecconi, S.; Miglietta, S.; Familiari, G.; Nottola, S.A. Fine morphological assessment of quality of human mature oocytes after slow freezing or vitrification with a closed device: A comparative analysis. Reprod. Biol. Endocrinol. 2014, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- De Santis, L.; Nottola, S.A.; Coticchio, G.; Borini, A.; Iussig, B.; Miglietta, S.; Macchiarelli, G. Type of protein supplement in cryopreservation solutions impacts on the degree of ultrastructural damage in frozen-thawed human oocytes. Cryobiology 2020, 95, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10, 2484. [Google Scholar] [CrossRef] [PubMed]
- Trebichalská, Z.; Kyjovská, D.; Kloudová, S.; Otevřel, P.; Hampl, A.; Holubcová, Z. Cytoplasmic maturation in human oocytes: An ultrastructural study. Biol. Reprod. 2021, 104, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, S.; Macchiarelli, G.; Micara, G.; Linari, A.; Boninsegna, C.; Aragona, C.; Rossi, G.; Cecconi, S.; Nottola, S.A. Ultrastructural markers of quality are impaired in human metaphase II aged oocytes: A comparison between reproductive and in vitro aging. J. Assist. Reprod. Genet. 2015, 32, 1343–1358. [Google Scholar] [CrossRef]
- Nottola, S.A.; Albani, E.; Coticchio, G.; Palmerini, M.G.; Lorenzo, C.; Scaravelli, G.; Borini, A.; Levi-Setti, P.E.; Macchiarelli, G. Freeze/thaw stress induces organelle remodeling and membrane recycling in cryopreserved human mature oocytes. J. Assist. Reprod. Genet. 2016, 33, 1559–1570. [Google Scholar] [CrossRef] [PubMed]
- Motta, P.M.; Nottola, S.A.; Micara, G.; Familiari, G. Ultrastructure of human unfertilized oocytes and polyspermic embryos in an IVF-ET program. Ann. N. Y. Acad. Sci. 1988, 541, 367–383. [Google Scholar] [CrossRef]
- Familiari, G.; Heyn, R.; Relucenti, M.; Nottola, S.A.; Sathananthan, A.H. Ultrastructural dynamics of human reproduction, from ovulation to fertilization and early embryo development. Int. Rev. Cytol. 2006, 249, 53–141. [Google Scholar] [CrossRef]
- Sousa, M.; Barros, A.; Silva, J.; Tesarik, J. Developmental changes in calcium content of ultrastructurally distinct subcellular compartments of preimplantation human embryos. Mol. Hum. Reprod. 1997, 3, 83–90. [Google Scholar] [CrossRef]
- Cheng, J.P.; Lane, J.D. Organelle dynamics and membrane trafficking in apoptosis and autophagy. Histol. Histopathol. 2010, 25, 1457–1472. [Google Scholar] [CrossRef]
- Farkas, Á.; Farkas, G. Effects of Spaceflight on Human Skin. Pharm. Physiol. 2021, 34, 239–245. [Google Scholar] [CrossRef]
- Takahashi, H.; Nakamura, A.; Shimizu, T. Simulated microgravity accelerates aging of human skeletal muscle myoblasts at the single cell level. Biochem. Biophys. Res. Commun. 2021, 578, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Feger, B.J.; Thompson, J.W.; Dubois, L.G.; Kommaddi, R.P.; Foster, M.W.; Mishra, R.; Shenoy, S.K.; Shibata, Y.; Kidane, Y.H.; Moseley, M.A.; et al. Microgravity induces proteomics changes involved in endoplasmic reticulum stress and mitochondrial protection. Sci. Rep. 2016, 6, 34091. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, H.; Liu, Z.; Lin, L.; Wang, L.; Xie, M.; Li, M.; Zhang, J.; Zhang, R. Endoplasmic reticulum stress-dependent activation of iNOS/NO-NF-κB signaling and NLRP3 inflammasome contributes to endothelial inflammation and apoptosis associated with microgravity. FASEB J. 2020, 34, 10835–10849. [Google Scholar] [CrossRef]
- Palmerini, M.G.; Nottola, S.A.; Leoni, G.G.; Succu, S.; Borshi, X.; Berlinguer, F.; Naitana, S.; Bekmukhambetov, Y.; Macchiarelli, G. In vitro maturation is slowed in prepubertal lamb oocytes: Ultrastructural evidences. Reprod. Biol. Endocrinol. 2014, 12, 115. [Google Scholar] [CrossRef] [PubMed]
- Reader, K.L.; Stanton, J.A.L.; Juengel, J.L. The Role of Oocyte Organelles in Determining Developmental Competence. Biology 2017, 6, 35. [Google Scholar] [CrossRef]
- Youle, R.J.; van der Bliek, A.M. Mitochondrial Fission, Fusion, and Stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Reddy, P.H.; Iijima, M.; Sesaki, H. Mitochondrial Division and Fusion in Metabolism. Curr. Opin. Cell Biol. 2015, 33, 111–118. [Google Scholar] [CrossRef]
- Suen, D.F.; Narendra, D.P.; Youle, R.J. Mitochondrial dynamics and apoptosis. Genes. Dev. 2017, 30, 327–349. [Google Scholar] [CrossRef]
- Jeong, A.J.; Kim, Y.J.; Lim, M.H.; Lee, H.; Noh, K.; Kim, B.-H.; Chung, J.W.; Cho, C.-H.; Kim, S.; Ye, S.-K. Microgravity induces autophagy via mitochondrial dysfunction in human Hodgkin’s lymphoma cells. Sci. Rep. 2018, 8, 14646. [Google Scholar] [CrossRef]
- Locatelli, L.; Cazzaniga, A.; De Palma, C.; Castiglioni, S.; Maier, J.A.M. Mitophagy contributes to endothelial adaptation to simulated microgravity. FASEB J. 2020, 34, 1833–1845. [Google Scholar] [CrossRef] [PubMed]
- Hoppeler, H.; Fluck, M. Plasticity of skeletal muscle mitochondria: Structure and function. Med. Sci. Sports Exerc. 2003, 35, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Adams, S.H.; Hoppel, C.L.; Lok, K.H.; Zhao, L.; Wong, S.W.; Minkler, P.E.; Hwang, D.H.; Newman, J.W.; Garvey, W.T. Plasma acylcarnitine profiles suggest incomplete long-chain fatty acid beta-oxidation and altered tricarboxylic acid cycle activity in type 2 diabetic African American women. J. Nutr. 2009, 139, 1073–1081. [Google Scholar] [CrossRef]
- Motta, P.M.; Nottola, S.A.; Makabe, S.; Heyn, R. Mitochondrial morphology in human fetal and adult female germ cells. Hum. Reprod. 2000, 2, 129–147. [Google Scholar] [CrossRef] [PubMed]
- Uva, B.M.; Masini, M.A.; Sturla, M.; Prato, P.; Passalacqua, M.; Giuliani, M.; Tagliafierro, G.; Strollo, F. Clinoration-induced weightlessness Influences the cytoskeleton of glial cells in culture. Brain Res. 2002, 934, 132–139. [Google Scholar] [CrossRef]
- de Bruin, J.P.; Dorland, M.; Spek, E.R.; Posthuma, G.; van Haaften, M.; Looman, C.W.; te Velde, E.R. Age-related changes in the ultrastructure of the resting follicle pool in human ovaries. Biol. Reprod. 2004, 70, 419–424. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphometric Parameters | Control Oocytes | RPM Oocytes | p-Values of Significance |
|---|---|---|---|
| Mean ± SD | Mean ± SD | ||
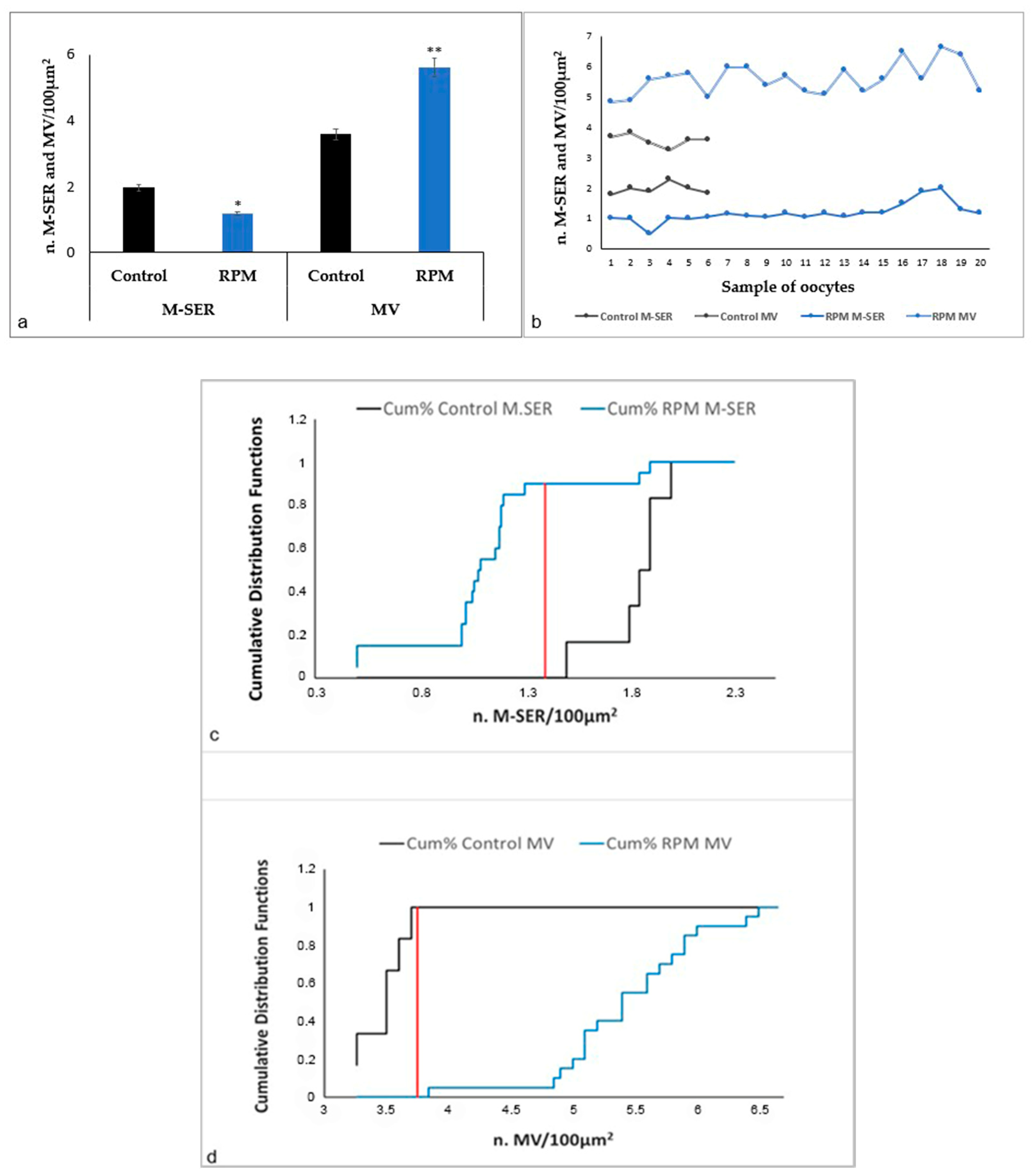
| n. M-SER/100 µm2 | 1.97 ± 0.16 | 1.18 ± 0.31 | p < 0.05 |
| n. MV/100 µm2 | 3.58 ± 0.17 | 5.61 ± 0.52 | p < 0.01 |
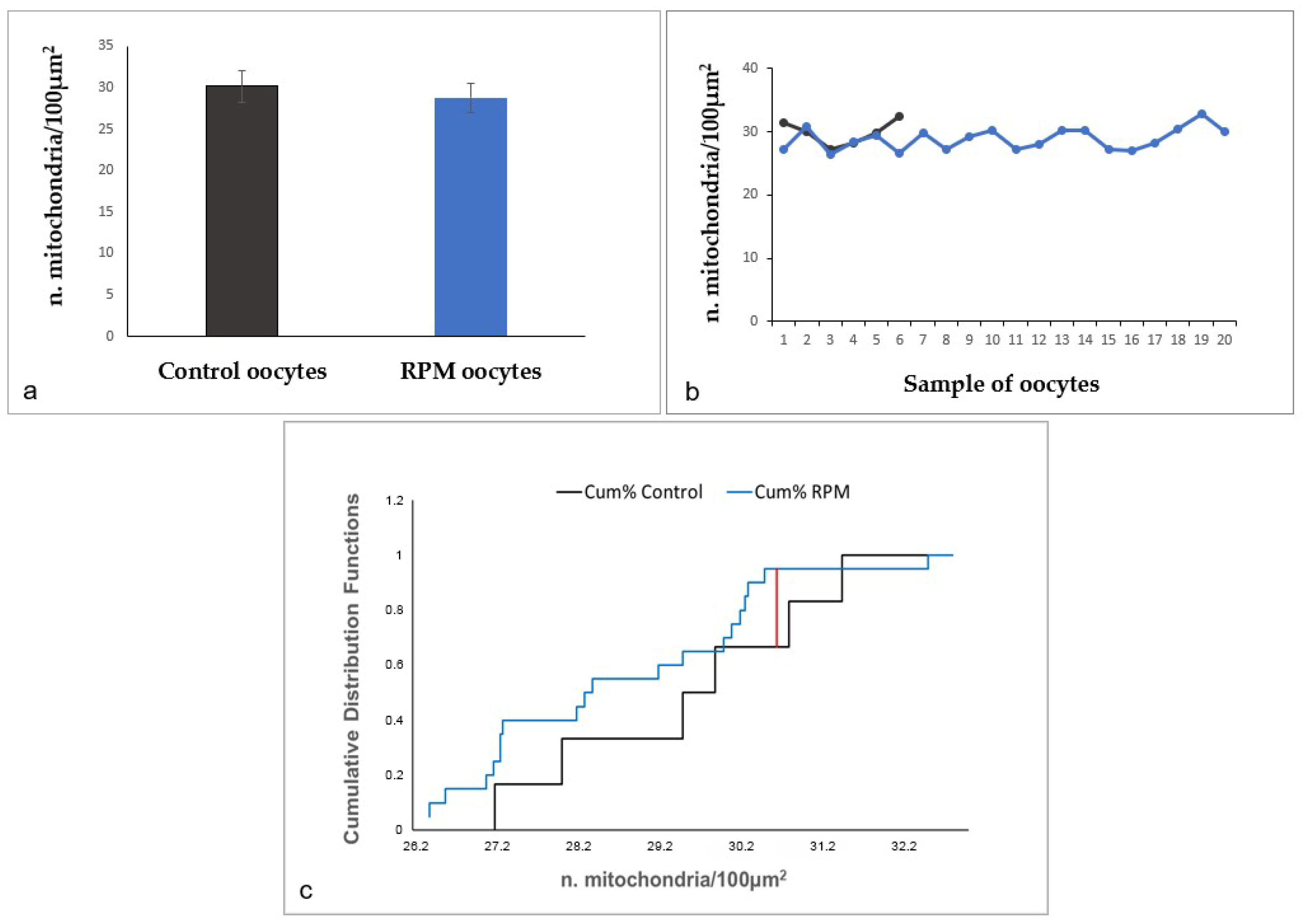
| n. mitochondria/100 µm2 | 29.88 ± 1.94 | 28.86 ± 1.73 | p = 0.29 |
| n. CG/10 µm | 8.61 ± 0.29 | 7.80 ± 0.35 | p = 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miglietta, S.; Cristiano, L.; Espinola, M.S.B.; Masiello, M.G.; Micara, G.; Battaglione, E.; Linari, A.; Palmerini, M.G.; Familiari, G.; Aragona, C.; et al. Effects of Simulated Microgravity In Vitro on Human Metaphase II Oocytes: An Electron Microscopy-Based Study. Cells 2023, 12, 1346. https://doi.org/10.3390/cells12101346
Miglietta S, Cristiano L, Espinola MSB, Masiello MG, Micara G, Battaglione E, Linari A, Palmerini MG, Familiari G, Aragona C, et al. Effects of Simulated Microgravity In Vitro on Human Metaphase II Oocytes: An Electron Microscopy-Based Study. Cells. 2023; 12(10):1346. https://doi.org/10.3390/cells12101346
Chicago/Turabian StyleMiglietta, Selenia, Loredana Cristiano, Maria Salomé B. Espinola, Maria Grazia Masiello, Giulietta Micara, Ezio Battaglione, Antonella Linari, Maria Grazia Palmerini, Giuseppe Familiari, Cesare Aragona, and et al. 2023. "Effects of Simulated Microgravity In Vitro on Human Metaphase II Oocytes: An Electron Microscopy-Based Study" Cells 12, no. 10: 1346. https://doi.org/10.3390/cells12101346