
Synaptopodin Regulates Denervation-Induced Plasticity at Hippocampal Mossy Fiber Synapses
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Preparation of Tissue Cultures
2.3. Entorhinal Cortex Lesion (ECL)
2.4. Whole-Cell Patch-Clamp Recordings
2.5. Paired Whole-Cell Patch-Clamp Recordings of Hippocampal Mossy Fiber Synapses
2.6. Post Hoc Staining and Imaging
2.7. Serial Block-Face Scanning Electron Microscopy (SBF-SEM)
2.8. Transmission Electron Microscopy—Mossy Fiber Synapse Visualization
2.9. Synaptopodin Co-Immunoprecipitation (SP-coIP), RNA-Seq Analysis and Negative Contrast Electron Microscopy
2.10. Regional mRNA Library Preparations and Transcriptome Analysis
2.11. Experimental Design and Statistical Analysis
2.12. Data Availability
2.13. Digital Illustrations
3. Results
3.1. Entorhinal Cortex Lesion Leads to Excitatory Synaptic Strengthening in Both dGCs and CA3-PCs
3.2. Partial Denervation Does Not Cause Changes in Intrinsic Cellular Properties
3.3. Entorhinal Cortex Lesion Induces Ultrastructural Changes in Dendritic Spines
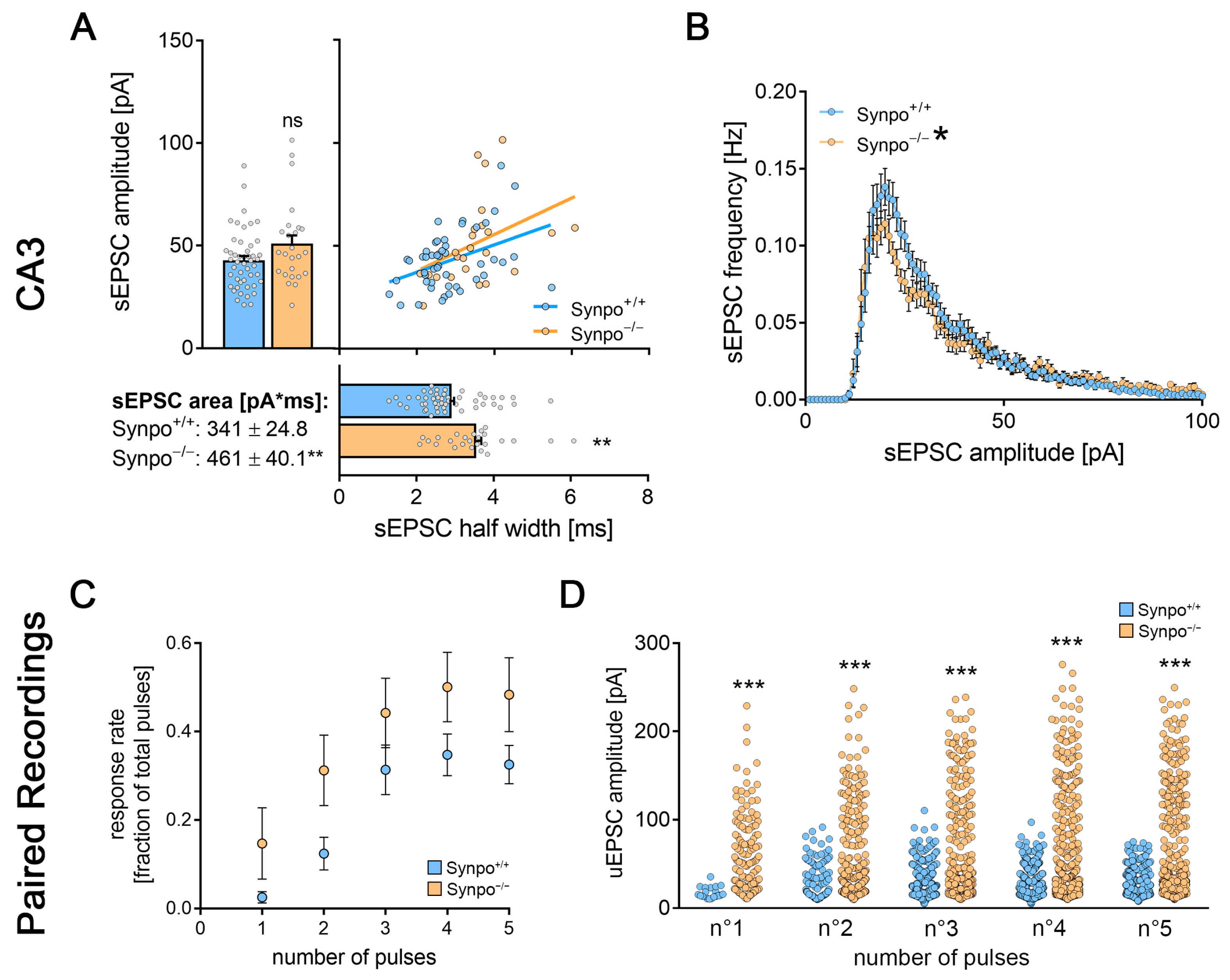
3.4. Entorhinal Cortex Lesion Induces Changes in Synaptic Transmission at Hippocampal Mossy Fiber Synapses
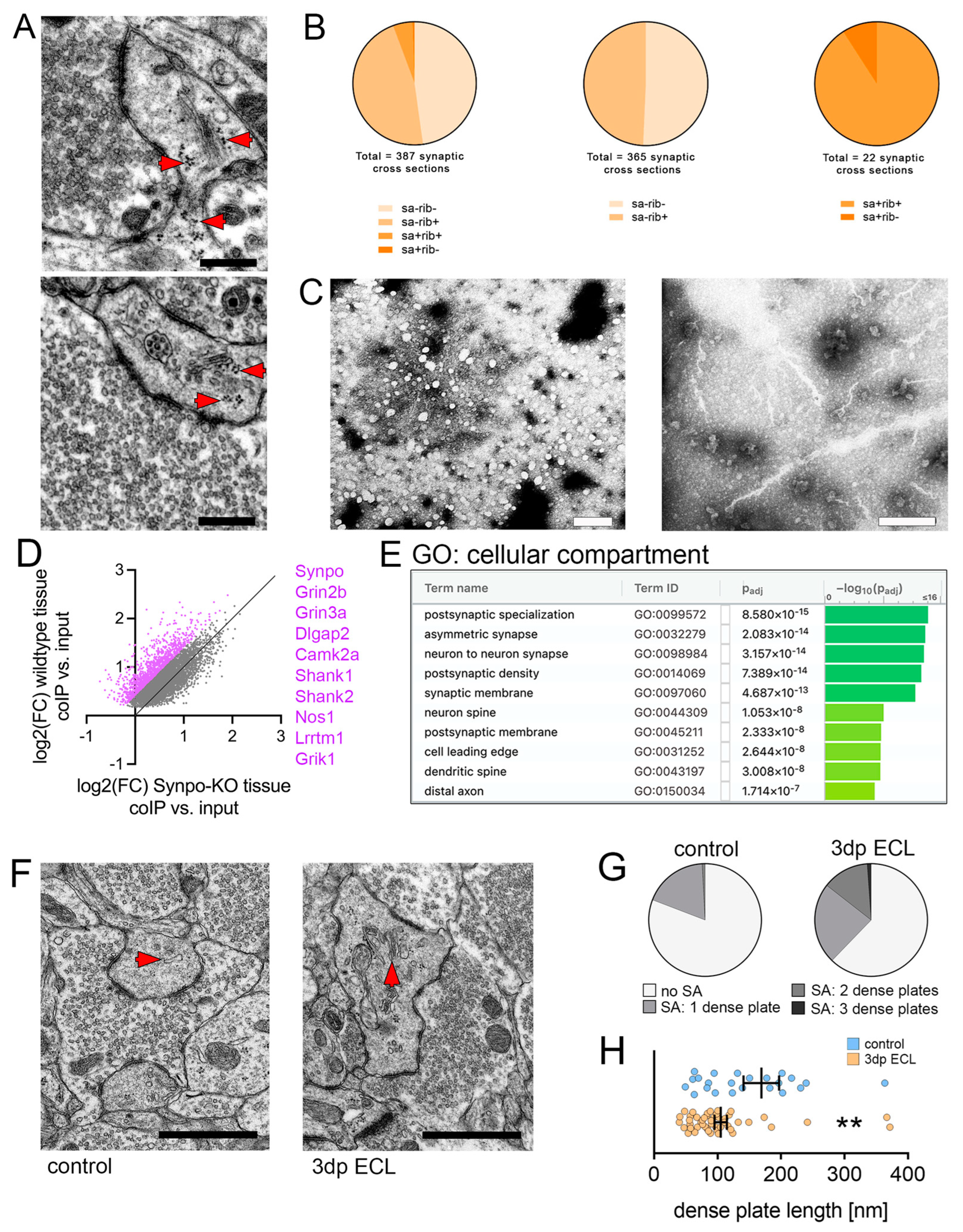
3.5. Synaptopodin and the Spine Apparatus Organelle Are Associated with Dendritic Ribosomes and Synapse-Related mRNAs
3.6. Ultrastructural Reorganization of the Spine Apparatus Organelle
3.7. Synaptopodin-Deficient Tissue Cultures Show Deficits in Lesion-Induced Synaptic Plasticity upon Partial Denervation
3.8. Entorhinal Cortex Lesion Causes Synaptopodin-Dependent Changes in the Region-Specific Hippocampal Transcriptome
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mesulam, M.M.; Lalehzari, N.; Rahmani, F.; Ohm, D.; Shahidehpour, R.; Kim, G.; Gefen, T.; Weintraub, S.; Bigio, E.; Geula, C. Cortical cholinergic denervation in primary progressive aphasia with Alzheimer pathology. Neurology 2019, 92, e1580–e1588. [Google Scholar] [CrossRef] [PubMed]
- Turrigiano, G.G.; Leslie, K.R.; Desai, N.S.; Rutherford, L.C.; Nelson, S.B. Activity-dependent scaling of quantal amplitude in neocortical neurons. Nature 1998, 391, 892–896. [Google Scholar] [CrossRef] [PubMed]
- Riegle, K.C.; Meyer, R.L. Rapid homeostatic plasticity in the intact adult visual system. J. Neurosci. 2007, 27, 10556–10567. [Google Scholar] [CrossRef] [PubMed]
- Weng, F.J.; Garcia, R.I.; Lutzu, S.; Alvina, K.; Zhang, Y.; Dushko, M.; Ku, T.; Zemoura, K.; Rich, D.; Garcia-Dominguez, D.; et al. Npas4 Is a Critical Regulator of Learning-Induced Plasticity at Mossy Fiber-CA3 Synapses during Contextual Memory Formation. Neuron 2018, 97, 1137–1152.e5. [Google Scholar] [CrossRef]
- Chicurel, M.E.; Harris, K.M. Three-dimensional analysis of the structure and composition of CA3 branched dendritic spines and their synaptic relationships with mossy fiber boutons in the rat hippocampus. J. Comp. Neurol. 1992, 325, 169–182. [Google Scholar] [CrossRef]
- Jedlicka, P.; Schwarzacher, S.W.; Winkels, R.; Kienzler, F.; Frotscher, M.; Bramham, C.R.; Schultz, C.; Bas Orth, C.; Deller, T. Impairment of in vivo theta-burst long-term potentiation and network excitability in the dentate gyrus of synaptopodin-deficient mice lacking the spine apparatus and the cisternal organelle. Hippocampus 2009, 19, 130–140. [Google Scholar] [CrossRef]
- Speranza, L.; Inglebert, Y.; De Sanctis, C.; Wu, P.Y.; Kalinowska, M.; McKinney, R.A.; Francesconi, A. Stabilization of Spine Synaptopodin by mGluR1 Is Required for mGluR-LTD. J. Neurosci. 2022, 42, 1666–1678. [Google Scholar] [CrossRef]
- Vlachos, A.; Ikenberg, B.; Lenz, M.; Becker, D.; Reifenberg, K.; Bas-Orth, C.; Deller, T. Synaptopodin regulates denervation-induced homeostatic synaptic plasticity. Proc. Natl. Acad. Sci. USA 2013, 110, 8242–8247. [Google Scholar] [CrossRef]
- Hu, H.T.; Lin, Y.J.; Wang, U.T.; Lee, S.P.; Liou, Y.H.; Chen, B.C.; Hsueh, Y.P. Autism-related KLHL17 and SYNPO act in concert to control activity-dependent dendritic spine enlargement and the spine apparatus. PLoS Biol. 2023, 21, e3002274. [Google Scholar] [CrossRef]
- Wingo, A.P.; Dammer, E.B.; Breen, M.S.; Logsdon, B.A.; Duong, D.M.; Troncosco, J.C.; Thambisetty, M.; Beach, T.G.; Serrano, G.E.; Reiman, E.M.; et al. Large-scale proteomic analysis of human brain identifies proteins associated with cognitive trajectory in advanced age. Nat. Commun. 2019, 10, 1619. [Google Scholar] [CrossRef]
- Lee, K.J.; Queenan, B.N.; Rozeboom, A.M.; Bellmore, R.; Lim, S.T.; Vicini, S.; Pak, D.T. Mossy fiber-CA3 synapses mediate homeostatic plasticity in mature hippocampal neurons. Neuron 2013, 77, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Maus, L.; Lee, C.; Altas, B.; Sertel, S.M.; Weyand, K.; Rizzoli, S.O.; Rhee, J.; Brose, N.; Imig, C.; Cooper, B.H. Ultrastructural Correlates of Presynaptic Functional Heterogeneity in Hippocampal Synapses. Cell Rep. 2020, 30, 3632–3643.e8. [Google Scholar] [CrossRef] [PubMed]
- Lenz, M.; Galanis, C.; Kleidonas, D.; Fellenz, M.; Deller, T.; Vlachos, A. Denervated mouse dentate granule cells adjust their excitatory but not inhibitory synapses following in vitro entorhinal cortex lesion. Exp. Neurol. 2019, 312, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, A.; Bas Orth, C.; Schneider, G.; Deller, T. Time-lapse imaging of granule cells in mouse entorhino-hippocampal slice cultures reveals changes in spine stability after entorhinal denervation. J. Comp. Neurol. 2012, 520, 1891–1902. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, A.; Becker, D.; Jedlicka, P.; Winkels, R.; Roeper, J.; Deller, T. Entorhinal denervation induces homeostatic synaptic scaling of excitatory postsynapses of dentate granule cells in mouse organotypic slice cultures. PLoS ONE 2012, 7, e32883. [Google Scholar] [CrossRef] [PubMed]
- Prang, P.; Del Turco, D.; Deller, T. Associational sprouting in the mouse fascia dentata after entorhinal lesion in vitro. Brain Res. 2003, 978, 205–212. [Google Scholar] [CrossRef]
- Jedlicka, P.; Deller, T. Understanding the role of synaptopodin and the spine apparatus in Hebbian synaptic plasticity—New perspectives and the need for computational modeling. Neurobiol. Learn. Mem. 2017, 138, 21–30. [Google Scholar] [CrossRef]
- Lenz, M.; Eichler, A.; Kruse, P.; Muellerleile, J.; Deller, T.; Jedlicka, P.; Vlachos, A. All-trans retinoic acid induces synaptopodin-dependent metaplasticity in mouse dentate granule cells. Elife 2021, 10, e71983. [Google Scholar] [CrossRef]
- Lenz, M.; Kruse, P.; Eichler, A.; Straehle, J.; Beck, J.; Deller, T.; Vlachos, A. All-trans retinoic acid induces synaptic plasticity in human cortical neurons. Elife 2021, 10, e63026. [Google Scholar] [CrossRef]
- Deller, T.; Korte, M.; Chabanis, S.; Drakew, A.; Schwegler, H.; Stefani, G.G.; Zuniga, A.; Schwarz, K.; Bonhoeffer, T.; Zeller, R.; et al. Synaptopodin-deficient mice lack a spine apparatus and show deficits in synaptic plasticity. Proc. Natl. Acad. Sci. USA 2003, 100, 10494–10499. [Google Scholar] [CrossRef]
- Korkotian, E.; Frotscher, M.; Segal, M. Synaptopodin regulates spine plasticity: Mediation by calcium stores. J. Neurosci. 2014, 34, 11641–11651. [Google Scholar] [CrossRef] [PubMed]
- Maggio, N.; Vlachos, A. Tumor necrosis factor (TNF) modulates synaptic plasticity in a concentration-dependent manner through intracellular calcium stores. J. Mol. Med. 2018, 96, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, A.; Korkotian, E.; Schonfeld, E.; Copanaki, E.; Deller, T.; Segal, M. Synaptopodin regulates plasticity of dendritic spines in hippocampal neurons. J. Neurosci. 2009, 29, 1017–1033. [Google Scholar] [CrossRef] [PubMed]
- Sutton, M.A.; Ito, H.T.; Cressy, P.; Kempf, C.; Woo, J.C.; Schuman, E.M. Miniature neurotransmission stabilizes synaptic function via tonic suppression of local dendritic protein synthesis. Cell 2006, 125, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Del Turco, D.; Deller, T. Organotypic entorhino-hippocampal slice cultures—A tool to study the molecular and cellular regulation of axonal regeneration and collateral sprouting in vitro. Methods Mol. Biol. 2007, 399, 55–66. [Google Scholar] [CrossRef]
- Vlachos, A.; Muller-Dahlhaus, F.; Rosskopp, J.; Lenz, M.; Ziemann, U.; Deller, T. Repetitive magnetic stimulation induces functional and structural plasticity of excitatory postsynapses in mouse organotypic hippocampal slice cultures. J. Neurosci. 2012, 32, 17514–17523. [Google Scholar] [CrossRef]
- Tani, H.; Dulla, C.G.; Farzampour, Z.; Taylor-Weiner, A.; Huguenard, J.R.; Reimer, R.J. A local glutamate-glutamine cycle sustains synaptic excitatory transmitter release. Neuron 2014, 81, 888–900. [Google Scholar] [CrossRef]
- Beike, L.; Wrede, C.; Hegermann, J.; Lopez-Rodriguez, E.; Kloth, C.; Gauldie, J.; Kolb, M.; Maus, U.A.; Ochs, M.; Knudsen, L. Surfactant dysfunction and alveolar collapse are linked with fibrotic septal wall remodeling in the TGF-beta1-induced mouse model of pulmonary fibrosis. Lab. Investig. 2019, 99, 830–852. [Google Scholar] [CrossRef]
- Henze, D.A.; McMahon, D.B.; Harris, K.M.; Barrionuevo, G. Giant miniature EPSCs at the hippocampal mossy fiber to CA3 pyramidal cell synapse are monoquantal. J. Neurophysiol. 2002, 87, 15–29. [Google Scholar] [CrossRef]
- Cardona, A.; Saalfeld, S.; Schindelin, J.; Arganda-Carreras, I.; Preibisch, S.; Longair, M.; Tomancak, P.; Hartenstein, V.; Douglas, R.J. TrakEM2 software for neural circuit reconstruction. PLoS ONE 2012, 7, e38011. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Gruning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed]
- Galaxy, C. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2022 update. Nucleic Acids Res. 2022, 50, W345–W351. [Google Scholar] [CrossRef]
- Goecks, J.; Nekrutenko, A.; Taylor, J.; Galaxy, T. Galaxy: A comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 2010, 11, R86. [Google Scholar] [CrossRef] [PubMed]
- Batut, B.; van den Beek, M.; Doyle, M.A.; Soranzo, N. RNA-Seq Data Analysis in Galaxy. Methods Mol. Biol. 2021, 2284, 367–392. [Google Scholar] [CrossRef]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]
- Strehl, A.; Galanis, C.; Radic, T.; Schwarzacher, S.W.; Deller, T.; Vlachos, A. Dopamine Modulates Homeostatic Excitatory Synaptic Plasticity of Immature Dentate Granule Cells in Entorhino-Hippocampal Slice Cultures. Front. Mol. Neurosci. 2018, 11, 303. [Google Scholar] [CrossRef]
- Lipp, H.P.; Schwegler, H.; Heimrich, B.; Driscoll, P. Infrapyramidal mossy fibers and two-way avoidance learning: Developmental modification of hippocampal circuitry and adult behavior of rats and mice. J. Neurosci. 1988, 8, 1905–1921. [Google Scholar] [CrossRef]
- Vandael, D.; Borges-Merjane, C.; Zhang, X.; Jonas, P. Short-Term Plasticity at Hippocampal Mossy Fiber Synapses Is Induced by Natural Activity Patterns and Associated with Vesicle Pool Engram Formation. Neuron 2020, 107, 509–521.e7. [Google Scholar] [CrossRef]
- Bissen, D.; Kracht, M.K.; Foss, F.; Hofmann, J.; Acker-Palmer, A. EphrinB2 and GRIP1 stabilize mushroom spines during denervation-induced homeostatic plasticity. Cell Rep. 2021, 34, 108923. [Google Scholar] [CrossRef]
- Platschek, S.; Cuntz, H.; Vuksic, M.; Deller, T.; Jedlicka, P. A general homeostatic principle following lesion induced dendritic remodeling. Acta Neuropathol. Commun. 2016, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Barnes, S.J.; Franzoni, E.; Jacobsen, R.I.; Erdelyi, F.; Szabo, G.; Clopath, C.; Keller, G.B.; Keck, T. Deprivation-Induced Homeostatic Spine Scaling In Vivo Is Localized to Dendritic Branches that Have Undergone Recent Spine Loss. Neuron 2017, 96, 871–882.e5. [Google Scholar] [CrossRef] [PubMed]
- Hengen, K.B.; Lambo, M.E.; Van Hooser, S.D.; Katz, D.B.; Turrigiano, G.G. Firing rate homeostasis in visual cortex of freely behaving rodents. Neuron 2013, 80, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Lambo, M.E.; Turrigiano, G.G. Synaptic and intrinsic homeostatic mechanisms cooperate to increase L2/3 pyramidal neuron excitability during a late phase of critical period plasticity. J. Neurosci. 2013, 33, 8810–8819. [Google Scholar] [CrossRef] [PubMed]
- Falahati, H.; Wu, Y.; Feuerer, V.; Simon, H.G.; De Camilli, P. Proximity proteomics of synaptopodin provides insight into the molecular composition of the spine apparatus of dendritic spines. Proc. Natl. Acad. Sci. USA 2022, 119, e2203750119. [Google Scholar] [CrossRef]
- Haber, M.; Zhou, L.; Murai, K.K. Cooperative astrocyte and dendritic spine dynamics at hippocampal excitatory synapses. J. Neurosci. 2006, 26, 8881–8891. [Google Scholar] [CrossRef]
- Heller, J.P.; Rusakov, D.A. The Nanoworld of the Tripartite Synapse: Insights from Super-Resolution Microscopy. Front. Cell. Neurosci. 2017, 11, 374. [Google Scholar] [CrossRef]
- Frotscher, M.; Heimrich, B. Lamina-specific synaptic connections of hippocampal neurons in vitro. J. Neurobiol. 1995, 26, 350–359. [Google Scholar] [CrossRef]
- Frotscher, M.; Heimrich, B.; Deller, T. Sprouting in the hippocampus is layer-specific. Trends Neurosci. 1997, 20, 218–223. [Google Scholar] [CrossRef]
- Frotscher, M.; Heimrich, B.; Deller, T.; Nitsch, R. Understanding the cortex through the hippocampus: Lamina-specific connections of the rat hippocampal neurons. J. Anat. 1995, 187 Pt 3, 539–545. [Google Scholar]
- Lenz, M.; Eichler, A.; Kruse, P.; Galanis, C.; Kleidonas, D.; Andrieux, G.; Boerries, M.; Jedlicka, P.; Muller, U.; Deller, T.; et al. The Amyloid Precursor Protein Regulates Synaptic Transmission at Medial Perforant Path Synapses. J. Neurosci. 2023, 43, 5290–5304. [Google Scholar] [CrossRef] [PubMed]
- Gulfo, M.C.; Lebowitz, J.J.; Ramos, C.; Hwang, D.W.; Nasrallah, K.; Castillo, P.E. Dopamine D2 receptors in hilar mossy cells regulate excitatory transmission and hippocampal function. Proc. Natl. Acad. Sci. USA 2023, 120, e2307509120. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.A.; Albrecht, A.; Caliskan, G.; Muller, B.; Demiray, Y.E.; Ludewig, S.; Meis, S.; Faber, N.; Hartig, R.; Schraven, B.; et al. HIPP neurons in the dentate gyrus mediate the cholinergic modulation of background context memory salience. Nat. Commun. 2017, 8, 189. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, R.E.; Rawson, J.M.; Eaton, B.A. An age-dependent change in the set point of synaptic homeostasis. J. Neurosci. 2014, 34, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Acsady, L.; Kamondi, A.; Sik, A.; Freund, T.; Buzsaki, G. GABAergic cells are the major postsynaptic targets of mossy fibers in the rat hippocampus. J. Neurosci. 1998, 18, 3386–3403. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.L.; Molina-Porcel, L.; Corrada, M.M.; Raible, K.; Lee, E.B.; Lee, V.M.; Kawas, C.H.; Trojanowski, J.Q. Perforant path synaptic loss correlates with cognitive impairment and Alzheimer’s disease in the oldest-old. Brain 2014, 137, 2578–2587. [Google Scholar] [CrossRef]
- Thal, D.R.; Holzer, M.; Rub, U.; Waldmann, G.; Gunzel, S.; Zedlick, D.; Schober, R. Alzheimer-related tau-pathology in the perforant path target zone and in the hippocampal stratum oriens and radiatum correlates with onset and degree of dementia. Exp. Neurol. 2000, 163, 98–110. [Google Scholar] [CrossRef]
- Hunsaker, M.R.; Rosenberg, J.S.; Kesner, R.P. The role of the dentate gyrus, CA3a,b, and CA3c for detecting spatial and environmental novelty. Hippocampus 2008, 18, 1064–1073. [Google Scholar] [CrossRef]
- Marr, D. Simple memory: A theory for archicortex. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1971, 262, 23–81. [Google Scholar] [CrossRef]
- Viana da Silva, S.; Zhang, P.; Haberl, M.G.; Labrousse, V.; Grosjean, N.; Blanchet, C.; Frick, A.; Mulle, C. Hippocampal Mossy Fibers Synapses in CA3 Pyramidal Cells Are Altered at an Early Stage in a Mouse Model of Alzheimer’s Disease. J. Neurosci. 2019, 39, 4193–4205. [Google Scholar] [CrossRef]
- Villanueva-Castillo, C.; Tecuatl, C.; Herrera-Lopez, G.; Galvan, E.J. Aging-related impairments of hippocampal mossy fibers synapses on CA3 pyramidal cells. Neurobiol. Aging 2017, 49, 119–137. [Google Scholar] [CrossRef] [PubMed]
- De Bruyckere, E.; Simon, R.; Nestel, S.; Heimrich, B.; Katzel, D.; Egorov, A.V.; Liu, P.; Jenkins, N.A.; Copeland, N.G.; Schwegler, H.; et al. Stability and Function of Hippocampal Mossy Fiber Synapses Depend on Bcl11b/Ctip2. Front. Mol. Neurosci. 2018, 11, 103. [Google Scholar] [CrossRef] [PubMed]
- Vandael, D.; Okamoto, Y.; Jonas, P. Transsynaptic modulation of presynaptic short-term plasticity in hippocampal mossy fiber synapses. Nat. Commun. 2021, 12, 2912. [Google Scholar] [CrossRef] [PubMed]
- Vyleta, N.P.; Borges-Merjane, C.; Jonas, P. Plasticity-dependent, full detonation at hippocampal mossy fiber-CA3 pyramidal neuron synapses. Elife 2016, 5, e17977. [Google Scholar] [CrossRef] [PubMed]
- Vyleta, N.P.; Jonas, P. Loose coupling between Ca2+ channels and release sensors at a plastic hippocampal synapse. Science 2014, 343, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Henze, D.A.; Wittner, L.; Buzsaki, G. Single granule cells reliably discharge targets in the hippocampal CA3 network in vivo. Nat. Neurosci. 2002, 5, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Vandael, D.; Okamoto, Y.; Borges-Merjane, C.; Vargas-Barroso, V.; Suter, B.A.; Jonas, P. Subcellular patch-clamp techniques for single-bouton stimulation and simultaneous pre- and postsynaptic recording at cortical synapses. Nat. Protoc. 2021, 16, 2947–2967. [Google Scholar] [CrossRef]
- Gundlfinger, A.; Leibold, C.; Gebert, K.; Moisel, M.; Schmitz, D.; Kempter, R. Differential modulation of short-term synaptic dynamics by long-term potentiation at mouse hippocampal mossy fibre synapses. J. Physiol. 2007, 585, 853–865. [Google Scholar] [CrossRef]
- Zalutsky, R.A.; Nicoll, R.A. Comparison of two forms of long-term potentiation in single hippocampal neurons. Science 1990, 248, 1619–1624. [Google Scholar] [CrossRef]
- Leutgeb, J.K.; Leutgeb, S.; Moser, M.B.; Moser, E.I. Pattern separation in the dentate gyrus and CA3 of the hippocampus. Science 2007, 315, 961–966. [Google Scholar] [CrossRef]
- O’Reilly, R.C.; McClelland, J.L. Hippocampal conjunctive encoding, storage, and recall: Avoiding a trade-off. Hippocampus 1994, 4, 661–682. [Google Scholar] [CrossRef] [PubMed]
- Prince, L.Y.; Bacon, T.J.; Tigaret, C.M.; Mellor, J.R. Neuromodulation of the Feedforward Dentate Gyrus-CA3 Microcircuit. Front. Synaptic Neurosci. 2016, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Monday, H.R.; Kharod, S.C.; Yoon, Y.J.; Singer, R.H.; Castillo, P.E. Presynaptic FMRP and local protein synthesis support structural and functional plasticity of glutamatergic axon terminals. Neuron 2022, 110, 2588–2606.e6. [Google Scholar] [CrossRef] [PubMed]
- Orlando, M.; Dvorzhak, A.; Bruentgens, F.; Maglione, M.; Rost, B.R.; Sigrist, S.J.; Breustedt, J.; Schmitz, D. Recruitment of release sites underlies chemical presynaptic potentiation at hippocampal mossy fiber boutons. PLoS Biol. 2021, 19, e3001149. [Google Scholar] [CrossRef]
- Jedlicka, P.; Vlachos, A.; Schwarzacher, S.W.; Deller, T. A role for the spine apparatus in LTP and spatial learning. Behav. Brain Res. 2008, 192, 12–19. [Google Scholar] [CrossRef]
- Czarnecki, K.; Haas, C.A.; Bas Orth, C.; Deller, T.; Frotscher, M. Postnatal development of synaptopodin expression in the rodent hippocampus. J. Comp. Neurol. 2005, 490, 133–144. [Google Scholar] [CrossRef]
- Dubes, S.; Soula, A.; Benquet, S.; Tessier, B.; Poujol, C.; Favereaux, A.; Thoumine, O.; Letellier, M. miR-124-dependent tagging of synapses by synaptopodin enables input-specific homeostatic plasticity. EMBO J. 2022, 41, e109012. [Google Scholar] [CrossRef]
- Yap, K.; Drakew, A.; Smilovic, D.; Rietsche, M.; Paul, M.H.; Vuksic, M.; Del Turco, D.; Deller, T. The actin-modulating protein synaptopodin mediates long-term survival of dendritic spines. Elife 2020, 9, e62944. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | 2hp ECL | 3dp ECL | |
|---|---|---|---|
| DG proximal | 0.1 [0.07–0.2] μm2 (ncontrol = 183) | 0.1 [0.08–0.1] μm2 (n2hpl = 297) ns | 0.09 [0.06–0.1] μm2 (n3dpl = 231) ns |
| DG distal | 0.2 [0.0975–0.3] μm2 (ncontrol = 62) | 0.1 [0.09–0.2] μm2 (n2hpl = 89) * | 0.09 [0.06–0.1] μm2 (n3dpl = 48) *** |
| CA3 proximal | 0.1 [0.07–0.2] μm2 (ncontrol = 212) | 0.1 [0.0685–0.2] μm2 (n2hpl = 229) ns | 0.1 [0.06–0.3] μm2 (n3dpl = 224) * |
| CA3 distal | 0.1 [0.08–0.2] μm2 (ncontrol = 100) | 0.09 [0.07–0.1] μm2 (n2hpl = 121) ns | 0.08 [0.06–0.1] μm2 (n3dpl = 120) *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kruse, P.; Brandes, G.; Hemeling, H.; Huang, Z.; Wrede, C.; Hegermann, J.; Vlachos, A.; Lenz, M. Synaptopodin Regulates Denervation-Induced Plasticity at Hippocampal Mossy Fiber Synapses. Cells 2024, 13, 114. https://doi.org/10.3390/cells13020114
Kruse P, Brandes G, Hemeling H, Huang Z, Wrede C, Hegermann J, Vlachos A, Lenz M. Synaptopodin Regulates Denervation-Induced Plasticity at Hippocampal Mossy Fiber Synapses. Cells. 2024; 13(2):114. https://doi.org/10.3390/cells13020114
Chicago/Turabian StyleKruse, Pia, Gudrun Brandes, Hanna Hemeling, Zhong Huang, Christoph Wrede, Jan Hegermann, Andreas Vlachos, and Maximilian Lenz. 2024. "Synaptopodin Regulates Denervation-Induced Plasticity at Hippocampal Mossy Fiber Synapses" Cells 13, no. 2: 114. https://doi.org/10.3390/cells13020114