
The Sensitization of Triple-Negative Breast Cancers to Poly ADP Ribose Polymerase Inhibition Independent of BRCA1/2 Mutation Status by Chemically Modified microRNA-489
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Reagents
2.2. Cell Viability Assay and Drug Synergy Assay
2.3. Immunofluorescence Staining
2.4. Western Blot Analysis
2.5. RNA-Seq Analysis
2.6. Organoid Culture and Drug Treatment
2.7. siRNA Transfection
2.8. Quantitative Real-Time PCR (qRT-PCR) of NEK8
2.9. Soft Agar Colony Assay
3. Results
3.1. CMM489 Synergizes with Olaparib as a Combination Treatment Partner in TNBC Cells Regardless of the BRCA Mutation Status
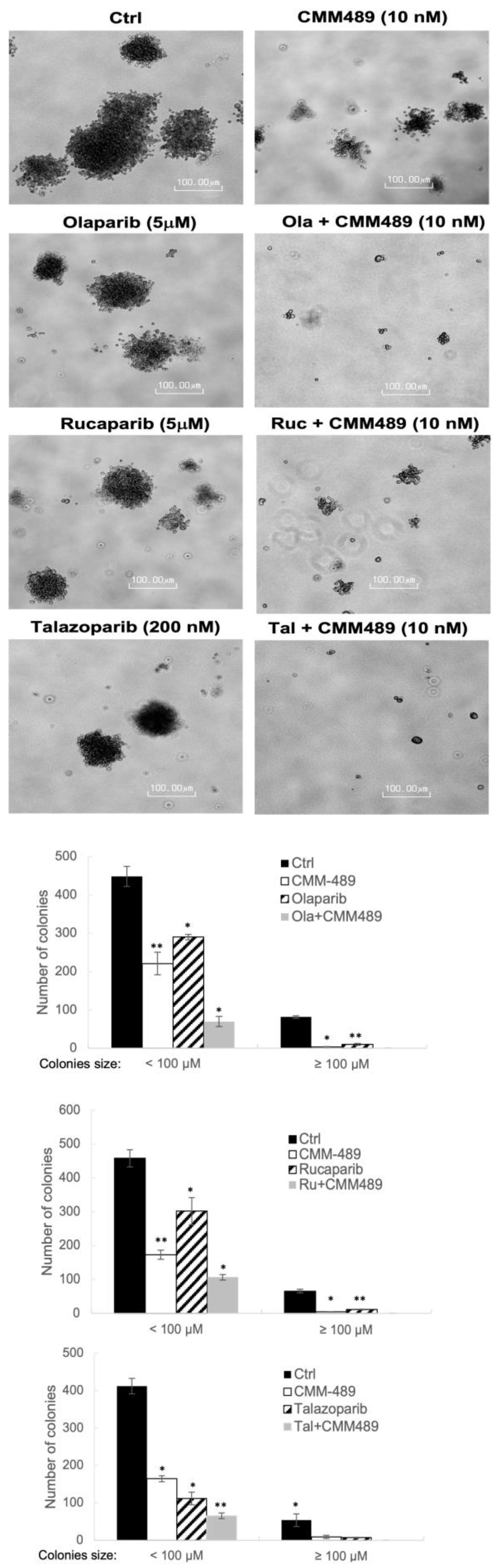
3.2. CMM489 Synergizes with Multiple FDA-Approved PARP Inhibitors in Colony-Forming Assays
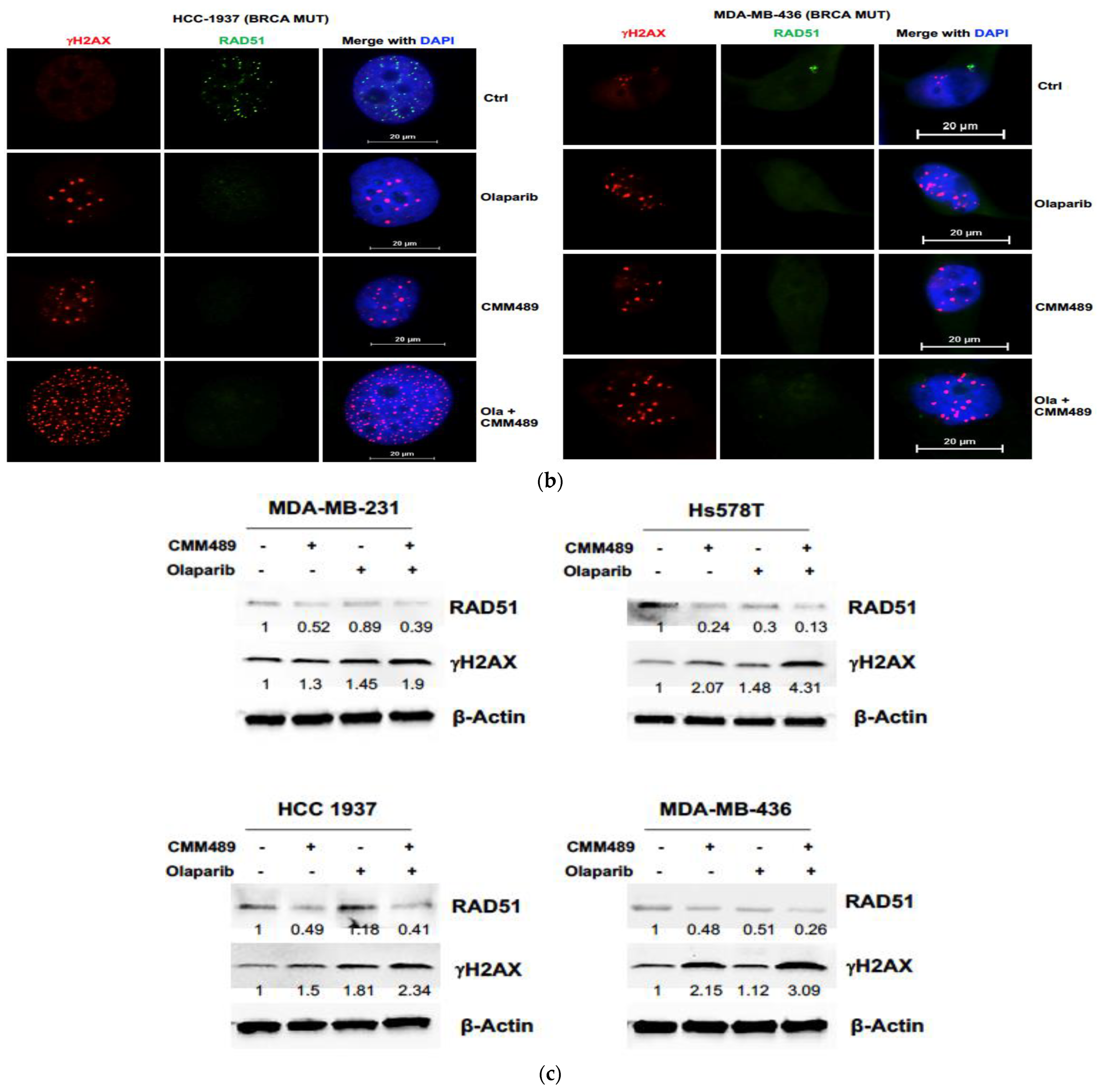
3.3. Homologous Recombination Deficiency (HRD) Is the Key Mechanism of Synergy between CMM489 and Olaparib
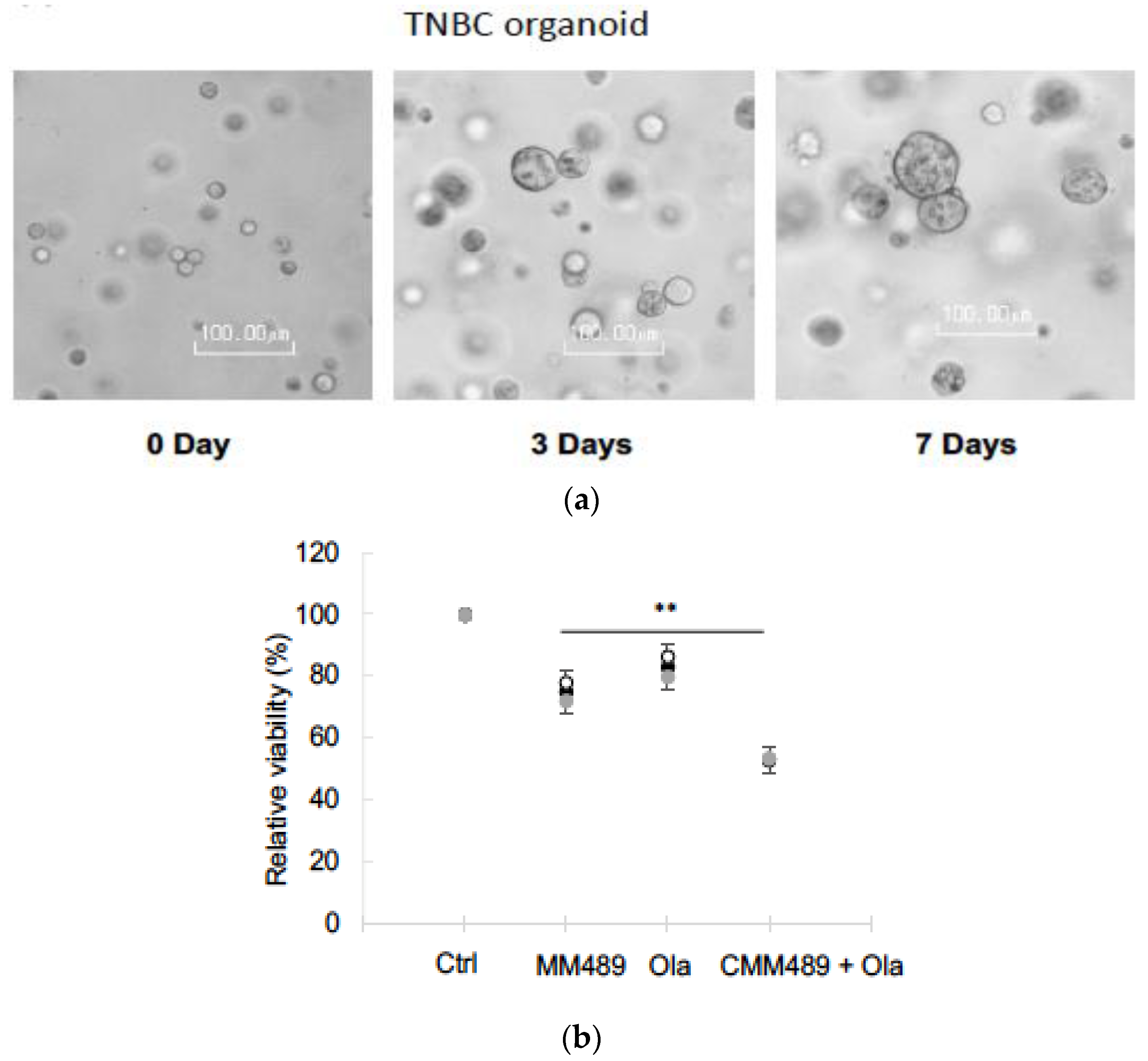
3.4. Synergistic Effects of CMM489 and Olaparib Were Observed in TNBC Organoid Models
3.5. RNA-Seq Analysis Identifies NEK8 as a Potential CMM489 Target Gene That Mediates Homologous Recombination
3.6. Knockdown of NEK8 Expression by siRNA Mimics the Action of CMM489
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mayer, I.A.; Abramson, V.G.; Lehmann, B.D.; Pietenpol, J.A. New Strategies for Triple-Negative Breast Cancer—Deciphering the Heterogeneity. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, G.; Balko, J.M.; Mayer, I.A.; Sanders, M.E.; Gianni, L. Triple-Negative Breast Cancer: Challenges and Opportunities of a Heterogeneous Disease. Nat. Rev. Clin. Oncol. 2016, 13, 674–690. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Xiang, L.; Li, T.; Bai, Z. Cancer Hallmarks, Biomarkers and Breast Cancer Molecular Subtypes. J. Cancer 2016, 7, 1281–1294. [Google Scholar] [CrossRef] [PubMed]
- Abramson, V.G.; Lehmann, B.D.; Ballinger, T.J.; Pietenpol, J.A. Subtyping of Triple-Negative Breast Cancer: Implications for Therapy. Cancer 2015, 121, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, I.S. Molecular Testing in Breast Cancer: A Guide to Current Practices. Arch. Pathol. Lab. Med. 2016, 140, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Collignon, J.; Lousberg, L.; Schroeder, H.; Jerusalem, G. Triple-Negative Breast Cancer: Treatment Challenges and Solutions. Breast Cancer Targets Ther. 2016, 8, 93–107. [Google Scholar] [CrossRef]
- Chartron, E.; Theillet, C.; Guiu, S.; Jacot, W. Targeting Homologous Repair Deficiency in Breast and Ovarian Cancers: Biological Pathways, Preclinical and Clinical Data. Crit. Rev. Oncol. Hematol. 2019, 133, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Brandsma, I.; Fleuren, E.D.G.; Williamson, C.T.; Lord, C.J. Directing the Use of DDR Kinase Inhibitors in Cancer Treatment. Expert Opin. Investig. Drugs 2017, 26, 1341–1355. [Google Scholar] [CrossRef]
- Norquist, B.; Wurz, K.A.; Pennil, C.C.; Garcia, R.; Gross, J.; Sakai, W.; Karlan, B.Y.; Taniguchi, T.; Swisher, E.M. Secondary Somatic Mutations Restoring BRCA1/2 Predict Chemotherapy Resistance in Hereditary Ovarian Carcinomas. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2011, 29, 3008–3015. [Google Scholar] [CrossRef]
- Lord, C.J.; Ashworth, A. Mechanisms of Resistance to Therapies Targeting BRCA-Mutant Cancers. Nat. Med. 2013, 19, 1381–1388. [Google Scholar] [CrossRef]
- Brown, J.S.; O’Carrigan, B.; Jackson, S.P.; Yap, T.A. Targeting DNA Repair in Cancer: Beyond PARP Inhibitors. Cancer Discov. 2017, 7, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.; Tutt, A.; Ashworth, A. Targeting the DNA Repair Defect of BRCA Tumours. Curr. Opin. Pharmacol. 2005, 5, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Bryant, H.E.; Schultz, N.; Thomas, H.D.; Parker, K.M.; Flower, D.; Lopez, E.; Kyle, S.; Meuth, M.; Curtin, N.J.; Helleday, T. Specific Killing of BRCA2-Deficient Tumours with Inhibitors of Poly(ADP-Ribose) Polymerase. Nature 2005, 434, 913–917. [Google Scholar] [CrossRef]
- Lord, C.J.; McDonald, S.; Swift, S.; Turner, N.C.; Ashworth, A. A High-Throughput RNA Interference Screen for DNA Repair Determinants of PARP Inhibitor Sensitivity. DNA Repair. 2008, 7, 2010–2019. [Google Scholar] [CrossRef] [PubMed]
- Audeh, M.W.; Carmichael, J.; Penson, R.T.; Friedlander, M.; Powell, B.; Bell-McGuinn, K.M.; Scott, C.; Weitzel, J.N.; Oaknin, A.; Loman, N.; et al. Oral Poly(ADP-Ribose) Polymerase Inhibitor Olaparib in Patients with BRCA1 or BRCA2 Mutations and Recurrent Ovarian Cancer: A Proof-of-Concept Trial. Lancet 2010, 376, 245–251. [Google Scholar] [CrossRef]
- Gelmon, K.A.; Tischkowitz, M.; Mackay, H.; Swenerton, K.; Robidoux, A.; Tonkin, K.; Hirte, H.; Huntsman, D.; Clemons, M.; Gilks, B.; et al. Olaparib in Patients with Recurrent High-Grade Serous or Poorly Differentiated Ovarian Carcinoma or Triple-Negative Breast Cancer: A Phase 2, Multicentre, Open-Label, Non-Randomised Study. Lancet Oncol. 2011, 12, 852–861. [Google Scholar] [CrossRef]
- Engel, C.; Rhiem, K.; Hahnen, E.; Loibl, S.; Weber, K.E.; Seiler, S.; Zachariae, S.; Hauke, J.; Wappenschmidt, B.; Waha, A.; et al. Prevalence of Pathogenic BRCA1/2 Germline Mutations among 802 Women with Unilateral Triple-Negative Breast Cancer without Family Cancer History. BMC Cancer 2018, 18, 265. [Google Scholar] [CrossRef]
- Tutt, A.; Robson, M.; Garber, J.E.; Domchek, S.M.; Audeh, M.W.; Weitzel, J.N.; Friedlander, M.; Arun, B.; Loman, N.; Schmutzler, R.K.; et al. Oral Poly(ADP-Ribose) Polymerase Inhibitor Olaparib in Patients with BRCA1 or BRCA2 Mutations and Advanced Breast Cancer: A Proof-of-Concept Trial. Lancet 2010, 376, 235–244. [Google Scholar] [CrossRef]
- Robson, M.; Goessl, C.; Domchek, S. Olaparib for Metastatic Germline BRCA-Mutated Breast Cancer. N. Engl. J. Med. 2017, 377, 1792–1793. [Google Scholar] [CrossRef]
- Rivero, J.D.; Kohn, E.C. PARP Inhibitors: The Cornerstone of DNA Repair-Targeted Therapies. Oncology 2017, 31, 265–273. [Google Scholar]
- Martin, L.P.; Hamilton, T.C.; Schilder, R.J. Platinum Resistance: The Role of DNA Repair Pathways. Clin. Cancer Res. 2008, 14, 1291–1295. [Google Scholar] [CrossRef] [PubMed]
- Belli, C.; Duso, B.A.; Ferraro, E.; Curigliano, G. Homologous Recombination Deficiency in Triple Negative Breast Cancer. Breast 2019, 45, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Hunia, J.; Gawalski, K.; Szredzka, A.; Suskiewicz, M.J.; Nowis, D. The Potential of PARP Inhibitors in Targeted Cancer Therapy and Immunotherapy. Front. Mol. Biosci. 2022, 9, 1073797. [Google Scholar] [CrossRef] [PubMed]
- You, K.S.; Yi, Y.W.; Cho, J.; Park, J.-S.; Seong, Y.-S. Potentiating Therapeutic Effects of Epidermal Growth Factor Receptor Inhibition in Triple-Negative Breast Cancer. Pharmaceuticals 2021, 14, 589. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Keyomarsi, K. PARP Inhibitors as Single Agents and in Combination Therapy: The Most Promising Treatment Strategies in Clinical Trials for BRCA-Mutant Ovarian and Triple-Negative Breast Cancers. Expert Opin. Investig. Drugs 2022, 31, 607–631. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; McDonnell, K.; Choi, H.; Gao, D.; Hahn, M.; Joshi, N.; Park, S.-M.; Catena, R.; Do, Y.; Brazin, J.; et al. Suppression of miRNA-708 by Polycomb Group Promotes Metastases by Calcium-Induced Cell Migration. Cancer Cell 2013, 23, 63–76. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.W.; Xing, A.Y.; Xiang, S.; Shi, D.B.; Liu, L.; Li, Y.X.; Gao, P. Suppression of SPIN1-Mediated PI3K-Akt Pathway by miR-489 Increases Chemosensitivity in Breast Cancer. J. Pathol. 2016, 239, 459–472. [Google Scholar] [CrossRef]
- Jiang, L.; He, D.; Yang, D.; Chen, Z.; Pan, Q.; Mao, A.; Cai, Y.; Li, X.; Xing, H.; Shi, M.; et al. MiR-489 Regulates Chemoresistance in Breast Cancer via Epithelial Mesenchymal Transition Pathway. FEBS Lett. 2014, 588, 2009–2015. [Google Scholar] [CrossRef]
- Soung, Y.H.; Chung, H.; Yan, C.; Fesler, A.; Kim, H.; Oh, E.-S.; Ju, J.; Chung, J. Therapeutic Potential of Chemically Modified miR-489 in Triple-Negative Breast Cancers. Cancers 2020, 12, 2209. [Google Scholar] [CrossRef]
- Yi, Y.W.; Park, J.-S.; Kwak, S.-J.; Seong, Y.-S. Co-Treatment with BEZ235 Enhances Sensitivity of BRCA1-Negative Breast Cancer Cells to Olaparib. Anticancer Res. 2015, 35, 3829–3838. [Google Scholar]
- Keung, M.Y.; Wu, Y.; Badar, F.; Vadgama, J.V. Response of Breast Cancer Cells to PARP Inhibitors Is Independent of BRCA Status. J. Clin. Med. 2020, 9, 940. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Chen, L.; Huang, B.; Li, X.; Yang, L.; Hu, X.; Jiang, Y.; Shao, Z.; Wang, Z. Efficacy and Mechanism of the Combination of PARP and CDK4/6 Inhibitors in the Treatment of Triple-Negative Breast Cancer. J. Exp. Clin. Cancer Res. 2021, 40, 122. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.F.; Cruz, C.; Greifenberg, A.K.; Dust, S.; Stover, D.G.; Chi, D.; Primack, B.; Cao, S.; Bernhardy, A.J.; Coulson, R.; et al. CDK12 Inhibition Reverses De Novo and Acquired PARP Inhibitor Resistance in BRCA Wild-Type and Mutated Models of Triple-Negative Breast Cancer. Cell Rep. 2016, 17, 2367–2381. [Google Scholar] [CrossRef] [PubMed]
- Kondo, J.; Inoue, M. Application of Cancer Organoid Model for Drug Screening and Personalized Therapy. Cells 2019, 8, 470. [Google Scholar] [CrossRef]
- Liu, L.; Yu, L.; Li, Z.; Li, W.; Huang, W. Patient-Derived Organoid (PDO) Platforms to Facilitate Clinical Decision Making. J. Transl. Med. 2021, 19, 40–49. [Google Scholar] [CrossRef]
- Abeyta, A.; Castella, M.; Jacquemont, C.; Taniguchi, T. NEK8 Regulates DNA Damage-Induced RAD51 Foci Formation and Replication Fork Protection. Cell Cycle Georget. Tex 2017, 16, 335–347. [Google Scholar] [CrossRef]
- Spies, J.; Waizenegger, A.; Barton, O.; Sürder, M.; Wright, W.D.; Heyer, W.-D.; Löbrich, M. Nek1 Regulates Rad54 to Orchestrate Homologous Recombination and Replication Fork Stability. Mol. Cell 2016, 62, 903–917. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Subtype | BRCA Mutation Status | Olaparib Sensitivity |
|---|---|---|---|
| MDA-MB-231 | Mesenchymal Stem-like | Wild type | No |
| Hs578T | Mesenchymal Stem-like | Wild type | Yes |
| BT-549 | Mesenchymal | Wild type | No |
| MDA-MB-436 | Mesenchymal Stem-like | Mutant (5396 + 1G > A) | Yes |
| HCC-1937 | Basal-like | Mutant (5382insC) | No |
| Gene | log2 Fold Change | p-Value | Gene Description |
|---|---|---|---|
| TMEM63A | −1.112 | 1.4 × 10−12 | transmembrane protein 63A |
| ADM2 | −1.569 | 6.9 × 10−5 | adrenomedullin 2 |
| TNFSF18 | −2.008 | 8.5 × 10−5 | TNF superfamily member 18 |
| SYT2 | −1.774 | 1.6 × 10−4 | synaptotagmin 2 |
| YPEL1 | −4.198 | 4.3 × 10−2 | yippee-like 1 |
| DBH | −1.888 | 4.9 × 10−4 | dopamine beta-hydroxylase |
| NEK8 | −1.389 | 1.7 × 10−3 | NIMA related kinase 8 |
| LPAR2 | −1.376 | 3.1 × 10−3 | lysophosphatidic acid receptor 2 |
| NEDD9 | −1.625 | 6.7 × 10−3 | neural precursor cell expressed, developmentally downregulated 9 |
| CALCR | −1.719 | 7.1 × 10−3 | calcitonin receptor |
| ESPN | −2.026 | 7.5 × 10−3 | espin |
| TNS4 | −1.458 | 1.3 × 10−2 | tensin 4 |
| STAB2 | −1.912 | 1.3 × 10−2 | stabilin 2 |
| PIP5KL1 | −1.572 | 1.5 × 10−2 | phosphatidylinositol-4-phosphate 5-kinase like 1 |
| CCR7 | −2.177 | 1.9 × 10−2 | C-C motif chemokine receptor 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soung, Y.H.; Ju, J.; Chung, J. The Sensitization of Triple-Negative Breast Cancers to Poly ADP Ribose Polymerase Inhibition Independent of BRCA1/2 Mutation Status by Chemically Modified microRNA-489. Cells 2024, 13, 49. https://doi.org/10.3390/cells13010049
Soung YH, Ju J, Chung J. The Sensitization of Triple-Negative Breast Cancers to Poly ADP Ribose Polymerase Inhibition Independent of BRCA1/2 Mutation Status by Chemically Modified microRNA-489. Cells. 2024; 13(1):49. https://doi.org/10.3390/cells13010049
Chicago/Turabian StyleSoung, Young Hwa, Jingfang Ju, and Jun Chung. 2024. "The Sensitization of Triple-Negative Breast Cancers to Poly ADP Ribose Polymerase Inhibition Independent of BRCA1/2 Mutation Status by Chemically Modified microRNA-489" Cells 13, no. 1: 49. https://doi.org/10.3390/cells13010049