

Embryonic Ethanol but Not Cannabinoid Exposure Affects Zebrafish Cardiac Development via Agrin and Sonic Hedgehog Interaction


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Ethanol Treatment of Zebrafish Embryo
2.3. Cannabinoid Treatment of Zebrafish Embryos
2.4. Antisense Morpholino Injection
2.5. Midbrain–Hindbrain Boundary Analysis
2.6. Whole-Mount In Situ Hybridization
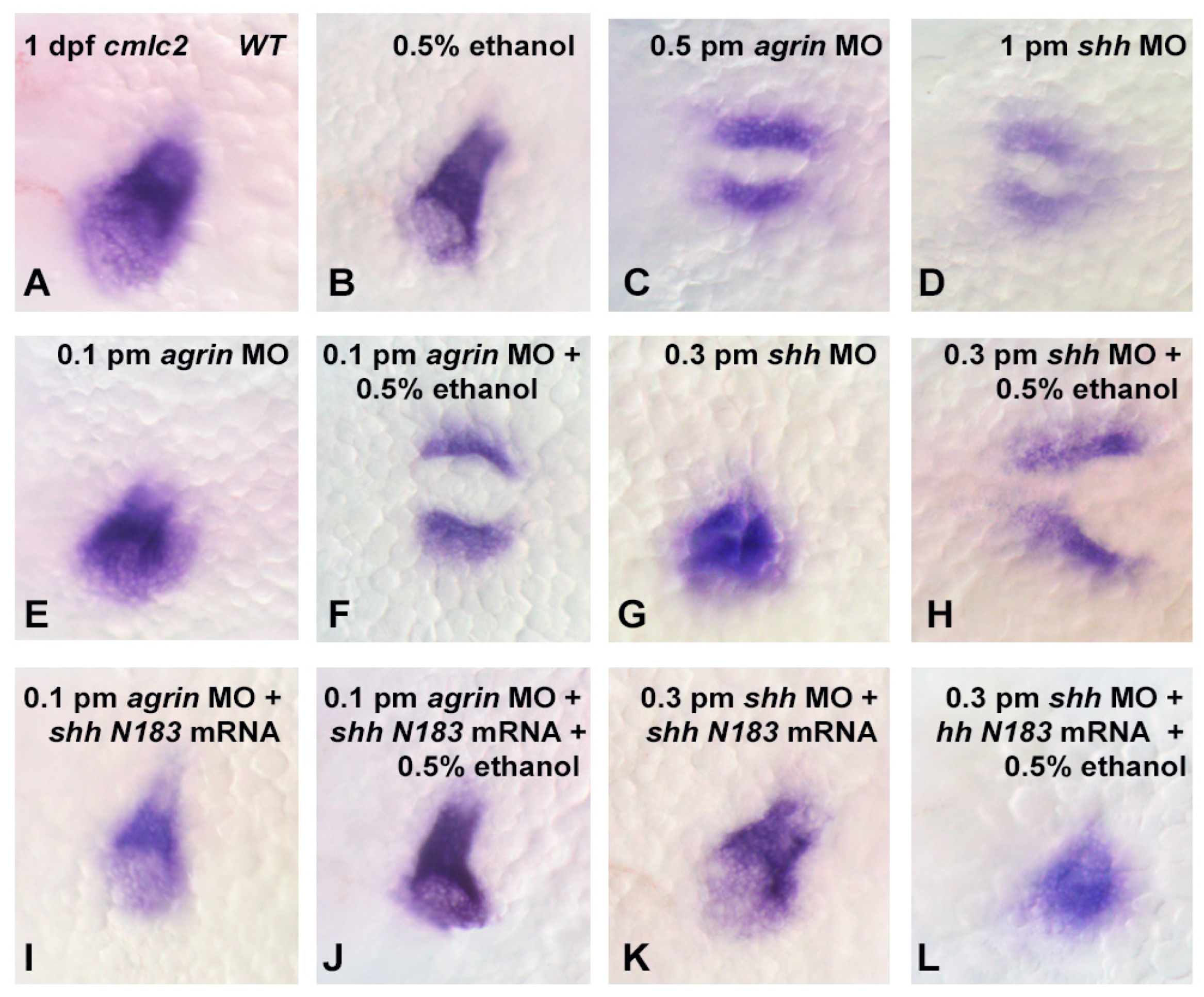
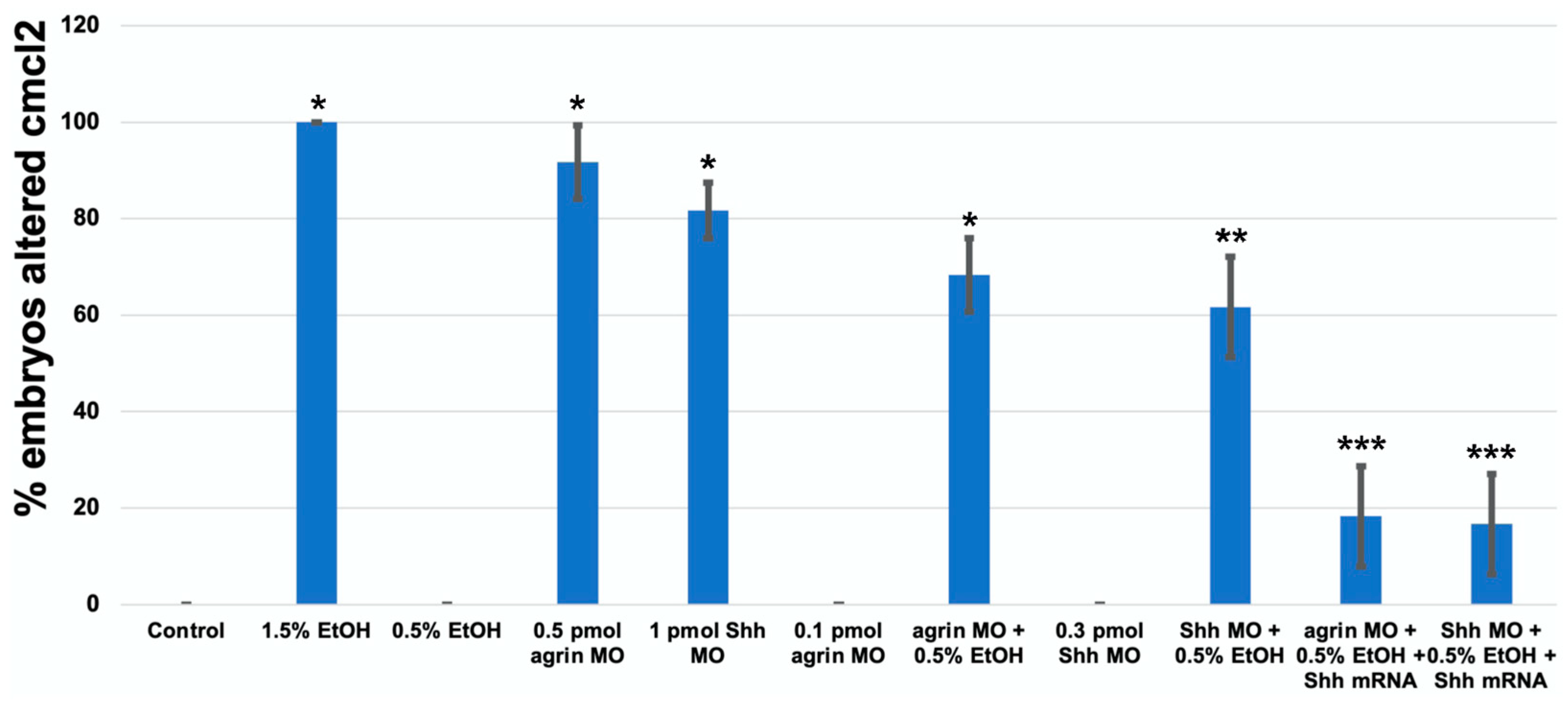
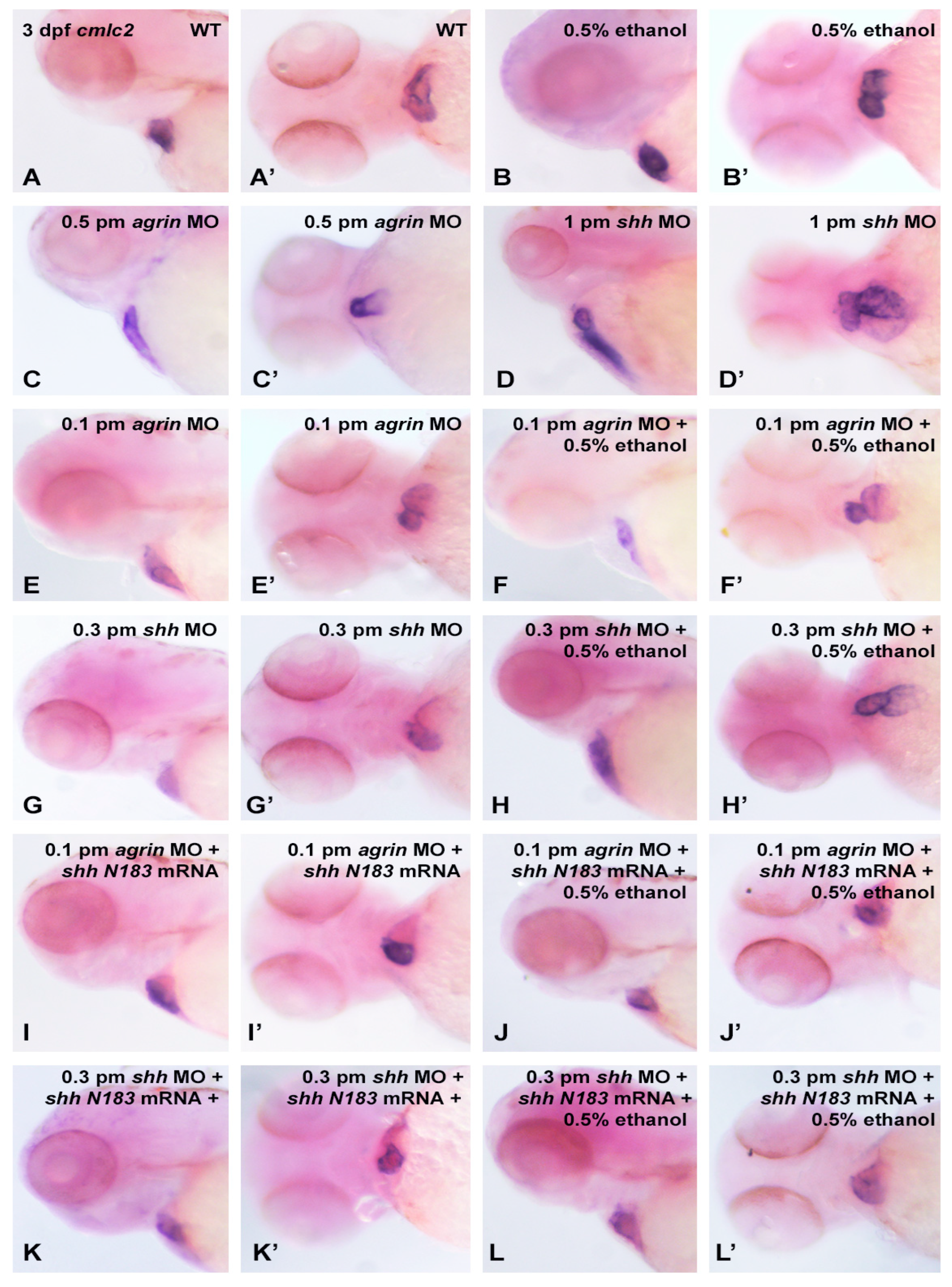
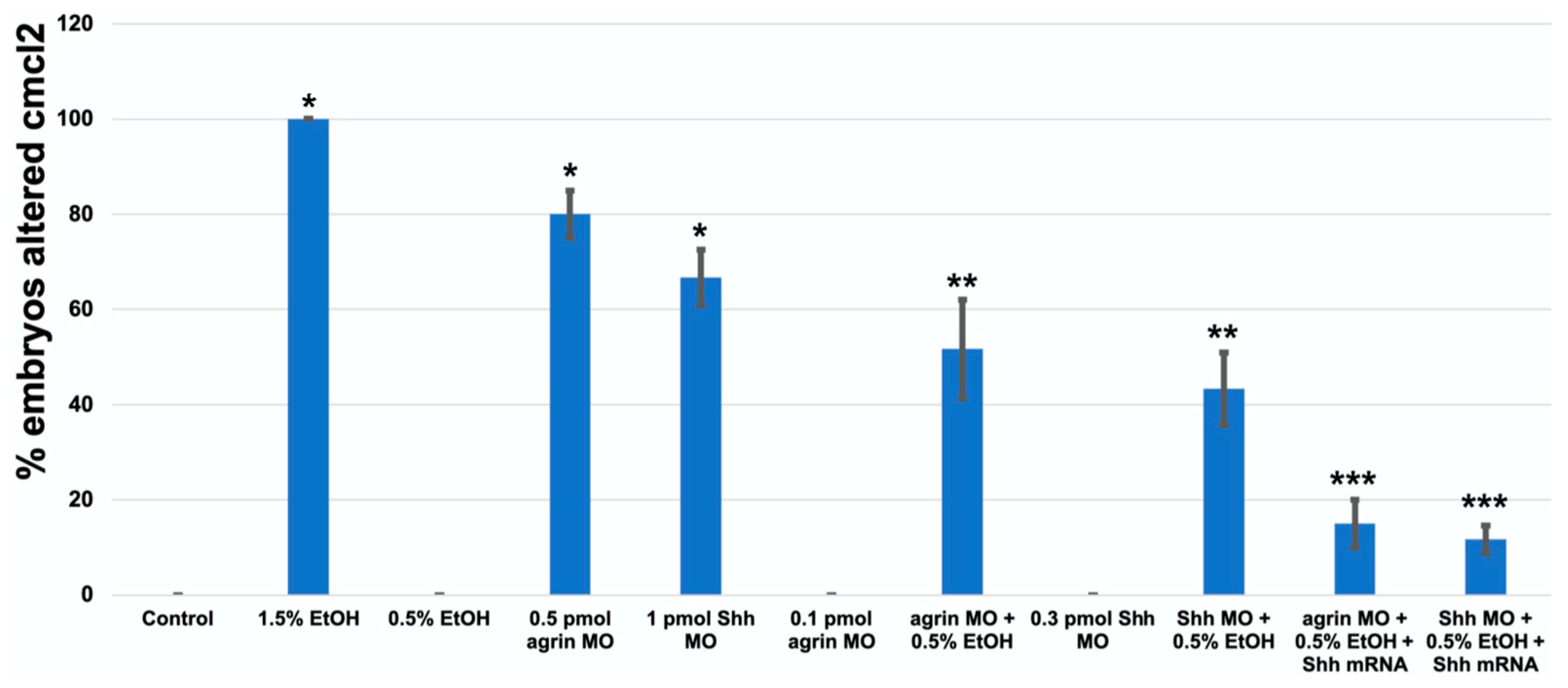
3. Results
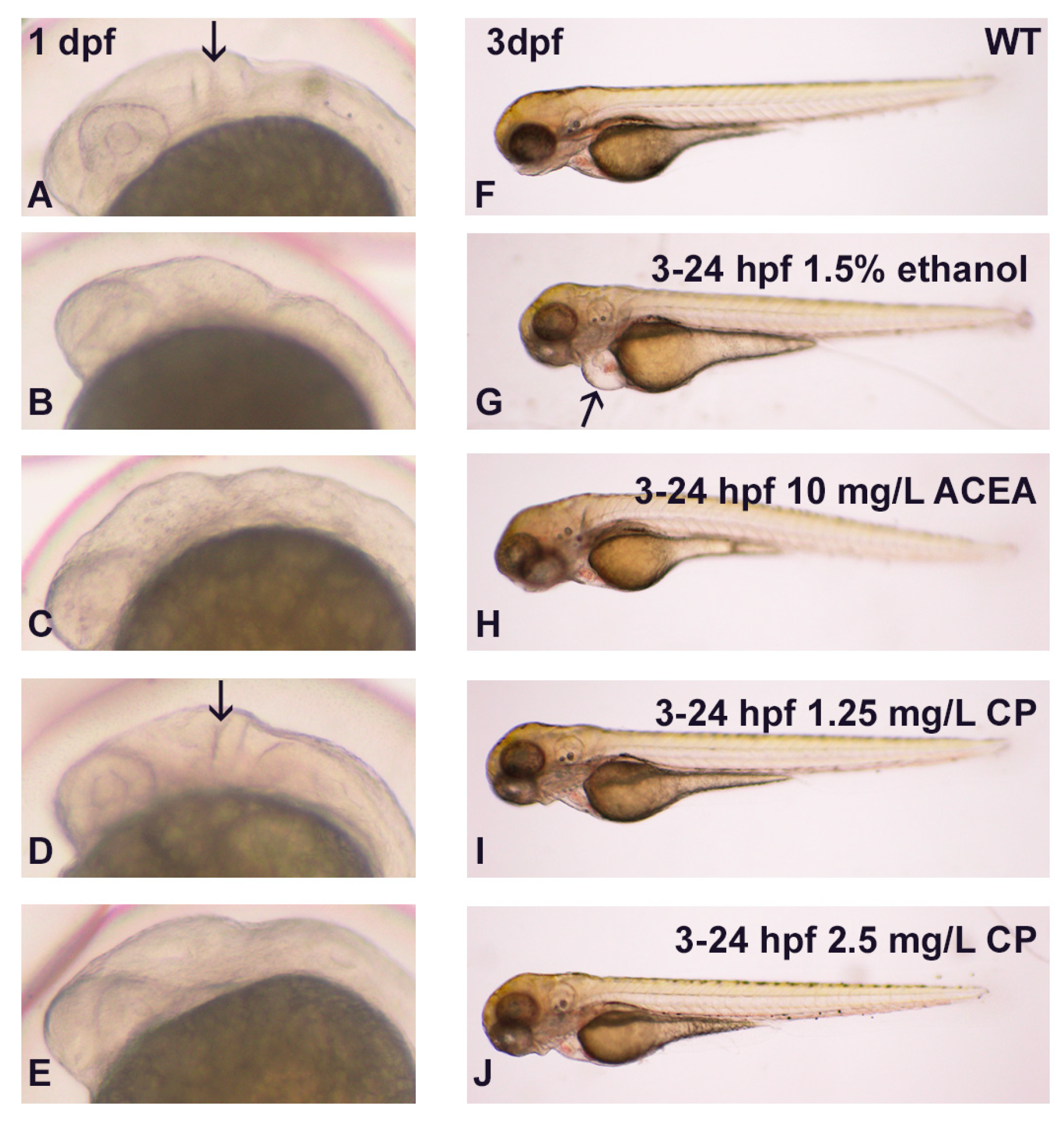
3.1. Effects of Ethanol and Cannabinoids on Zebrafish Embryo Development
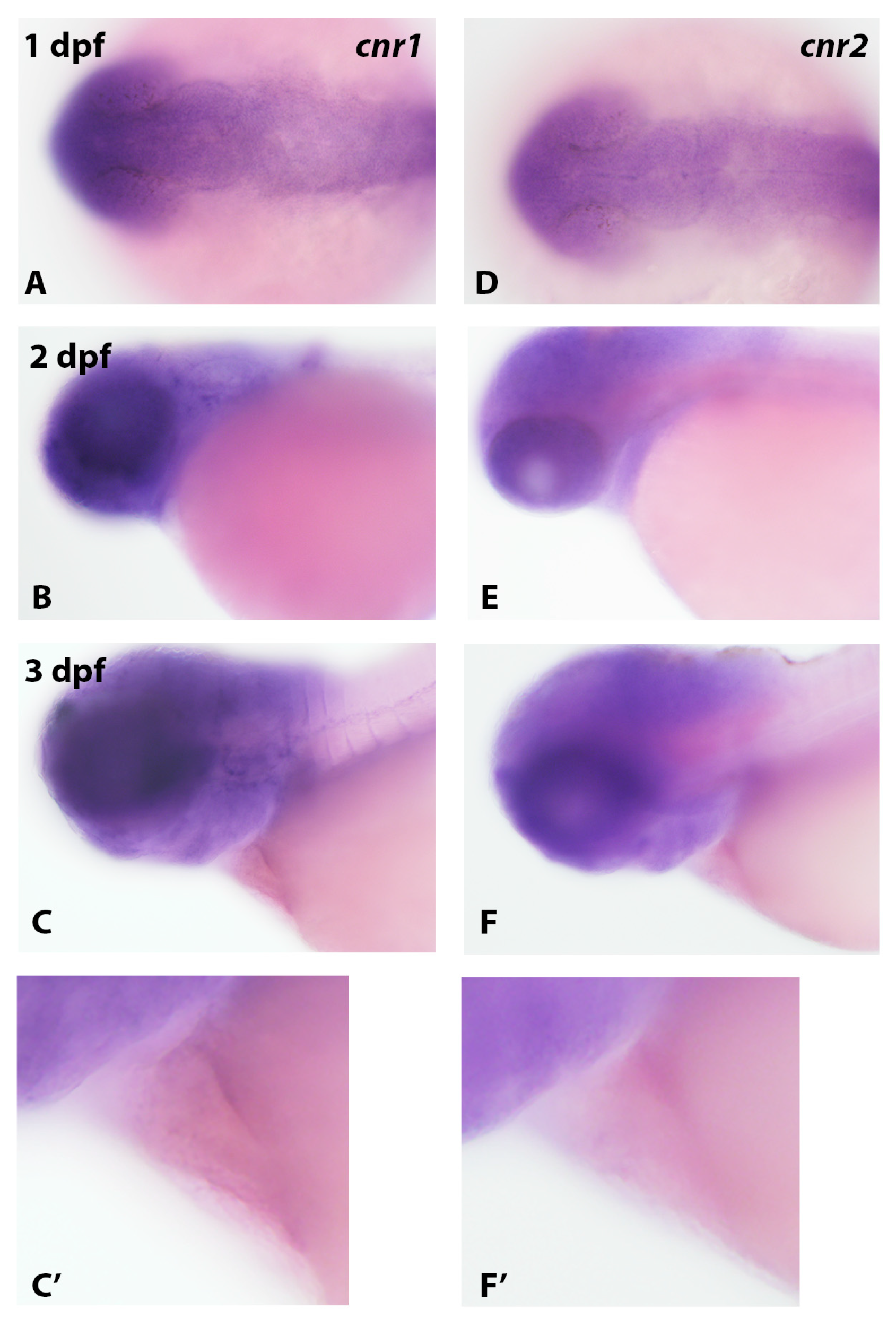
3.2. Analysis of Cannabinoid Receptor Gene Expression in Zebrafish Embryos Using In Situ Hybridization
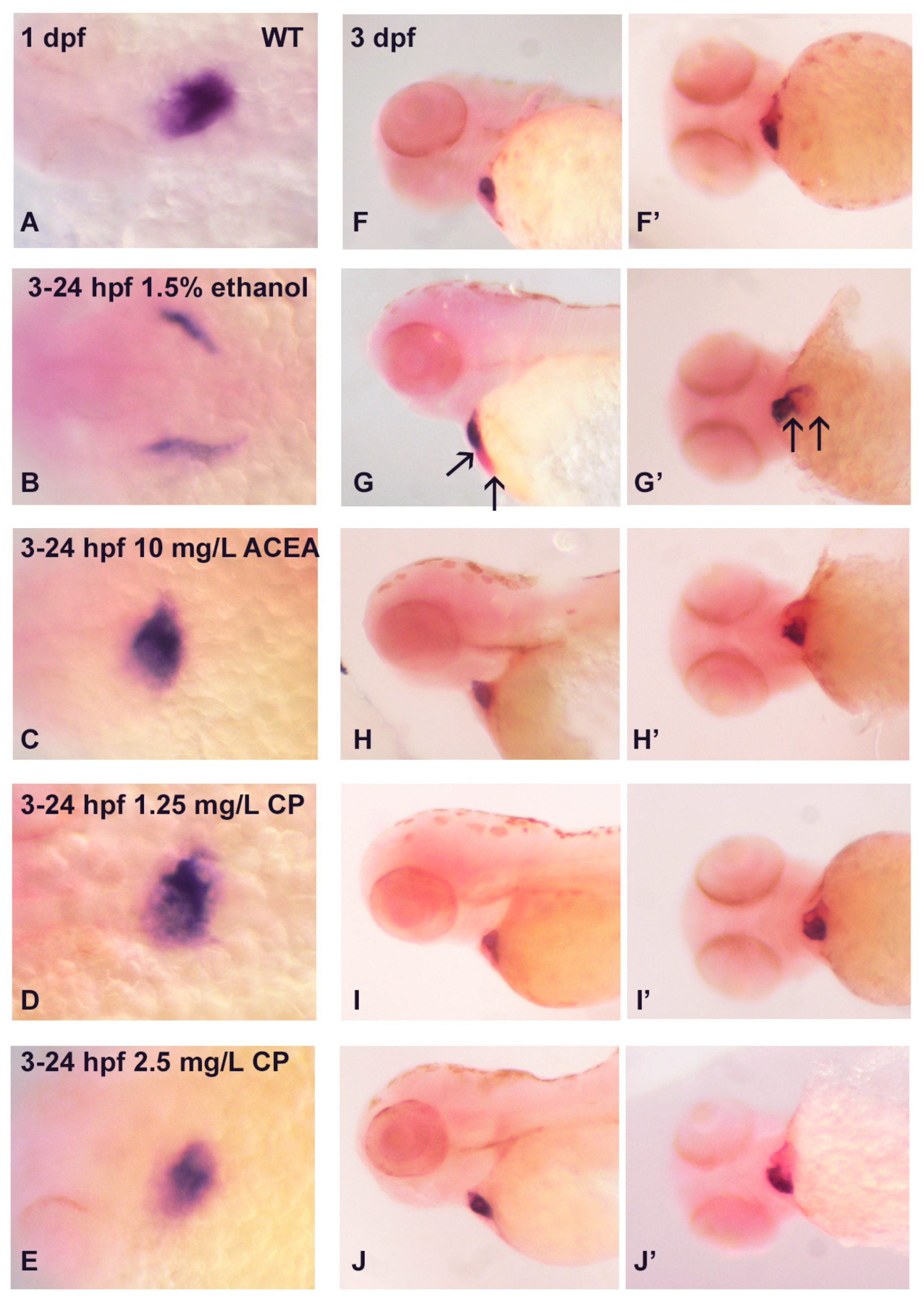
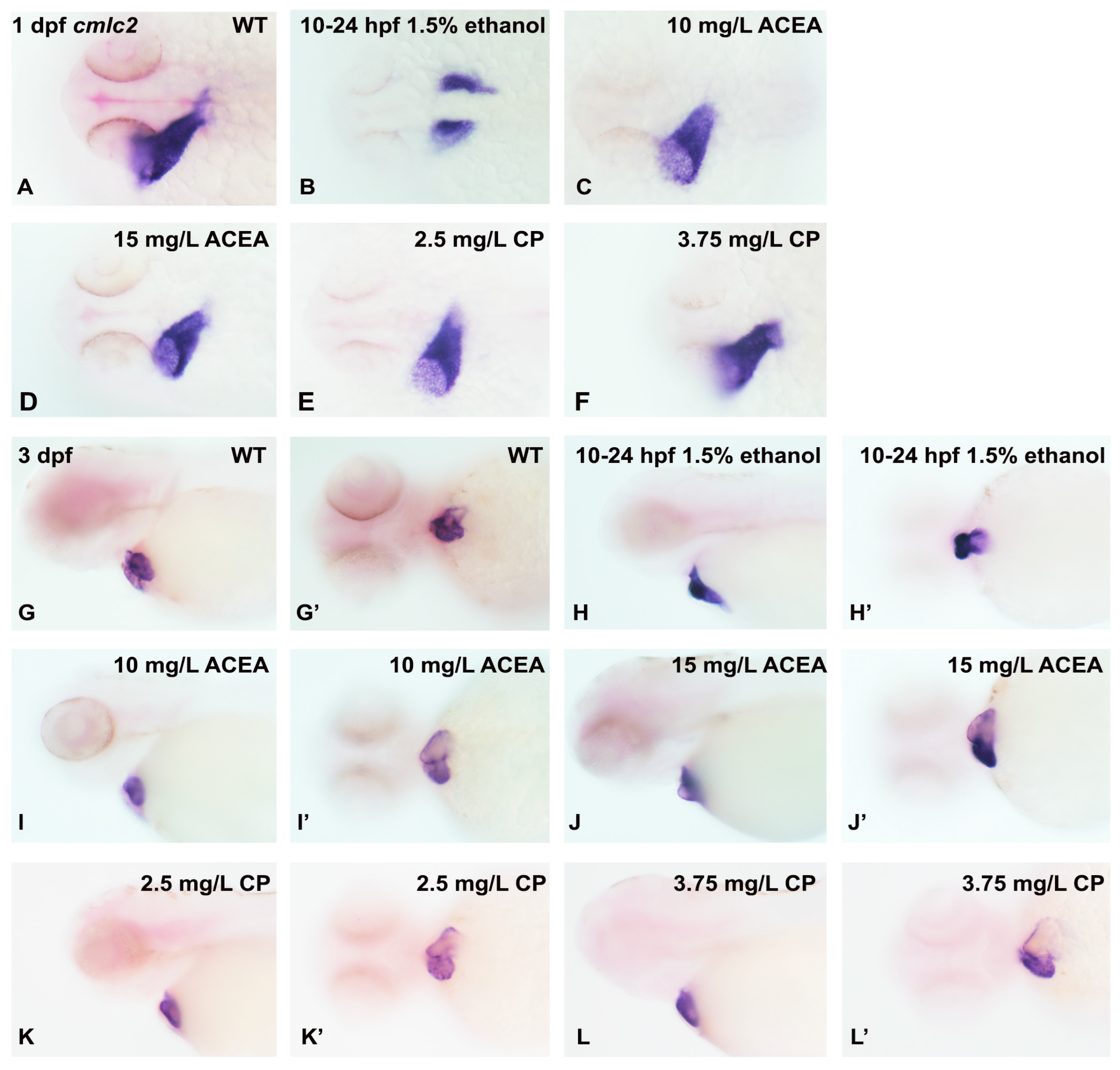
3.3. Does Ethanol Impact Cardiac Development via Impaired Extracellular Matrix Signaling?
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- May, P.A.; Baete, A.; Russo, J.; Elliott, A.J.; Blankenship, J.; Kalberg, W.O.; Buckley, D.; Brooks, M.; Hasken, J.; Abdul-Rahman, O.; et al. Prevalence and characteristics of fetal alcohol spectrum disorders. Pediatrics 2014, 134, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Probst, C.; Gmel, G.; Rehm, J.; Burd, L.; Popova, S. Global prevalence of fetal alcohol spectrum disorder among children and youth: A systematic review and meta-analysis. JAMA Pediatr. 2017, 171, 948–956. [Google Scholar] [CrossRef]
- Sulik, K.K.; Johnston, M.C.; Webb, M.A. Fetal alcohol syndrome: Embryogenesis in a mouse model. Science 1981, 214, 36–938. [Google Scholar] [CrossRef] [PubMed]
- Parnell, S.E.; O’Leary-Moore, S.K.; Godin, E.A.; Dehart, D.B.; Johnson, B.W.; Allan Johnson, G.; Styner, M.A.; Sulik, K.K. Magnetic resonance microscopy defines ethanol-induced brain abnormalities in prenatal mice: Effects of acute insult on gestational day 8. Alcohol. Clin. Exp. Res. 2009, 33, 1001–1011. [Google Scholar] [CrossRef]
- Godin, E.A.; O’Leary-Moore, S.K.; Khan, A.A.; Parnell, S.E.; Ament, J.J.; Dehart, D.B.; Johnson, B.W.; Allan Johnson, G.; Styner, M.A.; Sulik, K.K. Magnetic resonance microscopy defines ethanol-induced brain abnormalities in prenatal mice: Effects of acute insult on gestational day 7. Alcohol. Clin. Exp. Res. 2010, 34, 98–111. [Google Scholar] [CrossRef]
- Sarmah, S.; Marrs, J.A. Complex cardiac defects after ethanol exposure during discrete cardiogenic events in zebrafish: Prevention with folic acid. Dev. Dyn. 2013, 242, 1184–1201. [Google Scholar] [CrossRef]
- Sarmah, S.; Muralidharan, P.; Marrs, J.A. Embryonic ethanol exposure dysregulates Bmp and Notch signaling, leading to persistent atrio-ventricular valve defects in zebrafish. PLoS ONE 2016, 11, e0162105. [Google Scholar] [CrossRef]
- Subbanna, S.; Shivakumar, M.; Psychoyos, D.; Xie, S.; Basavarajappa, B.S. Anandamide CB1 receptor signaling contributes to postnatal ethanol-induced neonatal neurodegeneration, adult synaptic, and memory deficits. J. Neurosci. 2013, 33, 6350–6366. [Google Scholar] [CrossRef]
- Boa-Amponsem, O.; Zhang, C.; Mukhopadhyay, S.; Ardrey, A.; Cole, G.J. Ethanol and cannabinoids interact to alter behavior in a zebrafish fetal alcohol spectrum disorder model. Birth Defects Res. 2019, 111, 775–788. [Google Scholar] [CrossRef]
- Fish, E.W.; Murdaugh, L.B.; Zhang, C.; Boschen, K.E.; Boa-Amponsem, O.; Mendoza-Romero, H.N.; Tarpley, M.; Chdid, L.; Mukhopadhyay, S.; Cole, G.J.; et al. Cannabinoids Exacerbate Alcohol Teratogenesis by a CB1-Hedgehog Interaction. Sci. Rep. 2019, 9, 16057. [Google Scholar] [CrossRef] [PubMed]
- Sobrian, S.K. Developmental cannabinoid exposure: New perspectives on outcomes and mechanisms. Neurotoxicol. Teratol. 2016, 58, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Young-Wolff, K.C.; Tucker, L.-Y.; Alexeeff, S.; Armstrong, M.A.; Conway, A.; Weisner, C.; Goler, N. Trends in Self-reported and Biochemically Tested Marijuana Use Among Pregnant Females in California From 2009–2016. JAMA 2017, 318, 2490–2491. [Google Scholar] [CrossRef]
- Hansen, H.H.; Krutz, B.; Sifringer, M.; Stefovska, V.; Bittigau, P.; Pragst, F.; Marsicano, G.; Lutz, B.; Ikonomidou, C. Cannabinoids enhance susceptibility of immature brain to ethanol neurotoxicity. Ann. Neurol. 2008, 64, 42–52. [Google Scholar] [CrossRef]
- Gilbert, M.T.; Sulik, K.K.; Fish, E.W.; Baker, L.K.; Dehart, D.B.; Parnell, S.E. Dose-dependent teratogenicity of the synthetic cannabinoid CP-55,940 in mice. Neurotoxicol. Teratol. 2016, 58, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Aoto, K.; Shikata, Y.; Higashiyama, D.; Shiota, K.; Motoyama, J. Fetal ethanol exposure activates protein kinase A and impairs Shh expression in prechordal mesendoderm cells in the pathogenesis of holoprosencephaly. Birth Defects Res. Part A Clin. Mol. Teratol. 2008, 82, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Rubert, G.; Minana, R.; Pascual, M.; Guerri, C. Ethanol exposure during embryogenesis decreases the radial glial progenitor pool and affects the generation of neurons and astrocytes. J. Neurosci. Res. 2006, 84, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Yang, H.T.; Zdanowicz, M.; Sicklick, J.K.; Qi, Y.; Camp, T.J.; Diehl, A.M. Fetal alcohol exposure impairs Hedgehog cholesterol modification and signaling. Lab. Investig. 2007, 87, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Ahlgren, S.C.; Thakur, V.; Bronner-Fraser, M. Sonic hedgehog rescues cranial neural crest from cell death induced by ethanol exposure. Proc. Natl. Acad. Sci. USA 2002, 99, 10476–10481. [Google Scholar] [CrossRef]
- Loucks, E.J.; Ahlgren, S.C. Deciphering the role of Shh signaling in axial defects produced by ethanol exposure. Birth Defects Res. Part A Clin. Mol. Teratol. 2009, 85, 556–567. [Google Scholar] [CrossRef]
- Zhang, C.; Turton, Q.M.; MacKinnon, S.; Sulik, K.K.; Cole, G.J. Agrin function associated with ocular development is a target of ethanol exposure in embryonic zebrafish. Birth Defects Res. Part A Clin. Mol. Teratol. 2011, 91, 129–141. [Google Scholar] [CrossRef]
- Zhang, C.; Ojiaku, P.; Cole, G.J. Forebrain and hindbrain development in zebrafish is sensitive to ethanol exposure involving agrin, Fgf and Sonic hedgehog function. Birth Defects Res. Part A Clin. Mol. Teratol. 2013, 97, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.; Zhang, C.; Boa-Amponsem, O.; Mackinnon, S.; Cole, G.J. Long-term behavioral changes as a result of acute ethanol exposure in zebrafish: Evidence for a role for sonic hedgehog but not retinoic acid signaling. Neurotoxicol. Teratol. 2017, 61, 66–73. [Google Scholar] [CrossRef]
- Boa-Amponsem, O.; Zhang, C.; Burton, D.F.; Williams, K.P.; Cole, G.J. Ethanol and cannabinoids regulate zebrafish GABAergic neuron development and behavior in a sonic hedgehog and fibroblast growth factor dependent mechanism. Alcohol. Clin. Exp. Res. 2020, 44, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Krauss, R.S. Cdon mutation and fetal ethanol exposure synergize to produce midline signaling defects and holoprosencephaly spectrum disorders in mice. PLoS Genet. 2012, 8, e1002999. [Google Scholar] [CrossRef] [PubMed]
- Kietzman, H.W.; Everson, J.L.; Sulik, K.K.; Lipinski, R.J. The teratogenic effects of prenatal ethanol exposure are exacerbated by sonic hedgehog or GLI2 haploinsufficiency in the mouse. PLoS ONE 2014, 9, e89448. [Google Scholar] [CrossRef] [PubMed]
- Fish, E.W.; Murdaugh, L.B.; Sulik, K.K.; Williams, K.P.; Parnell, S.E. Genetic vulnerabilities to prenatal alcohol exposure: Limb defects in sonic hedgehog and GLI2 heterozygous mice. Birth Defects Res. 2017, 109, 860–865. [Google Scholar] [CrossRef]
- Kahn, B.M.; Corman, T.S.; Lovelace, K.; Hong, M.; Krauss, R.S.; Epstein, D.J. Prenatal ethanol exposure in mice phenocpoies Cdon mutation by impeding Shh function in the etiology of optic nerve hypoplasia. Dis. Model. Mech. 2017, 10, 29–37. [Google Scholar]
- Khaliullina, H.; Bilgin, M.; Sampaio, J.L.; Shevchenko, A.; Eaton, S. Endocannabinoids are conserved inhibitors of the Hedgehog pathway. Proc. Natl. Acad. Sci. USA 2015, 112, 3415–3420. [Google Scholar] [CrossRef] [PubMed]
- Dyer, L.A.; Kirby, M.L. The role of secondary heart field in cardiac development. Dev. Biol. 2009, 336, 137–144. [Google Scholar] [CrossRef]
- Migliarini, B.; Carnevali, O. A novel role for the endocannabinoid system during zebrafish development. Mol. Cell. Endocrinol. 2009, 299, 172–177. [Google Scholar] [CrossRef]
- Kim, M.J.; Liu, I.H.; Song, Y.; Lee, J.A.; Halfter, W.; Balice-Gordon, R.J.; Linney, E.; Cole, G.J. Agrin is required for posterior development and axon pathway formation in embryonic zebrafish. Glycobiology 2007, 17, 231–247. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Cotman, S.L.; Halfter, W.; Cole, G.J. The heparan sulfate proteoglycan agrin modulates neurite outgrowth mediated by FGF-2. J. Neurobiol. 2003, 55, 261–277. [Google Scholar] [PubMed]
- Bassat, E.; Mutlak, Y.E.; Genzelinakh, A.; Shadrin, I.Y.; Baruch Umansky, K.; Yifa, O.; Kain, D.; Rajchman, D.; Leach, J.; Riabov Bassat, D.; et al. The extracellular matrix protein agrin promotes heart regeneration in mice. Nature 2017, 547, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Frazier, J.M.; Chen, H.; Liu, Y.; Lee, J.A.; Cole, G.J. Molecular and morphological changes in zebrafish following transient ethanol exposure during defined developmental stages. Neurotoxicol. Teratol. 2014, 44, 70–80. [Google Scholar] [CrossRef]
- Liu, I.H.; Zhang, C.; Kim, M.J.; Cole, G.J. Retina development in zebrafish requires the heparan sulfate proteoglycan agrin. Dev. Neurobiol. 2008, 68, 877–898. [Google Scholar] [CrossRef]
- Yelon, D.; Horne, S.A.; Stainier, D.Y.R. Restricted expression of cardiac myosin genes reveals regulated aspects of heart tube assembly in zebrafish. Dev. Biol. 1999, 214, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.K.; Rosales, C.B.; Center, K.E.; Nuñez, A.; Gibson, S.J.; Christ, C.; Ehiri, J.E. Prenatal exposure to cannabis and maternal and child health outcomes: A systematic review and meta-analysis. BMJ Open 2016, 6, e009986. [Google Scholar] [CrossRef]
- Rajesh, M.; Batkai, S.; Kechrid, M.; Mukhopadhyay, P.; Lee, W.-S.; Horvath, B.; Holovac, E.; Cinar, R.; Liaudet, L.; Mackie, K.; et al. Cannabinoid 1receptor promotes cardiac dysfunction, oxidative stress, inflammation, and fibrosis in diabetic cardiomyopathy. Diabetes 2012, 61, 716–727. [Google Scholar] [CrossRef]
- Diez del Corral, R.; Storey, K.G. Opposing FGF and retinoid pathways: A signaling switch that controls differentiation and patterning onset in the extending vertebrate body axis. Bioessays 2004, 26, 857–869. [Google Scholar] [CrossRef]
- Guzzetta, A.; Koska, M.; Rowton, M.; Sullivan, K.R.; Jacobs-Li, J.; Kweon, J.; Hidalgo, H.; Eckart, H.; Hoffmann, A.D.; Back, R.; et al. Hedgehog-FGF signaling axis patterns anterior mesoderm during gastrulation. Proc. Natl. Acad. Sci. USA 2020, 117, 15712–15723. [Google Scholar] [CrossRef]
- Chrisman, K.; Kenney, R.; Comin, J.; Thal, T.; Suchocki, L.; Yueh, Y.G.; Gardner, D.P. Gestational ethanol exposure disrupts the expression of FGF8 and Sonic hedgehog during limb patterning. Birth Defects Res. Part A Clin. Mol. Teratol. 2004, 70, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Halfter, W.; Schurer, B.; Yip, J.; Yip, L.; Tsen, G.; Lee, J.-A.; Cole, G.J. Distribution and substrate properties of agrin, a heparan sulfate proteoglycan of developing axonal pathways. J. Comp. Neurol. 1997, 383, 1–17. [Google Scholar] [CrossRef]
- Tsen, G.; Halfter, W.; Kröger, S.; Cole, G.J. Agrin is a heparan sulfate proteoglycan. J. Biol. Chem. 1995, 270, 3392–3399. [Google Scholar] [CrossRef]
- Cotman, S.L.; Halfter, W.; Cole, G.J. Identification of extracellular matrix ligands for the heparan sulfate proteoglycan agrin. Exp. Cell Res. 1999, 249, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Robles, E.; Staub, W.; Gosse, N.J.; Cole, G.J.; Baier, H. Assembly of lamina-specific neuronal connections by a Slit gradient anchored to type IV collagen. Cell 2011, 146, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Devetzis, V.; Daryadel, A.; Roumeliotis, S.; Theodoridis, M.; Wagner, C.A.; Hettwer, S.; Huynh-Do, U.; Ploumis, P.; Arampatzis, S. C-termina fragment of agrin (CAF): A novel marker for progression of kidney disease in type 2 diabetes. PLoS ONE 2015, 10, e0143524. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Ezem, N.; Mackinnon, S.; Cole, G.J. Embryonic Ethanol but Not Cannabinoid Exposure Affects Zebrafish Cardiac Development via Agrin and Sonic Hedgehog Interaction. Cells 2023, 12, 1327. https://doi.org/10.3390/cells12091327
Zhang C, Ezem N, Mackinnon S, Cole GJ. Embryonic Ethanol but Not Cannabinoid Exposure Affects Zebrafish Cardiac Development via Agrin and Sonic Hedgehog Interaction. Cells. 2023; 12(9):1327. https://doi.org/10.3390/cells12091327
Chicago/Turabian StyleZhang, Chengjin, Natalie Ezem, Shanta Mackinnon, and Gregory J. Cole. 2023. "Embryonic Ethanol but Not Cannabinoid Exposure Affects Zebrafish Cardiac Development via Agrin and Sonic Hedgehog Interaction" Cells 12, no. 9: 1327. https://doi.org/10.3390/cells12091327