
The Role of Eosinophil-Derived Neurotoxin and Vascular Endothelial Growth Factor in the Pathogenesis of Eosinophilic Asthma

Abstract
:1. Introduction
1.1. Eosinophilic Asthma
1.2. Pathophysiology of Asthma
1.2.1. Th2 Response
1.2.2. Th1 Response
1.2.3. Th17 Response
1.3. Aim of the Study
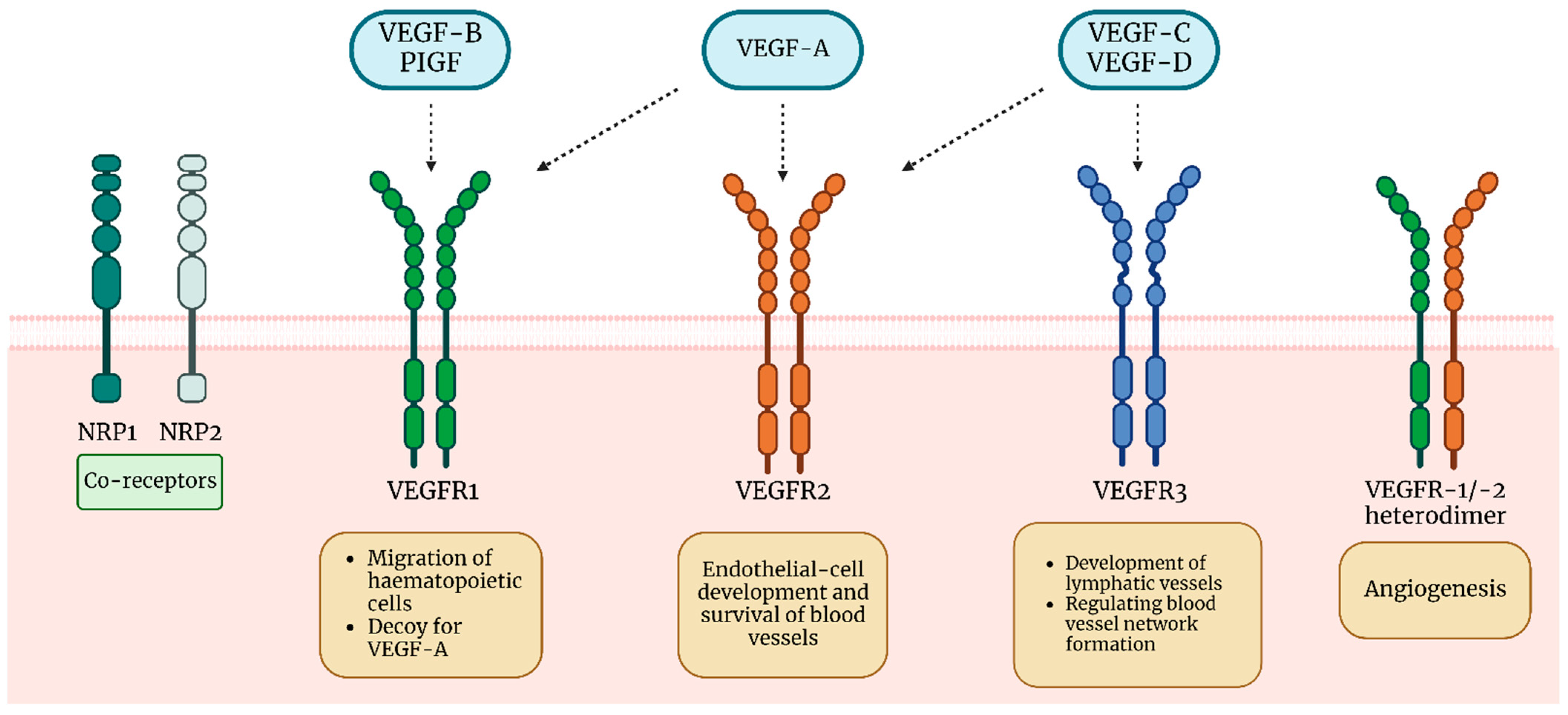
2. VEGF
VEGF in Asthma Pathogenesis
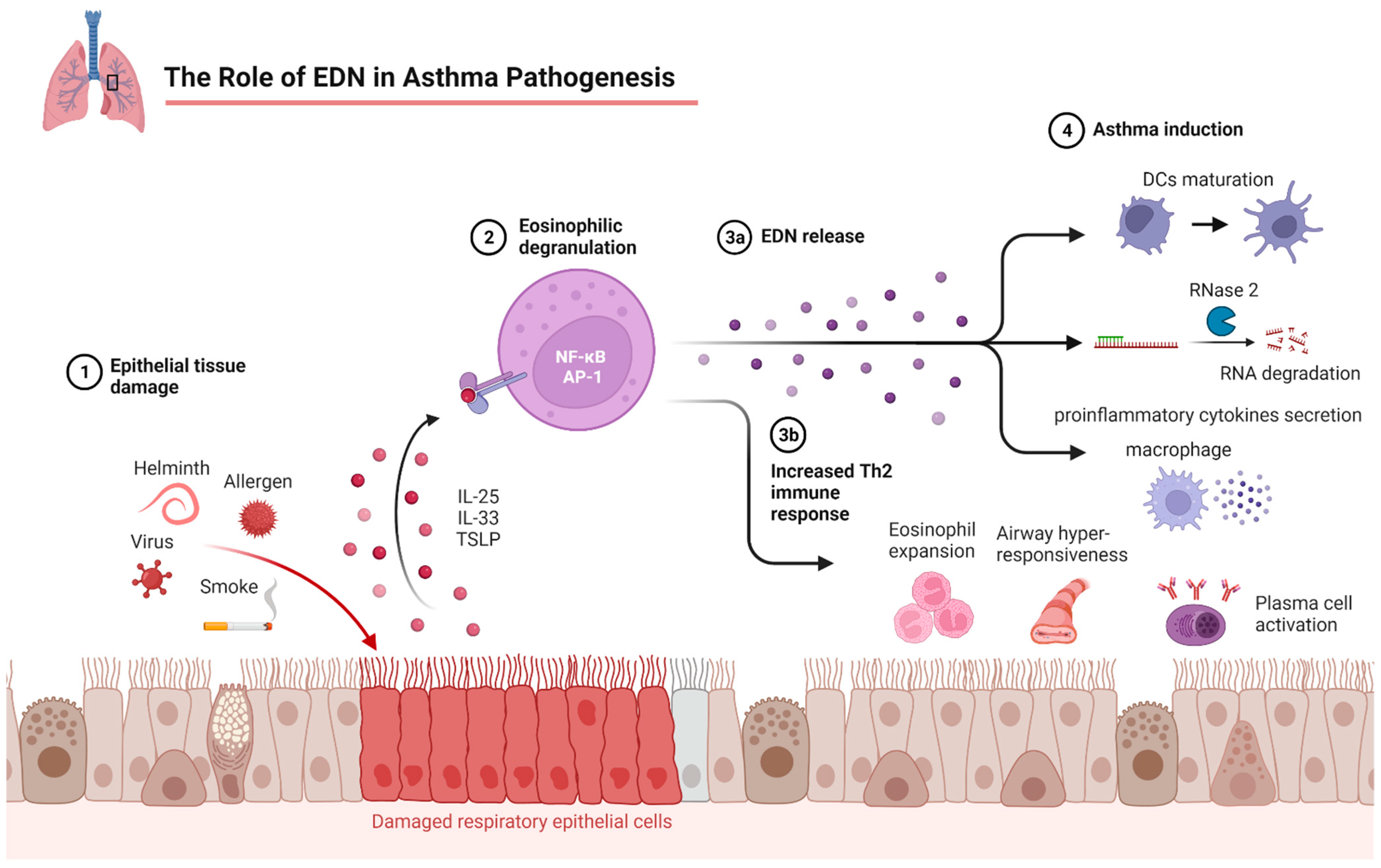
3. EDN
3.1. EDN in Asthma Pathogenesis
3.2. EDN vs. Blood Eosinophil Counts in Asthma Control Status
4. Putative Anti-VEGF and Anti-Eosinophil Drugs in Asthma Therapy
4.1. Anti-VEGF

{kind=link}
{kind=link}
| Drug | Mechanism | Response to Treatment | Adverse Events/Toxicity | Dose | Dose Dependence | Remarks | References |
|---|---|---|---|---|---|---|---|
| Budesonide | ↓VEGF ↓VEGFR-1 | ↑FEV(1) ↓airway hyperresponsiveness | well-tolerated | 100 μg of budesonide + 12 μg of formoterol twice daily or 400 μg of budesonide plus placebo twice daily | at follow-up, clinical and functional outcome measures were better in the group treated with low-dose budesonide + formoterol, compared to the higher dose of budesonide | drug in use | [77,137,138,158] |
| Licochalcone A | ↓VEGFR-2 ↓ERK1/2 ↓caveolin-1 | ND | ND | 10, 20 and 30 μM of licochalcone A per 50 ng/mL of VEGF | the higher doses had a better effect | more studies on humans need to be conducted | [79] |
| Silver nanoparticles | ↓VEGF ↓EGFR ↓MUC5AC | ND | research on mice showed increased pro-inflammatory T17 responses | silver NPs were administered for 48 h with different doses (10, 20, 50, 100, 200, or 500 μM) | elevated levels of HIF-1α, VEGF, and PI3K were significantly reduced in a dose-dependent manner | antiangiogenic effect, potential toxicity depending on genotype and phenotype | [144,145,159] |
| Bevacizumab | ↓VEGF-A ↓VEGFR-2 | ND | reversible bronchospasm | 5 mg/kg body weight | ND | further investigation of the signaling pathway involved in cytokine release and immunoregulation is required | [142,160] |
| Azithromycin | ↓HIF-1α ↓VEGF | ↓airway reactivity and lesions ↓exacerbations and improved quality of life when treated with oral azithromycin in adults with persistent symptomatic asthma ↓respiratory infections |
well-tolerated, increased risk of cardiac torsades des pointes in patients at risk (prolongation of QT interval) and diarrhea | 500 mg three times per week | long-term, low-dose azithromycin reduces asthma exacerbations | the long-term effects on community microbial resistance require further studies | [150,161,162] |
| Diosmetin | ↓VEGF ↓TGF-β1 | ↓airway remodeling ↓fibrogenesis | ND | 0.5 and 0.1 mg/kg | the high dose of diosmetin (0.5 mg/kg) significantly decreased the numbers of total cells, eosinophils, and neutrophils, whereas the low dose of diosmetin (0.1 mg/kg) had only a slight effect | antiproliferative effect on airway smooth muscle cells, antioxidant activity; more research needs to be conducted | [141,163,164] |
| Vitamin D | ↓VEGF-induced respiratory smooth muscle cell proliferation ↓disintegrin and metalloproteinase (ADAM33) | add-on therapy enhances corticosteroid responsiveness | in case of an overdose | 20, 50, 100 nM | inhibited the release of VEGF from ASM cells stimulated with TGF-β1 in a dose-dependent manner | supplementation did not prevent severe asthma exacerbations in children; more research needs to be conducted | [154,165,166] |
| Epigallocatechin gallate | ↓HIF-1α/VEGFA-mediated M2 macrophage skewing in mice ↓TGF-β1 | ND | ND | 10 mg/kg and 50 mg/kg | results indicate that EGCG, especially high-dose EGCG, could alleviate lung injury caused by PM2.5 exposure to asthma | exhibits antiviral, antibacterial, antioxidative, anticancer and chemopreventive activities; more research needs to be conducted | [148,149,167,168] |
| IGFBP-3 | ↓HIF-1α/HIF-2α ↓IGF-I ↓VEGF ↓NF-κB signaling pathway | ↓airway inflammation ↓airway hyperresponsiveness | ND | 10 μg/kg or 50 μg/kg body weight per day | higher dose had a better effect | more research needs to be conducted | [147,169] |
| Neovastat (AE-941) | ↓HIF-2α ↓VEGF | ND | ND | 5 mg/kg | ND | anticancer, antimetastatic antiangiogenic activity; more research needs to be conducted | [155,170] |
| Endostatin/Fc | ↓VEGFR-1,-2,-3 | ↓airway inflammation ↓airway hyperresponsiveness | ND | 20 mg/kg twice a day | ND | antiangiogenic activity, in use in other diseases, possible use in the treatment of asthma requires further study | [156] |
| Montelukast | ↓VEGF ↓IL-6 ↓IL-4 ↓IL-13 ↓eotaxin | ↓airway inflammation ↓airway hyperresponsiveness ↓mucus production ↓pulmonary fibrosis | agitation, anxiety, depression, sleep disturbance, hallucinations, suicidal thinking and suicidality, tremor, dizziness, drowsiness, neuropathies, seizures, anaphylaxis, and eosinophilic infiltration | 2 mg, 10 mg, 50 mg | 10 and 50 mg doses were associated with similar improvement, while the 2 mg dose was less effective and not significantly different from the placebo | IL-6 appears to have a greater effect in blocking irritant-induced asthma | [139,171,172] |
| PEDF | ↓VEGF | ↓airway inflammation ↓airway hyperresponsiveness ↓airway remodeling | ND | 50 or 100 µg/kg body weight | PEDF inhibited the release of VEGF from BEAS-2B cells stimulated with IL-1β in a dose-dependent manner | antiangiogenic, anticancer, and pro-differentiation factor; more research needs to be conducted | [146,173] |
| Melatonin | ↓VEGF ↓TRPV1 channel ↓MUC5AC ↓MAPK signaling | ↓airway inflammation ↓airway hyperresponsiveness ↓airway remodeling | ND | 15 mg/kg body weight | ND | could antagonize ozone-exacerbated asthma, antioxidative, antiproliferative factor; more research needs to be conducted | [157,174,175] |
4.2. Anti-Eosinophil Drugs
| Drug | Mechanism | Response to Treatment | Adverse Events/Toxicity Reported | Remarks | Dose | Dose Dependence | References |
|---|---|---|---|---|---|---|---|
| Budesonide | ↓IL-5 ↓TEC ↓IFN-γ ↓EDN | ↑FEV(1) ↓airway hyperresponsiveness | well-tolerated | effective only in non-smokers | 400 μg/d, 800 μg/d | 400 μg/d reduced the number of IL-5-responsive progenitor cells in the bone marrow at baseline; 800 μg/d reduced circulating eosinophils and serum levels of IL-5, as well as the recruitment of eosinophils to the airway after the allergen challenge | [183,184,185] |
| Benralizumab | ↓IL-5 ↓TEC ↓EDN ↓ECP | ↑FEV(1) ↓AER | well tolerated, injection-site reaction | recommended for 12+ years old children and adults with severe eosinophilic asthma | 25, 30, 100, or 200 mg every 4 weeks | blood eosinophils and sera EDN concentrations were significantly decreased after benralizuma treatment (25, 100, or 200 mg) relative to baseline and were dosage-independent (p < 0.05) | [13,186,187,188,189,190] |
| Mepolizumab | ↓IL-5 ↓EDN | ↑FEV(1), ↓AER improved ACQ | nasopharyngitis, headache, injection-site reaction | recommended for 12+ years old children and adults with severe eosinophilic asthma, men or people having BMI ≥30 had a lesser response | 75 mg, 100, 250 mg, 750 mg every 4 weeks | dose of 750 mg achieved the greatest reduction, dose-related reduction in sputum eosinophils | [13,191,192,193,194,195] |
| Reslizumab | ↓IL-5 ↓EDN ↓TEC | ↑FEV(1), ↓AER | anaphylaxis | recommended for adults with severe eosinophilic asthma | 100 mg, 3 mg/kg intravenously every 4 weeks | no significant difference | [13,177,190,191,196,197] |
| Dupilumab | ↓IL-4 ↓IL-13 ↓serum IgE, ↓plasma eotaxin-3 | ↑FEV(1), ↓AER ↓FeNO, | injection-site reaction, nasopharyngitis, injection-site erythema, bronchitis, hypereosinophilia | recommended for 12+ years old children and adults with severe eosinophilic asthma, especially effective in patients with high levels of blood eosinophils and FENO | 100 mg for those weighing ≤30 kg, 200 mg every 2 weeks, 300 mg every 2 weeks | no significant difference | [13,190,198,199,200,201] |
| Lebrikizumab | ↓IL-13 | ↑FEV(1) ↓AER | injection-site reaction | did not always show a significant reduction in asthma exacerbations in biomarker-high patients |
37.5 mg, 125 mg, 250 mg | 37.5 mg (81% reduction of exacerbation rate), 125 mg (77% reduction), 250 mg (no significant reduction) | [202,203,204,205] |
| Tralokinumab | ↓IL-13 | ↑FEV(1) ↑FVC ↓AER | well-tolerated | promising results in atopic dermatitis | 150 mg, 300 mg, 600 mg subcutaneously every 2 weeks | only tralokinumab 600 mg improved FEV1 significantly | [206,207,208] |
| Omalizumab | ↓EDN ↓serum IgE | ↑FEV(1), ↑FVC ↑PEF ↓AER | anaphylaxis | recommended for adults with severe eosinophilic asthma | 75–375 mg subcutaneously every two or four weeks | the basis for dose and frequency is calculated considering patient weight and pretreatment total IgE serum levels; a minimum dose of 0.008 mg/kg of body weight per IgE (IU/mL) every 2 weeks or 0.016 mg/kg per IgE (IU/mL) every 4 weeks | [13,129,209,210,211] |
| Tezepelumab | ↓TSLP ↓IL-5 ↓IL-13 ↓IgE | ↑FEV(1) ↓AER ↓FeNO | nasopharyngitis, upper respiratory tract infection, headache, injection-site reaction | add-on maintenance treatment for 12+ years old children and adults | 70 mg, 210 mg, 280 mg every 4 weeks | annualized asthma exacerbation rates at week 52 were 0.27 (70 mg), 0.20 (210 mg), and 0.23 (280 mg), compared with 0.72 in the placebo group; 210 mg dose reduced exacerbation rates by 64–82% | [206,212,213] |
| Fevipiprant | ↓PGD2 | ↑FEV(1) ↓AER improved ACQ | well-tolerated | not recommended for routine use, further studies are needed | 150 mg/d, 450 mg/d | RR of asthma exacerbation: 0.77 (0.61–0.97) for 450 mg compared with placebo; 0.86 (0.69–1.08) for 150 mg dose | [214] |
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vos, T.; Abajobir, A.A.; Abbafati, C.; Abbas, K.M.; Abate, K.H.; Abd-Allah, F.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F.; Aboyans, V.; et al. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 328 Diseases and Injuries for 195 Countries, 1990-2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Adeloye, D.; Salim, H.; Dos Santos, J.P.; Campbell, H.; Sheikh, A.; Rudan, I. Global, Regional, and National Prevalence of Asthma in 2019: A Systematic Analysis and Modelling Study. J. Glob. Health 2022, 12, 04052. [Google Scholar] [CrossRef] [PubMed]
- Abbafati, C.; Abbas, K.M.; Abbasi-Kangevari, M.; Abd-Allah, F.; Abdelalim, A.; Abdollahi, M.; Abdollahpour, I.; Abegaz, K.H.; Abolhassani, H.; Aboyans, V.; et al. Global Burden of 369 Diseases and Injuries in 204 Countries and Territories, 1990-2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- Lee, Y.J.; Fujisawa, T.; Kim, C.K. Biomarkers for Recurrent Wheezing and Asthma in Preschool Children. Allergy Asthma Immunol. Res. 2019, 11, 16–28. [Google Scholar] [CrossRef]
- Wu, T.D.; Brigham, E.P.; McCormack, M.C. Asthma in the Primary Care Setting. Med. Clin. N. Am. 2019, 103, 435–452. [Google Scholar] [CrossRef]
- Gans, M.D.; Gavrilova, T. Understanding the Immunology of Asthma: Pathophysiology, Biomarkers, and Treatments for Asthma Endotypes. Paediatr. Respir. Rev. 2020, 36, 118–127. [Google Scholar] [CrossRef]
- Ntontsi, P.; Photiades, A.; Zervas, E.; Xanthou, G.; Samitas, K. Genetics and Epigenetics in Asthma. Int. J. Mol. Sci. 2021, 22, 2412. [Google Scholar] [CrossRef]
- Maslan, J.; Mims, J.W. What Is Asthma? Pathophysiology, Demographics, and Health Care Costs. Otolaryngol. Clin. N. Am. 2014, 47, 13–22. [Google Scholar] [CrossRef]
- Habib, N.; Pasha, M.A.; Tang, D.D. Current Understanding of Asthma Pathogenesis and Biomarkers. Cells 2022, 11, 2764. [Google Scholar] [CrossRef]
- Popović-Grle, S.; Štajduhar, A.; Lampalo, M.; Rnjak, D. Biomarkers in Different Asthma Phenotypes. Genes 2021, 12, 801. [Google Scholar] [CrossRef]
- Schoettler, N.; Strek, M.E. Recent Advances in Severe Asthma: From Phenotypes to Personalized Medicine. Chest 2020, 157, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Lee, H.K. The Role of CD4+ T Cells and Microbiota in the Pathogenesis of Asthma. Int. J. Mol. Sci. 2021, 22, 11822. [Google Scholar] [CrossRef] [PubMed]
- Agache, I.; Akdis, C.A.; Akdis, M.; Canonica, G.W.; Casale, T.; Chivato, T.; Corren, J.; Chu, D.K.; Del Giacco, S.; Eiwegger, T.; et al. EAACI Biologicals Guidelines-Recommendations for Severe Asthma. Allergy 2021, 76, 14–44. [Google Scholar] [CrossRef] [PubMed]
- Carr, T.F.; Zeki, A.A.; Kraft, M. Eosinophilic and Noneosinophilic Asthma. Am. J. Respir. Crit. Care Med. 2018, 197, 22–37. [Google Scholar] [CrossRef]
- Rothenberg, M.E.; Hogan, S.P. The Eosinophil. Annu. Rev. Immunol. 2006, 24, 147–174. [Google Scholar] [CrossRef]
- Jacobsen, E.A.; Ochkur, S.I.; Lee, N.A.; Lee, J.J. Eosinophils and Asthma. Curr. Allergy Asthma Rep. 2007, 7, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, M. Targeting Eosinophil Biology in Asthma Therapy. Am. J. Respir. Cell Mol. Biol. 2011, 45, 667–674. [Google Scholar] [CrossRef]
- Karawajczyk, M.; Peterson, C.G.B.; Venge, P.; Garcia, R.C. An Extragranular Compartment of Blood Eosinophils Contains Eosinophil Protein X/Eosinophil-Derived Neurotoxin (EPX/EDN). Inflammation 2013, 36, 320–329. [Google Scholar] [CrossRef]
- Tsuda, T.; Maeda, Y.; Nishide, M.; Koyama, S.; Hayama, Y.; Nojima, S.; Takamatsu, H.; Okuzaki, D.; Kinehara, Y.; Kato, Y.; et al. Eosinophil-Derived Neurotoxin Enhances Airway Remodeling in Eosinophilic Chronic Rhinosinusitis and Correlates with Disease Severity. Int. Immunol. 2019, 31, 33–40. [Google Scholar] [CrossRef]
- Mims, J.W. Asthma: Definitions and Pathophysiology. Int. Forum Allergy Rhinol. 2015, 5 (Suppl. S1), S2–S6. [Google Scholar] [CrossRef]
- Woodruff, P.G.; Modrek, B.; Choy, D.F.; Jia, G.; Abbas, A.R.; Ellwanger, A.; Arron, J.R.; Koth, L.L.; Fahy, J.V. T-Helper Type 2-Driven Inflammation Defines Major Subphenotypes of Asthma. Am. J. Respir. Crit. Care Med. 2009, 180, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Fahy, J.V. Type 2 Inflammation in Asthma—Present in Most, Absent in Many. Nat. Rev. Immunol. 2015, 15, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, R.K.; Al Heialy, S.; Hamid, Q. Role of IL-17 in Asthma Pathogenesis and Its Implications for the Clinic. Expert Rev. Respir. Med. 2019, 13, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The Cytokines of Asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef]
- Yamauchi, K.; Ogasawara, M. The Role of Histamine in the Pathophysiology of Asthma and the Clinical Efficacy of Antihistamines in Asthma Therapy. Int. J. Mol. Sci. 2019, 20, 1733. [Google Scholar] [CrossRef]
- Varricchi, G.; Bagnasco, D.; Borriello, F.; Heffler, E.; Canonica, G.W. Interleukin-5 Pathway Inhibition in the Treatment of Eosinophilic Respiratory Disorders: Evidence and Unmet Needs. Curr. Opin. Allergy Clin. Immunol. 2016, 16, 186–200. [Google Scholar] [CrossRef]
- Conde, E.; Bertrand, R.; Balbino, B.; Bonnefoy, J.; Stackowicz, J.; Caillot, N.; Colaone, F.; Hamdi, S.; Houmadi, R.; Loste, A.; et al. Dual Vaccination against IL-4 and IL-13 Protects against Chronic Allergic Asthma in Mice. Nat. Commun. 2021, 12, 2574. [Google Scholar] [CrossRef]
- McGrath, K.W.; Icitovic, N.; Boushey, H.A.; Lazarus, S.C.; Sutherland, E.R.; Chinchilli, V.M.; Fahy, J.V. A Large Subgroup of Mild-to-Moderate Asthma Is Persistently Noneosinophilic. Am. J. Respir. Crit. Care Med. 2012, 185, 612–619. [Google Scholar] [CrossRef]
- Knochelmann, H.M.; Dwyer, C.J.; Bailey, S.R.; Amaya, S.M.; Elston, D.M.; Mazza-McCrann, J.M.; Paulos, C.M. When Worlds Collide: Th17 and Treg Cells in Cancer and Autoimmunity. Cell. Mol. Immunol. 2018, 15, 458–469. [Google Scholar] [CrossRef]
- Sieck, G.C.; Dogan, M.; Young-Soo, H.; Osorio Valencia, S.; Delmotte, P. Mechanisms Underlying TNFα-Induced Enhancement of Force Generation in Airway Smooth Muscle. Physiol. Rep. 2019, 7, e14220. [Google Scholar] [CrossRef]
- Niessen, N.M.; Gibson, P.G.; Baines, K.J.; Barker, D.; Yang, I.A.; Upham, J.W.; Reynolds, P.N.; Hodge, S.; James, A.L.; Jenkins, C.; et al. Sputum TNF Markers Are Increased in Neutrophilic and Severe Asthma and Are Reduced by Azithromycin Treatment. Allergy 2021, 76, 2090–2101. [Google Scholar] [CrossRef] [PubMed]
- Raundhal, M.; Morse, C.; Khare, A.; Oriss, T.B.; Milosevic, J.; Trudeau, J.; Huff, R.; Pilewski, J.; Holguin, F.; Kolls, J.; et al. High IFN-γ and Low SLPI Mark Severe Asthma in Mice and Humans. J. Clin. Investig. 2015, 125, 3037–3050. [Google Scholar] [CrossRef] [PubMed]
- Hudey, S.N.; Ledford, D.K.; Cardet, J.C. Mechanisms of Non-Type 2 Asthma. Curr. Opin. Immunol. 2020, 66, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Al-Alwan, L.; Risse, P.A.; Roussel, L.; Rousseau, S.; Halayko, A.J.; Martin, J.G.; Hamid, Q.; Eidelman, D.H. TH17 Cytokines Induce Human Airway Smooth Muscle Cell Migration. J. Allergy Clin. Immunol. 2011, 127, 1046–1053.e2. [Google Scholar] [CrossRef]
- Kudo, M.; Melton, A.C.; Chen, C.; Engler, M.B.; Huang, K.E.; Ren, X.; Wang, Y.; Bernstein, X.; Li, J.T.; Atabai, K.; et al. IL-17A Produced by Aβ T Cells Drives Airway Hyper-Responsiveness in Mice and Enhances Mouse and Human Airway Smooth Muscle Contraction. Nat. Med. 2012, 18, 547–554. [Google Scholar] [CrossRef]
- Hynes, G.M.; Hinks, T.S.C. The Role of Interleukin-17 in Asthma: A Protective Response? ERJ Open Res. 2020, 6, 00364–02019. [Google Scholar] [CrossRef]
- Kim, C.K. Eosinophil-Derived Neurotoxin: A Novel Biomarker for Diagnosis and Monitoring of Asthma. Korean J. Pediatr. 2013, 56, 8–12. [Google Scholar] [CrossRef]
- Lee, C.G.; Ma, B.; Takyar, S.; Ahangari, F.; DelaCruz, C.; He, C.H.; Elias, J.A. Studies of Vascular Endothelial Growth Factor in Asthma and Chronic Obstructive Pulmonary Disease. Proc. Am. Thorac. Soc. 2011, 8, 512–515. [Google Scholar] [CrossRef]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Morita, T. Molecular and Functional Diversity of Vascular Endothelial Growth Factors. Mol. Divers. 2006, 10, 515–527. [Google Scholar] [CrossRef]
- Voelkel, N.F.; Vandivier, R.W.; Tuder, R.M. Vascular Endothelial Growth Factor in the Lung. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L209–L221. [Google Scholar] [CrossRef] [PubMed]
- Tuder, R.M.; Yun, J.H. Vascular Endothelial Growth Factor of the Lung: Friend or Foe. Curr. Opin. Pharmacol. 2008, 8, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Vitt, U.A.; Hsu, S.Y.; Hsueh, A.J.W. Evolution and Classification of Cystine Knot-Containing Hormones and Related Extracellular Signaling Molecules. Mol. Endocrinol. 2001, 15, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.; Acharya, K.R. Tying the Knot: The Cystine Signature and Molecular-Recognition Processes of the Vascular Endothelial Growth Factor Family of Angiogenic Cytokines. FEBS J. 2011, 278, 4304–4322. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhu, X.; Cui, H.; Shi, J.; Yuan, G.; Shi, S.; Hu, Y. The Role of the VEGF Family in Coronary Heart Disease. Front. Cardiovasc. Med. 2021, 8, 738325. [Google Scholar] [CrossRef]
- Mayer, G. Capillary Rarefaction, Hypoxia, VEGF and Angiogenesis in Chronic Renal Disease. Nephrol. Dial. Transpl. 2011, 26, 1132–1137. [Google Scholar] [CrossRef]
- Long, H.Q.; Li, G.S.; Cheng, X.; Xu, J.H.; Li, F.B. Role of Hypoxia-Induced VEGF in Blood-Spinal Cord Barrier Disruption in Chronic Spinal Cord Injury. Chin. J. Traumatol. 2015, 18, 293–295. [Google Scholar] [CrossRef]
- Uemura, A.; Fruttiger, M.; D’Amore, P.A.; De Falco, S.; Joussen, A.M.; Sennlaub, F.; Brunck, L.R.; Johnson, K.T.; Lambrou, G.N.; Rittenhouse, K.D.; et al. VEGFR1 Signaling in Retinal Angiogenesis and Microinflammation. Prog. Retin. Eye Res. 2021, 84, 100954. [Google Scholar] [CrossRef]
- Baldwin, M.E.; Halford, M.M.; Roufail, S.; Williams, R.A.; Hibbs, M.L.; Grail, D.; Kubo, H.; Stacker, S.A.; Achen, M.G. Vascular Endothelial Growth Factor D Is Dispensable for Development of the Lymphatic System. Mol. Cell. Biol. 2005, 25, 2441–2449. [Google Scholar] [CrossRef]
- Ferrara, N. Role of Vascular Endothelial Growth Factor in the Regulation of Angiogenesis. Kidney Int. 1999, 56, 794–814. [Google Scholar] [CrossRef]
- Elebiyo, T.C.; Rotimi, D.; Evbuomwan, I.O.; Maimako, R.F.; Iyobhebhe, M.; Ojo, O.A.; Oluba, O.M.; Adeyemi, O.S. Reassessing Vascular Endothelial Growth Factor (VEGF) in Anti-Angiogenic Cancer Therapy. Cancer Treat. Res. Commun. 2022, 32, 100620. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Hristova, K. Receptor Tyrosine Kinase Transmembrane Domains: Function, Dimer Structure and Dimerization Energetics. Cell Adh. Migr. 2010, 4, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S. Vascular Endothelial Growth Factor (VEGF), VEGF Receptors and Their Inhibitors for Antiangiogenic Tumor Therapy. Biol. Pharm. Bull. 2011, 34, 1785–1788. [Google Scholar] [CrossRef] [PubMed]
- Secker, G.A.; Harvey, N.L. Regulation of VEGFR Signalling in Lymphatic Vascular Development and Disease: An Update. Int. J. Mol. Sci. 2021, 22, 7760. [Google Scholar] [CrossRef] [PubMed]
- Cébe-Suarez, S.; Zehnder-Fjällman, A.; Ballmer-Hofer, K. The Role of VEGF Receptors in Angiogenesis; Complex Partnerships. Cell. Mol. Life Sci. 2006, 63, 601–615. [Google Scholar] [CrossRef]
- Leppänen, V.M.; Tvorogov, D.; Kisko, K.; Prota, A.E.; Jeltsch, M.; Anisimov, A.; Markovic-Mueller, S.; Stuttfeld, E.; Goldie, K.N.; Ballmer-Hofer, K.; et al. Structural and Mechanistic Insights into VEGF Receptor 3 Ligand Binding and Activation. Proc. Natl. Acad. Sci. USA 2013, 110, 12960–12965. [Google Scholar] [CrossRef]
- Stuttfeld, E.; Ballmer-Hofer, K. Structure and Function of VEGF Receptors. IUBMB Life 2009, 61, 915–922. [Google Scholar] [CrossRef]
- Fuh, G.; Li, B.; Crowley, C.; Cunningham, B.; Wells, J.A. Requirements for Binding and Signaling of the Kinase Domain Receptor for Vascular Endothelial Growth Factor. J. Biol. Chem. 1998, 273, 11197–11204. [Google Scholar] [CrossRef]
- Takahashi, H.; Shibuya, M. The Vascular Endothelial Growth Factor (VEGF)/VEGF Receptor System and Its Role under Physiological and Pathological Conditions. Clin. Sci. 2005, 109, 227–241. [Google Scholar] [CrossRef]
- Simons, M.; Gordon, E.; Claesson-Welsh, L. Mechanisms and Regulation of Endothelial VEGF Receptor Signalling. Nat. Rev. Mol. Cell Biol. 2016, 17, 611–625. [Google Scholar] [CrossRef]
- Tammela, T.; Zarkada, G.; Wallgard, E.; Murtomäki, A.; Suchting, S.; Wirzenius, M.; Waltari, M.; Hellström, M.; Schomber, T.; Peltonen, R.; et al. Blocking VEGFR-3 Suppresses Angiogenic Sprouting and Vascular Network Formation. Nature 2008, 454, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Jena, M.K.; Sharma, N.R.; Petitt, M.; Maulik, D.; Nayak, N.R. Pathogenesis of Preeclampsia and Therapeutic Approaches Targeting the Placenta. Biomolecules 2020, 10, 953. [Google Scholar] [CrossRef] [PubMed]
- Goldman, C.K.; Kendall, R.L.; Cabrera, G.; Soroceanu, L.; Heike, Y.; Gillespie, G.Y.; Siegal, G.P.; Mao, X.; Bett, A.J.; Huckle, W.R.; et al. Paracrine Expression of a Native Soluble Vascular Endothelial Growth Factor Receptor Inhibits Tumor Growth, Metastasis, and Mortality Rate. Proc. Natl. Acad. Sci. USA 1998, 95, 8795–8800. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, T.; Saito-Takatsuji, H.; Yoshitomi, Y.; Yonekura, H. Role of Arginine Methylation in Alternative Polyadenylation of VEGFR-1 (Flt-1) Pre-MRNA. Int. J. Mol. Sci. 2020, 21, 6460. [Google Scholar] [CrossRef]
- Rahimi, N. Vascular Endothelial Growth Factor Receptors: Molecular Mechanisms of Activation and Therapeutic Potentials. Exp. Eye Res. 2006, 83, 1005–1016. [Google Scholar] [CrossRef]
- Rahimi, N. VEGFR-1 and VEGFR-2: Two Non-Identical Twins with a Unique Physiognomy. Front. Biosci. 2006, 11, 818–829. [Google Scholar] [CrossRef]
- Tjwa, M.; Luttun, A.; Autiero, M.; Carmeliet, P. VEGF and PlGF: Two Pleiotropic Growth Factors with Distinct Roles in Development and Homeostasis. Cell Tissue Res. 2003, 314, 5–14. [Google Scholar] [CrossRef]
- Geretti, E.; Klagsbrun, M. Neuropilins: Novel Targets for Anti-Angiogenesis Therapies. Cell Adh. Migr. 2007, 1, 56–61. [Google Scholar] [CrossRef]
- Niland, S.; Eble, J.A. Neuropilins in the Context of Tumor Vasculature. Int. J. Mol. Sci. 2019, 20, 639. [Google Scholar] [CrossRef]
- Dumbrăveanu, L.; Cușnir, V.; Bobescu, D. A Review of Neovascular Glaucoma. Etiopathogenesis and Treatment. Rom. J. Ophthalmol. 2021, 65, 315–329. [Google Scholar] [CrossRef]
- Gomułka, K.; Liebhart, J.; Gładysz, U.; Mȩdrala, W. VEGF Serum Concentration and Irreversible Bronchoconstriction in Adult Asthmatics. Adv. Clin. Exp. Med. 2019, 28, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, M.; Nakamura, Y.; Hamid, Q.A. Gene Expression of Vascular Endothelial Growth Factor and Its Receptors and Angiogenesis in Bronchial Asthma. J. Allergy Clin. Immunol. 2001, 107, 1034–1038. [Google Scholar] [CrossRef] [PubMed]
- Meyer, N.; Christoph, J.; Makrinioti, H.; Indermitte, P.; Rhyner, C.; Soyka, M.; Eiwegger, T.; Chalubinski, M.; Wanke, K.; Fujita, H.; et al. Inhibition of Angiogenesis by IL-32: Possible Role in Asthma. J. Allergy Clin. Immunol. 2012, 129, 964–973.e7. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Guisa, J.M.; Powers, C.; File, D.; Cochrane, E.; Jimenez, N.; Debley, J.S. Airway Epithelial Cells from Asthmatic Children Differentially Express Proremodeling Factors. J. Allergy Clin. Immunol. 2012, 129, 990–997.e6. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, M.; Takahashi, M.; Aoike, N. Expression of Vascular Endothelial Growth Factor, Basic Fibroblast Growth Factor, and Angiogenin Immunoreactivity in Asthmatic Airways and Its Relationship to Angiogenesis. J. Allergy Clin. Immunol. 2001, 107, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lin, J.; Ren, Z.; Li, X. Predictive Effect of Exhaled NO and VEGF Expression Levels on the Severity of Bronchial Asthma and Airway Inflammation. Int. J. Clin. Exp. Pathol. 2018, 11, 5401–5406. [Google Scholar]
- Wang, K.; Liu, C.T.; Wu, Y.H.; Feng, Y.L.; Bai, H.L. Budesonide/Formoterol Decreases Expression of Vascular Endothelial Growth Factor (VEGF) and VEGF Receptor 1 within Airway Remodelling in Asthma. Adv. Ther. 2008, 25, 342–354. [Google Scholar] [CrossRef]
- Kim, S.H.; Pei, Q.M.; Jiang, P.; Liu, J.; Sun, R.F.; Qian, X.J.; Liu, J.B. Upregulation of MUC5AC by VEGF in Human Primary Bronchial Epithelial Cells: Implications for Asthma. Respir. Res. 2019, 20, 282. [Google Scholar] [CrossRef]
- Kim, S.H.; Pei, Q.M.; Jiang, P.; Yang, M.; Qian, X.J.; Liu, J.B. Role of Licochalcone A in VEGF-Induced Proliferation of Human Airway Smooth Muscle Cells: Implications for Asthma. Growth Factors 2017, 35, 39–47. [Google Scholar] [CrossRef]
- Wang, H.; Keiser, J.A. Vascular Endothelial Growth Factor Upregulates the Expression of Matrix Metalloproteinases in Vascular Smooth Muscle Cells: Role of Flt-1. Circ. Res. 1998, 83, 832–840. [Google Scholar] [CrossRef]
- Pei, Q.M.; Jiang, P.; Yang, M.; Qian, X.J.; Liu, J.B.; Zheng, H.; Zhao, L.H.; Kim, S.H. Upregulation of a Disintegrin and Metalloproteinase-33 by VEGF in Human Airway Smooth Muscle Cells: Implications for Asthma. Cell Cycle 2016, 15, 2819–2826. [Google Scholar] [CrossRef] [PubMed]
- Clifford, R.L.; John, A.E.; Brightling, C.E.; Knox, A.J. Abnormal Histone Methylation Is Responsible for Increased Vascular Endothelial Growth Factor 165a Secretion from Airway Smooth Muscle Cells in Asthma. J. Immunol. 2012, 189, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Chalubinski, M.; Rhyner, C.; Indermitte, P.; Meyer, N.; Ferstl, R.; Treis, A.; Gomez, E.; Akkaya, A.; O’Mahony, L.; et al. Claudin-1 Expression in Airway Smooth Muscle Exacerbates Airway Remodeling in Asthmatic Subjects. J. Allergy Clin. Immunol. 2011, 127, 1612–1621.e8. [Google Scholar] [CrossRef] [PubMed]
- de Paulis, A.; Prevete, N.; Fiorentino, I.; Rossi, F.W.; Staibano, S.; Montuori, N.; Ragno, P.; Longobardi, A.; Liccardo, B.; Genovese, A.; et al. Expression and Functions of the Vascular Endothelial Growth Factors and Their Receptors in Human Basophils. J. Immunol. 2006, 177, 7322–7331. [Google Scholar] [CrossRef]
- Sumbayev, V.V.; Nicholas, S.A.; Streatfield, C.L.; Gibbs, B.F. Involvement of Hypoxia-Inducible Factor-1 HiF(1alpha) in IgE-Mediated Primary Human Basophil Responses. Eur. J. Immunol. 2009, 39, 3511–3519. [Google Scholar] [CrossRef]
- Corrigan, C.J.; Wang, W.; Meng, Q.; Fang, C.; Wu, H.; Reay, V.; Lv, Z.; Fan, Y.; An, Y.; Wang, Y.H.; et al. T-Helper Cell Type 2 (Th2) Memory T Cell-Potentiating Cytokine IL-25 Has the Potential to Promote Angiogenesis in Asthma. Proc. Natl. Acad. Sci. USA 2011, 108, 1579–1584. [Google Scholar] [CrossRef]
- Bucher, F.; Lee, J.; Shin, S.; Kim, M.S.; Oh, Y.S.; Ha, S.; Zhang, H.; Yea, K. Interleukin-5 Suppresses Vascular Endothelial Growth Factor-Induced Angiogenesis through STAT5 Signaling. Cytokine 2018, 110, 397–403. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Wang, L.; Chen, S.; Tian, B.; Huang, K.; Corrigan, C.J.; Ying, S.; Wang, W.; Wang, C. IL-33 Initiates Vascular Remodelling in Hypoxic Pulmonary Hypertension by up-Regulating HIF-1α and VEGF Expression in Vascular Endothelial Cells. EBioMedicine 2018, 33, 196–210. [Google Scholar] [CrossRef]
- Lee, C.G.; Link, H.; Baluk, P.; Homer, R.J.; Chapoval, S.; Bhandari, V.; Kang, M.J.; Cohn, L.; Kim, Y.K.; McDonald, D.M.; et al. Vascular Endothelial Growth Factor (VEGF) Induces Remodeling and Enhances TH2-Mediated Sensitization and Inflammation in the Lung. Nat. Med. 2004, 10, 1095–1103. [Google Scholar] [CrossRef]
- Meyer, N.; Akdis, C.A. Vascular Endothelial Growth Factor as a Key Inducer of Angiogenesis in the Asthmatic Airways. Curr. Allergy Asthma Rep. 2013, 13, 1–9. [Google Scholar] [CrossRef]
- Psarras, S.; Volonaki, E.; Skevaki, C.L.; Xatzipsalti, M.; Bossios, A.; Pratsinis, H.; Tsigkos, S.; Gourgiotis, D.; Constantopoulos, A.G.; Papapetropoulos, A.; et al. Vascular Endothelial Growth Factor-Mediated Induction of Angiogenesis by Human Rhinoviruses. J. Allergy Clin. Immunol. 2006, 117, 291–297. [Google Scholar] [CrossRef] [PubMed]
- De Silva, D.; Dagher, H.; Ghildyal, R.; Lindsay, M.; Li, X.; Freezer, N.J.; Wilson, J.W.; Bardin, P.G. Vascular Endothelial Growth Factor Induction by Rhinovirus Infection. J. Med. Virol. 2006, 78, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.Y.; Zhao, G.L.; Fu, M.F. Polymorphisms in the Vascular Endothelial Growth Factor (VEGF) Gene Associated with Asthma. Genet. Mol. Res. 2016, 15, 15027880. [Google Scholar] [CrossRef]
- Sharma, S.; Murphy, A.J.; Soto-Quiros, M.E.; Avila, L.; Klanderman, B.J.; Sylvia, J.S.; Celedón, J.C.; Raby, B.A.; Weiss, S.T. Association of VEGF Polymorphisms with Childhood Asthma, Lung Function and Airway Responsiveness. Eur. Respir. J. 2009, 33, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Simpson, A.; Custovic, A.; Tepper, R.; Graves, P.; Stern, D.A.; Jones, M.; Hankinson, J.; Curtin, J.A.; Wu, J.; Blekic, M.; et al. Genetic Variation in Vascular Endothelial Growth Factor-a and Lung Function. Am. J. Respir. Crit. Care Med. 2012, 185, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Balantic, M.; Rijavec, M.; Kavalar, M.S.; Suskovic, S.; Silar, M.; Kosnik, M.; Korosec, P. Asthma Treatment Outcome in Children Is Associated with Vascular Endothelial Growth Factor A (VEGFA) Polymorphisms. Mol. Diagn. Ther. 2012, 16, 173–180. [Google Scholar] [CrossRef]
- Boix, E.; Nogués, M.V. Mammalian Antimicrobial Proteins and Peptides: Overview on the RNase A Superfamily Members Involved in Innate Host Defence. Mol. Biosyst. 2007, 3, 317–335. [Google Scholar] [CrossRef]
- Rosenberg, H.F.; Ackerman, S.J.; Tenen, D.G. Human Eosinophil Cationic Protein. Molecular Cloning of a Cytotoxin and Helminthotoxin with Ribonuclease Activity. J. Exp. Med. 1989, 170, 163–176. [Google Scholar] [CrossRef]
- Rosenberg, H.F. RNase A Ribonucleases and Host Defense: An Evolving Story. J. Leukoc. Biol. 2008, 83, 1079–1087. [Google Scholar] [CrossRef]
- Hogan, S.P.; Rosenberg, H.F.; Moqbel, R.; Phipps, S.; Foster, P.S.; Lacy, P.; Kay, A.B.; Rothenberg, M.E. Eosinophils: Biological Properties and Role in Health and Disease. Clin. Exp. Allergy 2008, 38, 709–750. [Google Scholar] [CrossRef]
- Sur, S.; Glitz, D.G.; Kita, H.; Kujawa, S.M.; Peterson, E.A.; Weiler, D.A.; Kephart, G.M.; Wagner, J.M.; George, T.J.; Gleich, G.J.; et al. Localization of Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein in Neutrophilic Leukocytes. J. Leukoc. Biol. 1998, 63, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Rosenberg, H.F.; Chen, Q.; Dyer, K.D.; Kurosaka, K.; Oppenheim, J.J. Eosinophil-Derived Neurotoxin (EDN), an Antimicrobial Protein with Chemotactic Activities for Dendritic Cells. Blood 2003, 102, 3396–3403. [Google Scholar] [CrossRef] [PubMed]
- Domachowske, J.B.; Dyer, K.D.; Bonville, C.A.; Rosenberg, H.F. Recombinant Human Eosinophil-Derived Neurotoxin/RNase 2 Functions as an Effective Antiviral Agent against Respiratory Syncytial Virus. J. Infect. Dis. 1998, 177, 1458–1464. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Domachowske, J.B. Eosinophils, Eosinophil Ribonucleases, and Their Role in Host Defense against Respiratory Virus Pathogens. J. Leukoc. Biol. 2001, 70, 691–698. [Google Scholar] [CrossRef]
- Harrison, A.M.; Bonville, C.A.; Rosenberg, H.F.; Domachowske, J.B. Respiratory Syncytical Virus-Induced Chemokine Expression in the Lower Airways: Eosinophil Recruitment and Degranulation. Am. J. Respir. Crit. Care Med. 1999, 159, 1918–1924. [Google Scholar] [CrossRef]
- Dosanjh, A. Eosinophil-Derived Neurotoxin and Respiratory Tract Infection and Inflammation: Implications for COVID-19 Management. J. Interferon Cytokine Res. 2020, 40, 443–445. [Google Scholar] [CrossRef]
- Dill-McFarland, K.A.; Schwartz, J.T.; Zhao, H.; Shao, B.; Fulkerson, P.C.; Altman, M.C.; Gill, M.A. Eosinophil-Mediated Suppression and Anti-IL-5 Enhancement of Plasmacytoid Dendritic Cell Interferon Responses in Asthma. J. Allergy Clin. Immunol. 2022, 150, 666–675. [Google Scholar] [CrossRef]
- Phipps, S.; Howard, D.R.; Werder, R.B. Eosinophils Apply a Handbrake to Plasmacytoid Dendritic Cell Antiviral Immunity in Asthma. J. Allergy Clin. Immunol. 2022, 150, 589–591. [Google Scholar] [CrossRef]
- Rosenberg, H.F.; Dyer, K.D.; Foster, P.S. Eosinophils: Changing Perspectives in Health and Disease. Nat. Rev. Immunol. 2013, 13, 9–22. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Q.; Rosenberg, H.F.; Rybak, S.M.; Newton, D.L.; Wang, Z.Y.; Fu, Q.; Tchernev, V.T.; Wang, M.; Schweitzer, B.; et al. Human Ribonuclease A Superfamily Members, Eosinophil-Derived Neurotoxin and Pancreatic Ribonuclease, Induce Dendritic Cell Maturation and Activation. J. Immunol. 2004, 173, 6134–6142. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Q.; Shao, B.S.; Zhang, P.; Kurosaka, K.; Caspi, R.R.; Michalek, S.M.; Rosenberg, H.F.; Zhang, N.; Oppenheim, J.J. Eosinophil-Derived Neurotoxin Acts as an Alarmin to Activate the TLR2-MyD88 Signal Pathway in Dendritic Cells and Enhances Th2 Immune Responses. J. Exp. Med. 2008, 205, 79–90. [Google Scholar] [CrossRef]
- Oppenheim, J.J.; Yang, D. Alarmins: Chemotactic Activators of Immune Responses. Curr. Opin. Immunol. 2005, 17, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Han, Z.; Oppenheim, J.J. Alarmins and Immunity. Immunol. Rev. 2017, 280, 41–56. [Google Scholar] [CrossRef]
- Kim, C.K.; Callaway, Z.; Park, J.S.; Kwon, E. Utility of Serum Eosinophil-Derived Neurotoxin (EDN) Measurement by ELISA in Young Children with Asthma. Allergol. Int. 2017, 66, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Venge, P. Monitoring the Allergic Inflammation. Allergy 2004, 59, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Quoc, Q.L.; Moon, J.Y.; Lee, D.H.; Ban, G.Y.; Kim, S.H.; Park, H.S. Role of Thymus and Activation-Regulated Chemokine in Allergic Asthma. J. Asthma Allergy 2022, 15, 157–167. [Google Scholar] [CrossRef]
- Yancey, S.W.; Keene, O.N.; Albers, F.C.; Ortega, H.; Bates, S.; Bleecker, E.R.; Pavord, I. Biomarkers for Severe Eosinophilic Asthma. J. Allergy Clin. Immunol. 2017, 140, 1509–1518. [Google Scholar] [CrossRef]
- Nelson, R.K.; Bush, A.; Stokes, J.; Nair, P.; Akuthota, P. Eosinophilic Asthma. J. allergy Clin. Immunol. Pract. 2020, 8, 465–473. [Google Scholar] [CrossRef]
- Crimi, E.; Spanevello, A.; Neri, M.; Ind, P.W.; Rossi, G.A.; Brusasco, V. Dissociation between Airway Inflammation and Airway Hyperresponsiveness in Allergic Asthma. Am. J. Respir. Crit. Care Med. 1998, 157, 4–9. [Google Scholar] [CrossRef]
- Jatakanon, A.; Lim, S.; Barnes, P.J. Changes in Sputum Eosinophils Predict Loss of Asthma Control. Am. J. Respir. Crit. Care Med. 2000, 161, 64–72. [Google Scholar] [CrossRef]
- An, J.; Lee, J.H.; Sim, J.H.; Song, W.J.; Kwon, H.S.; Cho, Y.S.; Moon, H.B.; Kim, C.K.; Kim, T.B. Serum Eosinophil-Derived Neurotoxin Better Reflect Asthma Control Status Than Blood Eosinophil Counts. J. Allergy Clin. Immunol. Pract. 2020, 8, 2681–2688.e1. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.K.; Kita, H.; Callaway, Z.; Kim, H.B.; Choi, J.; Fujisawa, T.; Shin, B.M.; Koh, Y.Y. The Roles of a Th2 Cytokine and CC Chemokine in Children with Stable Asthma: Potential Implication in Eosinophil Degranulation. Pediatr. Allergy Immunol. 2010, 21 Pt 2, e697–e704. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.K.; Callaway, Z.; Kim, D.W.; Kita, H. Eosinophil Degranulation Is More Important than Eosinophilia in Identifying Asthma in Chronic Cough. J. Asthma 2011, 48, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lee, J.H.; Yang, E.M.; Kwon, E.M.; Jung, C.G.; Kim, S.C.; Choi, Y.; Cho, Y.S.; Kim, C.K.; Park, H.S. Serum Levels of Eosinophil-Derived Neurotoxin: A Biomarker for Asthma Severity in Adult Asthmatics. Allergy Asthma Immunol. Res. 2019, 11, 394–405. [Google Scholar] [CrossRef]
- Soo Kim, H.; Yang, H.J.; Song, D.J.; Ju Lee, Y.; In Suh, D.; Yeon Shim, J.; Yoo, Y.; Keun Kim, C.; Min Ahn, Y.; Tack Kim, J. Eosinophil-Derived Neurotoxin: An Asthma Exacerbation Biomarker in Children. Allergy Asthma Proc. 2022, 43, 133–139. [Google Scholar] [CrossRef]
- Dharmage, S.C.; Perret, J.L.; Custovic, A. Epidemiology of Asthma in Children and Adults. Front. Pediatr. 2019, 7, 246. [Google Scholar] [CrossRef]
- Kim, C.K.; Seo, J.K.; Ban, S.H.; Fujisawa, T.; Kim, D.W.; Callaway, Z. Eosinophil-Derived Neurotoxin Levels at 3 Months Post-Respiratory Syncytial Virus Bronchiolitis Are a Predictive Biomarker of Recurrent Wheezing. Biomarkers 2013, 18, 230–235. [Google Scholar] [CrossRef]
- Granger, V.; Zerimech, F.; Arab, J.; Siroux, V.; De Nadai, P.; Tsicopoulos, A.; Matran, R.; Akiki, Z.; Nadif, R. Blood Eosinophil Cationic Protein and Eosinophil-Derived Neurotoxin Are Associated with Different Asthma Expression and Evolution in Adults. Thorax 2022, 77, 552–562. [Google Scholar] [CrossRef]
- Gon, Y.; Ito, R.; Hattori, T.; Hiranuma, H.; Kumasawa, F.; Kozu, Y.; Endo, D.; Koyama, D.; Shintani, Y.; Eriko, T.; et al. Serum Eosinophil-Derived Neurotoxin: Correlation with Persistent Airflow Limitation in Adults with House-Dust Mite Allergic Asthma. Allergy Asthma Proc. 2015, 36, e113–e120. [Google Scholar] [CrossRef]
- Bellin, M.H.; Osteen, P.; Kub, J.; Bollinger, M.E.; Tsoukleris, M.; Chaikind, L.; Butz, A.M. Stress and Quality of Life in Urban Caregivers of Children With Poorly Controlled Asthma: A Longitudinal Analysis. J. Pediatr. Health Care 2015, 29, 536–546. [Google Scholar] [CrossRef]
- Hoch, H.E.; Houin, P.R.; Stillwell, P.C. Asthma in Children: A Brief Review for Primary Care Providers. Pediatr. Ann. 2019, 48, e103–e109. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, P.R.; Liu, A.H.; Borres, M.P.; Södergren, E.; Iachetti, F.; Casale, T.B. Asthma and Allergy: Unravelling a Tangled Relationship with a Focus on New Biomarkers and Treatment. Int. J. Mol. Sci. 2022, 23, 3881. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.K.; Callaway, Z.; Park, J.S.; Nishimori, H.; Ogino, T.; Nagao, M.; Fujisawa, T. Montelukast Reduces Serum Levels of Eosinophil-Derived Neurotoxin in Preschool Asthma. Allergy Asthma Immunol. Res. 2018, 10, 686–697. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W.; Lee, K.E.; Kim, E.S.; Song, T.W.; Sohn, M.H.; Kim, K.E. Serum Eosinophil-Derived Neurotoxin (EDN) in Diagnosis and Evaluation of Severity and Bronchial Hyperresponsiveness in Childhood Asthma. Lung 2007, 185, 97–103. [Google Scholar] [CrossRef]
- Hirst, S.J.; Lee, T.H. Airway Smooth Muscle as a Target of Glucocorticoid Action in the Treatment of Asthma. Am. J. Respir. Crit. Care Med. 1998, 158 Pt 3, S201–S206. [Google Scholar] [CrossRef]
- Ora, J.; Calzetta, L.; Matera, M.G.; Cazzola, M.; Rogliani, P. Advances with Glucocorticoids in the Treatment of Asthma: State of the Art. Expert Opin. Pharmacother. 2020, 21, 2305–2316. [Google Scholar] [CrossRef]
- Wen, F.Q.; Liu, X.; Manda, W.; Terasaki, Y.; Kobayashi, T.; Abe, S.; Fang, Q.; Ertl, R.; Manouilova, L.; Rennard, S.I. TH2 Cytokine-Enhanced and TGF-β-Enhanced Vascular Endothelial Growth Factor Production by Cultured Human Airway Smooth Muscle Cells Is Attenuated by IFN-γ and Corticosteroids. J. Allergy Clin. Immunol. 2003, 111, 1307–1318. [Google Scholar] [CrossRef]
- Skevaki, C.L.; Christodoulou, I.; Spyridaki, I.S.; Tiniakou, I.; Georgiou, V.; Xepapadaki, P.; Kafetzis, D.A.; Papadopoulos, N.G. Budesonide and Formoterol Inhibit Inflammatory Mediator Production by Bronchial Epithelial Cells Infected with Rhinovirus. Clin. Exp. Allergy 2009, 39, 1700–1710. [Google Scholar] [CrossRef]
- Hamamoto, Y.; Ano, S.; Allard, B.; O’Sullivan, M.; McGovern, T.K.; Martin, J.G. Montelukast Reduces Inhaled Chlorine Triggered Airway Hyperresponsiveness and Airway Inflammation in the Mouse. Br. J. Pharmacol. 2017, 174, 3346–3358. [Google Scholar] [CrossRef]
- Türkeli, A.; Yilmaz, Ö.; Karaman, M.; Kanik, E.; Firinci, F.; İnan, S.; Yüksel, H. Anti-VEGF Treatment Suppresses Remodeling Factors and Restores Epithelial Barrier Function through the E-Cadherin/β-Catenin Signaling Axis in Experimental Asthma Models. Exp. Ther. Med. 2021, 22, 689. [Google Scholar] [CrossRef]
- Ge, A.; Liu, Y.; Zeng, X.; Kong, H.; Ma, Y.; Zhang, J.; Bai, F.; Huang, M. Effect of Diosmetin on Airway Remodeling in a Murine Model of Chronic Asthma. Acta Biochim. Biophys. Sin. 2015, 47, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Dong, H.; Zou, M.; Luo, L.; Hu, Y.; Xie, Z.; Le, Y.; Liu, L.; Zou, F.; Cai, S. Bevacizumab Reduced Auto-Phosphorylation of VEGFR2 to Protect HDM-Induced Asthma Mice. Biochem. Biophys. Res. Commun. 2016, 478, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Bolandi, S.M.; Abdolmaleki, Z.; Assarehzadegan, M.A. Anti-Angiogenic Properties of Bevacizumab Improve Respiratory System Inflammation in Ovalbumin-Induced Rat Model of Asthma. Inflammation 2021, 44, 2463–2475. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Park, J.W.; Cha, H.R.; Jung, S.Y.; Lee, J.E.; Jung, S.S.; Kim, J.O.; Kim, S.Y.; Lee, C.S.; Park, H.S. Silver Nanoparticles Modify VEGF Signaling Pathway and Mucus Hypersecretion in Allergic Airway Inflammation. Int. J. Nanomed. 2012, 7, 1329–1343. [Google Scholar] [CrossRef]
- Kalishwaralal, K.; Banumathi, E.; Pandian, S.B.R.K.; Deepak, V.; Muniyandi, J.; Eom, S.H.; Gurunathan, S. Silver Nanoparticles Inhibit VEGF Induced Cell Proliferation and Migration in Bovine Retinal Endothelial Cells. Colloids Surf. B. Biointerfaces 2009, 73, 51–57. [Google Scholar] [CrossRef]
- Zha, W.; Su, M.; Huang, M.; Cai, J.; Du, Q. Administration of Pigment Epithelium-Derived Factor Inhibits Airway Inflammation and Remodeling in Chronic OVA-Induced Mice via VEGF Suppression. Allergy Asthma Immunol. Res. 2016, 8, 161–169. [Google Scholar] [CrossRef]
- Kim, S.R.; Lee, K.S.; Lee, K.B.; Lee, Y.C. Recombinant IGFBP-3 Inhibits Allergic Lung Inflammation, VEGF Production, and Vascular Leak in a Mouse Model of Asthma. Allergy 2012, 67, 869–877. [Google Scholar] [CrossRef]
- Yang, N.; Li, X. Epigallocatechin Gallate Relieves Asthmatic Symptoms in Mice by Suppressing HIF-1α/VEGFA-Mediated M2 Skewing of Macrophages. Biochem. Pharmacol. 2022, 202, 115112. [Google Scholar] [CrossRef]
- Li, Y.Z.; Chen, L.X.; Guo, F.F.; Cao, Y.; Hu, W.; Shi, Y.; Lin, X.C.; Hou, J.; Li, L.P.; Ding, X.F.; et al. Effects of Epigallocatechin-3-Gallate on the HMGB1/RAGE Pathway in PM2.5-Exposed Asthmatic Rats. Biochem. Biophys. Res. Commun. 2019, 513, 898–903. [Google Scholar] [CrossRef]
- Zhao, X.; Yu, F.Q.; Huang, X.J.; Xu, B.Y.; Li, Y.L.; Zhao, X.Y.; Guo, H.F.; Luan, B. Azithromycin Influences Airway Remodeling in Asthma via the PI3K/Akt/MTOR/HIF-1α/VEGF Pathway. J. Biol. Regul. Homeost. Agents 2018, 32, 1079–1088. [Google Scholar]
- Willems-Widyastuti, A.; Vanaudenaerde, B.M.; Vos, R.; Dilisen, E.; Verleden, S.E.; De Vleeschauwer, S.I.; Vaneylen, A.; Mooi, W.J.; de Boer, W.I.; Sharma, H.S.; et al. Azithromycin Attenuates Fibroblast Growth Factors Induced Vascular Endothelial Growth Factor via P38(MAPK) Signaling in Human Airway Smooth Muscle Cells. Cell Biochem. Biophys. 2013, 67, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.G.; Yang, I.A.; Upham, J.W.; Reynolds, P.N.; Hodge, S.; James, A.L.; Jenkins, C.; Peters, M.J.; Marks, G.B.; Baraket, M.; et al. Efficacy of Azithromycin in Severe Asthma from the AMAZES Randomised Trial. ERJ Open Res. 2019, 5, 00056–02019. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, J.J.; Jat, K.R.; Sankar, J.; Lodha, R.; Iyer, V.K.; Gautam, H.; Sood, S.; Kabra, S.K. Azithromycin for Poorly Controlled Asthma in Children: A Randomized Controlled Trial. Chest 2022, 161, 1456–1464. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Pei, Q.M.; Jiang, P.; Yang, M.; Qian, X.J.; Liu, J.B. Effect of Active Vitamin D3 on VEGF-Induced ADAM33 Expression and Proliferation in Human Airway Smooth Muscle Cells: Implications for Asthma Treatment. Respir. Res. 2017, 18, 7. [Google Scholar] [CrossRef]
- Lee, S.Y.; Chung, S.M. Neovastat (AE-941) Inhibits the Airway Inflammation via VEGF and HIF-2 Alpha Suppression. Vascul. Pharmacol. 2007, 47, 313–318. [Google Scholar] [CrossRef]
- Suzaki, Y.; Hamada, K.; Sho, M.; Ito, T.; Miyamoto, K.; Akashi, S.; Kashizuka, H.; Ikeda, N.; Nakajima, Y.; Iwase, M.; et al. A Potent Antiangiogenic Factor, Endostatin Prevents the Development of Asthma in a Murine Model. J. Allergy Clin. Immunol. 2005, 116, 1220–1227. [Google Scholar] [CrossRef]
- Yu, Q.; Yu, X.; Zhong, X.; Ma, Y.; Wu, Y.; Bian, T.; Huang, M.; Zeng, X. Melatonin Modulates Airway Smooth Muscle Cell Phenotype by Targeting the STAT3/Akt/GSK-3β Pathway in Experimental Asthma. Cell Tissue Res. 2020, 380, 129–142. [Google Scholar] [CrossRef]
- Kips, J.C.; O’Connor, B.J.; Inman, M.D.; Svensson, K.; Pauwels, R.A.; O’Byrne, P.M. A Long-Term Study of the Antiinflammatory Effect of Low-Dose Budesonide plus Formoterol versus High-Dose Budesonide in Asthma. Am. J. Respir. Crit. Care Med. 2000, 161 Pt 1, 996–1001. [Google Scholar] [CrossRef]
- Nicholas, T.P.; Haick, A.K.; Bammler, T.K.; Workman, T.W.; Kavanagh, T.J.; Faustman, E.M.; Gharib, S.A.; Altemeier, W.A. The Effects of Genotype × Phenotype Interactions on Transcriptional Response to Silver Nanoparticle Toxicity in Organotypic Cultures of Murine Tracheal Epithelial Cells. Toxicol. Sci. 2020, 173, 131–143. [Google Scholar] [CrossRef]
- Taniguchi, H.; Tsuchida, T.; Nakamura, Y.; Motoshima, K.; Mizoguchi, K.; Kohno, S. Severe, but Manageable Hypoxia Caused by Bronchospasm Induced by Bevacizumab. Respirol. Case Rep. 2013, 1, 14–16. [Google Scholar] [CrossRef]
- Parnham, M.J.; Haber, V.E.; Giamarellos-Bourboulis, E.J.; Perletti, G.; Verleden, G.M.; Vos, R. Azithromycin: Mechanisms of Action and Their Relevance for Clinical Applications. Pharmacol. Ther. 2014, 143, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.G.; Yang, I.A.; Upham, J.W.; Reynolds, P.N.; Hodge, S.; James, A.L.; Jenkins, C.; Peters, M.J.; Marks, G.B.; Baraket, M.; et al. Effect of Azithromycin on Asthma Exacerbations and Quality of Life in Adults with Persistent Uncontrolled Asthma (AMAZES): A Randomised, Double-Blind, Placebo-Controlled Trial. Lancet 2017, 390, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Ge, A.; Ma, Y.; Liu, Y.N.; Li, Y.S.; Gu, H.; Zhang, J.X.; Wang, Q.X.; Zeng, X.N.; Huang, M. Diosmetin Prevents TGF-Β1-Induced Epithelial-Mesenchymal Transition via ROS/MAPK Signaling Pathways. Life Sci. 2016, 153, 1–8. [Google Scholar] [CrossRef]
- Villa, P.; Cova, D.; De Francesco, L.; Guaitani, A.; Palladini, G.; Perego, R. Protective Effect of Diosmetin on in Vitro Cell Membrane Damage and Oxidative Stress in Cultured Rat Hepatocytes. Toxicology 1992, 73, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Forno, E.; Bacharier, L.B.; Phipatanakul, W.; Guilbert, T.W.; Cabana, M.D.; Ross, K.; Covar, R.; Gern, J.E.; Rosser, F.J.; Blatter, J.; et al. Effect of Vitamin D3 Supplementation on Severe Asthma Exacerbations in Children With Asthma and Low Vitamin D Levels: The VDKA Randomized Clinical Trial. JAMA 2020, 324, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; King, T.S.; McKenzie, M.; Bacharier, L.B.; Dixon, A.E.; Codispoti, C.D.; Dunn, R.M.; Grossman, N.L.; Lugogo, N.L.; Ramratnam, S.K.; et al. Vitamin D3 Therapy in Patients with Asthma Complicated by Sinonasal Disease: Secondary Analysis of the Vitamin D Add-on Therapy Enhances Corticosteroid Responsiveness in Asthma Trial. J. Allergy Clin. Immunol. 2016, 138, 589–592.e2. [Google Scholar] [CrossRef]
- Shan, L.; Kang, X.; Liu, F.; Cai, X.; Han, X.; Shang, Y. Epigallocatechin Gallate Improves Airway Inflammation through TGF-β1 Signaling Pathway in Asthmatic Mice. Mol. Med. Rep. 2018, 18, 2088–2096. [Google Scholar] [CrossRef]
- Yang, N.; Zhang, H.; Cai, X.; Shang, Y. Epigallocatechin-3-Gallate Inhibits Inflammation and Epithelial-mesenchymal Transition through the PI3K/AKT Pathway via Upregulation of PTEN in Asthma. Int. J. Mol. Med. 2018, 41, 818–828. [Google Scholar] [CrossRef]
- Lee, H.; Kim, S.R.; Oh, Y.; Cho, S.H.; Schleimer, R.P.; Lee, Y.C. Targeting Insulin-like Growth Factor-I and Insulin-like Growth Factor-Binding Protein-3 Signaling Pathways. A Novel Therapeutic Approach for Asthma. Am. J. Respir. Cell Mol. Biol. 2014, 50, 667–677. [Google Scholar] [CrossRef]
- Lee, S.Y.; Paik, S.Y.; Chung, S.M. Neovastat (AE-941) Inhibits the Airway Inflammation and Hyperresponsiveness in a Murine Model of Asthma. J. Microbiol. 2005, 43, 11–16. [Google Scholar] [CrossRef]
- Shin, I.S.; Jeon, W.Y.; Shin, H.K.; Lee, M.Y. Effects of Montelukast on Subepithelial/Peribronchial Fibrosis in a Murine Model of Ovalbumin Induced Chronic Asthma. Int. Immunopharmacol. 2013, 17, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Calapai, G.; Casciaro, M.; Miroddi, M.; Calapai, F.; Navarra, M.; Gangemi, S. Montelukast-Induced Adverse Drug Reactions: A Review of Case Reports in the Literature. Pharmacology 2014, 94, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Ek, E.T.H.; Dass, C.R.; Choong, P.F.M. PEDF: A Potential Molecular Therapeutic Target with Multiple Anti-Cancer Activities. Trends Mol. Med. 2006, 12, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, X.; Yang, X.; Liu, X.; Zeng, Y.; Li, J. Melatonin Antagonizes Ozone-Exacerbated Asthma by Inhibiting the TRPV1 Channel and Stabilizing the Nrf2 Pathway. Environ. Sci. Pollut. Res. Int. 2021, 28, 59858–59867. [Google Scholar] [CrossRef]
- Shin, I.S.; Park, J.W.; Shin, N.R.; Jeon, C.M.; Kwon, O.K.; Lee, M.Y.; Kim, H.S.; Kim, J.C.; Oh, S.R.; Ahn, K.S. Melatonin Inhibits MUC5AC Production via Suppression of MAPK Signaling in Human Airway Epithelial Cells. J. Pineal Res. 2014, 56, 398–407. [Google Scholar] [CrossRef]
- Bel, E.H.; ten Brinke, A. New Anti-Eosinophil Drugs for Asthma and COPD: Targeting the Trait! Chest 2017, 152, 1276–1282. [Google Scholar] [CrossRef]
- Jang, J.H.; Woo, S.D.; Lee, Y.; Kim, C.K.; Shin, Y.S.; Ye, Y.M.; Park, H.S. Changes in Type 2 Biomarkers After Anti-IL5 Treatment in Patients With Severe Eosinophilic Asthma. Allergy Asthma Immunol. Res. 2021, 13, 330–338. [Google Scholar] [CrossRef]
- Nair, P.; Ochkur, S.I.; Protheroe, C.; Radford, K.; Efthimiadis, A.; Lee, N.A.; Lee, J.J. Eosinophil Peroxidase in Sputum Represents a Unique Biomarker of Airway Eosinophilia. Allergy 2013, 68, 1177–1184. [Google Scholar] [CrossRef]
- Rutten, B.; Young, S.; Rhedin, M.; Olsson, M.; Kurian, N.; Syed, F.; Beech, A.; Fidock, M.; Newbold, P.; Singh, D.; et al. Eosinophil-Derived Neurotoxin: A Biologically and Analytically Attractive Asthma Biomarker. PLoS ONE 2021, 16, e0246627. [Google Scholar] [CrossRef]
- Agache, I.; Beltran, J.; Akdis, C.; Akdis, M.; Canelo-Aybar, C.; Canonica, G.W.; Casale, T.; Chivato, T.; Corren, J.; Del Giacco, S.; et al. Efficacy and Safety of Treatment with Biologicals (Benralizumab, Dupilumab, Mepolizumab, Omalizumab and Reslizumab) for Severe Eosinophilic Asthma. A Systematic Review for the EAACI Guidelines—Recommendations on the Use of Biologicals in Severe Asthma. Allergy 2020, 75, 1023–1042. [Google Scholar] [CrossRef]
- Pham, T.H.; Damera, G.; Newbold, P.; Ranade, K. Reductions in Eosinophil Biomarkers by Benralizumab in Patients with Asthma. Respir. Med. 2016, 111, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Haktanir Abul, M.; Phipatanakul, W. Severe Asthma in Children: Evaluation and Management. Allergol. Int. 2019, 68, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.A.B.; Busse, W.W.; Jarjour, N.N. Inhaled Budesonide Decreases Airway Inflammatory Response to Allergen. Am. J. Respir. Crit. Care Med. 2000, 162 Pt 1, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.; Dahl, R.; Karlström, R.; Peterson, C.G.B.; Venge, P. Eosinophil and Neutrophil Activity in Asthma in a One-Year Trial with Inhaled Budesonide. The Impact of Smoking. Am. J. Respir. Crit. Care Med. 1996, 153, 1519–1529. [Google Scholar] [CrossRef]
- Chipps, B.E.; Albers, F.C.; Reilly, L.; Johnsson, E.; Cappelletti, C.; Papi, A. Efficacy and Safety of As-Needed Albuterol/Budesonide versus Albuterol in Adults and Children Aged ≥4 Years with Moderate-to-Severe Asthma: Rationale and Design of the Randomised, Double-Blind, Active-Controlled MANDALA Study. BMJ Open Respir. Res. 2021, 8, e001077. [Google Scholar] [CrossRef]
- Kavanagh, J.E.; Hearn, A.P.; Dhariwal, J.; d’Ancona, G.; Douiri, A.; Roxas, C.; Fernandes, M.; Green, L.; Thomson, L.; Nanzer, A.M.; et al. Real-World Effectiveness of Benralizumab in Severe Eosinophilic Asthma. Chest 2021, 159, 496–506. [Google Scholar] [CrossRef]
- Makiya, M.A.; Khoury, P.; Kuang, F.L.; Mata, A.D.; Mahmood, S.; Bowman, A.; Espinoza, D.; Kovacs, N.; Brown, T.; Holland, N.; et al. Urine Eosinophil-Derived Neurotoxin: A Potential Marker of Activity in Select Eosinophilic Disorders. Allergy 2023, 78, 258–269. [Google Scholar] [CrossRef]
- Guntur, V.P.; Manka, L.A.; Denson, J.L.; Dunn, R.M.; Dollin, Y.T.; Gill, M.; Kolakowski, C.; Strand, M.J.; Wechsler, M.E. Benralizumab as a Steroid-Sparing Treatment Option in Eosinophilic Granulomatosis with Polyangiitis. J. Allergy Clin. Immunol. Pract. 2021, 9, 1186–1193.e1. [Google Scholar] [CrossRef]
- Korn, S.; Bourdin, A.; Chupp, G.; Cosio, B.G.; Arbetter, D.; Shah, M.; Gil, E.G. Integrated Safety and Efficacy Among Patients Receiving Benralizumab for Up to 5 Years. J. Allergy Clin. Immunol. Pract. 2021, 9, 4381–4392.e4. [Google Scholar] [CrossRef]
- Kwah, J.H.; Peters, A.T. Asthma in Adults: Principles of Treatment. Allergy Asthma Proc. 2019, 40, 396–402. [Google Scholar] [CrossRef]
- Charles, D.; Shanley, J.; Temple, S.N.; Rattu, A.; Khaleva, E.; Roberts, G. Real-World Efficacy of Treatment with Benralizumab, Dupilumab, Mepolizumab and Reslizumab for Severe Asthma: A Systematic Review and Meta-Analysis. Clin. Exp. Allergy 2022, 52, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Chapman, K.R.; Albers, F.C.; Chipps, B.; Muñoz, X.; Devouassoux, G.; Bergna, M.; Galkin, D.; Azmi, J.; Mouneimne, D.; Price, R.G.; et al. The Clinical Benefit of Mepolizumab Replacing Omalizumab in Uncontrolled Severe Eosinophilic Asthma. Allergy 2019, 74, 1716–1726. [Google Scholar] [CrossRef] [PubMed]
- Ortega, H.G.; Liu, M.C.; Pavord, I.D.; Brusselle, G.G.; FitzGerald, J.M.; Chetta, A.; Humbert, M.; Katz, L.E.; Keene, O.N.; Yancey, S.W.; et al. Mepolizumab Treatment in Patients with Severe Eosinophilic Asthma. N. Engl. J. Med. 2014, 371, 1198–1207. [Google Scholar] [CrossRef] [PubMed]
- Harvey, E.S.; Langton, D.; Katelaris, C.; Stevens, S.; Farah, C.S.; Gillman, A.; Harrington, J.; Hew, M.; Kritikos, V.; Radhakrishna, N.; et al. Mepolizumab Effectiveness and Identification of Super-Responders in Severe Asthma. Eur. Respir. J. 2020, 55, 1902420. [Google Scholar] [CrossRef]
- Pavord, I.D.; Korn, S.; Howarth, P.; Bleecker, E.R.; Buhl, R.; Keene, O.N.; Ortega, H.; Chanez, P. Mepolizumab for Severe Eosinophilic Asthma (DREAM): A Multicentre, Double-Blind, Placebo-Controlled Trial. Lancet 2012, 380, 651–659. [Google Scholar] [CrossRef]
- Pérez de Llano, L.A.; Cosío, B.G.; Lobato Astiárraga, I.; Soto Campos, G.; Tejedor Alonso, M.Á.; Marina Malanda, N.; Padilla Galo, A.; Urrutia Landa, I.; Michel de la Rosa, F.J.; García-Moguel, I. Asthma Control in Patients with Severe Eosinophilic Asthma Treated with Reslizumab: Spanish Real-Life Data. J. Asthma Allergy 2022, 15, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.; Zangrilli, J.; Wechsler, M.E.; Bateman, E.D.; Brusselle, G.G.; Bardin, P.; Murphy, K.; Maspero, J.F.; O’Brien, C.; Korn, S. Reslizumab for Inadequately Controlled Asthma with Elevated Blood Eosinophil Counts: Results from Two Multicentre, Parallel, Double-Blind, Randomised, Placebo-Controlled, Phase 3 Trials. Lancet. Respir. Med. 2015, 3, 355–366. [Google Scholar] [CrossRef]
- Wechsler, M.E.; Ford, L.B.; Maspero, J.F.; Pavord, I.D.; Papi, A.; Bourdin, A.; Watz, H.; Castro, M.; Nenasheva, N.M.; Tohda, Y.; et al. Long-Term Safety and Efficacy of Dupilumab in Patients with Moderate-to-Severe Asthma (TRAVERSE): An Open-Label Extension Study. Lancet. Respir. Med. 2022, 10, 11–25. [Google Scholar] [CrossRef]
- Bacharier, L.B.; Maspero, J.F.; Katelaris, C.H.; Fiocchi, A.G.; Gagnon, R.; de Mir, I.; Jain, N.; Sher, L.D.; Mao, X.; Liu, D.; et al. Dupilumab in Children with Uncontrolled Moderate-to-Severe Asthma. N. Engl. J. Med. 2021, 385, 2230–2240. [Google Scholar] [CrossRef]
- Dupin, C.; Belhadi, D.; Guilleminault, L.; Gamez, A.S.; Berger, P.; De Blay, F.; Bonniaud, P.; Leroyer, C.; Mahay, G.; Girodet, P.O.; et al. Effectiveness and Safety of Dupilumab for the Treatment of Severe Asthma in a Real-Life French Multi-Centre Adult Cohort. Clin. Exp. Allergy 2020, 50, 789–798. [Google Scholar] [CrossRef]
- Harb, H.; Chatila, T.A. Mechanisms of Dupilumab. Clin. Exp. Allergy 2020, 50, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Szefler, S.J.; Roberts, G.; Rubin, A.S.; Zielen, S.; Kuna, P.; Alpan, O.; Anzures-Cabrera, J.; Chen, Q.; Holweg, C.T.J.; Kaminski, J.; et al. Efficacy, Safety, and Tolerability of Lebrikizumab in Adolescent Patients with Uncontrolled Asthma (ACOUSTICS). Clin. Transl. Allergy 2022, 12, e12176. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.D.; Gonzalez Edick, M.; Ferrando, R.E.; Solon, M.; Baca, M.; Mesh, K.; Bradding, P.; Gauvreau, G.M.; Sumino, K.; FitzGerald, J.M.; et al. A Randomized, Placebo-Controlled Trial Evaluating Effects of Lebrikizumab on Airway Eosinophilic Inflammation and Remodelling in Uncontrolled Asthma (CLAVIER). Clin. Exp. Allergy 2020, 50, 1342–1351. [Google Scholar] [CrossRef] [PubMed]
- Hanania, N.A.; Korenblat, P.; Chapman, K.R.; Bateman, E.D.; Kopecky, P.; Paggiaro, P.; Yokoyama, A.; Olsson, J.; Gray, S.; Holweg, C.T.J.; et al. Efficacy and Safety of Lebrikizumab in Patients with Uncontrolled Asthma (LAVOLTA I and LAVOLTA II): Replicate, Phase 3, Randomised, Double-Blind, Placebo-Controlled Trials. Lancet. Respir. Med. 2016, 4, 781–796. [Google Scholar] [CrossRef]
- Hanania, N.A.; Noonan, M.; Corren, J.; Korenblat, P.; Zheng, Y.; Fischer, S.K.; Cheu, M.; Putnam, W.S.; Murray, E.; Scheerens, H.; et al. Lebrikizumab in Moderate-to-Severe Asthma: Pooled Data from Two Randomised Placebo-Controlled Studies. Thorax 2015, 70, 748–756. [Google Scholar] [CrossRef]
- Kardas, G.; Panek, M.; Kuna, P.; Damiański, P.; Kupczyk, M. Monoclonal Antibodies in the Management of Asthma: Dead Ends, Current Status and Future Perspectives. Front. Immunol. 2022, 13, 983852. [Google Scholar] [CrossRef]
- Edris, A.; De Feyter, S.; Maes, T.; Joos, G.; Lahousse, L. Monoclonal Antibodies in Type 2 Asthma: A Systematic Review and Network Meta-Analysis. Respir. Res. 2019, 20, 179. [Google Scholar] [CrossRef]
- Piper, E.; Brightling, C.; Niven, R.; Oh, C.; Faggioni, R.; Poon, K.; She, D.; Kell, C.; May, R.D.; Geba, G.P.; et al. A Phase II Placebo-Controlled Study of Tralokinumab in Moderate-to-Severe Asthma. Eur. Respir. J. 2013, 41, 330–338. [Google Scholar] [CrossRef]
- Gon, Y.; Maruoka, S.; Mizumura, K. Omalizumab and IgE in the Control of Severe Allergic Asthma. Front. Pharmacol. 2022, 13, 839011. [Google Scholar] [CrossRef]
- Kotoulas, S.C.; Tsiouprou, I.; Fouka, E.; Pataka, A.; Papakosta, D.; Porpodis, K. Omalizumab: An Optimal Choice for Patients with Severe Allergic Asthma. J. Pers. Med. 2022, 12, 165. [Google Scholar] [CrossRef]
- Hanania, N.A.; Alpan, O.; Hamilos, D.L.; Condemi, J.J.; Reyes-Rivera, I.; Zhu, J.; Rosen, K.E.; Eisner, M.D.; Wong, D.A.; Busse, W. Omalizumab in Severe Allergic Asthma Inadequately Controlled with Standard Therapy: A Randomized Trial. Ann. Intern. Med. 2011, 154, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Dorey-Stein, Z.L.; Shenoy, K.V. Tezepelumab as an Emerging Therapeutic Option for the Treatment of Severe Asthma: Evidence to Date. Drug Des. Devel. Ther. 2021, 15, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Menzies-Gow, A.; Corren, J.; Bourdin, A.; Chupp, G.; Israel, E.; Wechsler, M.E.; Brightling, C.E.; Griffiths, J.M.; Hellqvist, Å.; Bowen, K.; et al. Tezepelumab in Adults and Adolescents with Severe, Uncontrolled Asthma. N. Engl. J. Med. 2021, 384, 1800–1809. [Google Scholar] [CrossRef] [PubMed]
- Jahangir, A.; Sattar, S.B.A.; Niazi, M.R.K.; Muhammad, M.; Jahangir, A.; Sahra, S.; Sharif, M.A.; Anwar, M.Y.; Chalhoub, M. Efficacy and Safety of Fevipiprant in Asthma: A Review and Meta-Analysis. Cureus 2022, 14, e24641. [Google Scholar] [CrossRef]
- Wojtukiewicz, M.Z.; Mysliwiec, M.; Sierko, E.; Sobierska, M.; Kruszewska, J.; Lipska, A.; Radziwon, P.; Tucker, S.C.; Honn, K.V. Elevated Microparticles, Thrombin-Antithrombin and VEGF Levels in Colorectal Cancer Patients Undergoing Chemotherapy. Pathol. Oncol. Res. 2020, 26, 2499–2507. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Wang, H.; Ye, Q. Increased Circulating Vascular Endothelial Growth Factor in Acute Myeloid Leukemia Patients: A Systematic Review and Meta-Analysis. Syst. Rev. 2020, 9, 103. [Google Scholar] [CrossRef]
- Filipiak, J.; Boinska, J.; Ziołkowska, K.; Zduńska, M.; Zarychta, E.; Rość, D. Assessment of Endothelial Progenitor Cells, VEGF-A and SDF-1α in Hodgkin’s Lymphoma. Blood Coagul. Fibrinolysis 2021, 32, 266–272. [Google Scholar] [CrossRef]
- Botelho, F.; Pina, F.; Lunet, N. VEGF and Prostatic Cancer: A Systematic Review. Eur. J. Cancer Prev. 2010, 19, 385–392. [Google Scholar] [CrossRef]
- Yuan, Q.; Sun, L.; Li, J.J.; An, C.H. Elevated VEGF Levels Contribute to the Pathogenesis of Osteoarthritis. BMC Musculoskelet. Disord. 2014, 15, 437. [Google Scholar] [CrossRef]
- Marneros, A.G. Increased VEGF-A Promotes Multiple Distinct Aging Diseases of the Eye through Shared Pathomechanisms. EMBO Mol. Med. 2016, 8, 208–231. [Google Scholar] [CrossRef]
- Shim, J.W.; Madsen, J.R. VEGF Signaling in Neurological Disorders. Int. J. Mol. Sci. 2018, 19, 275. [Google Scholar] [CrossRef]
- Zhou, W.; Liu, K.; Zeng, L.; He, J.; Gao, X.; Gu, X.; Chen, X.; Jing Li, J.; Wang, M.; Wu, D.; et al. Targeting VEGF-A/VEGFR2 Y949 Signaling-Mediated Vascular Permeability Alleviates Hypoxic Pulmonary Hypertension. Circulation 2022, 146, 1855–1881. [Google Scholar] [CrossRef] [PubMed]
- Karakioulaki, M.; Papakonstantinou, E.; Goulas, A.; Stolz, D. The Role of Atopy in COPD and Asthma. Front. Med. 2021, 8, 674742. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.M.; Arepalli, S.; Ehlers, J.P. Current and Future Anti-VEGF Agents for Neovascular Age-Related Macular Degeneration. J. Exp. Pharmacol. 2021, 13, 905–912. [Google Scholar] [CrossRef]
- Bahrami, B.; Hong, T.; Gilles, M.C.; Chang, A. Anti-VEGF Therapy for Diabetic Eye Diseases. Asia-Pac. J. Ophthalmol. 2017, 6, 535–545. [Google Scholar] [CrossRef]
- Chay, J.; Fenner, B.J.; Finkelstein, E.A.; Teo, K.Y.C.; Cheung, C.M.G. Real-World Cost-Effectiveness of Anti-VEGF Monotherapy and Combination Therapy for the Treatment of Polypoidal Choroidal Vasculopathy. Eye 2022, 36, 2265–2270. [Google Scholar] [CrossRef]
- Linghu, D.; Cheng, Y.; Zhu, X.; Deng, X.; Yin, H.; Jiang, Y.; Zhao, M.; Li, X.; Liang, J. Comparison of Intravitreal Anti-VEGF Agents With Laser Photocoagulation for Retinopathy of Prematurity of 1,627 Eyes in China. Front. Med. 2022, 9, 911095. [Google Scholar] [CrossRef]
- Sun, Y.; Liang, Y.; Zhou, P.; Wu, H.; Hou, X.; Ren, Z.; Li, X.; Zhao, M. Anti-VEGF Treatment Is the Key Strategy for Neovascular Glaucoma Management in the Short Term. BMC Ophthalmol. 2016, 16, 150. [Google Scholar] [CrossRef]
- Sperandio, R.C.; Pestana, R.C.; Miyamura, B.V.; Kaseb, A.O. Hepatocellular Carcinoma Immunotherapy. Annu. Rev. Med. 2022, 73, 267–278. [Google Scholar] [CrossRef]
- Qi, W.X.; Fu, S.; Zhang, Q.; Guo, X.M. Efficacy and Toxicity of Anti-VEGF Agents in Patients with Castration-Resistant Prostate Cancer: A Meta-Analysis of Prospective Clinical Studies. Asian Pac. J. Cancer Prev. 2014, 15, 8177–8182. [Google Scholar] [CrossRef]
- Mavissakalian, M.; Brady, S. The Current State of Biologic Therapies for Treatment of Refractory Asthma. Clin. Rev. Allergy Immunol. 2020, 59, 195–207. [Google Scholar] [CrossRef]
- Tan, L.D.; Nguyen, N.; Alismail, A.; Castro, M. Management of Uncontrolled Asthma: A Framework for Novel and Legacy Biologic Treatments. J. Asthma Allergy 2022, 15, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Pérez de Llano, L.; Cosío, B.G.; Iglesias, A.; de Las Cuevas, N.; Soler-Cataluña, J.J.; Izquierdo, J.L.; López-Campos, J.L.; Calero, C.; Plaza, V.; Miravitlles, M.; et al. Mixed Th2 and Non-Th2 Inflammatory Pattern in the Asthma–COPD Overlap: A Network Approach. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 13, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Rhee, Y.K. Overlap Between Asthma and COPD: Where the Two Diseases Converge. Allergy Asthma Immunol. Res. 2010, 2, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Louie, S.; Zeki, A.A.; Schivo, M.; Chan, A.L.; Yoneda, K.Y.; Avdalovic, M.; Morrissey, B.M.; Albertson, T.E. The Asthma–Chronic Obstructive Pulmonary Disease Overlap Syndrome: Pharmacotherapeutic Considerations. Expert Rev. Clin. Pharmacol. 2013, 6, 197–219. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Eapen, M.S.; Chia, C.; Gaikwad, A.V.; Wark, P.A.B.; Sohal, S.S. Pathogenesis, Clinical Features of Asthma COPD Overlap, and Therapeutic Modalities. Am. J. Physiol. Lung Cell. Mol. Physiol. 2022, 322, L64–L83. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tota, M.; Łacwik, J.; Laska, J.; Sędek, Ł.; Gomułka, K. The Role of Eosinophil-Derived Neurotoxin and Vascular Endothelial Growth Factor in the Pathogenesis of Eosinophilic Asthma. Cells 2023, 12, 1326. https://doi.org/10.3390/cells12091326
Tota M, Łacwik J, Laska J, Sędek Ł, Gomułka K. The Role of Eosinophil-Derived Neurotoxin and Vascular Endothelial Growth Factor in the Pathogenesis of Eosinophilic Asthma. Cells. 2023; 12(9):1326. https://doi.org/10.3390/cells12091326
Chicago/Turabian StyleTota, Maciej, Julia Łacwik, Julia Laska, Łukasz Sędek, and Krzysztof Gomułka. 2023. "The Role of Eosinophil-Derived Neurotoxin and Vascular Endothelial Growth Factor in the Pathogenesis of Eosinophilic Asthma" Cells 12, no. 9: 1326. https://doi.org/10.3390/cells12091326