
Promoting the Differentiation of Neural Progenitor Cells into Oligodendrocytes through the Induction of Olig2 Expression: A Transcriptomic Study Using RNA-seq Analysis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lentivirus Packaging
2.2. ioNPC Generation
2.3. qRT-PCR
2.4. Immunocytochemistry
2.5. RNA Sequencing
2.6. Differential Gene Expression Analysis
2.7. Gene Ontology Analysis
2.8. Assessment of Cell Lineage Gene Programs Using Gene Set Enrichment Analysis
3. Results
3.1. Morphological and Transcriptional Analyses of ioNPCs Reveals Progressive Oligodendrogenic Lineage Development following Doxycycline Administration
3.2. In Vitro Differentiation Analysis of the Doxycycline-Treated ioNPCs Reveals a Bias along the Oligodendrogenic Lineage
3.3. RNA Sequencing Analysis of the ioNPCs Reveals Differentially Expressed Genes Linked to the Commitment of the Oligodendrogenic Lineage
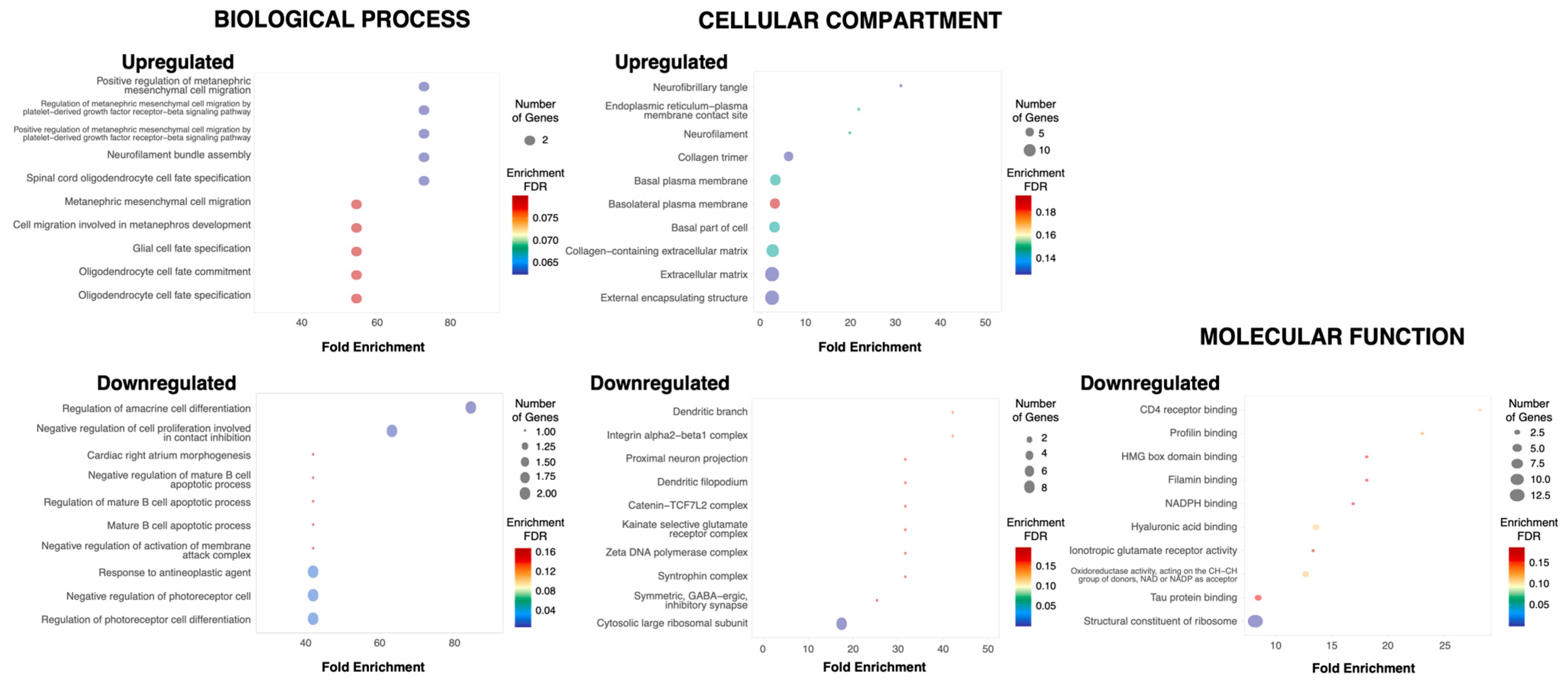
3.4. Gene Ontology Analysis Reveals That the Upregulated Genes in the ioNPCs Correspond to Pathways Related to Oligodendrocyte Development
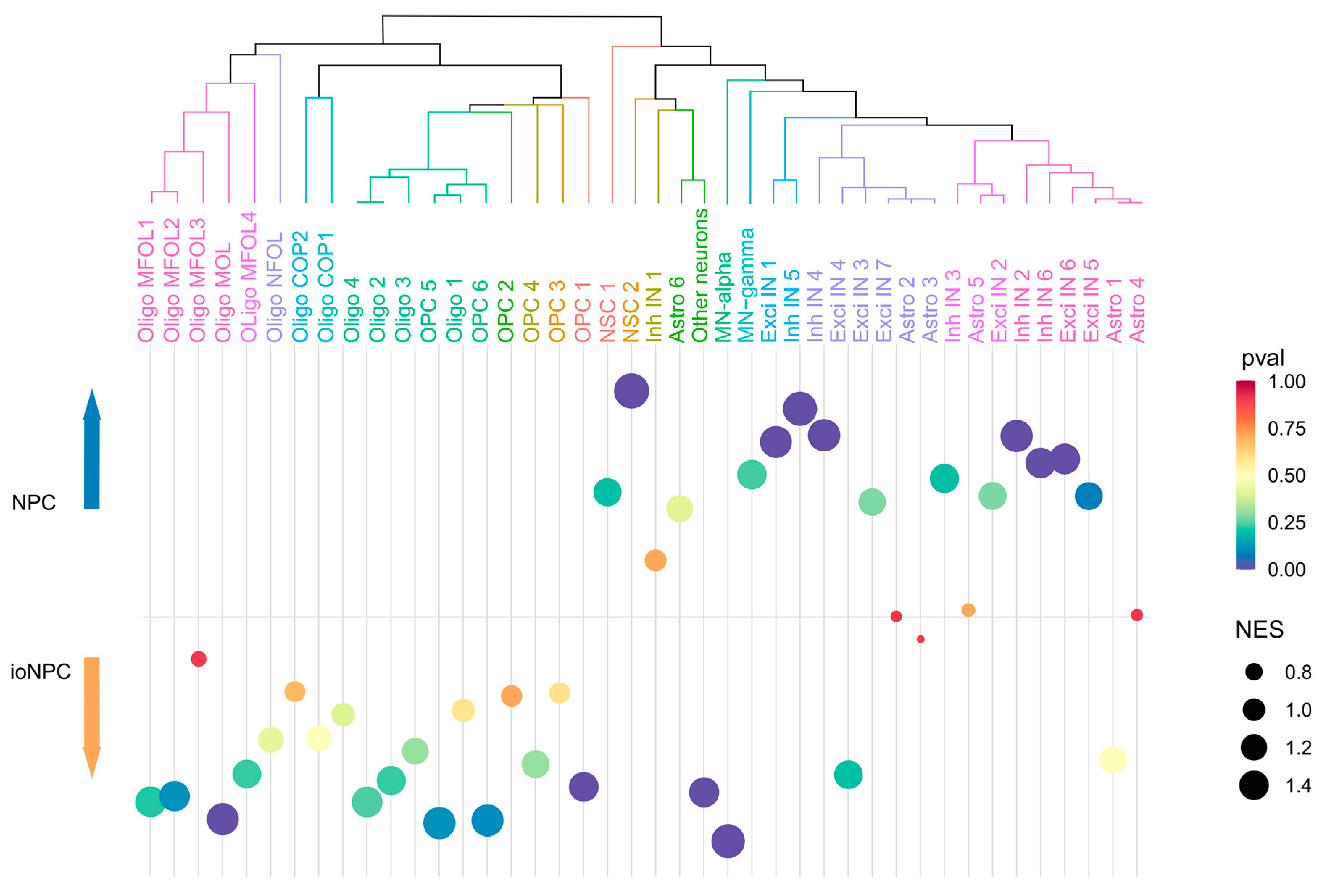
3.5. ioNPCs Exhibit Enrichment in the Gene Expression Signatures of Distinct Cells along the Oligodendrogenic Lineage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wellman, S.M.; Cambi, F.; Kozai, T.D. The Role of Oligodendrocytes and Their Progenitors on Neural Interface Technology: A Novel Perspective on Tissue Regeneration and Repair. Biomaterials 2018, 183, 200–217. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Zhang, Z.; Zhou, H.; Zhou, J. Differential Modulators of NG2-Glia Differentiation into Neurons and Glia and Their Crosstalk. Cell. Mol. Neurobiol. 2021, 41, 1–15. [Google Scholar] [CrossRef]
- Sakry, D.; Yigit, H.; Dimou, L.; Trotter, J. Oligodendrocyte Precursor Cells Synthesize Neuromodulatory Factors. PLoS ONE 2015, 10, e0127222. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Myelin Damage and Repair in Pathologic CNS: Challenges and Prospects. Front. Mol. Neurosci. 2015, 8, 35. [Google Scholar] [CrossRef]
- Khazaei, M.; Ahuja, C.S.; Nakashima, H.; Nagoshi, N.; Li, L.; Wang, J.; Chio, J.; Badner, A.; Seligman, D.; Ichise, A.; et al. GDNF Rescues the Fate of Neural Progenitor Grafts by Attenuating Notch Signals in the Injured Spinal Cord in Rodents. Sci. Transl. Med. 2020, 12, eaau3538. [Google Scholar] [CrossRef]
- Khazaei, M.; Ahuja, C.S.; Fehlings, M.G. Generation of Oligodendrogenic Spinal Neural Progenitor Cells from Human Induced Pluripotent Stem Cells. Curr. Protoc. Stem Cell. Biol. 2017, 42, 2D–20. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, N.; Khazaei, M.; Ahlfors, J.-E.; Ahuja, C.S.; Nori, S.; Wang, J.; Shibata, S.; Fehlings, M.G. Human Spinal Oligodendrogenic Neural Progenitor Cells Promote Functional Recovery after Spinal Cord Injury by Axonal Remyelination and Tissue Sparing. Stem Cells Transl. Med. 2018, 7, 806–818. [Google Scholar] [CrossRef]
- Nori, S.; Khazaei, M.; Ahuja, C.S.; Yokota, K.; Ahlfors, J.-E.; Liu, Y.; Wang, J.; Shibata, S.; Chio, J.; Hettiaratchi, M.H.; et al. Human Oligodendrogenic Neural Progenitor Cells Delivered with Chondroitinase ABC Facilitate Functional Repair of Chronic Spinal Cord Injury. Stem Cell. Rep. 2018, 11, 1433–1448. [Google Scholar] [CrossRef]
- Takebayashi, H.; Nabeshima, Y.; Yoshida, S.; Chisaka, O.; Ikenaka, K.; Nabeshima, Y. The Basic Helix-Loop-Helix Factor Olig2 Is Essential for the Development of Motoneuron and Oligodendrocyte Lineages. Curr. Biol. 2002, 12, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Zuchero, J.B.; Ahlenius, H.; Marro, S.; Ng, Y.H.; Vierbuchen, T.; Hawkins, J.S.; Geissler, R.; Barres, B.A.; Wernig, M. Generation of Oligodendroglial Cells by Direct Lineage Conversion. Nat. Biotechnol. 2013, 31, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, K.; Nobuta, H.; Yang, N.; Ang, C.E.; Huie, P.; Jordan, S.; Oldham, M.C.; Rowitch, D.H.; Wernig, M. Generation of Functional Human Oligodendrocytes from Dermal Fibroblasts by Direct Lineage Conversion. Development 2022, 149, dev199723. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.H.; Kim, B.G.; Kim, E.J.; Lee, S.I.; Joo, I.S.; Suh-Kim, H.; Sohn, S.; Kim, S.U. Transplantation of Human Neural Stem Cells Transduced with Olig2 Transcription Factor Improves Locomotor Recovery and Enhances Myelination in the White Matter of Rat Spinal Cord Following Contusive Injury. BMC Neurosci. 2009, 10, 117. [Google Scholar] [CrossRef]
- Chou, B.-K.; Mali, P.; Huang, X.; Ye, Z.; Dowey, S.N.; Resar, L.M.; Zou, C.; Zhang, Y.A.; Tong, J.; Cheng, L. Efficient Human IPS Cell Derivation by a Non-Integrating Plasmid from Blood Cells with Unique Epigenetic and Gene Expression Signatures. Cell Res. 2011, 21, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Chambers, S.M.; Fasano, C.A.; Papapetrou, E.P.; Tomishima, M.; Sadelain, M.; Studer, L. Highly Efficient Neural Conversion of Human ES and IPS Cells by Dual Inhibition of SMAD Signaling. Nat. Biotechnol. 2009, 27, 275–280. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A Graphical Gene-Set Enrichment Tool for Animals and Plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Artegiani, B.; Lyubimova, A.; Muraro, M.; van Es, J.H.; van Oudenaarden, A.; Clevers, H. A Single-Cell RNA Sequencing Study Reveals Cellular and Molecular Dynamics of the Hippocampal Neurogenic Niche. Cell Rep. 2017, 21, 3271–3284. [Google Scholar] [CrossRef] [PubMed]
- Weng, Q.; Wang, J.; Wang, J.; He, D.; Cheng, Z.; Zhang, F.; Verma, R.; Xu, L.; Dong, X.; Liao, Y.; et al. Single-Cell Transcriptomics Uncovers Glial Progenitor Diversity and Cell Fate Determinants during Development and Gliomagenesis. Cell Stem Cell 2019, 24, 707–723.e8. [Google Scholar] [CrossRef] [PubMed]
- Tirosh, I.; Venteicher, A.S.; Hebert, C.; Escalante, L.E.; Patel, A.P.; Yizhak, K.; Fisher, J.M.; Rodman, C.; Mount, C.; Filbin, M.G.; et al. Single-Cell RNA-Seq Supports a Developmental Hierarchy in Human Oligodendroglioma. Nature 2016, 539, 309–313. [Google Scholar] [CrossRef]
- Fan, X.; Dong, J.; Zhong, S.; Wei, Y.; Wu, Q.; Yan, L.; Yong, J.; Sun, L.; Wang, X.; Zhao, Y.; et al. Spatial Transcriptomic Survey of Human Embryonic Cerebral Cortex by Single-Cell RNA-Seq Analysis. Cell Res. 2018, 28, 730–745. [Google Scholar] [CrossRef]
- Russ, D.E.; Cross, R.B.P.; Li, L.; Koch, S.C.; Matson, K.J.E.; Yadav, A.; Alkaslasi, M.R.; Lee, D.I.; Le Pichon, C.E.; Menon, V.; et al. A Harmonized Atlas of Mouse Spinal Cord Cell Types and Their Spatial Organization. Nat. Commun. 2021, 12, 5722. [Google Scholar] [CrossRef]
- Rosenberg, A.B.; Roco, C.M.; Muscat, R.A.; Kuchina, A.; Sample, P.; Yao, Z.; Graybuck, L.T.; Peeler, D.J.; Mukherjee, S.; Chen, W.; et al. Single-Cell Profiling of the Developing Mouse Brain and Spinal Cord with Split-Pool Barcoding. Science 2018, 360, 176–182. [Google Scholar] [CrossRef]
- McCaughey-Chapman, A.; Connor, B. Cell Reprogramming for Oligodendrocytes: A Review of Protocols and Their Applications to Disease Modeling and Cell-Based Remyelination Therapies. J. Neurosci. Res. in press. 2023. [Google Scholar] [CrossRef]
- McIntyre, W.B.; Pieczonka, K.; Khazaei, M.; Fehlings, M.G. Regenerative Replacement of Neural Cells for Treatment of Spinal Cord Injury. Expert. Opin. Biol. Ther. 2021, 21, 1411–1427. [Google Scholar] [CrossRef]
- Patel, J.; Balabanov, R. Molecular Mechanisms of Oligodendrocyte Injury in Multiple Sclerosis and Experimental Autoimmune Encephalomyelitis. Int. J. Mol. Sci. 2012, 13, 10647–10659. [Google Scholar] [CrossRef] [PubMed]
- Hejrati, N.; McIntyre, W.B.; Pieczonka, K.; Ostmeier, S.; Ahuja, C.S.; Kwon, B.K.; Vaccaro, A.R.; Oner, F.C.; Fehlings, M.G. Chapter 31—Translational Research in Spinal Cord Injury—What Is in the Future? In Neural Repair and Regeneration after Spinal Cord Injury and Spine Trauma; Fehlings, M.G., Kwon, B.K., Vaccaro, A.R., Oner, F.C., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 587–602. ISBN 978-0-12-819835-3. [Google Scholar]
- Petersen, J.A.; Wilm, B.J.; von Meyenburg, J.; Schubert, M.; Seifert, B.; Najafi, Y.; Dietz, V.; Kollias, S. Chronic Cervical Spinal Cord Injury: DTI Correlates with Clinical and Electrophysiological Measures. J. Neurotrauma 2012, 29, 1556–1566. [Google Scholar] [CrossRef]
- Kisel, A.A.; Naumova, A.V.; Yarnykh, V.L. Macromolecular Proton Fraction as a Myelin Biomarker: Principles, Validation, and Applications. Front. Neurosci. 2022, 16, 819912. [Google Scholar] [CrossRef]
- Kolind, S.; Seddigh, A.; Combes, A.; Russell-Schulz, B.; Tam, R.; Yogendrakumar, V.; Deoni, S.; Sibtain, N.A.; Traboulsee, A.; Williams, S.C.R.; et al. Brain and Cord Myelin Water Imaging: A Progressive Multiple Sclerosis Biomarker. Neuroimage Clin. 2015, 9, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Argaw, A.T.; Gurfein, B.T.; Zameer, A.; Snyder, B.J.; Ge, C.; Lu, Q.R.; Rowitch, D.H.; Raine, C.S.; Brosnan, C.F.; et al. Notch1 Signaling Plays a Role in Regulating Precursor Differentiation during CNS Remyelination. Proc. Natl. Acad. Sci. USA 2009, 106, 19162–19167. [Google Scholar] [CrossRef]
- Fancy, S.P.J.; Baranzini, S.E.; Zhao, C.; Yuk, D.-I.; Irvine, K.-A.; Kaing, S.; Sanai, N.; Franklin, R.J.M.; Rowitch, D.H. Dysregulation of the Wnt Pathway Inhibits Timely Myelination and Remyelination in the Mammalian CNS. Genes Dev. 2009, 23, 1571–1585. [Google Scholar] [CrossRef] [PubMed]
- Kotter, M.R.; Li, W.-W.; Zhao, C.; Franklin, R.J.M. Myelin Impairs CNS Remyelination by Inhibiting Oligodendrocyte Precursor Cell Differentiation. J. Neurosci. 2006, 26, 328–332. [Google Scholar] [CrossRef]
- Baer, A.S.; Syed, Y.A.; Kang, S.U.; Mitteregger, D.; Vig, R.; ffrench-Constant, C.; Franklin, R.J.M.; Altmann, F.; Lubec, G.; Kotter, M.R. Myelin-Mediated Inhibition of Oligodendrocyte Precursor Differentiation Can Be Overcome by Pharmacological Modulation of Fyn-RhoA and Protein Kinase C Signalling. Brain 2009, 132, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, A.A.; Gowing, G.; Kobritz, N.; Savinoff, S.E.; Garcia, L.; Saxon, D.; Cho, N.; Kim, G.; Tom, C.M.; Park, H.; et al. Inducible Expression of GDNF in Transplanted IPSC-Derived Neural Progenitor Cells. Stem Cell Rep. 2018, 10, 1696–1704. [Google Scholar] [CrossRef] [PubMed]
- Nau, R.; Sörgel, F.; Eiffert, H. Penetration of Drugs through the Blood-Cerebrospinal Fluid/Blood-Brain Barrier for Treatment of Central Nervous System Infections. Clin. Microbiol. Rev. 2010, 23, 858–883. [Google Scholar] [CrossRef] [PubMed]
- Haddad, N.; Carr, M.; Balian, S.; Lannin, J.; Kim, Y.; Toth, C.; Jarvis, J. The Blood–Brain Barrier and Pharmacokinetic/Pharmacodynamic Optimization of Antibiotics for the Treatment of Central Nervous System Infections in Adults. Antibiotics 2022, 11, 1843. [Google Scholar] [CrossRef]
- Floriddia, E.M.; Lourenço, T.; Zhang, S.; van Bruggen, D.; Hilscher, M.M.; Kukanja, P.; Gonçalves Dos Santos, J.P.; Altınkök, M.; Yokota, C.; Llorens-Bobadilla, E.; et al. Distinct Oligodendrocyte Populations Have Spatial Preference and Different Responses to Spinal Cord Injury. Nat. Commun. 2020, 11, 5860. [Google Scholar] [CrossRef]
- Jäkel, S.; Agirre, E.; Mendanha Falcão, A.; van Bruggen, D.; Lee, K.W.; Knuesel, I.; Malhotra, D.; Ffrench-Constant, C.; Williams, A.; Castelo-Branco, G. Altered Human Oligodendrocyte Heterogeneity in Multiple Sclerosis. Nature 2019, 566, 543–547. [Google Scholar] [CrossRef]
- Marques, S.; Zeisel, A.; Codeluppi, S.; van Bruggen, D.; Mendanha Falcão, A.; Xiao, L.; Li, H.; Häring, M.; Hochgerner, H.; Romanov, R.A.; et al. Oligodendrocyte Heterogeneity in the Mouse Juvenile and Adult Central Nervous System. Science 2016, 352, 1326–1329. [Google Scholar] [CrossRef]
- Khandker, L.; Jeffries, M.A.; Chang, Y.-J.; Mather, M.L.; Evangelou, A.V.; Bourne, J.N.; Tafreshi, A.K.; Ornelas, I.M.; Bozdagi-Gunal, O.; Macklin, W.B.; et al. Cholesterol Biosynthesis Defines Oligodendrocyte Precursor Heterogeneity between Brain and Spinal Cord. Cell Rep. 2022, 38, 110423. [Google Scholar] [CrossRef]
- Bechler, M.E.; Byrne, L.; Ffrench-Constant, C. CNS Myelin Sheath Lengths Are an Intrinsic Property of Oligodendrocytes. Curr. Biol. 2015, 25, 2411–2416. [Google Scholar] [CrossRef]
- Hill, R.A.; Patel, K.D.; Medved, J.; Reiss, A.M.; Nishiyama, A. NG2 Cells in White Matter but Not Gray Matter Proliferate in Response to PDGF. J. Neurosci. 2013, 33, 14558–14566. [Google Scholar] [CrossRef]
- Lu, Q.R.; Yuk, D.; Alberta, J.A.; Zhu, Z.; Pawlitzky, I.; Chan, J.; McMahon, A.P.; Stiles, C.D.; Rowitch, D.H. Sonic Hedgehog--Regulated Oligodendrocyte Lineage Genes Encoding BHLH Proteins in the Mammalian Central Nervous System. Neuron 2000, 25, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.R.; Sun, T.; Zhu, Z.; Ma, N.; Garcia, M.; Stiles, C.D.; Rowitch, D.H. Common Developmental Requirement for Olig Function Indicates a Motor Neuron/Oligodendrocyte Connection. Cell 2002, 109, 75–86. [Google Scholar] [CrossRef]
- Cai, J.; Qi, Y.; Hu, X.; Tan, M.; Liu, Z.; Zhang, J.; Li, Q.; Sander, M.; Qiu, M. Generation of Oligodendrocyte Precursor Cells from Mouse Dorsal Spinal Cord Independent of Nkx6 Regulation and Shh Signaling. Neuron 2005, 45, 41–53. [Google Scholar] [CrossRef]
- Kessaris, N.; Fogarty, M.; Iannarelli, P.; Grist, M.; Wegner, M.; Richardson, W.D. Competing Waves of Oligodendrocytes in the Forebrain and Postnatal Elimination of an Embryonic Lineage. Nat. Neurosci. 2006, 9, 173–179. [Google Scholar] [CrossRef]
- Bergles, D.E.; Richardson, W.D. Oligodendrocyte Development and Plasticity. Cold Spring Harb. Perspect. Biol. 2016, 8, a020453. [Google Scholar] [CrossRef]
- Lee, S.-K.; Lee, B.; Ruiz, E.C.; Pfaff, S.L. Olig2 and Ngn2 Function in Opposition to Modulate Gene Expression in Motor Neuron Progenitor Cells. Genes Dev. 2005, 19, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Paes de Faria, J.; Andrew, P.; Nitarska, J.; Richardson, W.D. Phosphorylation Regulates OLIG2 Cofactor Choice and the Motor Neuron-Oligodendrocyte Fate Switch. Neuron 2011, 69, 918–929. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Long, Z.; Yang, C. The Effects of the Olig Family on the Regulation of Spinal Cord Development and Regeneration. Neurochem. Res. 2021, 46, 2776–2782. [Google Scholar] [CrossRef]
- Mie, M.; Kaneko, M.; Henmi, F.; Kobatake, E. Induction of Motor Neuron Differentiation by Transduction of Olig2 Protein. Biochem. Biophys. Res. Commun. 2012, 427, 531–536. [Google Scholar] [CrossRef]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic Spinal Cord Injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef]
- Du, Y.; Dreyfus, C.F. Oligodendrocytes as Providers of Growth Factors. J. Neurosci. Res. 2002, 68, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.; Majed, H.; Layfield, R.; Compston, A.; Chandran, S. Oligodendrocytes Promote Neuronal Survival and Axonal Length by Distinct Intracellular Mechanisms: A Novel Role for Oligodendrocyte-Derived Glial Cell Line-Derived Neurotrophic Factor. J. Neurosci. 2003, 23, 4967–4974. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Lercher, L.D.; Clinton, P.M.; Du, Y.; Livingston, D.L.; Vieira, C.; Yang, L.; Shen, M.M.; Dreyfus, C.F. The Trophic Role of Oligodendrocytes in the Basal Forebrain. J. Neurosci. 2003, 23, 5846–5853. [Google Scholar] [CrossRef]
- Lu, P.; Jones, L.L.; Snyder, E.Y.; Tuszynski, M.H. Neural Stem Cells Constitutively Secrete Neurotrophic Factors and Promote Extensive Host Axonal Growth after Spinal Cord Injury. Exp. Neurol. 2003, 181, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Pöyhönen, S.; Er, S.; Domanskyi, A.; Airavaara, M. Effects of Neurotrophic Factors in Glial Cells in the Central Nervous System: Expression and Properties in Neurodegeneration and Injury. Front. Physiol. 2019, 10, 486. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pieczonka, K.; Khazaei, M.; Fehlings, M.G. Promoting the Differentiation of Neural Progenitor Cells into Oligodendrocytes through the Induction of Olig2 Expression: A Transcriptomic Study Using RNA-seq Analysis. Cells 2023, 12, 1252. https://doi.org/10.3390/cells12091252
Pieczonka K, Khazaei M, Fehlings MG. Promoting the Differentiation of Neural Progenitor Cells into Oligodendrocytes through the Induction of Olig2 Expression: A Transcriptomic Study Using RNA-seq Analysis. Cells. 2023; 12(9):1252. https://doi.org/10.3390/cells12091252
Chicago/Turabian StylePieczonka, Katarzyna, Mohamad Khazaei, and Michael G. Fehlings. 2023. "Promoting the Differentiation of Neural Progenitor Cells into Oligodendrocytes through the Induction of Olig2 Expression: A Transcriptomic Study Using RNA-seq Analysis" Cells 12, no. 9: 1252. https://doi.org/10.3390/cells12091252