
Shining the Light on Astrocytic Ensembles


Instituto Cajal, Consejo Superior de Investigaciones Científicas (CSIC), 28002 Madrid, Spain
*
Author to whom correspondence should be addressed.
Cells 2023, 12(9), 1253; https://doi.org/10.3390/cells12091253
Submission received: 21 March 2023
/
Revised: 22 April 2023
/
Accepted: 24 April 2023
/
Published: 26 April 2023
(This article belongs to the Special Issue Glial Cells in Synaptic Plasticity)
Abstract
:While neurons have traditionally been considered the primary players in information processing, the role of astrocytes in this mechanism has largely been overlooked due to experimental constraints. In this review, we propose that astrocytic ensembles are active working groups that contribute significantly to animal conduct and suggest that studying the maps of these ensembles in conjunction with neurons is crucial for a more comprehensive understanding of behavior. We also discuss available methods for studying astrocytes and argue that these ensembles, complementarily with neurons, code and integrate complex behaviors, potentially specializing in concrete functions.
1. Introduction
The complex brain structure composed of neurons, astrocytes, and the remaining glial cells has been revealed to cooperate, share information, and code responses. This communication is accomplished by using different languages, such as flows of charge across neurons that lead to electrical potentials and ionic currents, oscillatory rhythms that direct synchronization and phasing, astrocytic and neuronal calcium transients, and the continuous chemical exchange of neurotransmitters, in which all involved parties find a common language to understand. Recently, research has been shedding light on the behavior of astrocyte networks as communicating systems that have their own rules and mechanisms [1,2,3].
Astrocytes play an important role in brain function as computing hubs. Their coding strategy relies on intracellular Ca2+ fluctuations that are very heterogeneous in space and time [4] that result from Ca2+ stores released mainly by inositol triphosphate (IP3) receptors located in the membrane of the endoplasmic reticulum (ER) [5,6]. These signals can range from single-cell spontaneous intrinsic Ca2+ variations that maintain different levels of activity [7] to intercellular Ca2+ flows that spread over long distances and can reach hundreds of cells [8,9] to transmembrane receptors-elicited Ca2+ signals that account for external stimuli [5,10]. The external executors are, substantially, neurotransmitters released synaptically. Bidirectionally, astrocytes can also release neuroactive molecules, such as glutamate, ATP, GABA, adenosine, or D-serine, that switch on neuronal receptors, establishing communication and revealing their involvement in the modulation of neuronal activity and synaptic transmission in different brain areas [11,12,13,14,15,16].
Therefore, astrocytes are significantly involved in synapse formation, remodeling, and plasticity [17,18,19], which is partly due to their evolutionarily conserved intricate branched morphology—proven both in mice and humans—[20,21]. The perisynaptic astrocytic processes (PAPs) that establish physical contact with pre- and post-synaptic terminals and form tripartite synapses [22] can undergo both gross and fine-scale structural changes analogous to those seen in neurons during plasticity modulation [23,24,25,26]. This has a functional meaning since not all synapses exhibit the presence of astrocytic processes, and it is a necessity dynamically shaped under development requirements, or in response to injury, among others [27,28,29,30]. As such, astrocytes play a key role in regulating learning and memory, which are foundational to complex behaviors. To identify the astrocytic elements involved in synaptic plasticity, studies have used electrophysiology, molecular genetics, and astrocytic knockout phenotypes, as well as examining determinant elements such as receptors (e.g., cannabinoid receptor CB1 [31], inositol trisphosphate receptor IP3Rs [32,33], GABAb [34], kinases [35]), cell adhesion molecules (e.g., connexins 43 and 30 [36], or neuroligins [37]), among other approaches [38,39].
In addition, astrocytes play an important role in intercellular information transmission through neuronal oscillations, which contribute to rhythmicity. They are essential participants in understanding the synchronous activities of central pattern generators (CPGs), which control behaviors such as respiration [40,41,42,43,44], locomotion [45,46,47,48,49], mastication [50,51,52], and the regulation of sleep patterns [53]. Astrocytes in the suprachiasmatic nucleus of the hypothalamus have been shown to independently encode circadian information and control the period of circadian oscillations through glutamate [54,55,56], and the inhibition of vesicular release in hippocampal astrocytes provokes disruption of gamma oscillatory activity [57]. In conclusion, the active role astrocytes play in neurotransmission is responsible for modulating the connectivity of neurons at the network level.
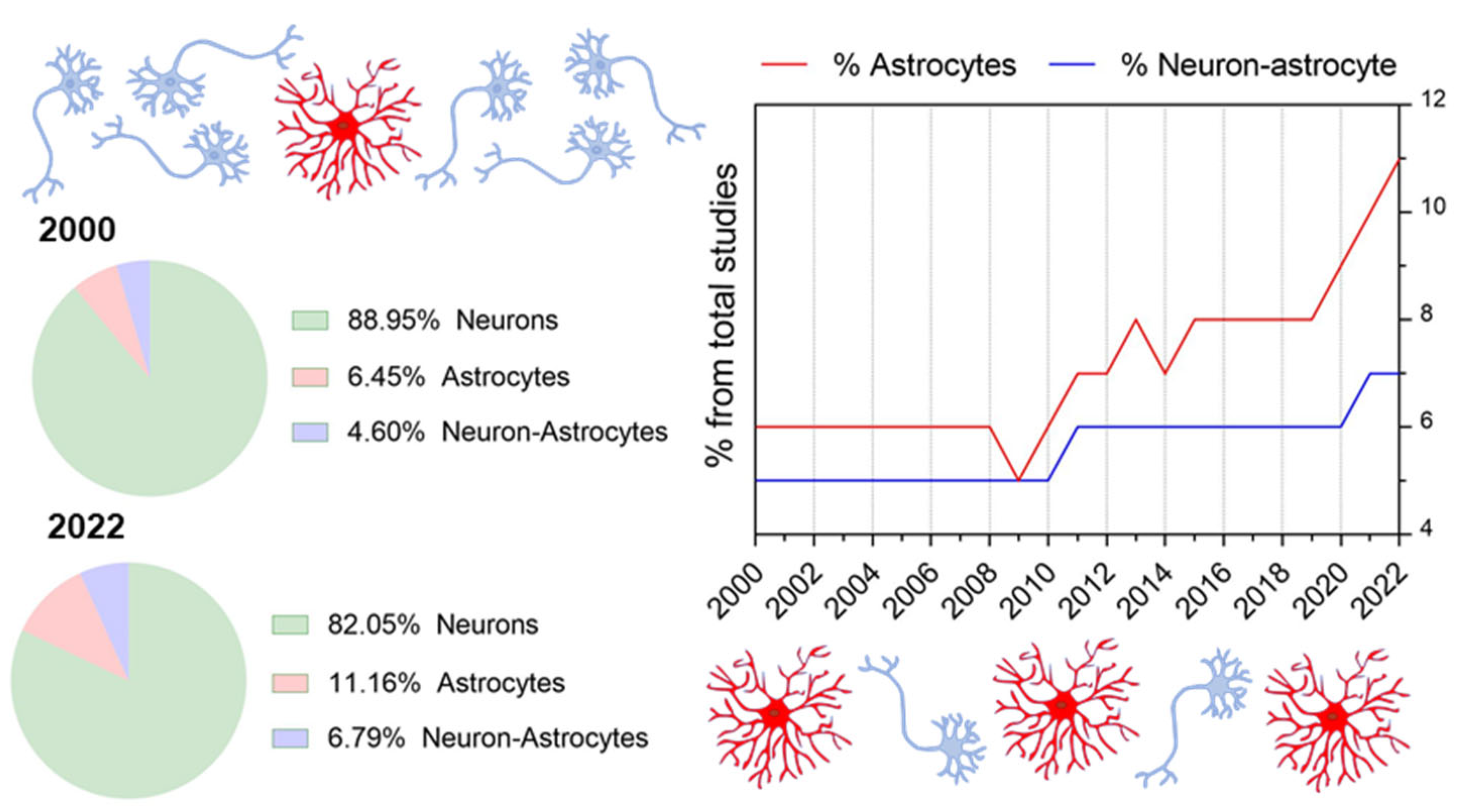
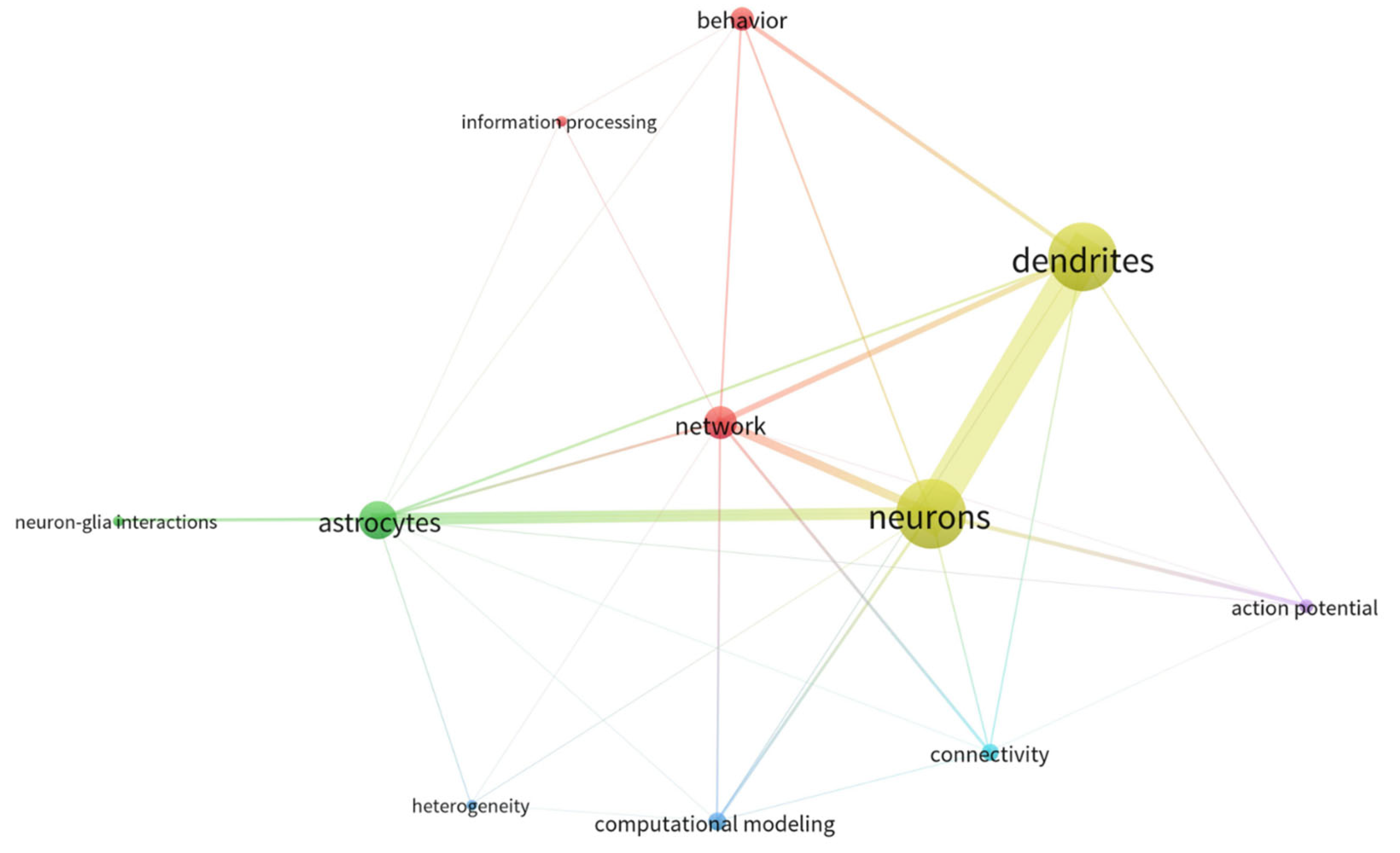
Therefore, despite the undeniably growing attention for the astrocytic field within the last several years (see Figure 1 and Figure 2 and Table 1), awareness of the existence and coordination of astrocyte working groups and their interaction with neuronal systems for behavioral encoding is still missing. Thus, whereas available studies generally focus on cause-effect astrocyte-neuron interactions or astrocyte function, almost none includes the panorama of network-interacting astrocytes with the view put in behavior. As a result, we detect a pivotal need to shape the concept of astrocytic ensembles, we discuss its supporting evidence and the conceptual relevance for implementing top-down study perspectives that ultimately allow functional correlates of astrocyte responses. Herein, we first provide an overview of the state-of-the-art methods available for studying astrocytes, showing the differences between bottom-up approaches (which are safe bets to build organized knowledge for deciphering the astrocyte’s role in the brain) and top-down perspectives (by setting the focus on astrocyte network dynamics). Next, from a systems neuroscience perspective, we discuss the features of astrocytes’ and neurons’ communication in the physiological brain as well as their connectivity patterns and argue that astrocytes’ response in behavior results from working ensembles. From that, we deduce that astrocytes may have the possibility to specialize, and we present current missing links to decipher astrocyte integration properties in physiological conditions. Lastly, we evaluate the most important concerns for the in vivo study of astrocytes.
2. Available Methods for Tracking Astrocytes
To reliably picture a system in continuous motion is an ambitious task. Herein, we review the methods available to study astrocytes and present the bottom-up and top-down study perspectives (see Table 2 and Table 3). The bottom-up approach integrates smaller pieces of information to form the bigger picture, while the top-down approach first identifies the bigger picture and then infers the relationships among the elements involved. The goal is to provide sufficient knowledge of the state-of-the-art techniques for visualizing and manipulating astrocytes to help researchers choose the right approach for their biological questions. Greater detail is beyond our scope and can be found elsewhere [6,58,59,60,61].
Structure, morphology, and diversity. The specialized and polarized end-foot structures of astrocytic processes vary in size, shape, and structure and require new labeling and imaging systems to accurately detect calcium transients in these fine processes [62]. This heterogeneity level leads to classification according to the diffraction-limited resolution required for its detection and, therefore, to their potential to be resolved using traditional microscopes. Discerning between adjacent astrocytes at single-cell resolution can be achieved with cell labeling and confocal microscopy by using lucifer yellow intracellular dye-filling through a sharp electrode; iontophoresis-based (which requires short tissue exposition to a fixative solution [63]); by expression of fluorescent reporters through virus delivery [64], transgenic mice lines [65], but also with electroporation [37]; or the sparse labeling method mosaicism with repeat frameshift (MORF) [66]. In addition, electron microscopy (EM) reconstructions also allow the detail of PAPs and end feet [67,68,69,70,71,72] but provide limited experimental flexibility due to requiring fixed tissue. By combining viral vector transduction to mark different cells with different fluorescent reporters, confocal or two-photon imaging, and the CLARITY technique, Goshen and Rafaeli [73] showed different astrocytes, their complete and detailed structure, and their spatial network interactions, resulting in a stationary way to represent large-scale volumes at single-scale resolutions. Spatio-temporal interactions between astrocyte processes and synapses have been detected with fluorescent proteins and a genetically targeted neuron–astrocyte assay (NAPA) that depends on resonance energy transfer (FRET) between the fluorophores expressed in neurons and the adjacent astrocytes [30,74].
Among the shaping factors of individual-cell structural and morphological diversities are genetic programs that give rise to determined molecular profiles [75]. The choice for resolving astrocytic regional and microenvironmental diversity are transcriptomic and proteomic analyses along with morphological reconstruction, which also allows for studying the occurring changes dependent on synapses, behavior, or experience [65,76,77,78]. Of concern is, therefore, the traditional limitation of molecular approaches to study transcriptomes and proteomes: the need to isolate cells (i.e., to undergo a purification process), which has been shown to change naturally occurring gene expression and does not allow spatial traceability. There are approaches to sequence RNA (RNA-seq), also single-cell (scRNA-seq) [79], including fluorescence-activated cell sorting (FACS) [80], immunopanning [81,82], and magnetic-activated cell sorting [83,84]. Translating ribosome affinity purification (TRAP) [85,86] does not require physical cell dissociation; however, the specific technique that allows for in vivo genetic targeting of selected cell populations is the protocol RiboTag [87], allowing labeling even the translatomes of subcellular compartments such as the end feet, and low-expressing genes [88], overcoming the need to having segregated cells. In regard to techniques that allow spatial transcriptomics [89,90,91], some already commercialized are Spatial Transcriptomics [92], GeoMx [93], and CosMx [94].
Indicators of Ca2+ activity. The spectrum of study possibilities commences with calcium-specific dyes (i.e., fluorescent reporters conjugated to a Ca2+ chelator such as Fluo-4, Rod-2-, or OregonGreen-BAPTA, among others [59,60]), which were the first used for measuring intracellular calcium [95,96]. In addition, there are genetically encoded Ca2+ indicators (GECIs) that bind to calcium and emit fluorescence (e.g., GCaMP [97]). Cre-dependent transgenic mice [98,99,100] and adeno-associated viruses (AAV) are available to express GECIs specifically in astrocytes by tackling specific astrocytic reporters (such as GFAP, S100β, GLAST, or GLT-1 [58,101,102]), with which it is possible to identify calcium signals across different areas, or within fine leaflets [103,104]. The combination of GECIs and optic fiber is a widely used approach to register deep-tissue measurements of calcium transients in freely behaving mice [59,103,105]. In regard to imaging, for instance, Georgiou et al. [106] used axoastrocytic AAV transfer and two-photon microscopy to study the functional-dependent organization of Ca2+ microdomains and were able to determine stability within occurring patterns. Additionally, in an in vivo study carried out by Li et al. [107], a combination of magnetic resonance imaging (MRI) with an AAV vector carrying a reporter was used to uncover dynamic Ca2+ changes in astrocytes. Furthermore, in our laboratory, we have recently adapted a functional tool to monitor astrocyte activity called CaMPARIGFAP, which is a GECI for astrocytes that undergoes irreversible fluorescence conversion when there is elevated intracellular Ca2+ and simultaneous delivery of 405 nm light [3]. Apart from allowing the study in a temporally controlled manner, it enables the labeling of a specific set of astrocytes that are active during a certain time window resulting in functional maps of the instantaneous picture of astrocyte activity. Indicators can also be genetically targeted at different signaling molecules such as second messengers or metabolites (i.e., ATP, serotonin, glucose, or glutamate, among others) to follow up the different input/output activities. Even though the simultaneous study of several molecules using different indicators can result in being restricted due to genetic constraints or fluorophores overlap, having the possibility of measuring intracellular signaling enables establishing behavioral correlates or gene expression changes and sets a step forward towards delineating the underlying computing principles.
Actuators. To interact with the system in an inhibitory or excitatory manner for studying the effects is an interesting approach for establishing connections between animal behavior, neural functioning, and astrocyte activity, despite the absence of strict physiological context [108]. Activation of astrocytes is traditionally performed with opto- and chemogenetic approaches such as designer receptors exclusively activated with designer drugs (DREADDs) [109,110], channelrhodopsin 2 (ChR2) [111], the Gq-dependent actuator melanopsin [112], the light-gated ionotropic glutamate receptor LiGluR [30,113], or the engineered light-sensitive GPCRs called opto-XRs [114]. As for inhibition, multiple agonists tackling different targets can be used depending on the source of the input signal (e.g., signals mediated by Gq GPCR [115] or Ca2+ release from intracellular stores in which IP3 type 2 receptors are involved [116]). With calcium extruders, which is a genetic approach, it is also possible to attenuate calcium signaling regardless of the source [117].
Advanced in vivo imaging technologies. Emerging imaging technologies are two-photon laser scanning microscopy (2PLSM)—which reaches significant tissue depths in the order of 500 nm—[118,119]; laser scanning confocal microscopy (LSCM)—suitable for more superficial areas—[120]; stimulation emission depletion (STED)—a right choice for fine process detection—[121]; or light sheet fluorescent microscopy (LSFM)—that allows acquisition of ultrafast calcium transients—[122,123]. For instance, using 2PLSM, Volterra et al. [124] tracked in 3D the Ca2+ activity of astrocytic processes in the hippocampus, revealing different types of events and correlating them with local synapses. Nevertheless, methods such as two-photon stimulation emission depletion (2P-STED) are emerging for in vivo exploration of interactions under physiological conditions, allowing correlation with activity dependence.
Analysis tools. Ultimately, the efficacy of manipulating and imaging methods will also depend on the development of analytical tools [58,60] to focus the resulting data toward predictive modeling. Available software for analyzing Ca 2+ signals includes Astrocyte Quantitative Analysis (AQuA) [125], which is a non-ROIs (regions-of-interest)-based analysis that captures the heterogeneous fluorescent dynamics, allows spatio-temporal quantification and is a good choice for examining dynamical properties of distinct calcium transients. In contrast, GECIquant [126] determines ROIs containing calcium changes based on a certain threshold value, permits differentiating between process-occurring activity or somatic fluctuations, and is better suited for examining localized calcium transients within single cells. An ROI-based machine learning method is CaSCaDe [127], which outputs a spatial map of activity and can be used to monitor single-cell calcium microdomains. Partition in Regular Quadrants (PRQ), which is ROI-based, also enables a spatial analysis to obtain activity maps [3] or the MATLAB custom program Calsee [128] to contour the territories of individual astrocytes and quantify region-specific calcium activity. The choice between ROI- or event-based methods should be made based on how limiting it is that ROIs are constant in shape, position, and overlap [125]. In terms of network analysis, Astral was designed to analyze astrocyte–astrocyte population interactions [129] and provides an event-based approach to measure single-cell events and their intercellular propagation.
To summarize, experimental top-down study methods in astrocytes have been very limited until recently, which has considerably slowed down the possibility of setting the focus on astrocyte network dynamics. However, now there are available techniques to combine the in vivo labeling of active astrocytic ensembles with quantitative measures of their activity (with advanced imaging or fiber photometry recordings). This enables tracking the astrocytic activity at the network level during a certain behavior but also deciphering the astroglial physiological activation patterns, which are essential for establishing relationships to infer the integration properties of astrocytes.

{kind=link}
{kind=link}
{kind=link}
Table 2.
Experimental techniques to label and visualize astrocytes.
| Experimental Techniques | ||
|---|---|---|
| Structure and/or Morphology of Astrocytes | References | |
| Labeling |
| [63] |
| [64] | |
| [65] | |
| [37] | |
| [66] | |
| Imaging |
| [67,68,69,70,71,72] |
| [118,119] | |
| [120] | |
| [121] | |
| [122,123] | |
| Labeling and Imaging |
| [73] |
| Labeling and Imaging and FRET Phenomenon |
| [30,74] |
| Genetic Programs and Molecular Profiles | ||
| RNA-Seq and Single Cell RNA-Seq |
| [80] |
| [81,82] | |
| [83,84] | |
| [85,86] | |
| [87,88] | |
| Spatial Transcriptomics |
| [92] |
| [93] | |
| [94] | |
| Activity of Astrocytes | ||
| Ca2+ Specific Dyes for Intracellular Measurements |
| [59,60] |
| Genetically Encoded Ca2+Indicators(GECIs) |
| [3,97] |
| [98,99,100] | |
| [58,101,102,106,107] | |
| Activation of Astrocytes |
| [109,110] |
| [111] | |
| [130] | |
| [30,113] | |
| [114] | |
| Inhibition of astrocytes |
| [115,116] |
| [117] | |
Table 3.
Analytical tools to study astrocytes.
| Analytical Tools | References | Properties |
|---|---|---|
| Astrocyte Quantitative Analysis (AQuA) | [125] |
|
| GECIquant | [126] |
|
| CaSCaDe | [127] |
|
| Partition in Regular Quadrants (PRQ) | [3] |
|
| Calsee | [128] |
|
| Astral | [129] |
|
3. Astrocytes Add Complexity and Increase Integration Versatility
3.1. Definition of Functional Astrocytic Ensembles
The central axis of systems neuroscience for understanding the biological complexity of the brain is to seek a systems-level description of the network interactions regardless of analyzing its individual components. To date, the discipline barely considers the role of glial cells [131]. In the following lines, we focus on the interactions between neuronal and astrocytic systems and on their different connectivity patterns, shaped by their distinct electrical, structural, and temporal features.
Neurons and astrocytes have developed their own individual attributes that lead to different network working principles. On the one hand, neurons have high electrical excitability, while astrocytes have the capacity for K+ buffering and, therefore, for controlling this electrical excitability of neurons. This difference in electrical barriers may allow the overall system to balance activation and achieve maximum efficiency. On the other hand, astrocytes form a continuous assembly due to their connection through gap junctions that facilitate intercellular coupling, control the flow of molecules, and allow the sharing of cytoplasm, leading to a reticular organization known as syncytium [132], while neurons form a network of independent connected units. The divergence in electrical and structural features between astrocytes and neurons is also found in the temporal resolution dynamics that provide each system with a separate, complementary field of action [31,124,133].
The modularity theory is well-assumed in most theoretical approaches in cognitive science to represent innate neural structures that work together to form functional circuits [134,135]. In fact, there are supportive studies that report, for instance, how (1) by externally triggering certain neuronal patterns, it is possible to induce motor tasks in humans [136], (2) by triggering the electrical encoding of facial identity, it is possible to reproduce it with a high degree of accuracy in macaques [137], or even (3) decision-making can be predictable from action potential sequences with a wise choice probability above 70% [138,139]. Nevertheless, none include the response refinement encoded by astrocytes, missing an important variable that should be considered indispensable [13,34,54,58,108,115,140,141,142,143].
There are no solid responses regarding how astrocytes encode outputs to influence behavior, despite the evidence showing their involvement. An example is found in the astrocytic implications of memory consolidation, a complex behavior that requires the integration of reasoning, comprehension, and learning in the case of humans. In 2012, Han et al. [31] showed that activation of astroglial cannabinoid receptors is sufficient to impair spatial working memory (SWM) and induce long-term depression (LTD). Recently, a study published by Becker et al. [133] showed how working memory representations are influenced by astrocyte signaling by developing a computational model of a neural network that considers astrocytes, which resulted in actions on presynaptic short-term plasticity (STP) dynamics by increasing its calcium binding rate. Namely, they show how the fast STP dynamics are modulated by the slow timescale of astrocytes that ranges from seconds to minutes suggesting a slow integrator role that also saves information about the active representation. Gordleeva et al. [144] go further in the modeling by considering astrocyte–astrocyte interactions and draw conclusions on their ability to store patterns to coherently perform synaptic changes, incorporating novel biophysical network mechanisms of memory formation. The latter is supported as well, among others [145,146,147], by Porto-Pazos et al. [148], who concluded that artificial neuron–glia networks (NGN) provide more efficient performance in comparison to sole artificial neural networks (NN) and that this increased complexity is a factor for network performance improvement.
Therefore, the conclusion that astrocytes are indispensable in brain function related to behavior is emerging in parallel to the growing evidence showing how astrocytes are organized into functional clusters. In the research carried out by Houades et al. [149], it is shown that astrocytes in the barrel cortex couple largely through gap junctions in the same barrel and scarcely between astrocytes from different barrels, which define local microcircuits. Apparently, the established connectivity is region-specific, revealing a non-homogeneous character of gap junctions and maybe interacting astrocytic subpopulations. Sul et al. [150] concluded that adjoining astrocytes in the hippocampus were functionally connected and that this recruitment was determined by certain threshold concentrations of the extracellular transmitters ATP and ACh. However, how this specificity is modulated remains unknown. Furthermore, in a recently published study by our laboratory by Serra et al. [3], the stimulation of three different glutamatergic nuclei resulted in differential astrocytic activity in the nucleus accumbens in a way that for each stimulated nucleus astrocytes not only responded in highly innervated areas, which supports the existence of different connected astrocyte subpopulations able to discern the input origin. This astrocytic discrimination between pathways was also previously observed by Perea and Araque [151]. Moreover, the encoding of spatial information by hippocampal astrocytes has been experimentally investigated by Curreli and colleagues [152], who conclude that the coding is not explained by mere visual cue information. They give evidence about how spatial information was processed differentially in distinct topographical locations of the same astrocyte and that it complemented neuron-encoded information in an excluding way. Doron et al. [2] enriched Curreli’s findings by tracking real-time calcium activity of astrocytes in the hippocampus also during a spatial event-related task. The results showed how astrocytic calcium activity increased after recognition of a learned reward location. The latter means that astrocytes code information levels since the increase was not appreciated in non-familiar environments. Importantly, they showed that the reconstruction of a mouse trajectory was feasible only with the activity of the astrocytic population.
Consequently, by combining the theoretical framework of the modular theory with the experimental findings that support the existence of subsets of astrocytes that activate in a behavioral paradigm, we detect there is a missing concept to refer to these groups of working astrocytes and, herein, we propose the term astrocytic ensembles (see Figure 3). This conceptualization will aid the phenomenon in being formulated into testable hypotheses and correlated with specific variables of interest. Lastly, an agreement in the understanding of such a key concept opens up a horizon of questions about the mechanism behind ensemble selection and whether it happens in all behavioral conditions, how the behavioral ensembles operate in an unconditioned neutral context, or whether gradients of positive or negative valences are encoded. The experimental approaches to these responses implicate the selective labeling of implicated ensembles and their reactivation or inhibition.
3.2. Could Astrocytes Undergo Specialization?
The study of astrocytes is conditioned by their heterogeneous nature [75,153,154,155]. Initially, two main astrocyte types were characterized, the fibrous and protoplasmic, found in white and gray matter, respectively [156]. However, currently, there is evidence showing the existence of several phenotypes with determined molecular profiles that differentially express ion channels [157], gap junction proteins [158], glutamate transporters [159,160], or receptors [161]. The individual cell identities are shaped by intrinsic factors (developmental patterning that results in a unique combination of transcription factors [156,162,163]) and extrinsic factors (the local environment drives functional adaptations, i.e., the mature phenotype is highly determined by the signaling from adjacent neurons, despite coming from the same progenitor pool [65,164]). What is more, the different phenotypes have been shown to be located according to the brain region’s functionality [65,78,163,165]. Therefore, astroglial cells exhibit high heterogeneity [154,155], not only in their morphology [65,75] and developmental origin [156,162,163] but also in their gene expression profiles [18,75,153] and functionalities [6,141,154,165], both within the same region and across different areas [65]. While the functional implications of this diversity in various neural circuits is a current matter of debate, astrocytes play critical roles in the formation, maturation, and maintenance of synapses in diverse brain regions [14,22,25], both in physiological and pathological conditions [75,166]. Astroglia are constantly facing changing environments, and this environmental degree of complexity increases throughout the phylogenetic scale. In this way, compared to rodent astrocytes, human astrocytes are larger, structurally more complex, and show a higher phenotypic diversity, and their Ca2+ signaling travels at larger velocities [167]. These features provide the human brain with increased functional competence and more complex computational mechanisms [167,168,169].
In the case of neurons, it is well known that they are specialized cells with defined functions [131,170], implying that they have well-conserved roles and can perform the same role in different physiological situations. By contrast, astrocytes show very variable capacities, which permits them to perform very different roles in different physiological situations. However, under the assumption that the difference between heterogeneity and specialization lies in the attribution of a functional role, we deduce that astrocytic ensembles are likely to be specialized for a determined function. In this line, how could, therefore, the dynamic nature of astrocytes’ cell identity be explained? Do ensembles lose their capacity to operate with such a margin of versatility? How is specialization controlled, and when is it triggered? Contemplating the limitations for capturing relevant physiological sub-populations of astrocytes and exploring alternative parameters to define functionally homogeneous populations to study their impact on neural circuits reveals the ever-evolving nature of neuroscience research.
3.3. The In Vivo Study of Astrocytic Processing in Behavior
There is no doubt that astrocytes play important roles in the production of cognition, emotion, sensory, and motor processing that give rise to complex behaviors [140,171]. The current knowledge of astrocytic calcium signaling mainly comes from studies of responses to specific ex vivo or in vitro stimulation of different brain regions [154], but in order to understand the correlation between behavior and astrocytic activity, in vivo studies in awake animals are requisite. Until now, the main limitation for attempting the in vivo tracking of astroglial Ca2+ activity has been the lack of available methods, and most studies were based on anesthetized animals [105], which have been useful to determine some encoded patterns of the processing strategies but leave unattended the physiological condition.
Astrocytes activate through intracellular calcium increases, which are their basic unit for communication [4,15]. Multiple parameters of the signaling encode information, such as the amplitude, duration, frequency, transient onset, or spatial pattern [172,173], which provides the astrocytes with a very high versatility of operation. For instance, studies in anesthetized mice have shown that astrocytes in the somatosensory cortex respond to foot stimulation by modulating their signal amplitude [174] or that the frequency of intense whisker stimulation is encoded in the number of recruited astrocytes in the barrel cortex [175]. However, general anesthesia has been shown to have a significant effect on calcium signaling, which can affect calcium transients and spontaneous activity [176,177,178]. To overcome these limitations, studies using GECIs and fiber optic recordings have been conducted in freely moving animals, bringing the challenge of interpreting the signals, as the recordings contain information from non-relevant sources. The fact that astrocytic activity could be masked in the awake in vivo state has been addressed by Curreli et al. [152]. They demonstrated that the signaling of hippocampal astrocytes encodes information regarding the animal position and that this information is complementary to that of neural place cells. Additionally, Doron et al. showed that hippocampal astrocytes ciphered a reward location only when the spatial context was familiar. Even more, they demonstrated that the activity of the astroglial population was sufficient to reconstruct trajectories in known environments [2]. In summary, there is evidence showing how astrocytic signals encode information complementarily to neurons and support their involvement in the refinement mechanisms of neuronal responses with integrative (and stabilizing) properties [12].
Therefore, these studies suggest that the existence of astrocytic networks enables complex integration in the cognitive function of spatial information coding, including the processing of episodic memory linked to specific locations and contexts. Importantly, these results reflect physiological astrocytic responses. Additionally, the ability of astrocytes to reflect the activity of neighboring neurons [175,178] and encode complementary information [152] may depend on the type of response elicited, either sensory-evoked or event-based. Further research is needed to better understand these differences and to monitor specific cell identities in specific networks for different tasks.
The potential to comprehend the computational principles of astrocytes is now within reach. Can we anticipate a uniform calculation between inputs and outputs within the same framework? How would the stimulation of astrocytes specifically impact the dynamics of neuronal efferents? Is there a noticeable behavioral effect after stimulating astrocytic ensembles in vivo? In light of the studies discussed in Section 3.1, where specific neuronal networks were selectively stimulated, resulting in a motor outcome [137,138,139], we hypothesize that triggering the selective activation of astrocytic ensembles could imply or enhance a behavioral circumstance, probably in the scope of a learned task due to the refinement and integrative character of their responses. For this, it is remarkable that the necessity of having new tools allows the tracking of the Ca2+ levels of astrocytic activities.
4. Concluding Remarks
Astrocytes are active cells implicated in the computational procedures of the brain. Based on recent studies showing that astrocytes work as networked systems and the modular nature of the brain, we propose the term “ensemble” to describe groups of astrocytes that come together to carry out a specific function. This concept opens up the possibility that these ensembles may develop specialized skills for performing that function. By making a systematic comparison of neurons and astrocytes, our argument rotates around the requirement of increasing the number of experimental studies that consider the existence of astrocytic ensembles since the number of currently available ones is very limited and computational descriptions need to be based on or supported by experimental facts. Despite the well-known fact that astrocytes play a role in animal behavior—their activity disruption being a cause of disease—how the coding of their responses occurs is not yet determined.
It is important to emphasize that astrocytes interact with other glial cells, including microglia [179,180,181] and oligodendrocytes [182,183], and also with immune cells [184,185]. These interactions play a crucial role in both normal and disease conditions [185,186,187]. Recent experimental evidence has confirmed the significant role of astrocyte-microglia communication [186], which has therapeutic potential for neurological disorders such as experimental autoimmune encephalomyelitis (EAE) and multiple sclerosis (MS) [188,189]. Therefore, studying the mechanisms of communication within astrocytic engrams and other glial cells will help us understand disease-promoting glial responses and develop new treatments. Similarly, the finding that astrocytes can be reprogrammed into neuronal progenitors and mature neurons in the CNS of adult mammals [190,191,192,193,194,195] raises questions about the role that ensembles may play in this orchestration. The molecular mechanisms, regulatory networks, and potential astrocytic ensembles that trigger this reprogramming are not yet clear, which could bring new insights into regeneration-based therapies.
The fact that we currently have methods to study astrocytic ensembles in mice and rats is not yet promising in the study of the human brain astrocyte ensembles due to their being highly invasive. It is, however, worth mentioning that some of the studies herein referenced were performed in humans or in non-human primates [104] due to their being non-invasive [68,136] or were with the informed consent of patients in surgeries of epilepsy cases and tumors [82].
The task of deciphering the results generated by neuronal–astrocyte networks is complex, involving the systematization of mechanisms or the creation of approximate fitting models. By adopting a top-down perspective and concentrating on the dynamics of astrocyte networks, it is possible to make headway in identifying the functional correlates of astrocyte responses. Such insights will establish the groundwork for more sophisticated investigations and contribute towards comprehending potential therapeutic targets aimed at repairing and restoring normal function in disease.
Author Contributions
L.D. and M.N. wrote the article. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ayudante de investigación de la Comunidad de Madrid 2020 fellowship (PEJ-2020-AI/BMD-18245) to L.D.; grants from Ministerio de Ciencia e Innovación (PID2021-122586NB-I00) and International Society for Neurochemistry ISN Career Development Grant (CDG) to M.N.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to E. Martin and G. Perea for their helpful comments. The authors would like to express their regret to any colleagues whose significant contributions were not referenced explicitly.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yu, X. Hippocampal Astrocytes Represent Navigation Space. PLoS Biol. 2022, 20, e3001568. [Google Scholar] [CrossRef] [PubMed]
- Doron, A.; Rubin, A.; Benmelech-Chovav, A.; Benaim, N.; Carmi, T.; Refaeli, R.; Novick, N.; Kreisel, T.; Ziv, Y.; Goshen, I. Hippocampal Astrocytes Encode Reward Location. Nature 2022, 609, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Serra, I.; Esparza, J.; Delgado, L.; Martín-Monteagudo, C.; Puigròs, M.; Podlesniy, P.; Trullás, R.; Navarrete, M. Ca2+-Modulated Photoactivatable Imaging Reveals Neuron-Astrocyte Glutamatergic Circuitries within the Nucleus Accumbens. Nat. Commun. 2022, 13, 5272. [Google Scholar] [CrossRef] [PubMed]
- Volterra, A.; Liaudet, N.; Savtchouk, I. Astrocyte Ca2+ Signalling: An Unexpected Complexity. Nat. Rev. Neurosci. 2014, 15, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium Signalling: Dynamics, Homeostasis and Remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; McCarthy, K.D. Astrocyte Calcium Signaling: From Observations to Functions and the Challenges Therein. Cold Spring Harb. Perspect. Biol. 2015, 7, a020404. [Google Scholar] [CrossRef]
- Nett, W.J.; Oloff, S.H.; Mccarthy, K.D. Hippocampal Astrocytes In Situ Exhibit Calcium Oscillations That Occur Independent of Neuronal Activity. J. Neurophysiol. 2002, 87, 528–537. [Google Scholar] [CrossRef]
- Ding, F.; O’Donnell, J.; Thrane, A.S.; Zeppenfeld, D.; Kang, H.; Xie, L.; Wang, F.; Nedergaard, M. A1-Adrenergic Receptors Mediate Coordinated Ca2+ Signaling of Cortical Astrocytes in Awake, Behaving Mice. Cell Calcium 2013, 54, 387–394. [Google Scholar] [CrossRef]
- Cornell-Bell, A.H.; Finkbeiner, S.M.; Cooper, M.S.; Smith, S.J. Glutamate Induces Calcium Waves in Cultured Astrocytes: Long-Range Glial Signaling. Science 1990, 247, 470–473. [Google Scholar] [CrossRef]
- Palygin, O.; Lalo, U.; Verkhratsky, A.; Pankratov, Y. Ionotropic NMDA and P2X1/5 Receptors Mediate Synaptically Induced Ca2+ Signalling in Cortical Astrocytes. Cell Calcium 2010, 48, 225–231. [Google Scholar] [CrossRef]
- Araque, A.; Carmignoto, G.; Haydon, P.G. Dynamic Signaling between Astrocytes and Neurons. Annu. Rev. Physiol. 2001, 63, 795–813. [Google Scholar] [CrossRef] [PubMed]
- Araque, A.; Carmignoto, G.; Haydon, P.G.; Oliet, S.H.R.; Robitaille, R.; Volterra, A. Gliotransmitters Travel in Time and Space. Neuron 2014, 81, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Halassa, M.M.; Haydon, P.G. Integrated Brain Circuits: Astrocytic Networks Modulate Neuronal Activity and Behavior. Annu. Rev. Physiol. 2010, 72, 335. [Google Scholar] [CrossRef] [PubMed]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite Synapses: Astrocytes Process and Control Synaptic Information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef]
- Volterra, A.; Meldolesi, J. Astrocytes, from Brain Glue to Communication Elements: The Revolution Continues. Nat. Rev. Neurosci. 2005, 6, 626–640. [Google Scholar] [CrossRef]
- Aguado, F.; Espinosa-Parrilla, J.F.; Carmona, M.A.; Soriano, E. Neuronal Activity Regulates Correlated Network Properties of Spontaneous Calcium Transients in Astrocytes In Situ. J. Neurosci. 2002, 22, 9430–9444. [Google Scholar] [CrossRef]
- Chung, W.S.; Allen, N.J.; Eroglu, C. Astrocytes Control Synapse Formation, Function, and Elimination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020370. [Google Scholar] [CrossRef]
- Baldwin, K.T.; Eroglu, C. Molecular Mechanisms of Astrocyte-Induced Synaptogenesis. Curr. Opin. Neurobiol. 2017, 45, 113–120. [Google Scholar] [CrossRef]
- Allen, N.J.; Eroglu, C. Cell Biology of Astrocyte-Synapse Interactions. Neuron 2017, 96, 697–708. [Google Scholar] [CrossRef]
- Stork, T.; Sheehan, A.; Tasdemir-Yilmaz, O.E.; Freeman, M.R. Neuron-Glia Interactions through the Heartless FGF Receptor Signaling Pathway Mediate Morphogenesis of Drosophila Astrocytes. Neuron 2014, 83, 388–403. [Google Scholar] [CrossRef]
- Oberheim, N.A.; Wang, X.; Goldman, S.; Nedergaard, M. Astrocytic Complexity Distinguishes the Human Brain. Trends Neurosci. 2006, 29, 547–553. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite Synapses: Glia, the Unacknowledged Partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Kleim, J.A.; Markham, J.A.; Vij, K.; Freese, J.L.; Ballard, D.H.; Greenough, W.T. Motor Learning Induces Astrocytic Hypertrophy in the Cerebellar Cortex. Behav. Brain Res. 2007, 178, 244–249. [Google Scholar] [CrossRef]
- Allen, N.J. Astrocyte Regulation of Synaptic Behavior. Annu. Rev. Cell Dev. Biol. 2014, 30, 439–463. [Google Scholar] [CrossRef] [PubMed]
- Bernardinelli, Y.; Muller, D.; Nikonenko, I. Astrocyte-Synapse Structural Plasticity. Neural Plast. 2014, 2014, 1. [Google Scholar] [CrossRef]
- Arizono, M.; Inavalli, V.V.G.K.; Bancelin, S.; Fernández-Monreal, M.; Nägerl, U.V. Super-Resolution Shadow Imaging Reveals Local Remodeling of Astrocytic Microstructures and Brain Extracellular Space after Osmotic Challenge. Glia 2021, 69, 1605–1613. [Google Scholar] [CrossRef]
- Theodosis, D.T.; Poulain, D.A.; Oliet, S.H.R. Activity-Dependent Structural and Functional Plasticity of Astrocyte-Neuron Interactions. Physiol. Rev. 2008, 88, 983–1008. [Google Scholar] [CrossRef] [PubMed]
- Hirrlinger, J.; Hülsmann, S.; Kirchhoff, F. Astroglial Processes Show Spontaneous Motility at Active Synaptic Terminals In Situ. Eur. J. Neurosci. 2004, 20, 2235–2239. [Google Scholar] [CrossRef]
- Xu-Friedman, M.A.; Harris, K.M.; Regehr, W.G. Three-Dimensional Comparison of Ultrastructural Characteristics at Depressing and Facilitating Synapses onto Cerebellar Purkinje Cells. J. Neurosci. 2001, 21, 6666–6672. [Google Scholar] [CrossRef]
- Bernardinelli, Y.; Randall, J.; Janett, E.; Nikonenko, I.; König, S.; Jones, E.V.; Flores, C.E.; Murai, K.K.; Bochet, C.G.; Holtmaat, A.; et al. Activity-Dependent Structural Plasticity of Perisynaptic Astrocytic Domains Promotes Excitatory Synapse Stability. Curr. Biol. 2014, 24, 1679–1688. [Google Scholar] [CrossRef]
- Han, J.; Kesner, P.; Metna-Laurent, M.; Duan, T.; Xu, L.; Georges, F.; Koehl, M.; Abrous, D.N.; Mendizabal-Zubiaga, J.; Grandes, P.; et al. Acute Cannabinoids Impair Working Memory through Astroglial CB1 Receptor Modulation of Hippocampal LTD. Cell 2012, 148, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Duarte, A.; Roberts, A.J.; Ouyang, K.; Sejnowski, T.J. Impairments in Remote Memory Caused by the Lack of Type 2 IP3 Receptors. Glia 2019, 67, 1976–1989. [Google Scholar] [CrossRef]
- Sherwood, M.W.; Arizono, M.; Panatier, A.; Mikoshiba, K.; Oliet, S.H.R. Astrocytic IP3Rs: Beyond IP3R2. Front. Cell Neurosci. 2021, 15, 695817. [Google Scholar] [CrossRef] [PubMed]
- Mederos, S.; Sánchez-Puelles, C.; Esparza, J.; Valero, M.; Ponomarenko, A.; Perea, G. GABAergic Signaling to Astrocytes in the Prefrontal Cortex Sustains Goal-Directed Behaviors. Nat. Neurosci. 2021, 24, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, M.; Cuartero, M.I.; Palenzuela, R.; Draffin, J.E.; Konomi, A.; Serra, I.; Colié, S.; Castaño-Castaño, S.; Hasan, M.T.; Nebreda, Á.R.; et al. Astrocytic P38α MAPK Drives NMDA Receptor-Dependent Long-Term Depression and Modulates Long-Term Memory. Nat. Commun. 2019, 10, 2968. [Google Scholar] [CrossRef]
- Lutz, S.E.; Zhao, Y.; Gulinello, M.; Lee, S.C.; Raine, C.S.; Brosnan, C.F. Deletion of Astrocyte Connexins 43 and 30 Leads to a Dysmyelinating Phenotype and Hippocampal CA1 Vacuolation. J. Neurosci. 2009, 29, 7743–7752. [Google Scholar] [CrossRef] [PubMed]
- Stogsdill, J.A.; Ramirez, J.; Liu, D.; Kim, Y.-H.; Baldwin, K.T.; Enustun, E.; Ejikeme, T.; Ji, R.-R.; Eroglu, C. Astrocytic Neuroligins Control Astrocyte Morphogenesis and Synaptogenesis. Nature 2017, 551, 192–197. [Google Scholar] [CrossRef]
- Kol, A.; Adamsky, A.; Groysman, M.; Kreisel, T.; London, M.; Goshen, I. Astrocytes Contribute to Remote Memory Formation by Modulating Hippocampal-Cortical Communication during Learning. Nat. Neurosci. 2020, 23, 1229–1239. [Google Scholar] [CrossRef]
- Vaccari Cardoso, B.; Shevelkin, A.V.; Terrillion, C.; Mychko, O.; Mosienko, V.; Kasparov, S.; Pletnikov, M.V.; Teschemacher, A.G. Reducing L-Lactate Release from Hippocampal Astrocytes by Intracellular Oxidation Increases Novelty Induced Activity in Mice. Glia 2021, 69, 1241–1250. [Google Scholar] [CrossRef]
- Beltrán-Castillo, S.; Olivares, M.J.; Contreras, R.A.; Zúñiga, G.; Llona, I.; Von Bernhardi, R.; Eugenín, J.L. D-Serine Released by Astrocytes in Brainstem Regulates Breathing Response to CO2 Levels. Nat. Commun. 2017, 8, 838. [Google Scholar] [CrossRef]
- Bhandare, A.; van de Wiel, J.; Roberts, R.; Braren, I.; Huckstepp, R.; Dale, N. Analyzing the Brainstem Circuits for Respiratory Chemosensitivity in Freely Moving Mice. ELife 2022, 11, e70671. [Google Scholar] [CrossRef] [PubMed]
- Czeisler, C.M.; Silva, T.M.; Fair, S.R.; Liu, J.; Tupal, S.; Kaya, B.; Cowgill, A.; Mahajan, S.; Silva, P.E.; Wang, Y.; et al. The Role of PHOX2B-derived Astrocytes in Chemosensory Control of Breathing and Sleep Homeostasis. J. Physiol. 2019, 597, 2225. [Google Scholar] [CrossRef]
- Okada, Y.; Sasaki, T.; Oku, Y.; Takahashi, N.; Seki, M.; Ujita, S.; Tanaka, K.F.; Matsuki, N.; Ikegaya, Y. Preinspiratory Calcium Rise in Putative Pre-Bötzinger Complex Astrocytes. J. Physiol. 2012, 590, 4933–4944. [Google Scholar] [CrossRef] [PubMed]
- Oku, Y.; Fresemann, J.; Miwakeichi, F.; Hülsmann, S. Respiratory Calcium Fluctuations in Low-Frequency Oscillating Astrocytes in the Pre-Bötzinger Complex. Respir. Physiol. Neurobiol. 2016, 226, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Bardoni, R.; Ghirri, A.; Zonta, M.; Betelli, C.; Vitale, G.; Ruggieri, V.; Sandrini, M.; Carmignoto, G. Glutamate-Mediated Astrocyte-to-Neuron Signalling in the Rat Dorsal Horn. J. Physiol. 2010, 588, 831–846. [Google Scholar] [CrossRef]
- Broadhead, M.J.; Miles, G.B. Bi-Directional Communication Between Neurons and Astrocytes Modulates Spinal Motor Circuits. Front. Cell. Neurosci. 2020, 14, 30. [Google Scholar] [CrossRef]
- Dombeck, D.A.; Khabbaz, A.N.; Collman, F.; Adelman, T.L.; Tank, D.W. Imaging Large-Scale Neural Activity with Cellular Resolution in Awake, Mobile Mice. Neuron 2007, 56, 43–57. [Google Scholar] [CrossRef]
- Hegyi, Z.; Oláh, T.; Koszeghy, Á.; Pisticelli, F.; Holló, K.; Pál, B.; Csernoch, L.; di Marzo, V.; Antal, M. CB 1 Receptor Activation Induces Intracellular Ca2+ Mobilization and 2-Arachidonoylglycerol Release in Rodent Spinal Cord Astrocytes. Sci. Rep. 2018, 8, 10562. [Google Scholar] [CrossRef]
- Sekiguchi, K.J.; Shekhtmeyster, P.; Merten, K.; Arena, A.; Cook, D.; Hoffman, E.; Ngo, A.; Nimmerjahn, A. Imaging Large-Scale Cellular Activity in Spinal Cord of Freely Behaving Mice. Nat. Commun. 2016, 7, 11450. [Google Scholar] [CrossRef]
- Brocard, F.; Verdier, D.; Arsenault, I.; Lund, J.P.; Kolta, A. Emergence of Intrinsic Bursting in Trigeminal Sensory Neurons Parallels the Acquisition of Mastication in Weanling Rats. J. Neurophysiol. 2006, 96, 2410–2424. [Google Scholar] [CrossRef]
- Condamine, S.; Lavoie, R.; Verdier, D.; Kolta, A. Functional Rhythmogenic Domains Defined by Astrocytic Networks in the Trigeminal Main Sensory Nucleus. Glia 2018, 66, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Morquette, P.; Verdier, D.; Kadala, A.; Féthière, J.; Philippe, A.G.; Robitaille, R.; Kolta, A. An Astrocyte-Dependent Mechanism for Neuronal Rhythmogenesis. Nat. Neurosci. 2015, 18, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Damulewicz, M.; Doktór, B.; Baster, Z.; Pyza, E. The Role of Glia Clocks in the Regulation of Sleep in Drosophila Melanogaster. J. Neurosci. 2022, 42, 6848–6860. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, M.; Edwards, M.D.; Patton, A.P.; Smyllie, N.J.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Cell-Autonomous Clock of Astrocytes Drives Circadian Behavior in Mammals. Science 2019, 363, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, M.; Patton, A.P.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Astrocytes Control Circadian Timekeeping in the Suprachiasmatic Nucleus via Glutamatergic Signaling. Neuron 2017, 93, 1420–1435.e5. [Google Scholar] [CrossRef] [PubMed]
- Patton, A.P.; Smyllie, N.J.; Chesham, J.E.; Hastings, M.H. Astrocytes Sustain Circadian Oscillation and Bidirectionally Determine Circadian Period, But Do Not Regulate Circadian Phase in the Suprachiasmatic Nucleus. J. Neurosci. 2022, 42, 5522–5537. [Google Scholar] [CrossRef]
- Lee, H.S.; Ghetti, A.; Pinto-Duarte, A.; Wang, X.; Dziewczapolski, G.; Galimi, F.; Huitron-Resendiz, S.; Piña-Crespo, J.C.; Roberts, A.J.; Verma, I.M.; et al. Astrocytes Contribute to Gamma Oscillations and Recognition Memory. Proc. Natl. Acad. Sci. USA 2014, 111, E3343. [Google Scholar] [CrossRef]
- Hirrlinger, J.; Nimmerjahn, A. A Perspective on Astrocyte Regulation of Neural Circuit Function and Animal Behavior. Glia 2022, 70, 1554–1580. [Google Scholar] [CrossRef]
- Yu, X.; Nagai, J.; Khakh, B.S. Improved Tools to Study Astrocytes. Nat. Rev. Neurosci. 2020, 21, 121–138. [Google Scholar] [CrossRef]
- Gorzo, K.A.; Gordon, G.R. Photonics Tools Begin to Clarify Astrocyte Calcium Transients. Neurophotonics 2022, 9, 021907. [Google Scholar] [CrossRef]
- Gorina, Y.V.; Salmina, A.B.; Erofeev, A.I.; Gerasimov, E.I.; Bolshakova, A.V.; Balaban, P.M.; Bezprozvanny, I.B.; Vlasova, O.L. Astrocyte Activation Markers. Biochemistry 2022, 87, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Bindocci, E.; Savtchouk, I.; Liaudet, N.; Becker, D.; Carriero, G.; Volterra, A. Three-Dimensional Ca2+ Imaging Advances Understanding of Astrocyte Biology. Science 2017, 356, eaai8185. [Google Scholar] [CrossRef] [PubMed]
- Bushong, E.A.; Martone, M.E.; Jones, Y.Z.; Ellisman, M.H. Protoplasmic Astrocytes in CA1 Stratum Radiatum Occupy Separate Anatomical Domains. J. Neurosci. 2002, 22, 183–192. [Google Scholar] [CrossRef]
- Holt, L.M.; Hernandez, R.D.; Pacheco, N.L.; Torres Ceja, B.; Hossain, M.; Olsen, M.L. Astrocyte Morphogenesis Is Dependent on BDNF Signaling via Astrocytic TrkB.T1. Elife 2019, 8, e44667. [Google Scholar] [CrossRef]
- Lanjakornsiripan, D.; Pior, B.J.; Kawaguchi, D.; Furutachi, S.; Tahara, T.; Katsuyama, Y.; Suzuki, Y.; Fukazawa, Y.; Gotoh, Y. Layer-Specific Morphological and Molecular Differences in Neocortical Astrocytes and Their Dependence on Neuronal Layers. Nat. Commun. 2018, 9, 1623. [Google Scholar] [CrossRef]
- Lu, X.H.; Yang, X.W. Genetically-Directed Sparse Neuronal Labeling in BAC Transgenic Mice through Mononucleotide Repeat Frameshift. Sci. Rep. 2017, 7, 43915. [Google Scholar] [CrossRef] [PubMed]
- Mathiisen, T.M.; Lehre, K.P.; Danbolt, N.C.; Ottersen, O.P. The Perivascular Astroglial Sheath Provides a Complete Covering of the Brain Microvessels: An Electron Microscopic 3D Reconstruction. Glia 2010, 58, 1094–1103. [Google Scholar] [CrossRef]
- Witcher, M.R.; Park, Y.D.; Lee, M.R.; Sharma, S.; Harris, K.M.; Kirov, S.A. Three-Dimensional Relationships Between Perisynaptic Astroglia and Human Hippocampal Synapses. Glia 2010, 58, 572. [Google Scholar] [CrossRef]
- Calì, C.; Agus, M.; Kare, K.; Boges, D.J.; Lehväslaiho, H.; Hadwiger, M.; Magistretti, P.J. 3D Cellular Reconstruction of Cortical Glia and Parenchymal Morphometric Analysis from Serial Block-Face Electron Microscopy of Juvenile Rat. Prog. Neurobiol. 2019, 183, 101696. [Google Scholar] [CrossRef]
- Watanabe, K.; Takeishi, H.; Hayakawa, T.; Sasaki, H. Three-Dimensional Organization of the Perivascular Glial Limiting Membrane and Its Relationship with the Vasculature: A Scanning Electron Microscope Study. Okajimas Folia Anat. Jpn. 2010, 87, 109–121. [Google Scholar] [CrossRef]
- Ventura, R.; Harris, K.M. Three-Dimensional Relationships between Hippocampal Synapses and Astrocytes. J. Neurosci. 1999, 19, 6897–6906. [Google Scholar] [CrossRef] [PubMed]
- Patrushev, I.; Gavrilov, N.; Turlapov, V.; Semyanov, A. Subcellular Location of Astrocytic Calcium Stores Favors Extrasynaptic Neuron-Astrocyte Communication. Cell Calcium 2013, 54, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Refaeli, R.; Goshen, I. Investigation of Spatial Interaction between Astrocytes and Neurons in Cleared Brains. J. Vis. Exp. 2022, 2022, e63679. [Google Scholar] [CrossRef]
- Octeau, J.C.; Chai, H.; Jiang, R.; Bonanno, S.L.; Martin, K.C.; Khakh, B.S. An Optical Neuron-Astrocyte Proximity Assay at Synaptic Distance Scales. Neuron 2018, 98, 49–66.e9. [Google Scholar] [CrossRef]
- Endo, F.; Kasai, A.; Soto, J.S.; Yu, X.; Qu, Z.; Hashimoto, H.; Gradinaru, V.; Kawaguchi, R.; Khakh, B.S. Molecular Basis of Astrocyte Diversity and Morphology across the CNS in Health and Disease. Science 2022, 378, 1631–1635. [Google Scholar] [CrossRef]
- Lavialle, M.; Aumann, G.; Anlauf, E.; Pröls, F.; Arpin, M.; Derouiche, A. Structural Plasticity of Perisynaptic Astrocyte Processes Involves Ezrin and Metabotropic Glutamate Receptors. Proc. Natl. Acad. Sci. USA 2011, 108, 12915–12919. [Google Scholar] [CrossRef]
- Perez-Alvarez, A.; Navarrete, M.; Covelo, A.; Martin, E.D.; Araque, A. Structural and Functional Plasticity of Astrocyte Processes and Dendritic Spine Interactions. J. Neurosci. 2014, 34, 12738–12744. [Google Scholar] [CrossRef]
- Chai, H.; Diaz-Castro, B.; Shigetomi, E.; Monte, E.; Octeau, J.C.; Yu, X.; Cohn, W.; Rajendran, P.S.; Vondriska, T.M.; Whitelegge, J.P.; et al. Neural Circuit-Specialized Astrocytes: Transcriptomic, Proteomic, Morphological, and Functional Evidence. Neuron 2017, 95, 531–549.e9. [Google Scholar] [CrossRef]
- Adil, A.; Kumar, V.; Jan, A.T.; Asger, M. Single-Cell Transcriptomics: Current Methods and Challenges in Data Acquisition and Analysis. Front. Neurosci. 2021, 15, 591122. [Google Scholar] [CrossRef]
- Cahoy, J.D.; Emery, B.; Kaushal, A.; Foo, L.C.; Zamanian, J.L.; Christopherson, K.S.; Xing, Y.; Lubischer, J.L.; Krieg, P.A.; Krupenko, S.A.; et al. A Transcriptome Database for Astrocytes, Neurons, and Oligodendrocytes: A New Resource for Understanding Brain Development and Function. J. Neurosci. 2008, 28, 264–278. [Google Scholar] [CrossRef]
- Barres, B.A.; Silverstein, B.E.; Corey, D.R.; Chun, L.L.Y. Immunological, Morphological, and Electrophysiological Variation among Retinal Ganglion Cells Purified by Panning. Neuron 1988, 1, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sloan, S.A.; Clarke, L.E.; Caneda, C.; Plaza, C.A.; Blumenthal, P.D.; Vogel, H.; Steinberg, G.K.; Edwards, M.S.B.; Li, G.; et al. Purification and Characterization of Progenitor and Mature Human Astrocytes Reveals Transcriptional and Functional Differences with Mouse. Neuron 2016, 89, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Holt, L.M.; Stoyanof, S.T.; Olsen, M.L. Magnetic Cell Sorting for In Vivo and In Vitro Astrocyte, Neuron, and Microglia Analysis. Curr. Protoc. Neurosci. 2019, 88, e71. [Google Scholar] [CrossRef]
- Holt, L.M.; Olsen, M.L. Novel Applications of Magnetic Cell Sorting to Analyze Cell-Type Specific Gene and Protein Expression in the Central Nervous System. PLoS ONE 2016, 11, e0150290. [Google Scholar] [CrossRef] [PubMed]
- Heiman, M.; Schaefer, A.; Gong, S.; Peterson, J.D.; Day, M.; Ramsey, K.E.; Suárez-Fariñas, M.; Schwarz, C.; Stephan, D.A.; Surmeier, D.J.; et al. A Translational Profiling Approach for the Molecular Characterization of CNS Cell Types. Cell 2008, 135, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.P.; Dougherty, J.D.; Heiman, M.; Schmidt, E.F.; Stevens, T.R.; Ma, G.; Bupp, S.; Shrestha, P.; Shah, R.D.; Doughty, M.L.; et al. Application of a Translational Profiling Approach for the Comparative Analysis of CNS Cell Types. Cell 2008, 135, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Sanz, E.; Bean, J.C.; Carey, D.P.; Quintana, A.; McKnight, G.S. RiboTag: Ribosomal Tagging Strategy to Analyze Cell-Type-Specific MRNA Expression In Vivo. Curr. Protoc. Neurosci. 2019, 88, e77. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Ferrer, I.; Khakh, B.S.; Díaz-Castro, B. Cell-Specific RNA Purification to Study Translatomes of Mouse Central Nervous System. STAR Protoc. 2022, 3, 101397. [Google Scholar] [CrossRef]
- Rao, A.; Barkley, D.; França, G.S.; Yanai, I. Exploring Tissue Architecture Using Spatial Transcriptomics. Nature 2021, 596, 211. [Google Scholar] [CrossRef]
- Dries, R.; Chen, J.; Del Rossi, N.; Khan, M.M.; Sistig, A.; Yuan, G.C. Advances in Spatial Transcriptomic Data Analysis. Genome Res. 2021, 31, 1706. [Google Scholar] [CrossRef]
- Williams, C.G.; Lee, H.J.; Asatsuma, T.; Vento-Tormo, R.; Haque, A. An Introduction to Spatial Transcriptomics for Biomedical Research. Genome Med 2022, 14, 68. [Google Scholar] [CrossRef] [PubMed]
- Ståhl, P.L.; Salmén, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; Giacomello, S.; Asp, M.; Westholm, J.O.; Huss, M.; et al. Visualization and Analysis of Gene Expression in Tissue Sections by Spatial Transcriptomics. Science 2016, 353, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Merritt, C.R.; Ong, G.T.; Church, S.E.; Barker, K.; Danaher, P.; Geiss, G.; Hoang, M.; Jung, J.; Liang, Y.; McKay-Fleisch, J.; et al. Multiplex Digital Spatial Profiling of Proteins and RNA in Fixed Tissue. Nat. Biotechnol. 2020, 38, 586–599. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Bhatt, R.; Brown, C.; Brown, E.A.; Buhr, D.L.; Chantranuvatana, K.; Danaher, P.; Dunaway, D.; Garrison, R.G.; Geiss, G.; et al. High-Plex Multiomic Analysis in FFPE at Subcellular Level by Spatial Molecular Imaging. Nat. Biotechnol. 2022, 40, 1794–1806. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A New Generation of Ca2+ Indicators with Greatly Improved Fluorescence Properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [CrossRef] [PubMed]
- Minta, A.; Kao, J.P.Y.; Tsien, R.Y. Fluorescent Indicators for Cytosolic Calcium Based on Rhodamine and Fluorescein Chromophores. J. Biol. Chem. 1989, 264, 8171–8178. [Google Scholar] [CrossRef] [PubMed]
- Nakai, J.; Ohkura, M.; Imoto, K. A High Signal-to-Noise Ca(2+) Probe Composed of a Single Green Fluorescent Protein. Nat. Biotechnol. 2001, 19, 137–141. [Google Scholar] [CrossRef]
- Redolfi, N.; Greotti, E.; Zanetti, G.; Hochepied, T.; Fasolato, C.; Pendin, D.; Pozzan, T. A New Transgenic Mouse Line for Imaging Mitochondrial Calcium Signals. Function 2021, 2, zqab012. [Google Scholar] [CrossRef]
- Srinivasan, R.; Lu, T.Y.; Chai, H.; Xu, J.; Huang, B.S.; Golshani, P.; Coppola, G.; Khakh, B.S. New Transgenic Mouse Lines for Selectively Targeting Astrocytes and Studying Calcium Signals in Astrocyte Processes In Situ and In Vivo. Neuron 2016, 92, 1181–1195. [Google Scholar] [CrossRef]
- Zhuo, L.; Theis, M.; Alvarez-Maya, I.; Brenner, M.; Willecke, K.; Messing, A. HGFAP-Cre Transgenic Mice for Manipulation of Glial and Neuronal Function In Vivo. Genesis 2001, 31, 85–94. [Google Scholar] [CrossRef]
- Vives, V.; Alonso, G.; Solal, A.C.; Joubert, D.; Legraverend, C. Visualization of S100B-Positive Neurons and Glia in the Central Nervous System of EGFP Transgenic Mice. J. Comp. Neurol. 2003, 457, 404–419. [Google Scholar] [CrossRef]
- Nolte, C.; Matyash, M.; Pivneva, T.; Schipke, C.G.; Ohlemeyer, C.; Hanisch, U.K.; Kirchhoff, F.; Kettenmann, H. GFAP Promoter-Controlled EGFP-Expressing Transgenic Mice: A Tool to Visualize Astrocytes and Astrogliosis in Living Brain Tissue. Glia 2001, 33, 72–86. [Google Scholar] [CrossRef]
- Shigetomi, E.; Bushong, E.A.; Haustein, M.D.; Tong, X.; Jackson-Weaver, O.; Kracun, S.; Xu, J.; Sofroniew, M.V.; Ellisman, M.H.; Khakh, B.S. Imaging Calcium Microdomains within Entire Astrocyte Territories and Endfeet with GCaMPs Expressed Using Adeno-Associated Viruses. J. Gen. Physiol. 2013, 141, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, P.A.; Bland, R.J.; Mouravlev, A.; Young, D.; During, M.J. Efficient Gene Delivery and Selective Transduction of Glial Cells in the Mammalian Brain by AAV Serotypes Isolated from Nonhuman Primates. Mol. Ther. 2009, 17, 1692–1702. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; He, W.; Yang, C.; Li, J.; Jian, T.; Liang, S.; Chen, T.; Feng, H.; Chen, X.; Liao, X.; et al. Monitoring Astrocytic Ca2+ Activity in Freely Behaving Mice. Front. Cell. Neurosci. 2020, 14, 410. [Google Scholar] [CrossRef]
- Georgiou, L.; Echeverría, A.; Georgiou, A.; Kuhn, B. Ca+ Activity Maps of Astrocytes Tagged by Axoastrocytic AAV Transfer. Sci. Adv. 2022, 8, 5371. [Google Scholar] [CrossRef]
- Li, M.; Liu, Z.; Wu, Y.; Zheng, N.; Liu, X.; Cai, A.; Zheng, D.; Zhu, J.; Wu, J.; Xu, L.; et al. In Vivo Imaging of Astrocytes in the Whole Brain with Engineered AAVs and Diffusion-Weighted Magnetic Resonance Imaging. Mol. Psychiatry 2022, 1–8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nagai, J.; Yu, X.; Papouin, T.; Cheong, E.; Freeman, M.R.; Monk, K.R.; Hastings, M.H.; Haydon, P.G.; Rowitch, D.; Shaham, S.; et al. Behaviorally Consequential Astrocytic Regulation of Neural Circuits. Neuron 2021, 109, 576–596. [Google Scholar] [CrossRef]
- Scofield, M.D.; Boger, H.A.; Smith, R.J.; Li, H.; Haydon, P.G.; Kalivas, P.W. Gq-DREADD Selectively Initiates Glial Glutamate Release and Inhibits Cue-Induced Cocaine Seeking. Biol. Psychiatry 2015, 78, 441. [Google Scholar] [CrossRef]
- Agulhon, C.; Boyt, K.M.; Xie, A.X.; Friocourt, F.; Roth, B.L.; Mccarthy, K.D. Modulation of the Autonomic Nervous System and Behaviour by Acute Glial Cell Gq Protein-Coupled Receptor Activation in Vivo. J. Physiol. 2013, 591, 5599. [Google Scholar] [CrossRef]
- Perea, G.; Yang, A.; Boyden, E.S.; Sur, M. Optogenetic Astrocyte Activation Modulates Response Selectivity of Visual Cortex Neurons In Vivo. Nat. Commun. 2014, 5, 3262. [Google Scholar] [CrossRef] [PubMed]
- Mederos, S.; Hernández-Vivanco, A.; Ramírez-Franco, J.; Martín-Fernández, M.; Navarrete, M.; Yang, A.; Boyden, E.S.; Perea, G. Melanopsin for Precise Optogenetic Activation of Astrocyte-Neuron Networks. Glia 2019, 67, 915–934. [Google Scholar] [CrossRef] [PubMed]
- Volgraf, M.; Gorostiza, P.; Numano, R.; Kramer, R.H.; Isacoff, E.Y.; Trauner, D. Allosteric Control of an Ionotropic Glutamate Receptor with an Optical Switch. Nat. Chem. Biol. 2006, 2, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Airan, R.D.; Thompson, K.R.; Fenno, L.E.; Bernstein, H.; Deisseroth, K. Temporally Precise In Vivo Control of Intracellular Signalling. Nature 2009, 458, 1025–1029. [Google Scholar] [CrossRef] [PubMed]
- Nagai, J.; Bellafard, A.; Qu, Z.; Yu, X.; Ollivier, M.; Gangwani, M.R.; Diaz-Castro, B.; Coppola, G.; Schumacher, S.M.; Golshani, P.; et al. Specific and Behaviorally Consequential Astrocyte G q GPCR Signaling Attenuation in Vivo with IβARK. Neuron 2021, 109, 2256–2274.e9. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zima, A.V.; Sheikh, F.; Blatter, L.A.; Chen, J. Endothelin-1–Induced Arrhythmogenic Ca2+ Signaling Is Abolished in Atrial Myocytes of Inositol-1,4,5-Trisphosphate(IP3)–Receptor Type 2–Deficient Mice. Circ. Res. 2005, 96, 1274–1281. [Google Scholar] [CrossRef]
- Yu, X.; Taylor, A.M.W.; Nagai, J.; Golshani, P.; Evans, C.J.; Coppola, G.; Khakh, B.S. Reducing Astrocyte Calcium Signaling In Vivo Alters Striatal Microcircuits and Causes Repetitive Behavior. Neuron 2018, 99, 1170–1187.e9. [Google Scholar] [CrossRef]
- Helmchen, F.; Denk, W. Deep Tissue Two-Photon Microscopy. Nat. Methods 2005, 2, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.T.; Theer, P.; Denk, W. Two-Photon Imaging to a Depth of 1000 Μm in Living Brains by Use of a Ti:Al2O3 Regenerative Amplifier. Opt. Lett. 2003, 28, 1022–1024. [Google Scholar] [CrossRef]
- Centonze, V.E.; White, J.G. Multiphoton Excitation Provides Optical Sections from Deeper within Scattering Specimens than Confocal Imaging. Biophys. J 1998, 75, 2015. [Google Scholar] [CrossRef] [PubMed]
- Tonnesen, J.; Nägerl, U.V. Two-Color STED Imaging of Synapses in Living Brain Slices. Methods Mol. Biol. 2013, 950, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Ritter, J.G.; Veith, R.; Veenendaal, A.; Siebrasse, J.P.; Kubitscheck, U. Light Sheet Microscopy for Single Molecule Tracking in Living Tissue. PLoS ONE 2010, 5, e11639. [Google Scholar] [CrossRef] [PubMed]
- Keller, P.J.; Dodt, H.U. Light Sheet Microscopy of Living or Cleared Specimens. Curr. Opin. Neurobiol. 2012, 22, 138–143. [Google Scholar] [CrossRef] [PubMed]
- di Castro, M.A.; Chuquet, J.; Liaudet, N.; Bhaukaurally, K.; Santello, M.; Bouvier, D.; Tiret, P.; Volterra, A. Local Ca2+ Detection and Modulation of Synaptic Release by Astrocytes. Nat. Neurosci. 2011, 14, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; DelRosso, N.V.; Vaidyanathan, T.V.; Cahill, M.K.; Reitman, M.E.; Pittolo, S.; Mi, X.; Yu, G.; Poskanzer, K.E. Accurate Quantification of Astrocyte and Neurotransmitter Fluorescence Dynamics for Single-Cell and Population-Level Physiology. Nat. Neurosci. 2019, 22, 1936–1944. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Huang, B.S.; Venugopal, S.; Johnston, A.D.; Chai, H.; Zeng, H.; Golshani, P.; Khakh, B.S. Ca2+ Signaling in Astrocytes from Ip3r2−/− Mice in Brain Slices and during Startle Responses in Vivo. Nat. Neurosci. 2015, 18, 708–717. [Google Scholar] [CrossRef]
- Agarwal, A.; Wu, P.H.; Hughes, E.G.; Fukaya, M.; Tischfield, M.A.; Langseth, A.J.; Wirtz, D.; Bergles, D.E. Transient Opening of the Mitochondrial Permeability Transition Pore Induces Microdomain Calcium Transients in Astrocyte Processes. Neuron 2017, 93, 587–605.e7. [Google Scholar] [CrossRef]
- Lines, J.; Martin, E.D.; Kofuji, P.; Aguilar, J.; Araque, A. Astrocytes Modulate Sensory-Evoked Neuronal Network Activity. Nat. Commun. 2020, 11, 3689. [Google Scholar] [CrossRef]
- Dzyubenko, E.; Prazuch, W.; Pillath-Eilers, M.; Polanska, J.; Hermann, D.M. Analysing Intercellular Communication in Astrocytic Networks Using “Astral”. Front. Cell. Neurosci. 2021, 15, 219. [Google Scholar] [CrossRef]
- Mederos, S.; González-Arias, C.; Perea, G. Melanopsin for Time-Controlling Activation of Astrocyte -Neuron Networks. Methods Mol. Biol. 2020, 2173, 53–69. [Google Scholar] [CrossRef]
- Kastanenka, K.V.; Moreno-Bote, R.; de Pittà, M.; Perea, G.; Eraso-Pichot, A.; Masgrau, R.; Poskanzer, K.E.; Galea, E. A Roadmap to Integrate Astrocytes into Systems Neuroscience. Glia 2020, 68, 5–26. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Khakh, B.S. SnapShot: Astrocyte Interactions. Cell 2022, 185, 220–220.e1. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Nold, A.; Tchumatchenko, T. Modulation of Working Memory Duration by Synaptic and Astrocytic Mechanisms. PLoS Comput. Biol. 2022, 18, e1010543. [Google Scholar] [CrossRef]
- Nande, A.; Dubinkina, V.; Ravasio, R.; Zhang, G.H.; Berman, G.J. Bottlenecks, Modularity, and the Neural Control of Behavior. Front. Behav. Neurosci. 2022, 16, 835753. [Google Scholar] [CrossRef]
- Shein-Idelson, M.; Cohen, G.; Ben-Jacob, E.; Hanein, Y. Modularity Induced Gating and Delays in Neuronal Networks. PLoS Comput. Biol. 2016, 12, e1004883. [Google Scholar] [CrossRef]
- Cangelosi, A.; Invitto, S. Human-Robot Interaction and Neuroprosthetics: A Review of New Technologies. IEEE Consum. Electron. Mag. 2017, 6, 24–33. [Google Scholar] [CrossRef]
- Chang, L.; Tsao, D.Y. The Code for Facial Identity in the Primate Brain. Cell 2017, 169, 1013–1028.e14. [Google Scholar] [CrossRef]
- Kiani, R.; Cueva, C.J.; Reppas, J.B.; Newsome, W.T. Dynamics of Neural Population Responses in Prefrontal Cortex Indicate Changes of Mind on Single Trials. Curr. Biol. 2014, 24, 1542–1547. [Google Scholar] [CrossRef]
- Nogueira, R.; Abolafia, J.M.; Drugowitsch, J.; Balaguer-Ballester, E.; Sanchez-Vives, M.V.; Moreno-Bote, R. Lateral Orbitofrontal Cortex Anticipates Choices and Integrates Prior with Current Information. Nat. Commun. 2017, 8, 14823. [Google Scholar] [CrossRef]
- Kofuji, P.; Araque, A. Annual Review of Neuroscience Astrocytes and Behavior. Annu. Rev. Neurosci. 2021, 44, 49–67. [Google Scholar] [CrossRef]
- Nimmerjahn, A.; Hirrlinger, J. Astrocyte Regulation of Neural Circuit Function and Animal Behavior. Glia 2022, 70, 1453–1454. [Google Scholar] [CrossRef] [PubMed]
- Tso, C.F.; Simon, T.; Greenlaw, A.C.; Puri, T.; Mieda, M.; Herzog, E.D. Astrocytes Regulate Daily Rhythms in the Suprachiasmatic Nucleus and Behavior. Curr. Biol. 2017, 27, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- van Horn, M.R.; Benfey, N.J.; Shikany, C.; Severs, L.J.; Deemyad, T. Neuron-Astrocyte Networking: Astrocytes Orchestrate and Respond to Changes in Neuronal Network Activity across Brain States and Behaviors. J. Neurophysiol. 2021, 126, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Gordleeva, S.Y.; Tsybina, Y.A.; Krivonosov, M.I.; Ivanchenko, M.V.; Zaikin, A.A.; Kazantsev, V.B.; Gorban, A.N. Modeling Working Memory in a Spiking Neuron Network Accompanied by Astrocytes. Front. Cell. Neurosci. 2021, 15, 86. [Google Scholar] [CrossRef] [PubMed]
- de Pittà, M.; Brunel, N. Modulation of Synaptic Plasticity by Glutamatergic Gliotransmission: A Modeling Study. Neural Plast. 2016, 2016, 7607924. [Google Scholar] [CrossRef]
- Sajedinia, Z.; Hélie, S. A New Computational Model for Astrocytes and Their Role in Biologically Realistic Neural Networks. Comput. Intell. Neurosci. 2018, 2018, 3689487. [Google Scholar] [CrossRef] [PubMed]
- Mesejo, P.; Ibáñez, O.; Fernández-Blanco, E.; Cedrón, F.; Pazos, A.; Porto-Pazos, A.B. 2nd Reading Artificial Neuron-Glia Networks Learning Approach Based on Cooperative Coevolution. Int. J. Neural. Syst. 2015, 25, 1550012. [Google Scholar] [CrossRef]
- Porto-Pazos, A.B.; Veiguela, N.; Mesejo, P.; Navarrete, M.; Alvarellos, A.; Ibáñez, O.; Pazos, A.; Araque, A. Artificial Astrocytes Improve Neural Network Performance. PLoS ONE 2011, 6, e19109. [Google Scholar] [CrossRef]
- Houades, V.; Koulakoff, A.; Ezan, P.; Seif, I.; Giaume, C. Gap Junction-Mediated Astrocytic Networks in the Mouse Barrel Cortex. J. Neurosci. 2008, 28, 5207–5217. [Google Scholar] [CrossRef]
- Sul, J.Y.; Orosz, G.; Givens, R.S.; Haydon, P.G.; Haydon, P.G. Astrocytic Connectivity in the Hippocampus. Neuron Glia Biol. 2004, 1, 3. [Google Scholar] [CrossRef]
- Perea, G.; Araque, A. Properties of Synaptically Evoked Astrocyte Calcium Signal Reveal Synaptic Information Processing by Astrocytes. J. Neurosci. 2005, 25, 2192–2203. [Google Scholar] [CrossRef] [PubMed]
- Curreli, S.; Bonato, J.; Romanzi, S.; Panzeri, S.; Fellin, T. Complementary Encoding of Spatial Information in Hippocampal Astrocytes. PLoS Biol. 2022, 20, e3001530. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, K.T. Molecular Diversity of Astrocytes. Science 2022, 378, 475–476. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; Sofroniew, M.V. Diversity of Astrocyte Functions and Phenotypes in Neural Circuits. Nat. Neurosci. 2015, 18, 942. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; Deneen, B. The Emerging Nature of Astrocyte Diversity. Annu. Rev. Neurosci. 2019, 42, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.H.; Raff’, M.C.; Abney, E.R.; Cohen, J.; Lindsay, R. Fibrous and Protoplasmic Astrocytes Are Biochemically and Developmentally Distinct. J. Neurosci. 1984, 4, 585–592. [Google Scholar] [CrossRef]
- Poopalasundaram, S.; Knott, C.; Shamotienko, O.G.; Foran, P.G.; Dolly, J.O.; Ghiani, C.A.; Gallo, V.; Wilkin, G.P. Glial Heterogeneity in Expression of the Inwardly Rectifying K Channel, Kir4.1, in Adult Rat CNS. Glia 2000, 30, 362–372. [Google Scholar] [CrossRef]
- Nagy, J.I.; Patel, D.; Ochalski, P.A.Y.; Stelmack, G.L. Connexin30 in Rodent, Cat and Human Brain: Selective Expression in Gray Matter Astrocytes, Co-Localization with Connexin43 at Gap Junctions and Late Developmental Appearance. Neuroscience 1999, 88, 447–468. [Google Scholar] [CrossRef]
- Goursaud, S.; Kozlova, E.N.; Maloteaux, J.M.; Hermans, E. Cultured Astrocytes Derived from Corpus Callosum or Cortical Grey Matter Show Distinct Glutamate Handling Properties. J. Neurochem. 2009, 108, 1442–1452. [Google Scholar] [CrossRef]
- Regan, M.R.; Huang, Y.H.; Yu, S.K.; Dykes-Hoberg, M.I.; Jin, L.; Watkins, A.M.; Bergles, D.E.; Rothstein, J.D. Variations in Promoter Activity Reveal a Differential Expression and Physiology of Glutamate Transporters by Glia in the Developing and Mature CNS. J. Neurosci. 2007, 27, 6607–6619. [Google Scholar] [CrossRef]
- Höft, S.; Griemsmann, S.; Seifert, G.; Steinhäuser, C. Heterogeneity in Expression of Functional Ionotropic Glutamate and GABA Receptors in Astrocytes across Brain Regions: Insights from the Thalamus. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130602. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, O.A.; Fuentealba, L.C.; Alvarez-Buylla, A.; Rowitch, D.H. Astrocyte Development and Heterogeneity. Cold Spring Harb. Perspect. Biol. 2014, 7, a020362. [Google Scholar] [CrossRef] [PubMed]
- Chaboub, L.S.; Deneen, B. Developmental Origins of Astrocyte Heterogeneity: The Final Frontier of CNS Development. Dev. Neurosci. 2012, 34, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Kita, Y.; Kawakami, K.; Takahashi, Y.; Murakami, F. Development of Cerebellar Neurons and Glias Revealed by in Utero Electroporation: Golgi-Like Labeling of Cerebellar Neurons and Glias. PLoS ONE 2013, 8, e70091. [Google Scholar] [CrossRef]
- Oberheim, N.A.; Goldman, S.A.; Nedergaard, M. Heterogeneity of Astrocytic Form and Function. Methods Mol. Biol. 2012, 814, 23–45. [Google Scholar] [CrossRef]
- Schirmer, L.; Schafer, D.P.; Bartels, T.; Rowitch, D.H.; Calabresi, P.A. Diversity and Function of Glial Cell Types in Multiple Sclerosis. Trends Immunol. 2021, 42, 228–247. [Google Scholar] [CrossRef] [PubMed]
- Oberheim, N.A.; Takano, T.; Han, X.; He, W.; Lin, J.H.C.; Wang, F.; Xu, Q.; Wyatt, J.D.; Pilcher, W.; Ojemann, J.G.; et al. Uniquely Hominid Features of Adult Human Astrocytes. J. Neurosci. 2009, 29, 3276–3287. [Google Scholar] [CrossRef]
- Falcone, C.; Martínez-Cerdeño, V. Astrocyte Evolution and Human Specificity. Neural Regen. Res. 2023, 18, 131. [Google Scholar] [CrossRef]
- Navarrete, M.; Perea, G.; Maglio, L.; Pastor, J.; García De Sola, R.; Araque, A. Astrocyte Calcium Signal and Gliotransmission in Human Brain Tissue. Cereb. Cortex 2013, 23, 1240–1246. [Google Scholar] [CrossRef]
- Zeisel, A.; Muñoz-Manchado, A.B.; Codeluppi, S.; Lönnerberg, P.; La Manno, G.; Juréus, A.; Marques, S.; Munguba, H.; He, L.; Betsholtz, C.; et al. Cell Types in the Mouse Cortex and Hippocampus Revealed by Single-Cell RNA-Seq. Science 2015, 347, 1138–1142. [Google Scholar] [CrossRef]
- Oliveira, J.F.; Sardinha, V.M.; Guerra-Gomes, S.; Araque, A.; Sousa, N. Do Stars Govern Our Actions? Astrocyte Involvement in Rodent Behavior. Trends Neurosci. 2015, 38, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Bergles, D.E. Large-Scale Recording of Astrocyte Activity. Curr. Opin. Neurobiol. 2015, 32, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.A.; Wang, X.; Cook, D.; Carey, E.M.; Nimmerjahn, A. Imaging Spinal Cord Activity in Behaving Animals. Exp. Neurol. 2019, 320, 112974. [Google Scholar] [CrossRef] [PubMed]
- Bekar, L.K.; He, W.; Nedergaard, M. Locus Coeruleus Alpha-Adrenergic-Mediated Activation of Cortical Astrocytes In Vivo. Cereb. Cortex 2008, 18, 2789–2795. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lou, N.; Xu, Q.; Tian, G.F.; Peng, W.G.; Han, X.; Kang, J.; Takano, T.; Nedergaard, M. Astrocytic Ca2+ Signaling Evoked by Sensory Stimulation in Vivo. Nat. Neurosci. 2006, 9, 816–823. [Google Scholar] [CrossRef]
- Thrane, A.S.; Thrane, V.R.; Zeppenfeld, D.; Lou, N.; Xu, Q.; Nagelhus, E.A.; Nedergaard, M. General Anesthesia Selectively Disrupts Astrocyte Calcium Signaling in the Awake Mouse Cortex. Proc. Natl. Acad. Sci. USA 2012, 109, 18974–18979. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Mukamel, E.A.; Schnitzer, M.J. Motor Behavior Activates Bergmann Glial Networks. Neuron 2009, 62, 400–412. [Google Scholar] [CrossRef]
- Sonoda, K.; Matsui, T.; Bito, H.; Ohki, K. Astrocytes in the Mouse Visual Cortex Reliably Respond to Visual Stimulation. Biochem. Biophys. Res. Commun. 2018, 505, 1216–1222. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic Reactive Astrocytes Are Induced by Activated Microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Rothhammer, V.; Borucki, D.M.; Tjon, E.C.; Takenaka, M.C.; Chao, C.C.; Ardura-Fabregat, A.; De Lima, K.A.; Gutiérrez-Vázquez, C.; Hewson, P.; Staszewski, O.; et al. Microglial Control of Astrocytes in Response to Microbial Metabolites. Nature 2018, 557, 724–728. [Google Scholar] [CrossRef]
- Vainchtein, I.D.; Chin, G.; Cho, F.S.; Kelley, K.W.; Miller, J.G.; Chien, E.C.; Liddelow, S.A.; Nguyen, P.T.; Nakao-Inoue, H.; Dorman, L.C.; et al. Astrocyte-Derived Interleukin-33 Promotes Microglial Synapse Engulfment and Neural Circuit Development. Science 2018, 359, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Tognatta, R.; Karl, M.T.; Fyffe-Maricich, S.L.; Popratiloff, A.; Garrison, E.D.; Schenck, J.K.; Abu-Rub, M.; Miller, R.H. Astrocytes Are Required for Oligodendrocyte Survival and Maintenance of Myelin Compaction and Integrity. Front. Cell. Neurosci. 2020, 14, 74. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Wang, X.; Yang, Y.; Fancy, S.P.J.; Yi, C.; Correspondence, J.N. Astrocyte Endfoot Formation Controls the Termination of Oligodendrocyte Precursor Cell Perivascular Migration during Development Graphical Abstract Highlights d Astrocyte Endfoot Placement Terminates Developmental OPC Perivascular Migration d Astrocyte-Derived Semaphorins 3a and 6a Act to Repel OPCs from Blood Vessels d OPC Detachment from Endothelium Permits Their Subsequent Differentiation. Neuron 2023, 111, 190–201.e8. [Google Scholar] [CrossRef] [PubMed]
- Sanmarco, L.M.; Polonio, C.M.; Wheeler, M.A.; Quintana, F.J. Functional Immune Cell-Astrocyte Interactions. J. Exp. Med. 2021, 218, e20202715. [Google Scholar] [CrossRef]
- Han, R.T.; Kim, R.D.; Molofsky, A.V.; Liddelow, S.A. Astrocyte-Immune Cell Interactions in Physiology and Pathology. Immunity 2021, 54, 211–224. [Google Scholar] [CrossRef]
- Wheeler, M.A.; Clark, I.C.; Lee, H.G.; Li, Z.; Linnerbauer, M.; Rone, J.M.; Blain, M.; Akl, C.F.; Piester, G.; Giovannoni, F.; et al. Droplet-Based Forward Genetic Screening of Astrocyte–Microglia Cross-Talk. Science 2023, 379, 1023. [Google Scholar] [CrossRef]
- Wheeler, M.A.; Jaronen, M.; Covacu, R.; Zandee, S.E.J.; Scalisi, G.; Rothhammer, V.; Tjon, E.C.; Chao, C.C.; Kenison, J.E.; Blain, M.; et al. Environmental Control of Astrocyte Pathogenic Activities in CNS Inflammation. Cell 2019, 176, 581–596.e18. [Google Scholar] [CrossRef]
- Clark, I.C.; Gutiérrez-Vázquez, C.; Wheeler, M.A.; Li, Z.; Rothhammer, V.; Linnerbauer, M.; Sanmarco, L.M.; Guo, L.; Blain, M.; Zandee, S.E.J.; et al. Barcoded Viral Tracing of Single-Cell Interactions in Central Nervous System Inflammation. Science 2021, 372, eabf1230. [Google Scholar] [CrossRef]
- Wang, L.L.; Zhang, C.L. Engineering New Neurons: In Vivo Reprogramming in Mammalian Brain and Spinal Cord. Cell Tissue Res. 2017, 371, 201–212. [Google Scholar] [CrossRef]
- Gascón, S.; Masserdotti, G.; Russo, G.L.; Götz, M. Direct Neuronal Reprogramming: Achievements, Hurdles, and New Roads to Success. Cell Stem Cell 2017, 21, 18–34. [Google Scholar] [CrossRef]
- Tai, W.; Xu, X.M.; Zhang, C.L. Regeneration through in Vivo Cell Fate Reprogramming for Neural Repair. Front. Cell. Neurosci. 2020, 14, 107. [Google Scholar] [CrossRef]
- Rivetti Di Val Cervo, P.; Romanov, R.A.; Spigolon, G.; Masini, D.; Martín-Montañez, E.; Toledo, E.M.; La Manno, G.; Feyder, M.; Pifl, C.; Ng, Y.H.; et al. Induction of Functional Dopamine Neurons from Human Astrocytes in Vitro and Mouse Astrocytes in a Parkinson’s Disease Model. Nat. Biotechnol. 2017, 35, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Tao, X.; Sui, M.; Cui, M.; Liu, D.; Wang, B.; Wang, T.; Zheng, Y.; Luo, J.; Mu, Y.; et al. Reprogramming Astrocytes to Motor Neurons by Activation of Endogenous Ngn2 and Isl1. Stem Cell Rep. 2021, 16, 1777–1791. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Xie, H.; Du, X.; Wang, L.; Jin, X.; Zhang, Q.; Han, Y.; Sun, S.; Wang, L.; Li, X.; et al. In Vivo Chemical Reprogramming of Astrocytes into Neurons. Cell Discov. 2021, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Chen, F.; Song, N.; Xie, J. In Vivo Direct Conversion of Astrocytes to Neurons Maybe a Potential Alternative Strategy for Neurodegenerative Diseases. Front. Aging Neurosci. 2021, 13, 474. [Google Scholar] [CrossRef]
Figure 1.
Percentage of studies focusing exclusively on Neurons (green), Astrocytes (red), and on Neurons and Astrocytes (blue) published in the years 2000 and 2022 and across time within the year range from 2000 to 2022. The total number accounts for the sum of Neuron, Astrocytes, and Neuron and Astrocytes studies. Data were extracted from the Scopus database.
Figure 1.
Percentage of studies focusing exclusively on Neurons (green), Astrocytes (red), and on Neurons and Astrocytes (blue) published in the years 2000 and 2022 and across time within the year range from 2000 to 2022. The total number accounts for the sum of Neuron, Astrocytes, and Neuron and Astrocytes studies. Data were extracted from the Scopus database.

Figure 2.
Map of bibliographical data with co-occurrence analysis of arbitrarily selected keywords (items) to show the relatedness of the keywords as a function of the number of documents in which they occur together, which is quantified by a weight value, and visually reflected in the thickness of the linking line, the closeness within items, and the label size.
Figure 2.
Map of bibliographical data with co-occurrence analysis of arbitrarily selected keywords (items) to show the relatedness of the keywords as a function of the number of documents in which they occur together, which is quantified by a weight value, and visually reflected in the thickness of the linking line, the closeness within items, and the label size.

Figure 3.
Schematic representation to illustrate the traditional perspective in which astrocytes respond homogeneously (left; neurons (blue) and astrocytes (gray)) and the view in which astrocytes respond in ensembles (right; neurons (blue), low-activity astrocytes (green), medium-activity astrocytes (yellow), high-activity astrocytes (red)).
Figure 3.
Schematic representation to illustrate the traditional perspective in which astrocytes respond homogeneously (left; neurons (blue) and astrocytes (gray)) and the view in which astrocytes respond in ensembles (right; neurons (blue), low-activity astrocytes (green), medium-activity astrocytes (yellow), high-activity astrocytes (red)).

Table 1.
Details of the bibliographical search seen in Figure 1.
Table 1.
Details of the bibliographical search seen in Figure 1.
| Bibliographic Search | |
|---|---|
| Source | Scopus |
| Data accessed | 26 October 2022 |
| Query | |
| Description | (neuron) OR (astrocyte) AND (mice, OR mouse, OR human) AND (dendrite) AND (process) AND (network, OR engram) |
| Documents found | 16.199 |
| Query Filters | |
| Years Range | 1980–2023 |
| Language | English |
| Map Analysis | |
| Software | 1.6.18 VOSviewer |
| Type | Co-occurrence |
| Counting Method | Full counting |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Delgado, L.; Navarrete, M. Shining the Light on Astrocytic Ensembles. Cells 2023, 12, 1253. https://doi.org/10.3390/cells12091253
AMA Style
Delgado L, Navarrete M. Shining the Light on Astrocytic Ensembles. Cells. 2023; 12(9):1253. https://doi.org/10.3390/cells12091253
Chicago/Turabian StyleDelgado, Laura, and Marta Navarrete. 2023. "Shining the Light on Astrocytic Ensembles" Cells 12, no. 9: 1253. https://doi.org/10.3390/cells12091253
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.