


miR-205 Regulates the Fusion of Porcine Myoblast by Targeting the Myomaker Gene
 , , , and
, , , and 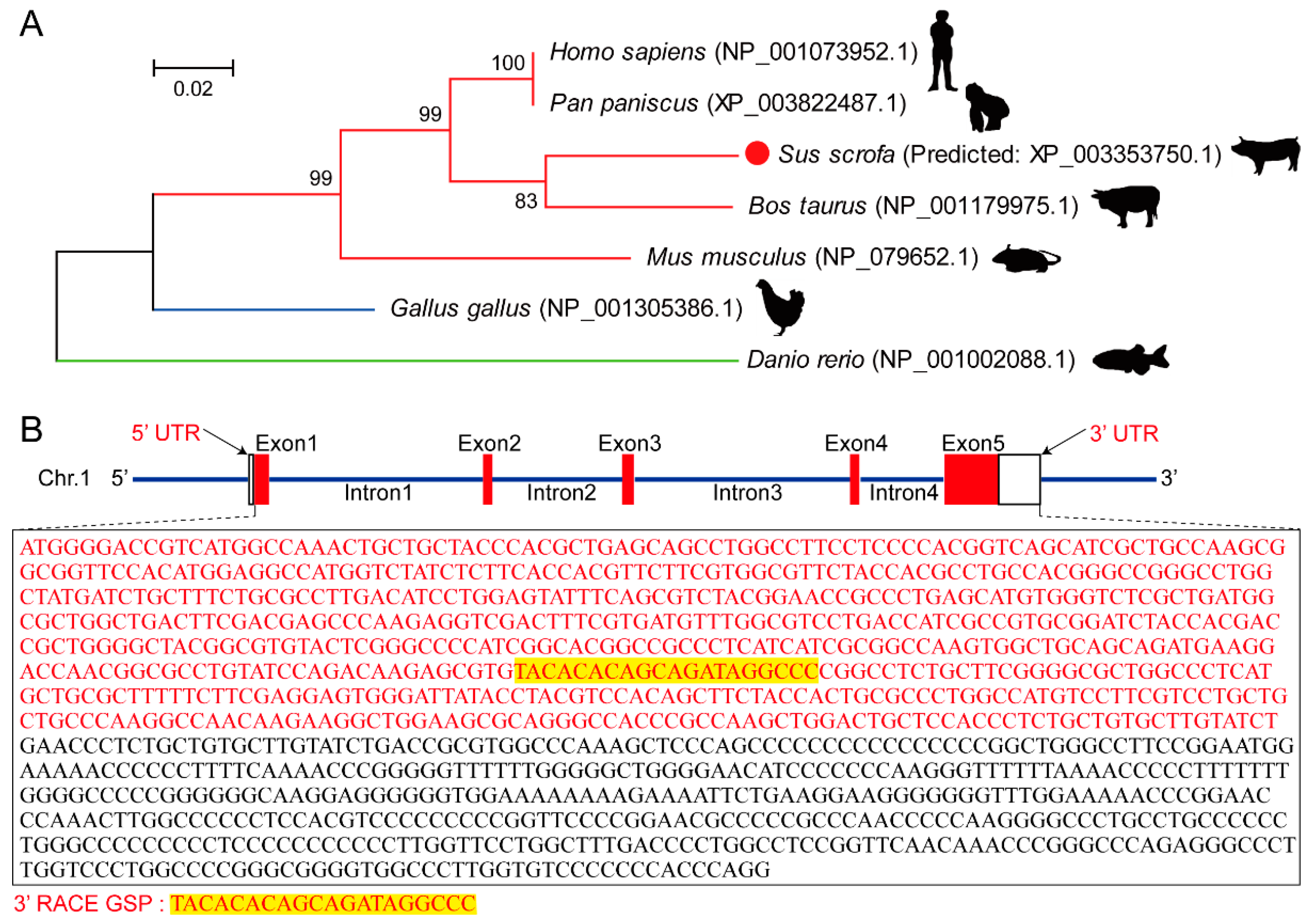{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Tissues Collection
2.2. Total RNA Isolation, Reverse Transcription, and qRT-PCR
2.3. The 3′ Rapid Amplification of cDNA Ends (3′ RACE) of Myomaker
2.4. Western Blotting Analysis
2.5. Primary Skeletal Muscle Satellite Cells (MSCs) Isolation, Purification, and Culture
2.6. Immunocytochemistry and Immunohistochemistry
2.7. Overexpressed Plasmid Construction and Transfection
2.8. Dual-Luciferase Reporter Assay
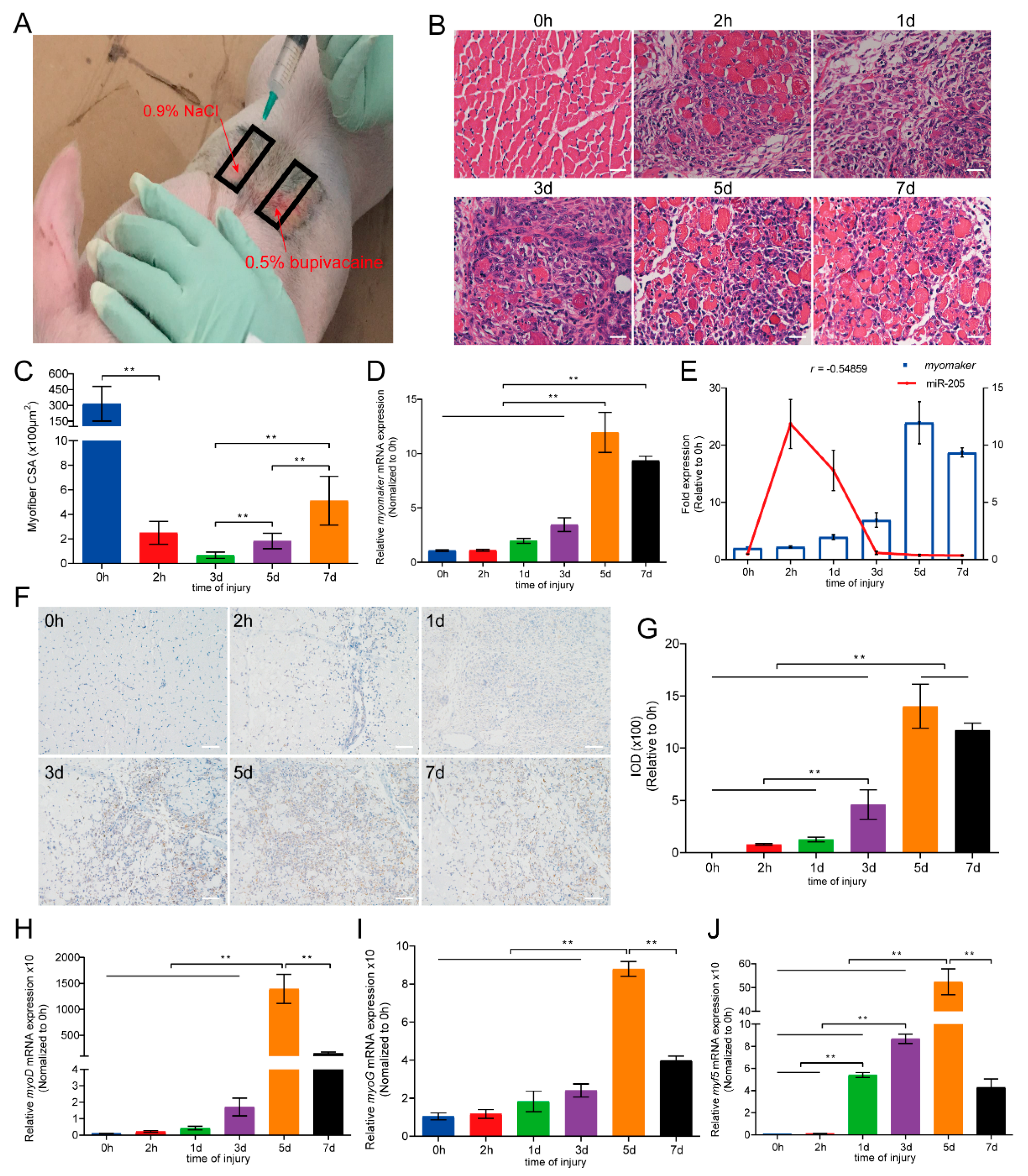
2.9. In Vivo Muscle Injury
2.10. Statistical Analysis
3. Results
3.1. Amino Acid Evolution Tree and Gene Structure of Myomaker
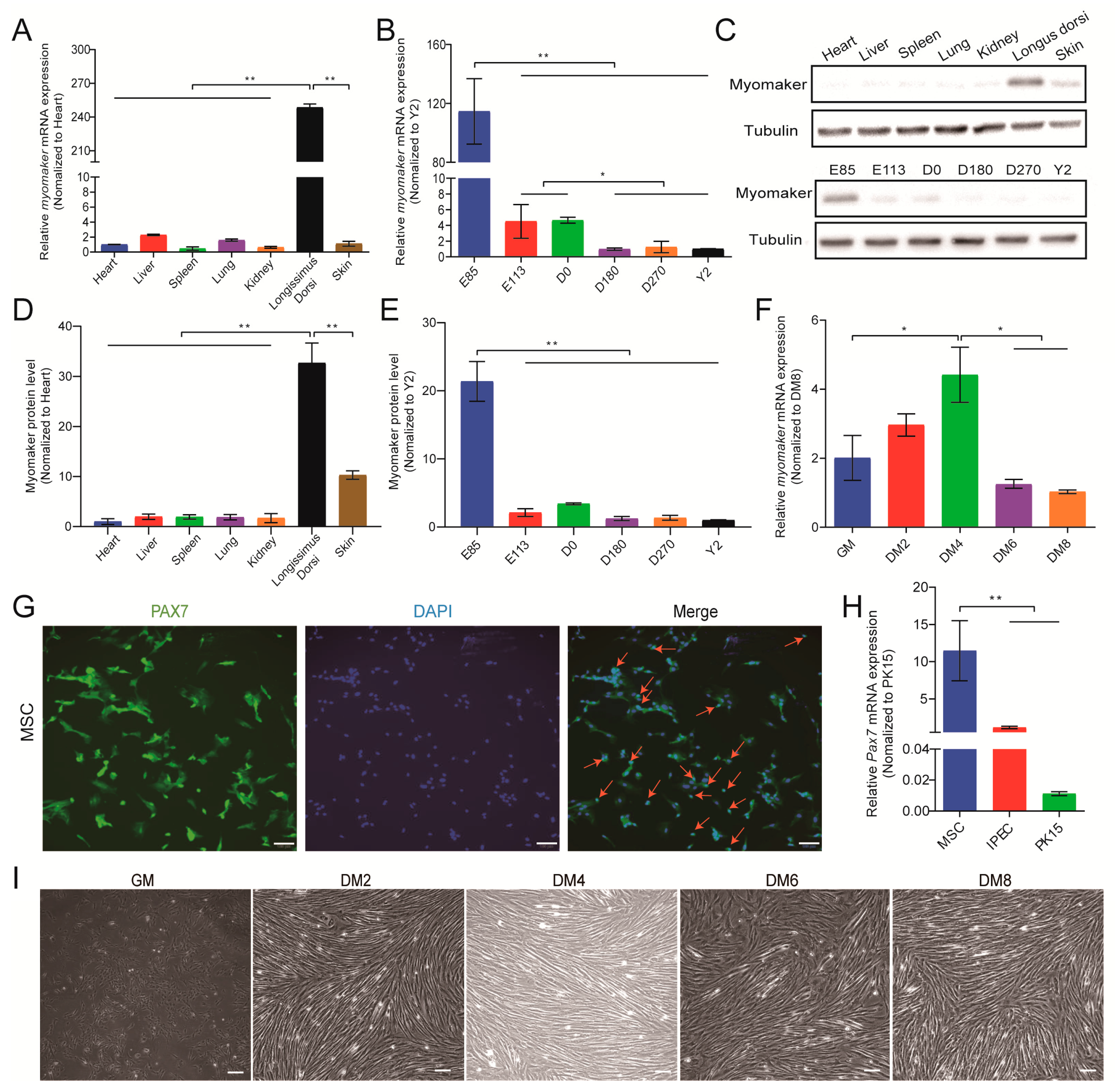
3.2. Myomaker Expression Pattern during Pig Skeletal Muscle Development
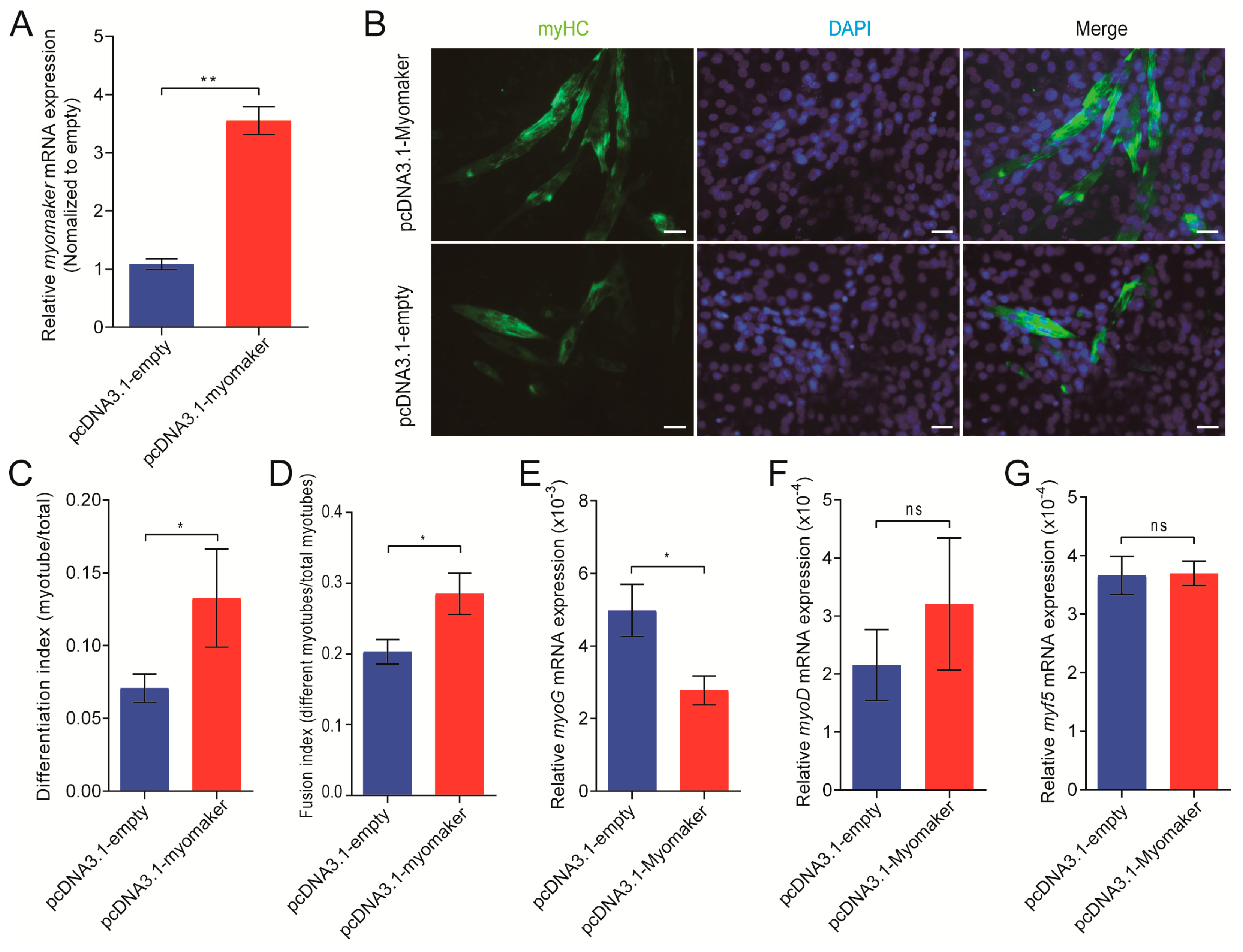
3.3. Myomaker Plays an Important Role in Porcine Primary Myoblast Fusion
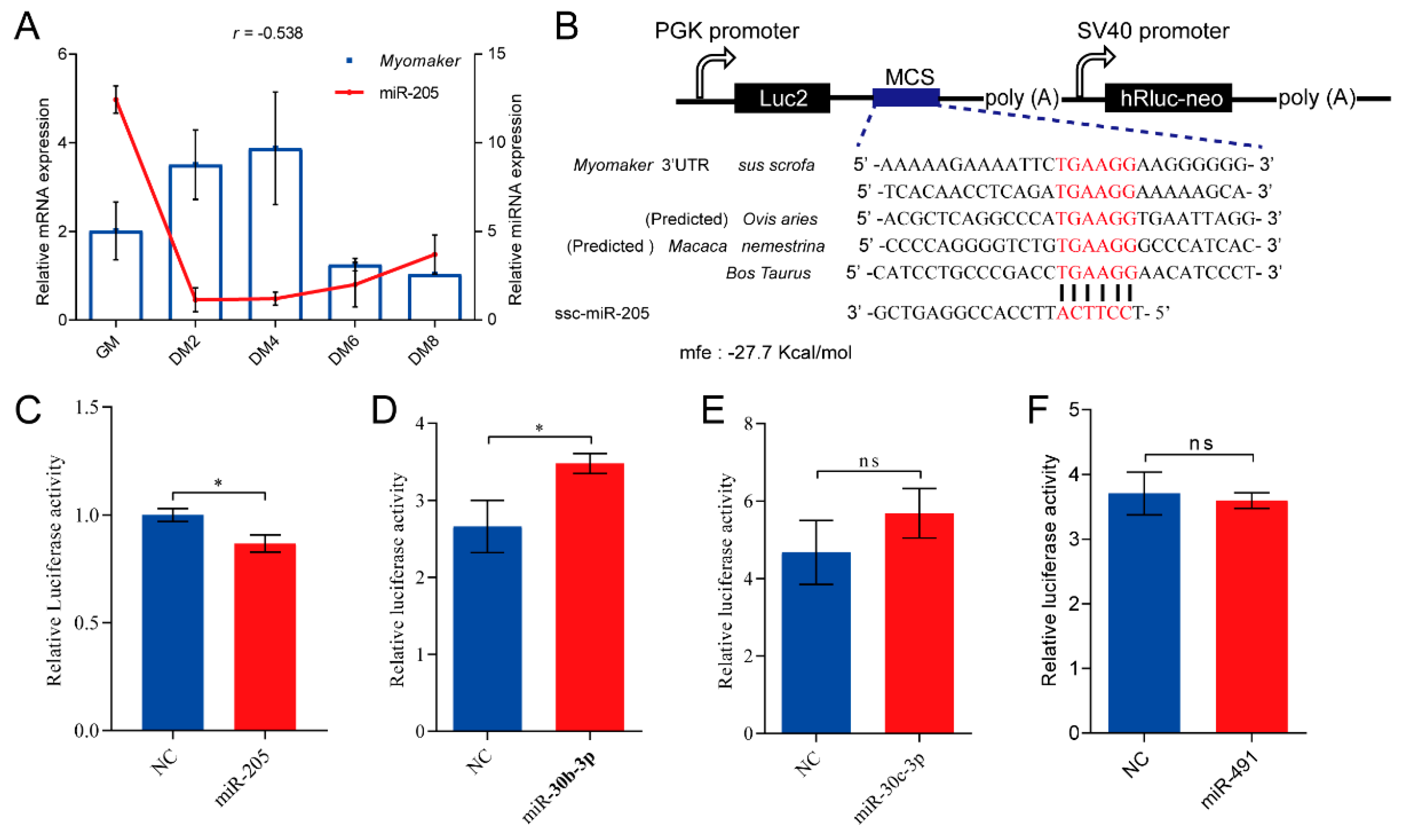
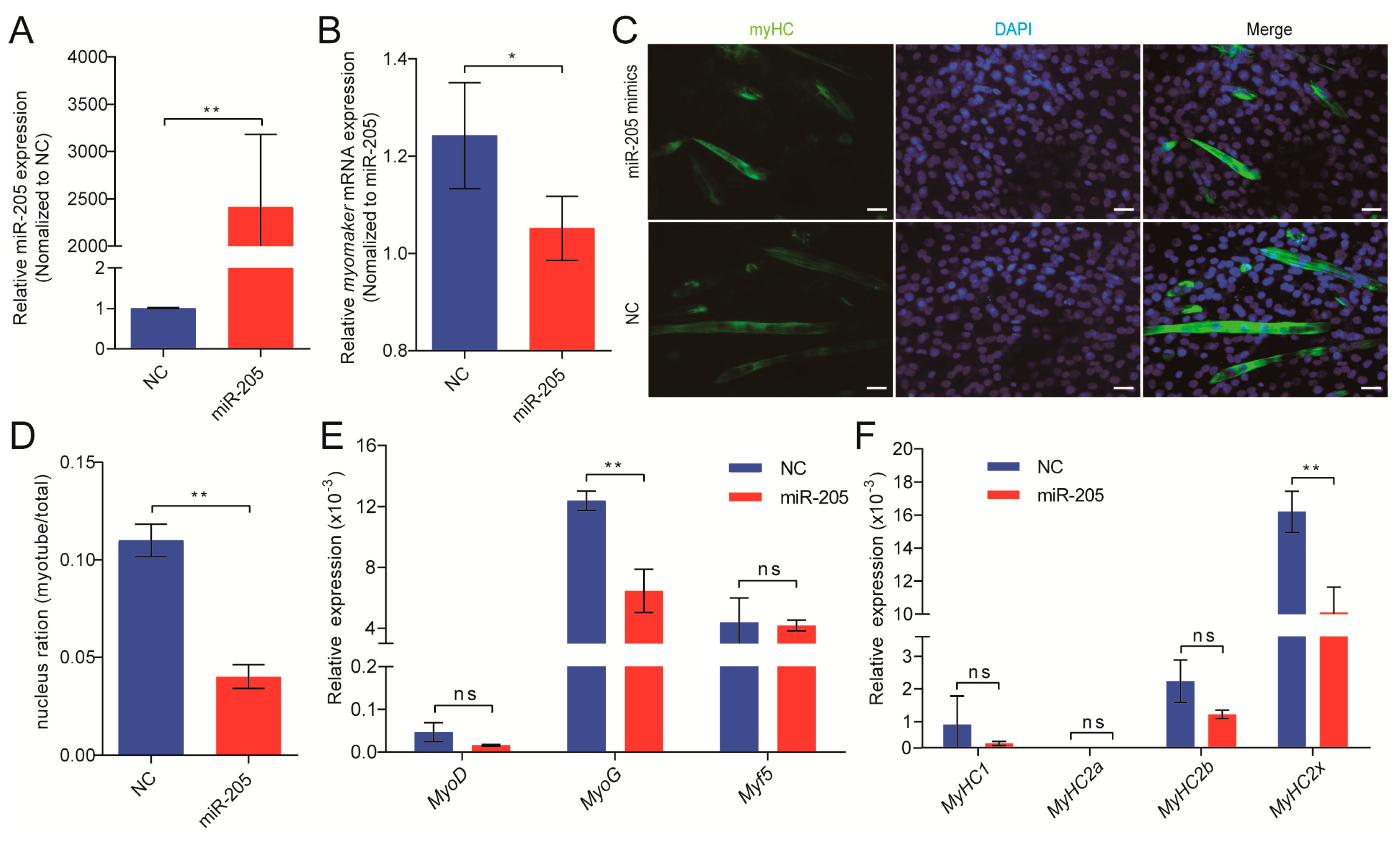
3.4. miR-205 Regulates Myomaker Expression in Pigs
3.5. miR-205 Inhibits Primary Myoblasts Fusion through Targeting Myomaker
3.6. Expression Profiles of Myomaker during Porcine Muscle Regeneration
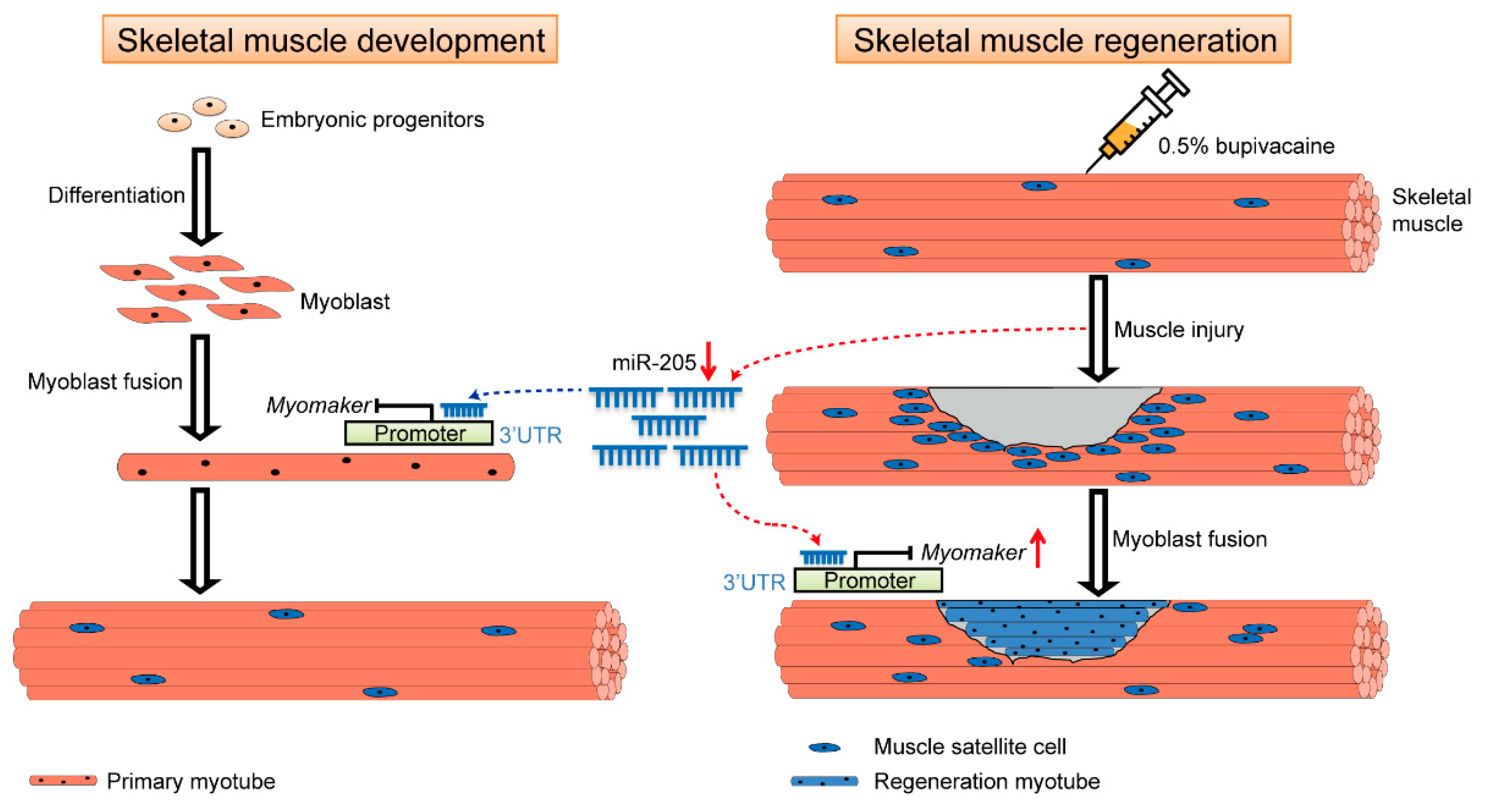
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buckingham, M. Myogenic progenitor cells and skeletal myogenesis in vertebrates. Curr. Opin. Genet. Dev. 2006, 16, 525–532. [Google Scholar] [CrossRef]
- Buckingham, M.; Relaix, F. The role of Pax genes in the development of tissues and organs: Pax3 and Pax7 regulate muscle progenitor cell functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 645–673. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef] [Green Version]
- Bharathy, N.; Ling, B.M.; Taneja, R. Epigenetic regulation of skeletal muscle development and differentiation. Subcell. Biochem. 2013, 61, 139–150. [Google Scholar]
- Cheung, T.H.; Rando, T.A. Molecular regulation of stem cell quiescence. Nat. Rev. Mol. Cell Biol. 2013, 14, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Millay, D.P.; O’Rourke, J.R.; Sutherland, L.B.; Bezprozvannaya, S.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Myomaker is a membrane activator of myoblast fusion and muscle formation. Nature 2013, 499, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Millay, D.P.; Sutherland, L.B.; Bassel-Duby, R.; Olson, E.N. Myomaker is essential for muscle regeneration. Genes Dev. 2014, 28, 1641–1646. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Wang, F.; Zhang, P.; Li, W.; Wang, J.; Li, J.; Liu, H.; Chen, X. miR-491 inhibits skeletal muscle differentiation through targeting myomaker. Arch. Biochem. Biophys. 2017, 625, 30–38. [Google Scholar] [CrossRef]
- Zhang, W.; Roy, S. Myomaker is required for the fusion of fast-twitch myocytes in the zebrafish embryo. Dev. Biol. 2017, 423, 24–33. [Google Scholar] [CrossRef]
- Luo, W.; Li, E.; Nie, Q.; Zhang, X. Myomaker, regulated by MYOD, MYOG and miR-140-3p, promotes chicken myoblast fusion. Int. J. Mol. Sci. 2015, 16, 26186–26201. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.H.; Liu, N.; Van Rooij, E.; Olson, E.N. MicroRNA control of muscle development and disease. Curr. Opin. Cell Biol. 2009, 21, 461–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goljanek-Whysall, K.; Sweetman, D.; Münsterberg, A.E. microRNAs in skeletal muscle differentiation and disease. Clin. Sci. 2012, 123, 611–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.Y.; Fu, L.L.; Cheng, H.J.; Zhao, S.H. Advances on microRNA in regulating mammalian skeletal muscle development. Yi Chuan 2017, 39, 1046–1053. [Google Scholar]
- He, K.; Ren, T.; Zhu, S.; Liang, S.; Zhao, A. Transiently expressed pattern during myogenesis and candidate miRNAs of Tmem8C in goose. J. Genet. 2017, 96, 39–46. [Google Scholar] [CrossRef]
- He, B.; Zheng, R.; Xiong, Y. Culture, identification and biological characteristics of skeletal muscle satellite cells of the neonatal pig. Acta Vet. Et Zootech. Sinica 2006, 37, 555. [Google Scholar]
- Johnson, S.E.; Allen, R.E. Proliferating cell nuclear antigen (PCNA) is expressed in activated rat skeletal muscle satellite cells. J. Cell Physiol. 1993, 15, 39–43. [Google Scholar] [CrossRef]
- Yablonka-Reuveni, Z.; Quinn, L.S.; Nameroff, M. Isolation and clonal analysis of satellite cells from chicken pectoralis muscle. Dev. Biol. 1987, 119, 252–259. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, C.; Kobayashi, S.; Nonaka, I. Comparison of behavior in muscle fiber regeneration after bupivacaine hydrochloride-and acid anhydride-induced myonecrosis. Acta Neuropathol. 1992, 83, 584–589. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. elife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Rehmsmeier, M.; Steffen, P.; Hochsmann, M.; Giegerich, R. Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef] [Green Version]
- Desvignes, T.; Batzel, P.; Berezikov, E.; Eilbeck, K.; Eppig, J.T.; McAndrews, M.S.; Singer, A.; Postlethwait, J.H. miRNA nomenclature: A view incorporating genetic origins, biosynthetic pathways, and sequence variants. Trends Genet. 2015, 31, 613–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piletič, K.; Kunej, T. Minimal standards for reporting microRNA: Target interactions. OMICS 2017, 21, 197–206. [Google Scholar] [CrossRef]
- Jurie, C.; Picard, B.; Geay, Y. Changes in the metabolic and contractile characteristics of muscle in male cattle between 10 and 16 months of age. Histochem. J. 1999, 31, 117–122. [Google Scholar] [CrossRef]
- Brooke, M.H.; Kaiser, K.K. Three “myosin adenosine triphosphatase” systems: The nature of their pH lability and sulfhydryl dependence. J. Histochem. Cytochem. 1970, 18, 670–672. [Google Scholar] [CrossRef]
- Rahelic, S.; Puac, S. Fibre types in longissimus dorsi from wild and highly selected pig breeds. Meat Sci. 1981, 5, 439–450. [Google Scholar] [CrossRef]
- Landemaine, A.; Rescan, P.Y.; Gabillard, J.C. Myomaker mediates fusion of fast myocytes in zebrafish embryos. Biochem. Biophys. Res. Commun. 2014, 451, 480–484. [Google Scholar] [CrossRef]
- Chen, Y.P.; Liu, T.; Zou, D.H.; Zhang, L.; Mao, Y.Q.; Huo, Z.J. Histomorphological assessment of a rat model of skeletal muscle injury induced by local injection of bupivacaine. Chin. J. Tissue Eng. Res. 2016, 20, 2615–2621. [Google Scholar]
- Nepomnyashchikh, L.M.; Bakarev, M.A.; Tsimmerman, V.G. Reparative reactions of the skeletal muscles in early aging OXYS rats with toxic metabolic injuries caused by bupivacaine. Bull. Exp. Biol. Med. 2007, 144, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt, J.D.; Woods, R.I. Hypertrophy of rat extensor digitorum longus muscle injected with bupivacaine. A sequential histochemical, immunohistochemical, histological and morphometric study. J. Anat. 1992, 181, 11. [Google Scholar] [PubMed]
- Burn, J.H.; Rand, M.J. The cause of the supersensitivity of smooth muscle to noradrenaline after sympathetic degeneration. J. Physiol. 1959, 147, 135. [Google Scholar] [CrossRef] [Green Version]
- Oz Gergin, O.; Bayram, A.; Gergin, I.S.; Aksu, R.; Yay, A.; Balcıoglu, E.; Polat, S.; Coşkun, G.; Soyer Sarıca, Z.; Yıldız, K. Comparison of myotoxic effects of levobupivacaine, bupivacaine and ropivacaine: Apoptotic activity and acute effect on pro-inflammatory cytokines. Biotech. Histochem. 2019, 94, 252–260. [Google Scholar] [CrossRef]
- Benoit, P.W.; Belt, W.D. Destruction and regeneration of skeletal muscle after treatment with a local anaesthetic, bupivacaine (Marcaine). J. Anat. 1970, 107, 547. [Google Scholar] [PubMed]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef]
- Van Rooij, E.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Hill, J.; Olson, E.N. Control of stress-depe ndent cardiac growth and gene expression by a microRNA. Science 2007, 316, 575–579. [Google Scholar] [CrossRef] [Green Version]
- Greene, S.B.; Herschkowitz, J.I.; Rosen, J.M. The ups and downs of miR-205: Identifying the roles of miR-205 in mammary gland development and breast cancer. RNA Biol. 2010, 7, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, K.; Isomoto, H.; Yamaguchi, N.; Inoue, N.; Machida, H.; Nakayama, T.; Hayashi, T.; Kunizaki, M.; Hidaka Shigekazu, T.; Masahiro, N.; et al. MiRNA-205 modulates cellular invasion and migration via regulating zinc finger E-box binding homeobox 2 expression in esophageal squamous cell carcinoma cells. J. Transl. Med. 2011, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Zhu, Y.; Zhou, X.; Zhang, J.; Sun, J.; Li, Z.; Jin, L.; Long, K.; Lu, L.; Ge, L. miR-205 Regulates the Fusion of Porcine Myoblast by Targeting the Myomaker Gene. Cells 2023, 12, 1107. https://doi.org/10.3390/cells12081107
Ma J, Zhu Y, Zhou X, Zhang J, Sun J, Li Z, Jin L, Long K, Lu L, Ge L. miR-205 Regulates the Fusion of Porcine Myoblast by Targeting the Myomaker Gene. Cells. 2023; 12(8):1107. https://doi.org/10.3390/cells12081107
Chicago/Turabian StyleMa, Jideng, Yan Zhu, Xiankun Zhou, Jinwei Zhang, Jing Sun, Zhengjie Li, Long Jin, Keren Long, Lu Lu, and Liangpeng Ge. 2023. "miR-205 Regulates the Fusion of Porcine Myoblast by Targeting the Myomaker Gene" Cells 12, no. 8: 1107. https://doi.org/10.3390/cells12081107