
Sclerotinia sclerotiorum (Lib.) de Bary: Insights into the Pathogenomic Features of a Global Pathogen

 , , , and
, , , and
Abstract
:1. Introduction
2. Unique Signs and Symptoms That Constitute Clinical Features of S. sclerotiorum
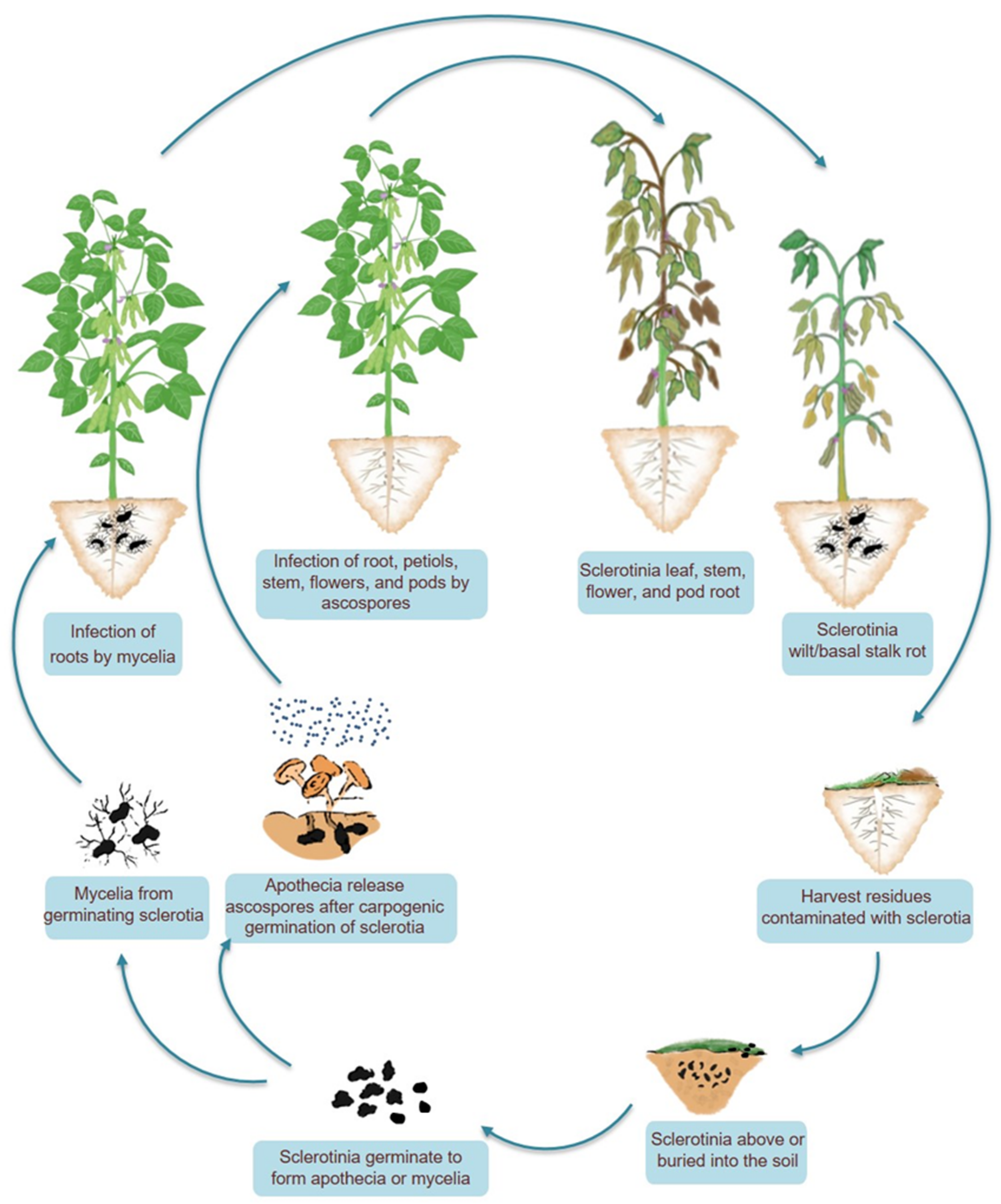
3. A Naive Lifecycle of a Deadly Pathogen
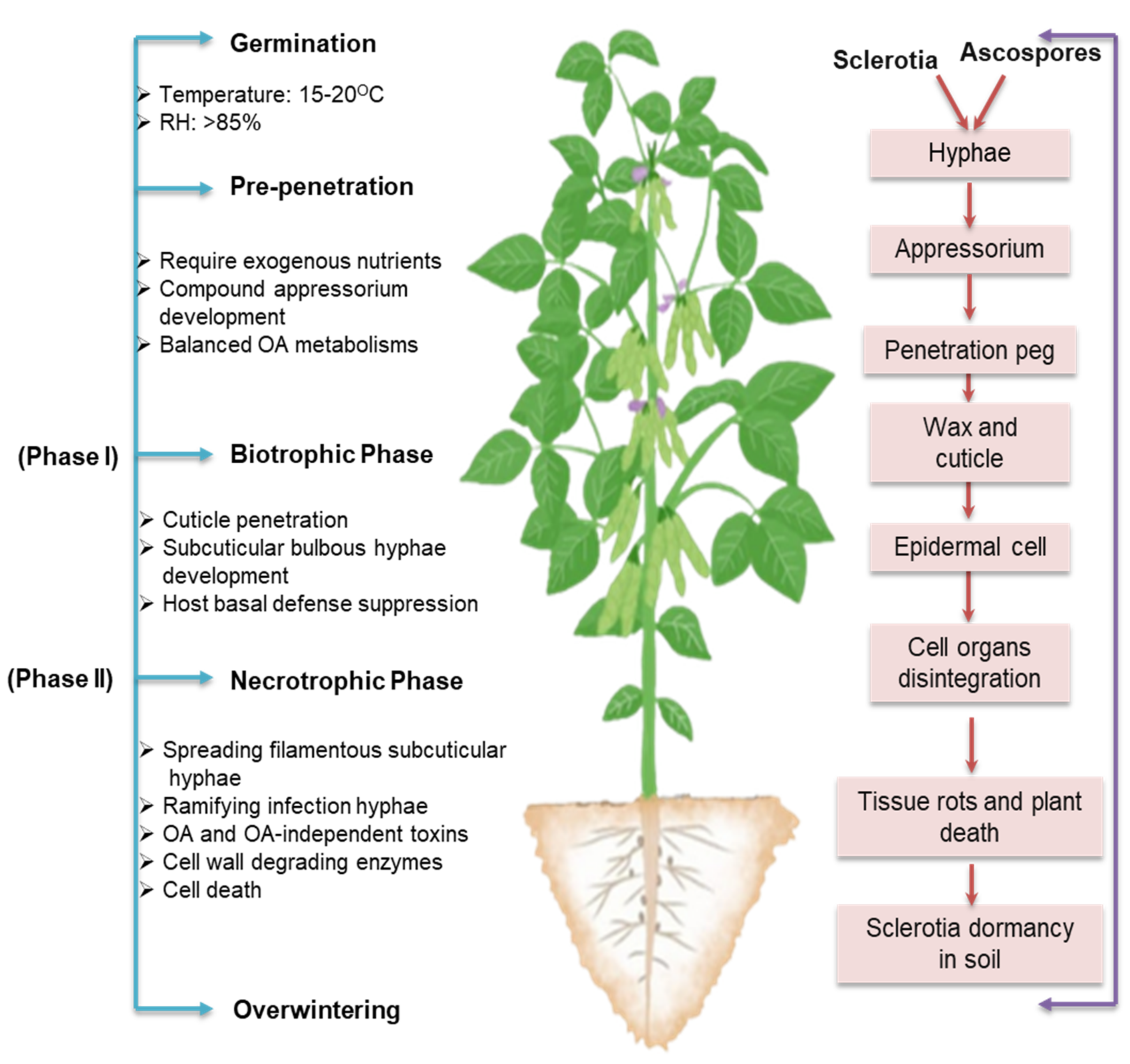
4. Two-Phase Infection Model: The Transition from Biotrophy to Necrotrophy
5. Privileges of Massive Arsenal to Subsist and Succeed against Host Defense
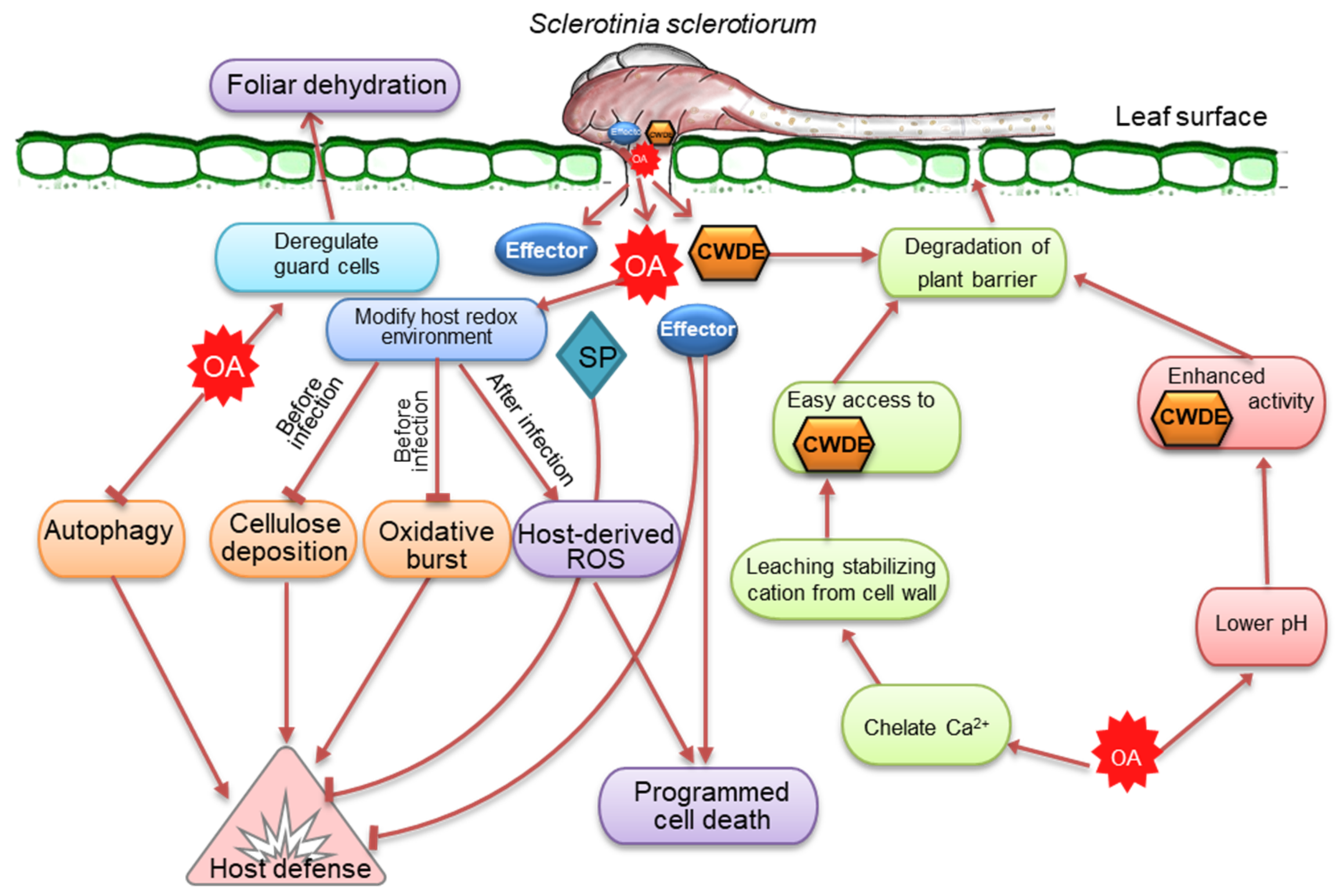
5.1. Oxalic Acid, a Versatile Toxin and Broad-Spectrum Pathogenicity Factor
5.2. Arsenal of Cell Wall-Degrading Enzymes Reflects Broad Host Preference
5.3. Secretory Effectors, Critical Components in the Necrotrophic Infection Strategy
6. An Array of Genes Regulating Growth, Development, and Virulence
7. cAMP as a Key Modulator
8. MAPK as Central Signaling Cascade
9. Sclerotinia Genome as a Treasure Trove
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lehner, M.S.; De Paula Junior, T.J.; Del Ponte, E.M.; Mizubuti, E.S.G.; Pethybridge, S.J. Independently founded populations of Sclerotinia sclerotiorum from a tropical and a temperate region have similar genetic structure. PLoS ONE 2017, 12, e0173915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahan, R.; Siddique, S.S.; Jannat, R.; Hossain, M.M. Cosmos white rot: First characterization, physiology, host range, disease resistance, and chemical control. J. Basic Microbiol. 2022, 62, 911–929. [Google Scholar] [CrossRef]
- Saharan, G.S.; Mehta, N. Sclerotinia Diseases of Crop Plants: Biology, Ecology and Disease Management; Springer: Berlin, Germany, 2008; ISBN 978-1-4020-8408-9. [Google Scholar]
- Prova, A.; Akanda, A.M.; Islam, S.; Hossain, M.M. First report of Sclerotinia sclerotiorum causing pod rot disease on okra in Bangladesh. Can. J. Plant Pathol. 2017, 39, 72–76. [Google Scholar] [CrossRef]
- Islam, M.R.; Akanda, A.M.; Hossain, M.M.; Hossain, M.M. First characterization of a newly emerging phytopathogen, Sclerotinia sclerotiorum causing white mold in pea. J. Basic Microbiol. 2021, 61, 923–939. [Google Scholar] [CrossRef] [PubMed]
- Prova, A.; Akanda, M.A.M.; Islam, S.; Sultana, F.; Islam, M.T.; Hossain, M.M. First report of stem and pod blight of hyacinth bean caused by Sclerotinia sclerotiorum in Bangladesh. J. Plant Pathol. 2014, 96, 603–611. [Google Scholar] [CrossRef]
- Choi, I.Y.; Kim, J.; Lee, W.H.; Cho, S.E.; Shin, H.D. First report of Sclerotinia stem rot caused by Sclerotinia sclerotiorum on Chinese chives in Korea. Plant Dis. 2017, 101, 1953. [Google Scholar] [CrossRef]
- Islam, M.R.; Prova, A.; Akanda, A.M.; Hossain, M.M. First report of white mould caused by Sclerotinia sclerotiorum on pea in Bangladesh. J. Plant. Pathol. 2020, 102, 941. [Google Scholar] [CrossRef] [Green Version]
- Boland, G.J.; Hall, R. Index of plant hosts of Sclerotinia sclerotiorum. Can. J. Plant Pathol. 1994, 16, 93–108. [Google Scholar] [CrossRef]
- Tian, B.; Xie, J.; Fu, Y.; Cheng, J.; Li, B.O.; Chen, T.; Zhao, Y.; Gao, Z.; Yang, P.; Barbetti, M.J.; et al. A cosmopolitan fungal pathogen of dicots adopts an endophytic lifestyle on cereal crops and protects them from major fungal diseases. ISME J. 2020, 14, 3120–3135. [Google Scholar] [CrossRef]
- Derbyshire, M.C.; Denton-Giles, M. The control of sclerotinia stem rot on oilseed rape (Brassica napus): Current practices and future opportunities. Plant Pathol. 2016, 65, 859–877. [Google Scholar] [CrossRef] [Green Version]
- Alkooranee, J.T.; Aledan, T.R.; Ali, A.K.; Lu, G.; Zhang, X.; Wu, J.; Fu, C.; Li, M. Detecting the hormonal pathways in oilseed rape behind induced systemic resistance by Trichoderma harzianum TH12 to Sclerotinia sclerotiorum. PLoS ONE 2017, 12, e0168850. [Google Scholar] [CrossRef] [Green Version]
- Albert, D.; Dumonceaux, T.; Carisse, O.; Beaulieu, C.; Filion, M. Combining desirable traits for a good biocontrol strategy against Sclerotinia sclerotiorum. Microorganisms 2022, 10, 1189. [Google Scholar] [CrossRef]
- Rollins, J.A.; Dickman, M.B. pH signaling in Sclerotinia sclerotiorum: Identification of a pacC/RIMI homolog. Appl. Environ. Microbiol. 2001, 67, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Heffer Link, V.; Johnson, K.B. White Mold. Plant Health Instructor. 2007. [Google Scholar] [CrossRef]
- Mathew, F.; Harveson, R.; Block, C.; Gulya, T.; Ryley, M.; Thompson, S.; Markell, S. Sclerotinia sclerotiorum diseases of sunflower (white mold). Plant Health Instructor. 2020. [Google Scholar] [CrossRef]
- Prova, A.; Akanda, A.M.; Islam, S.; Hossain, M.M. Characterization of Sclerotinia sclerotiorum, an emerging fungal pathogen causing blight in Hyacinth Bean (Lablab purpureus). Plant Pathol. J. 2018, 34, 367–380. [Google Scholar] [CrossRef]
- Ekins, M.G.; Aitken, E.A.B.; Goulter, K.C. Carpogenic germination of Sclerotinia minor and potential distribution in Australia. Australas. Plant Pathol. 2002, 31, 259–265. [Google Scholar] [CrossRef]
- Smolinska, U.; Kowalska, B. Biological control of the soil-borne fungal pathogen Sclerotinia sclerotiorum—A review. J. Plant Pathol. 2018, 100, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sutton, D.C.; Deverall, B.J. Studies on infection of bean (Phaseolus vulgaris) and soybean (Glycine max) by ascospores of Sclerotinia sclerotiorum. Plant Pathol. 1983, 32, 251–261. [Google Scholar] [CrossRef]
- Hegedus, D.D.; Rimmer, S.R. Sclerotinia sclerotiorum: When “to be or not to be” a pathogen? FEMS Microbiol Lett. 2005, 251, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Fernando, W.G.D.; Nakkeeran, S.; Zhang, Y. Ecofriendly Methods in Combating Sclerotinia sclerotiorum (Lib.) de Bary; Research Signpost: Trivandrum, India, 2004; Volume 661, pp. 329–347. ISBN 81-7736-217-8. [Google Scholar]
- Thomma, B.P.H.J. Alternaria spp. from general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, J.P.; Fawcett, L.; Anthony, S.G.; Young, C. A model for Sclerotinia sclerotiorum Infection and disease development in lettuce, based on the effects of temperature, relative humidity and ascospore density. PLoS ONE 2014, 9, e94049. [Google Scholar] [CrossRef] [PubMed]
- Rollins, J.A. The Sclerotinia sclerotiorum pac1 gene is required for sclerotial development and virulence. Mol. Plant Microb. Interact. 2003, 16, 785–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.F.; Rollins, J.A. Mechanisms of broad host range necrotrophic pathogenesis in Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1128–1140. [Google Scholar] [CrossRef] [Green Version]
- Tariq, V.N.; Jeffries, P. Appressorium formation by Sclerotinia sclerotiorum: Scanning electron microscopy. Trans. Br. Mycol. Soc. 1984, 82, 645–651. [Google Scholar] [CrossRef]
- Huang, L.; Buchenauer, H.; Han, Q.; Zhang, X.; Kang, Z. Ultrastructural and cytochemical studies on the infection process of Sclerotinia sclerotiorum in oilseed rape. J. Plant Dis. Prot. 2008, 115, 9–16. [Google Scholar] [CrossRef]
- Tariq, V.N.; Jeffries, P. Ultrastructure of penetration of Phaseolus spp. by Sclerotinia sclerotiorum. Can. J. Bot. 1986, 64, 2909–2915. [Google Scholar] [CrossRef]
- Garg, H.; Li, H.; Sivasithamparam, K.; Kuo, J.; Barbetti, M.J. The infection processes of Sclerotinia sclerotiorum in cotyledon tissue of a resistant and a susceptible genotype of Brassica napus. Ann. Bot. 2010, 106, 897–908. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Liberti, D.; Li, M.; Kim, Y.T.; Hutchens, A.; Wilson, R.; Rollins, J.A. Oxaloacetate acetylhydrolase gene mutants of Sclerotinia sclerotiorum do not accumulate oxalic acid but do produce limited lesions on host plants. Mol. Plant Pathol. 2015, 16, 559–571. [Google Scholar] [CrossRef]
- Kabbage, M.; Yarden, O.; Dickman, M.B. Pathogenic attributes of Sclerotinia sclerotiorum: Switching from a biotrophic to necrotrophic lifestyle. Plant Sci. 2015, 233, 53–60. [Google Scholar] [CrossRef]
- Seifbarghi, S.; Borhan, M.H.; Wei, Y.; Coutu, C.; Robinson, S.J.; Hegedus, D.D. Changes in the Sclerotinia sclerotiorum transcriptome during Infection of Brassica napus. BMC Genom. 2017, 18, 266. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.N.; Li, T.; Guo, X.J.; Li, M.; Liu, X.Y.; Cao, J.; Tan, X.L. Sclerotinia stem rot resistance in rapeseed: Recent progress and future prospects. J. Agric. Food Chem. 2021, 69, 2965–2978. [Google Scholar] [CrossRef]
- Williams, B.; Kabbage, M.; Kim, H.J.; Britt, R.; Dickman, M.B. Tipping the balance: Sclerotinia sclerotiorum secreted oxalic acid suppresses host defenses by manipulating the host redox environment. PLoS Pathog. 2011, 7, e1002107. [Google Scholar] [CrossRef] [Green Version]
- Chittem, K.; Yajima, W.R.; Goswami, R.S.; Del Río Mendoza, L.E. Transcriptome analysis of the plant pathogen Sclerotinia sclerotiorum interaction with resistant and susceptible canola (Brassica napus) lines. PLoS ONE 2020, 15, e0229844. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Li, J.; Yu, B.; Liu, L.; Zhang, X.; Liu, J.; Pan, H.; Zhang, Y. Transcription Factor SsSte12 was involved in mycelium growth and development in Sclerotinia sclerotiorum. Front. Microbiol. 2018, 9, 2476. [Google Scholar] [CrossRef]
- Derbyshire, M.C.; Newman, T.E.; Khentry, Y.; Owolabi, T.A. The evolutionary and molecular features of the broad-host-range plant pathogen Sclerotinia sclerotiorum. Mol. Plant Pathol. 2022, 23, 1075–1090. [Google Scholar] [CrossRef]
- Cessna, S.G.; Sears, V.E.; Low, P.S.; Dickman, M. Oxalic acid, a pathogenicity factor for Sclerotinia sclerotiorum, suppresses the oxidative burst of the host plant. Plant Cell. 2000, 12, 2191–2200. [Google Scholar] [CrossRef] [Green Version]
- Bateman, D.F.; Beer, S.V. Simultaneous production and synergistic action of oxalic acid and polygalacturonase during pathogenesis by Sclerotium rolfsii. Phytopathology 1965, 55, 204–211. [Google Scholar]
- Favaron, F.; Sella, L.; D’Ovidio, R. Relationships among endo-polygalacturonase, oxalate, pH, and plant polygalacturonase-inhibiting protein (PGIP) in the interaction between Sclerotinia sclerotiorum and soybean. Mol. Plant Microbe Interact. 2004, 17, 1402–1409. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, R.L.; Stotz, H.U. Oxalate production by Sclerotinia sclerotiorum deregulates guard cells during infection. Plant Physiol. 2004, 136, 3703–3711. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.S.; Min, J.Y.; Dickman, M.B. Oxalic acid is an elicitor of plant programmed cell death during Sclerotinia sclerotiorum disease development. Mol. Plant Microb. Interact. 2008, 21, 605–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heller, A.; Witt-Geiges, T. Oxalic acid has an additional, detoxifying function in Sclerotinia sclerotiorum pathogenesis. PLoS ONE 2013, 8, e72292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagundes-Nacarath, I.R.F.; Debona, D.; Oliveira, A.T.H.; Hawerroth, C.; Rodrigues, F.A. Biochemical responses of common bean to white mold potentiated by phosphites. Plant Physiol. Biochem. 2018, 132, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Dutton, M.V.; Evans, C.S. Oxalate production by fungi: Its role in pathogenicity and ecology in the soil environment. Can. J. Microbiol. 1996, 42, 881–895. [Google Scholar] [CrossRef]
- Liberti, D.; Rollins, J.A.; Dobinson, K.F. Peroxisomal carnitine acetyl transferase influences host colonization capacity in Sclerotinia sclerotiorum. Mol. Plant Microb. Interact. 2013, 26, 768–780. [Google Scholar] [CrossRef] [Green Version]
- Rollins, J.; Cuomo, C.; Dickman, M.; Kohn, L. Genomics of Sclerotinia sclerotiorum. In Genomics of Plant-Associated Fungi and Oomycetes: Dicot Pathogens; Dean, R.A., Lichens-Park, A., Kole, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–17. ISBN 978-3-662-44056-8. [Google Scholar]
- Magro, P.; Marciano, P.; Lenna, P. Enzymatic oxalate decarboxylation in isolates of Sclerotinia sclerotiorum. FEMS Microbiol. Lett. 1988, 49, 49–52. [Google Scholar] [CrossRef]
- Mäkelä, M.R.; Hildén, K.; Lundell, T.K. Oxalate decarboxylase: Biotechnological update and prevalence of the enzyme in filamentous fungi. Appl. Microbiol. Biotechnol. 2010, 87, 801–814. [Google Scholar] [CrossRef]
- Held, M.A.; Jiang, N.; Basu, D.; Showalter, A.M.; Faik, A. Plant Cell Wall Polysaccharides: Structure and Biosynthesis. In Polysaccharides; Ramawat, K., Mérillon, J.M., Eds.; Springer: Cham, Switzerland, 2015; ISBN 978-3-319-03751-6. [Google Scholar]
- Blanco-Ulate, B.; Morales-Cruz, A.; Amrine, K.C.H.; Labavitch, J.M.; Powell, A.L.T.; Cantu, D. Genome-wide transcriptional profiling of Botrytis cinerea genes targeting plant cell walls during infections of different hosts. Front. Plant Sci. 2014, 5, 435. [Google Scholar] [CrossRef] [Green Version]
- Amselem, J.; Cuomo, J.C.A.; van Kan, J.A.L.; Viaud, M.; Benito, E.P.; Couloux, A.; Coutinho, P.M.; de Vries, R.P.; Dyer, P.S.; Fillinger, S.; et al. Genomic analysis of the necrotrophic fungal pathogens Sclerotinia sclerotiorum and Botrytis cinerea. PLoS Genet. 2011, 7, e1002230. [Google Scholar] [CrossRef] [Green Version]
- Kars, I.; Van Kan, J.A. Extracellular enzymes and metabolites involved in pathogenesis of Botrytis. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 99–118. ISBN 978-1-4020-6586-6. [Google Scholar]
- Wagner, F.; Kusserow, H.; Schafer, W. Cloning and targeted disruption of two polygalacturonase genes in Penicillium olsonii. FEMS Microbiol. Lett. 2000, 186, 293–299. [Google Scholar] [CrossRef]
- Garcia-Maceira, F.I.; Di Pietro, A.; Huertas-Gonzalez, M.D.; Ruiz-Roldan, M.C.; Roncero, M.I.G. Molecular characterization of an endopolygalacturonase from Fusarium oxysporum expressed during early stages of infection. Appl. Environ. Microbiol. 2001, 67, 2191–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kars, I.; Krooshof, G.H.; Wagemakers, L.; Joosten, R.; Benen, J.A.; Van Kan, J.A. Necrotizing activity of five Botrytis cinerea endopolygalacturonases produced in Pichia pastoris. Plant J. 2005, 43, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.; Gerday, C.; Feller, G. Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. 2005, 29, 3–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Xiao, J.; Du, J.; Yang, Y.; Bi, C.; Qing, L. Disruption of the gene encoding endo-β-1, 4-xylanase affects the growth and virulence of Sclerotinia sclerotiorum. Front. Microbiol. 2016, 7, 1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heard, S.; Brown, N.A.; Hammond-Kosack, K. An interspecies comparative analysis of the predicted secretomes of the necrotrophic plant pathogens Sclerotinia sclerotiorum and Botrytis cinerea. PLoS ONE 2015, 10, e0130534. [Google Scholar] [CrossRef] [Green Version]
- Zuppini, A.; Navazio, L.; Sella, L.; Castiglioni, C.; Favaron, F.; Mariani, P. An endopolygalacturonase from Sclerotinia sclerotiorum induces calcium-mediated signaling and programmed cell death in soybean cells. Mol. Plant Microb. Interact. 2005, 18, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.X.; Ma, H.X.; Chen, Y. Sensitivity of Sclerotinia sclerotiorum to boscalid in Jiangsu Province of China. Crop Prot. 2009, 28, 882–886. [Google Scholar] [CrossRef]
- Dallal Bashi, Z.; Hegedus, D.D.; Buchwaldt, L.; Rimmer, S.R.; Borhan, M.H. Expression and regulation of Sclerotinia sclerotiorum necrosis and ethylene-inducing peptides (NEPs). Mol. Plant Pathol. 2010, 11, 43–53. [Google Scholar] [CrossRef]
- Zhu, W.; Wei, W.; Fu, Y.; Cheng, J.; Xie, J.; Li, G.; Yi, X.; Kang, Z.; Dickman, M.B.; Jiang, D. A secretory protein of necrotrophic fungus Sclerotinia sclerotiorum that suppresses host resistance. PLoS ONE 2013, 8, e53901. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Rollins, J.A. An oxalate decarboxylase gene functions in the early infection processes of Sclerotinia sclerotiorum. Phytopathology 2013, 103, 81. [Google Scholar]
- Zhang, H.; Wu, Q.; Cao, S.; Zhao, T.; Chen, L.; Zhuang, P.; Zhou, X.; Gao, Z. A novel protein elicitor (SsCut) from Sclerotinia sclerotiorum induces multiple defense responses in plants. Plant Mol. Biol. 2014, 86, 495–511. [Google Scholar] [CrossRef]
- Lyu, X.; Shen, C.; Fu, Y.; Xie, J.; Jiang, D.; Li, G.; Cheng, J. A small secreted virulence-related protein is essential for the necrotrophic interactions of Sclerotinia sclerotiorum with its host plants. PLoS Pathog. 2016, 12, e1005435. [Google Scholar] [CrossRef]
- Lyu, X.; Shen, C.; Fu, Y.; Xie, J.; Jiang, D.; Li, G.; Cheng, J. The microbial opsin homolog Sop1 is involved in Sclerotinia sclerotiorum development and environmental stress response. Front. Microbiol. 2016, 6, 1504. [Google Scholar] [CrossRef] [Green Version]
- Kabbage, M.; Williams, B.; Dickman, M.B. Cell death control: The interplay of apoptosis and autophagy in the pathogenicity of Sclerotinia sclerotiorum. PLoS Pathog. 2013, 9, e1003287. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Xie, J.; Cheng, J.; Li, G.; Yi, X.; Jiang, D.; Fu, Y. Novel secretory protein Ss-Caf1 of the plant-pathogenic fungus Sclerotinia sclerotiorum is required for host penetration and normal sclerotial development. Mol. Plant Microb. Interact. 2014, 27, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Xiao, J.; Yang, Y.; Bi, C.; Qing, L.; Tan, W. Ss-Bi1 encodes a putative BAX inhibitor-1 protein that is required for full virulence of Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2015, 90, 115–122. [Google Scholar] [CrossRef]
- Xu, L.; Chen, W. Random T-DNA mutagenesis identifies a Cu/Zn superoxide dismutase gene as a virulence factor of Sclerotinia sclerotiorum. Mol. Plant Microb. Interact. 2013, 26, 431–441. [Google Scholar] [CrossRef] [Green Version]
- Yarden, O.; Veluchamy, S.; Dickman, M.B.; Kabbage, M. Sclerotinia sclerotiorum catalase SCAT1 affects oxidative stress tolerance, regulates ergosterol levels and controls pathogenic development. Physiol. Mol. Plant Pathol. 2014, 85, 34–41. [Google Scholar] [CrossRef]
- Pan, Y.; Xu, Y.; Li, X.; Yao, C.; Gao, Z. SsPemG1 encodes an elicitor-homologous protein and regulates pathogenicity in Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2015, 92, 70–78. [Google Scholar] [CrossRef]
- Yang, G.; Tang, L.; Gong, Y.; Xie, J.; Fu, Y.; Jiang, D.; Li, G.; Collinge, D.B.; Chen, W.; Cheng, J. A cerato-platanin protein SsCP1 targets plant PR1 and contributes to virulence of Sclerotinia sclerotiorum. New Phytol. 2018, 217, 739–755. [Google Scholar] [CrossRef] [Green Version]
- Seifbarghi, S.; Borhan, M.H.; Wei, Y.; Ma, L.; Coutu, C.; Bekkaoui, D.; Hegedus, D.D. Receptor-Like Kinases BAK1 and SOBIR1 are required for necrotizing activity of a novel group of Sclerotinia sclerotiorum necrosis-inducing effectors. Front. Plant Sci. 2020, 11, 1021. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, Y. Trick or treat: Microbial pathogens evolved apoplastic effectors modulating plant susceptibility to infection. Mol. Plant Microb. Interact. 2018, 31, 6–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyon, K.; Balague, C.; Roby, D.; Raffaele, S. Secretome analysis reveals effector candidates associated with broad host range necrotrophy in the fungal plant pathogen Sclerotinia sclerotiorum. BMC Genom. 2014, 15, 336. [Google Scholar] [CrossRef] [Green Version]
- Derbyshire, M.; Denton-Giles, M.; Hegedus, D.; Seifbarghy, S.; Rollins, J.; Van Kan, J.; Seidl, M.F.; Faino, L.; Mbengue, M.; Navaud, O.; et al. The complete genome sequence of the Phytopathogenic fungus Sclerotinia sclerotiorum reveals insights into the genome architecture of broad host range pathogens. Genome Biol. Evol. 2017, 9, 593–618. [Google Scholar] [CrossRef]
- Harel, A.; Bercovich, S.; Yarden, O. Calcineurin is required for sclerotial development and pathogenicity of Sclerotinia sclerotiorum in an oxalic acid-independent manner. Mol. Plant Microb. Interact. 2006, 19, 682–693. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Chen, C.; Kabbage, M.; Dickman, M.B. Identification and characterization of Sclerotinia sclerotiorum NADPH oxidases. Appl. Environ. Microbiol. 2011, 77, 7721–7729. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Ge, C.; Liu, S.; Wang, J.; Zhou, M. A two-component histidine kinase Shk1 controls stress response, sclerotial formation and fungicide resistance in Sclerotinia sclerotiorum. Mol. Plant Pathol. 2013, 14, 708–718. [Google Scholar] [CrossRef]
- Qu, X.; Yu, B.; Liu, J.; Zhang, X.; Li, G.; Zhang, D.; Li, L.; Wang, X.; Wang, L.; Chen, J.; et al. MADS-Box transcription factor SsMADS is involved in regulating growth and virulence in Sclerotinia sclerotiorum. Int. J. Mol. Sci. 2014, 15, 8049–8062. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.B.; Ge, C.Y.; Zhou, M.G. Molecular and biochemical characterization of Sclerotinia sclerotiorum laboratory mutants resistant to dicarboximide and phenylpyrrole fungicides. J. Pest Sci. 2014, 87, 221–230. [Google Scholar] [CrossRef]
- Lyu, X.; Shen, C.; Fu, Y.; Xie, J.; Jiang, D.; Li, G.; Cheng, J. Comparative genomic and transcriptional analyses of the carbohydrate-active enzymes and secretomes of phytopathogenic fungi reveal their significant roles during infection and development. Sci. Rep. 2015, 5, 15565. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Moomaw, E.W.; Rollins, J.A. Fungal oxalate decarboxylase activity contributes to Sclerotinia sclerotiorum early infection by affecting both compound appressoria development and function. Mol. Plant Pathol. 2015, 16, 825–836. [Google Scholar] [CrossRef]
- Fan, H.; Yu, G.; Liu, Y.; Zhang, X.; Liu, J.; Zhang, Y.; Rollins, J.A.; Sun, F.; Pan, H. An atypical forkhead containing transcription factor SsFKH1 is involved in sclerotial formation and is essential for pathogenicity in Sclerotinia sclerotiorum. Mol. Plant Pathol. 2016, 18, 963–975. [Google Scholar] [CrossRef]
- Yu, Y.; Xiao, J.; Zhu, W.; Yang, Y.; Mei, J.; Bi, C.; Qian, W.; Qing, L.; Tan, W. Ss-Rhs1, a secretory Rhs repeat-containing protein, is required for the virulence of Sclerotinia sclerotiorum. Mol. Plant Pathol. 2017, 18, 1052–1061. [Google Scholar] [CrossRef]
- Liang, Y.; Xiong, W.; Steinkellner, S.; Feng, J. Deficiency of the melanin biosynthesis genes SCD1 and THR1 affects sclerotial development and vegetative growth, but not pathogenicity, in Sclerotinia sclerotiorum. Mol. Plant Pathol. 2018, 19, 1444–1453. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wang, Q.; Zhang, X.; Liu, J.; Zhang, Y.; Pan, H. Ssams2, a gene encoding GATA transcription factor, is required for appressoria formation and chromosome segregation in Sclerotinia sclerotiorum. Front. Microbiol. 2018, 9, 3031. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wang, Q.; Sun, Y.; Zhang, Y.; Zhang, X.; Liu, J.; Yu, G.; Pan, H. Sssfh1, a gene encoding a putative component of the RSC chromatin remodeling complex, is involved in hyphal growth, reactive oxygen species accumulation, and pathogenicity in Sclerotinia sclerotiorum. Front. Microbiol. 2018, 9, 1828. [Google Scholar] [CrossRef]
- Yu, Y.; Du, J.; Wang, Y.; Zhang, M.; Huang, Z.; Cai, J.; Fang, A.; Yang, Y.; Qing, L.; Bi, C.; et al. Survival factor 1 contributes to the oxidative stress response and is required for full virulence of Sclerotinia sclerotiorum. Mol. Plant Pathol. 2019, 20, 895–906. [Google Scholar] [CrossRef] [Green Version]
- Westrick, N.M.; Ranjan, A.; Jain, S.; Grau, C.R.; Smith, D.L.; Kabbage, M. Gene regulation of Sclerotinia sclerotiorum during Infection of Glycine max: On the road to pathogenesis. BMC Genom. 2019, 20, 157. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xiao, K.; Li, M.; Hu, H.; Zhang, X.; Liu, J.; Pan, H.; Zhang, Y. SsAGM1-mediated uridine diphosphate-n-acetylglucosamine synthesis is essential for development, stress response, and pathogenicity of Sclerotinia sclerotiorum. Front. Microbiol. 2022, 13, 938784. [Google Scholar] [CrossRef]
- Cong, J.; Xiao, K.; Jiao, W.; Zhang, C.; Zhang, X.; Liu, J.; Zhang, Y.; Pan, H. The Coupling between cell wall integrity mediated by MAPK Kinases and SsFkh1 is involved in sclerotia formation and pathogenicity of Sclerotinia sclerotiorum. Front. Microbiol. 2022, 13, 816091. [Google Scholar] [CrossRef] [PubMed]
- Mio, T.; Yamada-Okabe, T.; Arisawa, M.; Yamada-Okabe, H. Functional cloning and mutational analysis of the human cDNA for phosphoacetylglucosamine mutase: Identification of the amino acid residues essential for the catalysis. Biochim. Biophys. Acta Gene Struct. Exp. 2000, 1492, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Du, T.; Raimi, O.G.; Hurtado-Guerrero, R.; Marino, K.; Ibrahim, A.F.; Albarbarawi, O.; Ferguson, M.A.J.; Jin, C.; Van Aalten, D.M.F. Genetic and structural validation of Aspergillus fumigatus N-acetylphosphoglucosamine mutase as an antifungal target. Biosci. Rep. 2013, 33, e00063. [Google Scholar] [CrossRef] [PubMed]
- Rollins, J.A.; Dickman, M.B. Increase in endogenous and exogenous cyclic AMP levels inhibits sclerotial development in Sclerotinia sclerotiorum. Appl. Environ. Microbiol. 1998, 64, 2539–2544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Dickman, M.B. cAMP blocks MAPK activation and sclerotial development via Rap-1 in a PKA-independent manner in Sclerotinia sclerotiorum. Mol. Microbiol. 2005, 55, 299–311. [Google Scholar] [CrossRef]
- Doehlemann, G.; Berndt, P.; Hahn, M. Different signalling pathways involving a G alpha protein, cAMP and a MAP kinase control germination of Botrytis cinerea conidia. Mol. Microbiol. 2006, 59, 821–835. [Google Scholar] [CrossRef]
- Jurick II, W.M.; Dickman, M.B.; Rollins, J.A. Characterization and functional analysis of a cAMP-dependent protein kinase A catalytic subunit gene (pka1) in Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2004, 64, 155–163. [Google Scholar] [CrossRef]
- Jurick, W.M.; Rollins, J.A. Deletion of the adenylate cyclase (sac1) gene affects multiple developmental pathways and pathogenicity in Sclerotinia sclerotiorum. Fungal Genet. Biol. 2007, 44, 521–530. [Google Scholar] [CrossRef]
- Girard, V.; Fèvre, M.; Bruel, C. Involvement of cyclic AMP in the production of the acid protease Acp1 by Sclerotinia sclerotiorum. FEMS Microbiol. Lett. 2004, 237, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Hashiba, T.; Ishikawa, T. Effect of adenosine 30, 50-cyclic monophosphate on induction of sclerotia in Rhizoctonia solani. Phytopathology 1978, 68, 1723–1727. [Google Scholar] [CrossRef]
- Hadar, Y.; Pines, M.; Chet, I.; Henis, Y. The regulation of sclerotium initiation in sclerotium rolfsii by glucose and cyclic-AMP. Can. J. Microbiol. 1983, 29, 21–26. [Google Scholar] [CrossRef]
- Rosenberg, G.B.; Pall, M.L. Properties of two cyclic-nucleotide deficient mutants of Neurospora crassa. J. Bacteriol. 1979, 137, 1140–1144. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Harel, A.; Gorovoits, R.; Yarden, O.; Dickman, M.B. MAPK regulation of sclerotial development in Sclerotinia sclerotiorum is linked with pH and cAMP sensing. Mol. Plant Microb. Interact. 2004, 17, 404–413. [Google Scholar] [CrossRef] [Green Version]
- Harel, A.; Gorovits, R.; Yarden, O. Changes in protein kinase A activity accompany sclerotial development in Sclerotinia sclerotiorum. Phytopathology 2005, 95, 397–404. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef]
- Yu, P.L.; Rollins, J.A. The cAMP-dependent protein kinase A pathway perturbs autophagy and plays important roles in development and virulence of Sclerotinia sclerotiorum. Fungal Biol. 2022, 126, 20–34. [Google Scholar] [CrossRef]
- Erental, A.; Dickman, M.B.; Yarden, O. Sclerotial development in Sclerotinia sclerotiorum: Awakening molecular analysis of a “Dormant” structure. Fungal Bio. Rev. 2008, 22, 6–16. [Google Scholar] [CrossRef]
- Erental, A.; Harel, A.; Yarden, O. Type 2A phosphoprotein phosphatase is required for asexual development and pathogenesis of Sclerotinia sclerotiorum. Mol Plant Microb. Interact. 2007, 20, 944–954. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Cheng, X.; Liu, L.; Liu, S. Genome sequence resource for the plant pathogen Sclerotinia sclerotiorum WH6 isolated in China. Plant Dis. 2021, 105, 3720–3722. [Google Scholar] [CrossRef]
- Lv, Z.; He, Z.; Hao, L.; Kang, X.; Ma, B.; Li, H.; Luo, Y.; Yuan, J.; He, N. Genome sequencing analysis of Scleromitrula shiraiana, a causal agent of mulberry sclerotial disease with narrow host range. Front. Microbiol. 2021, 11, 603927. [Google Scholar] [CrossRef]
- Atwell, S.; Corwin, J.A.; Soltis, N.E.; Subedy, A.; Denby, K.J.; Kliebenstein, D.J. Whole genome resequencing of Botrytis cinerea isolates identifies high levels of standing diversity. Front. Microbiol. 2015, 6, 996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.R.; Pan, H.; et al. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 2005, 434, 980–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zampounis, A.; Pigné, S.; Dallery, J.F.; Wittenberg, A.H.; Zhou, S.; Schwartz, D.C.; Thon, M.R.; O’Connell, R.J. Genome sequence and annotation of Colletotrichum higginsianum, a causal agent of crucifer anthracnose disease. Genome Announc. 2016, 4, e00821-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buiate, E.A.S.; Xavier, K.V.; Moore, N.; Torres, M.F.; Farman, M.L.; Schardl, C.L.; Vaillancourt, L.J. A comparative genomic analysis of putative pathogenicity genes in the host-specific sibling species Colletotrichum graminicola and Colletotrichum sublineola. BMC Genom. 2017, 18, 67. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Liu, H.; Li, Z.; Ke, X.; Dou, D.; Gao, X.; Song, N.; Dai, Q.; Wu, Y.; Xu, J.R.; et al. Genome sequence of Valsa canker pathogens uncovers a potential adaptation of colonization of woody bark. New Phytol. 2015, 208, 1202–1216. [Google Scholar] [CrossRef]
- Spanu, P.D.; Abbott, J.C.; Amselem, J.; Burgis, T.A.; Soanes, D.M.; Stüber, K.; Ver Loren van Themaat, E.; Brown, J.K.; Butcher, S.A.; Gurr, S.J.; et al. Genome expansion and gene loss in powdery mildew fungi reveal tradeoffs in extreme parasitism. Science 2010, 330, 1543–1546. [Google Scholar] [CrossRef]
- Galagan, J.E.; Calvo, S.E.; Borkovich, K.A.; Selker, E.U.; Read, N.D.; Jaffe, D.; FitzHugh, W.; Ma, L.J.; Smirnov, S.; Purcell, S.; et al. The genome sequence of the filamentous fungus Neurospora crassa. Nature 2003, 422, 859–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, R.; Urban, M.; Hammond-Kosack, M.C.; Hassani-Pak, K.; Hammond-Kosack, K.E. The completed genome sequence of the pathogenic ascomycete fungus Fusarium graminearum. BMC Genom. 2015, 16, 544. [Google Scholar] [CrossRef] [Green Version]
- Galagan, J.E.; Calvo, S.E.; Cuomo, C.; Ma, L.J.; Wortman, J.R.; Batzoglou, S.; Lee, S.I.; Baştürkmen, M.; Spevak, C.C.; Clutterbuck, J.; et al. Sequencing of Aspergillus nidulans and comparative analysis with A. fumigatus and A. oryzae. Nature 2005, 438, 1105–1115. [Google Scholar] [CrossRef] [Green Version]
- Hane, J.K.; Lowe, R.G.; Solomon, P.S.; Tan, K.C.; Schoch, C.L.; Spatafora, J.W.; Crous, P.W.; Kodira, C.; Birren, B.W.; Galagan, J.E.; et al. Dothideomycete plant interactions illuminated by genome sequencing and EST analysis of the wheat pathogen Stagonospora nodorum. Plant Cell. 2007, 19, 3347–3368. [Google Scholar] [CrossRef] [Green Version]
- Wyatt, N.A.; Richards, J.K.; Brueggeman, R.S.; Friesen, T.L. Reference assembly and annotation of the Pyrenophora teres f. teres isolate 0-1. G3 2018, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Duplessis, S.; Cuomo, C.A.; Lin, Y.C.; Aerts, A.; Tisserant, E.; Veneault-Fourrey, C.; Joly, D.L.; Hacquard, S.; Amselem, J.; Cantarel, B.L.; et al. Obligate biotrophy features unraveled by the genomic analysis of rust fungi. Proc. Natl. Acad. Sci. USA 2011, 108, 9166–9171. [Google Scholar] [CrossRef] [Green Version]
- Andrew, M.; Barua, R.; Short, S.M.; Kohn, L.M. Evidence for a common toolbox based on necrotrophy in a fungal lineage spanning necrotrophs, biotrophs, endophytes, host generalists and specialists. PLoS ONE 2012, 7, e29943. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Annotation | Gene Functions | Reference |
|---|---|---|---|
| Sspac1 | pH-responsive transcription factor-encoding gene | Radial growth, sclerotial development (having the melanized rind layer), and virulence | [25] |
| Sscna1 | Catalytic subunit calcineurin-encoding gene | Virulence | [81] |
| PP2B | Type 2B phosphatase | Maturation of sclerotium | [81] |
| Ssnox1 | NADPH oxidase | Sclerotial formation | [82] |
| SsITL | Integrin-like protein | Virulence, hyphal polarity, and sclerotial formation | [64] |
| Sssod1 | Cu/Zn-superoxide dismutase | Hyphal growth, sclerotial formation, virulence, oxidative stress tolerance, host ROS repression, and fungicide resistance | [83] |
| SsMADS | MADS-box transcription factor | Aerial growth, sclerotial development, and virulence | [84] |
| Scat1 | Type A catalase | Radial growth, larger and melanized sclerotia, and osmotic stress tolerance | [73] |
| Shk1 | Histidine kinase | Radial growth, sclerotial formation, oxidative stresses, and glycerol accumulation | [85] |
| SsCVNH | Cysteine-rich, small secreted protein with a carbohydrate-binding module | Normal mycelial growth and sclerotial formation | [86] |
| Ssoah1 | Oxaloacetate acetylhydrolase | Radial growth, compound appressorium formation, sclerotial development, and pathogenesis | [31] |
| Ssodc1 | Oxalate decarboxylase | Vegetative hyphae, apothecia, early phases of compound appressorium formation, and during plant colonization | [87] |
| Ssodc2 | Oxalate decarboxylase | Middle-to-late phases of compound appressorium formation | [87] |
| Ss-Bi1 | BAX inhibitor-1 | Full virulence | [71] |
| Ss-sop1 | Microbial opsin homolog gene | Radial growth, sclerotial formation, tolerance to salt, and osmotic and cell wall stresses | [68] |
| Ss-Xyl1 | Endo-β-1, 4-xylanase | Normal colony morphology, radial growth, apothecia producing fertile sclerotial formation, and pathogenicity | [59] |
| Ss-SSVP1 | Small secreted virulence-related protein | Slightly necessary for radial growth | [67] |
| rhs1 | Rearrangement hotspot repeat-containing protein | Hyphal growth, sclerotial development, and virulence | [89] |
| SCD1 | Scytalone dehydratase | Vegetative growth and sclerotial development | [26] |
| THR1 | Trihydroxynaphthalene reductase | Vegetative growth and sclerotial development | [26] |
| Ss-ams2 | A protein predicted to contain GATA-box domain | Radial growth, appressorium formation, pathogenicity, and chromosome segregation | [91] |
| Ss-sfh1 | GATA transcription factor with an SNF5 domain | Radial growth, ROS accumulation, and virulence | [92] |
| Ss-Ste12 | Ste12 transcription factor | Mycelial growth, sclerotial development, appressorium formation, and penetration-dependent pathogenicity. | [37] |
| Ss-svf1 | Survival factor 1 homologue | Appressorium formation and virulence | [93] |
| Ss-rhs1 | Rhs repeat-containing protein | Sclerotial development and hyphal infection | [94] |
| Ss-Fkh1 | Forkhead transcription factor | Hyphal growth, sclerotial formation, and tolerance to oxidative and osmotic stresses | [88,96] |
| Ss-AGM1 | N-acetylglucosamine-phosphate mutase | Chitin production, cell wall integrity of mycelia, infection cushions, sclerotial formation, and virulence | [95] |
| Genomes | Phylum | Nutrition Mode | Genome Size (Mb) | GC (%) | Chromosome | Genes | Proteins | Genes/ Genome | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Sclerotinia sclerotiorum 1980 | Ascomycota | Necrotroph | 38.9 | 41.55 | 16 | 11,368 | 11,130 | 292.24 | [53] |
| Scleromitrula shiraiana SX-001 | Ascomycota | Necrotroph | 39.0 | 38.85 | n/a | 11,327 | 11,327 | 290.44 | [115] |
| Botrytis cinerea B05.10 | Ascomycota | Necrotroph | 42.6 | 42.00 | 18 | 14,262 | 13,703 | 334.79 | [116] |
| Magnaporthe oryzae 70-15 (MG8) | Ascomycota | Hemibiotroph | 41.0 | 51.59 | 7 | 13,184 | 12,989 | 321.56 | [117] |
| Colletotrichum higginsianum IMI 349063 | Ascomycota | Hemibiotroph | 50.7 | 54.41 | 11 | 14,650 | 14,650 | 288.95 | [118] |
| Colletotrichum graminicola M1.001 | Ascomycota | Hemibiotroph | 51.6 | 49.09 | n/a | 12,387 | 12,020 | 240.06 | [119] |
| Valsa mali 03-8 | Ascomycota | Necrotroph | 44.7 | 49.35 | 13 | 11,284 | 11,284 | 252.44 | [120] |
| Valsa pyri SXYL134 | Ascomycota | Necrotroph | 35.7 | 51.70 | n/a | 10,855 | 10,855 | 304.06 | [120] |
| Blumeria graminis f. sp. hordei DH14 | Ascomycota | Obligate biotroph | 118.7 | 45.40 | n/a | 7061 | 6495 | 59.49 | [121] |
| Neurospora crassa OR74A | Ascomycota | Saprotroph | 41.1 | 48.23 | 7 | 10,785 | 10,785 | 262.41 | [122] |
| Fusarium graminearum PH-1 | Ascomycota | Hemibiotroph | 38.0 | 48.15 | 4 | 14,145 | 14,143 | 372.24 | [123] |
| Aspergillus nidulans FGSC_A4 | Ascomycota | Saprotroph | 29.8 | 50.37 | 8 | 10,597 | 10,534 | 355.60 | [124] |
| Parastagonospora nodorum SN15 | Ascomycota | Necrotroph | 37.2 | 50.37 | n/a | 12,773 | 12,391 | 343.36 | [125] |
| Pyrenophora teres f. sp. teres 0-1 | Ascomycota | Necrotroph | 33.5 | 50.9 | n/a | 12,103 | 11,799 | 361.28 | [126] |
| Puccinia graminis f. sp. tritici CRL 75-36-700-3 | Basidiomycota | Obligate biotroph | 88.7 | 43.80 | n/a | 16,304 | 15,979 | 183.81 | [127] |
| Melampsora larici-populina 98AG31 | Basidiomycota | Obligate biotroph | 109.8 | 41.00 | 18 | 17,140 | 16,372 | 178.05 | [128] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.M.; Sultana, F.; Li, W.; Tran, L.-S.P.; Mostofa, M.G. Sclerotinia sclerotiorum (Lib.) de Bary: Insights into the Pathogenomic Features of a Global Pathogen. Cells 2023, 12, 1063. https://doi.org/10.3390/cells12071063
Hossain MM, Sultana F, Li W, Tran L-SP, Mostofa MG. Sclerotinia sclerotiorum (Lib.) de Bary: Insights into the Pathogenomic Features of a Global Pathogen. Cells. 2023; 12(7):1063. https://doi.org/10.3390/cells12071063
Chicago/Turabian StyleHossain, Md. Motaher, Farjana Sultana, Weiqiang Li, Lam-Son Phan Tran, and Mohammad Golam Mostofa. 2023. "Sclerotinia sclerotiorum (Lib.) de Bary: Insights into the Pathogenomic Features of a Global Pathogen" Cells 12, no. 7: 1063. https://doi.org/10.3390/cells12071063