

Chromatin Accessibility and Transcriptional Landscape during Inhibition of Salmonella enterica by Lactobacillus reuteri in IPEC-J2 Cells
 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culture
2.2. Zone of Inhibition Assay
2.3. Cell Viability Assay
2.4. Sample Collection
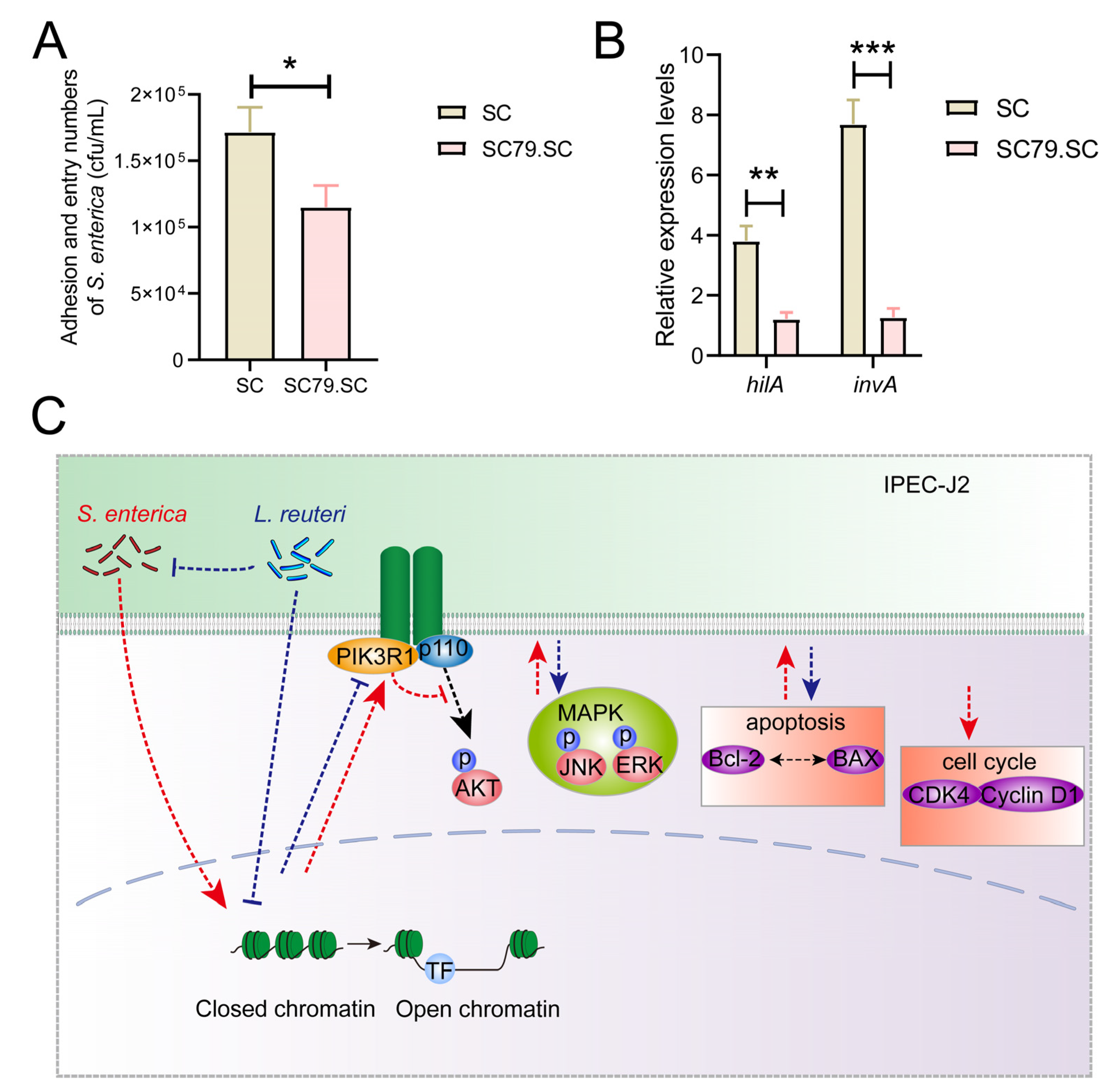
2.5. Assay of Adhesion and Entry Ability of Bacteria into IPEC-J2 Cells
2.6. Detection of Salmonella Virulence Factors hilA and invA via a qPCR Assay
2.7. RNA-Seq Library Construction
2.8. RNA-Seq Data Processing
2.9. Fuzzy C-Means Clustering
2.10. ATAC-Seq Library Construction
2.11. ATAC-Seq Data Analysis
2.12. Pathway Analysis
2.13. Integrative Analysis of ATAC-Seq and RNA-Seq Data
2.14. ChIP-qPCR
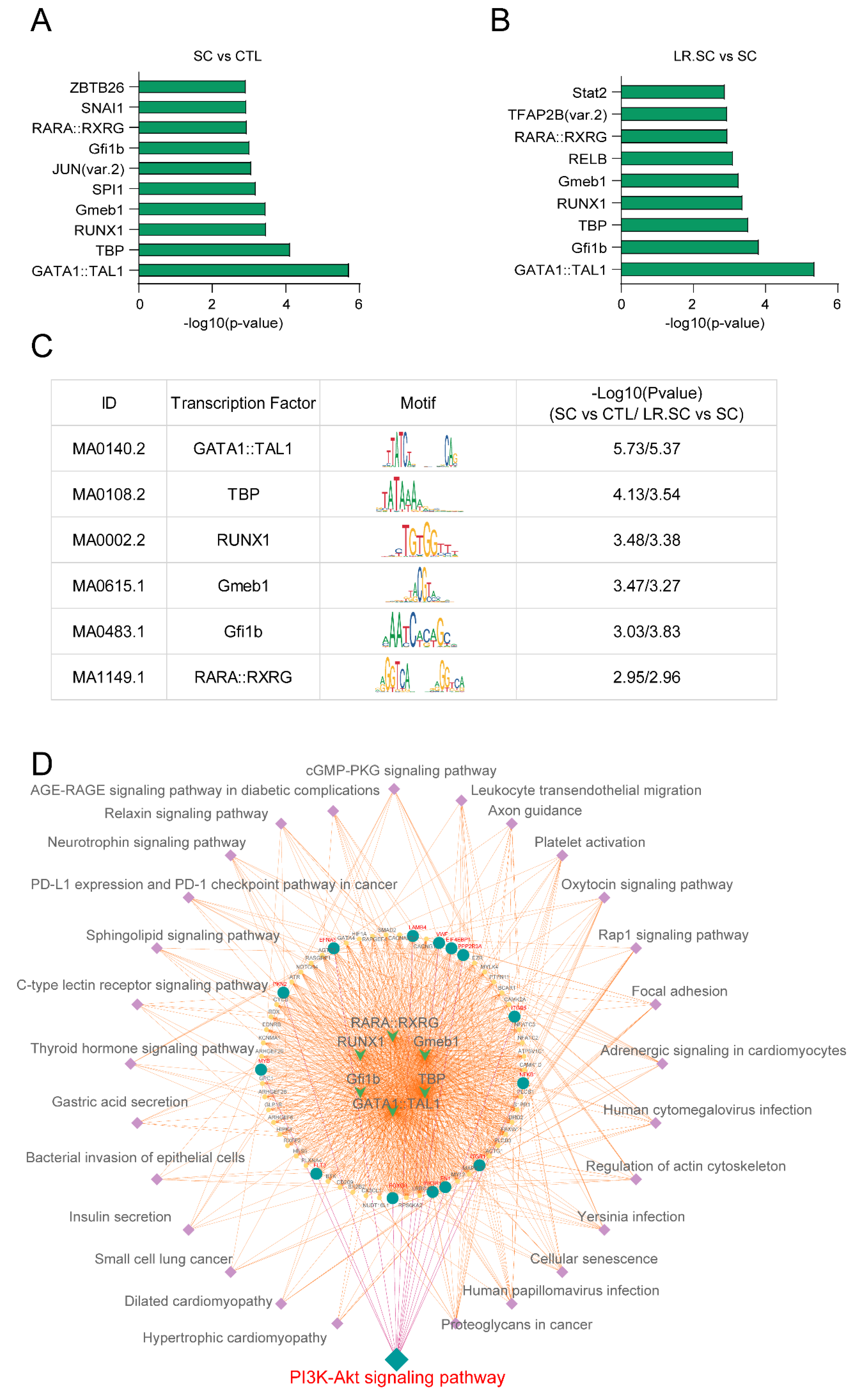
2.15. MEME Analysis
2.16. Quantitative Real-Time PCR (qRT-PCR)
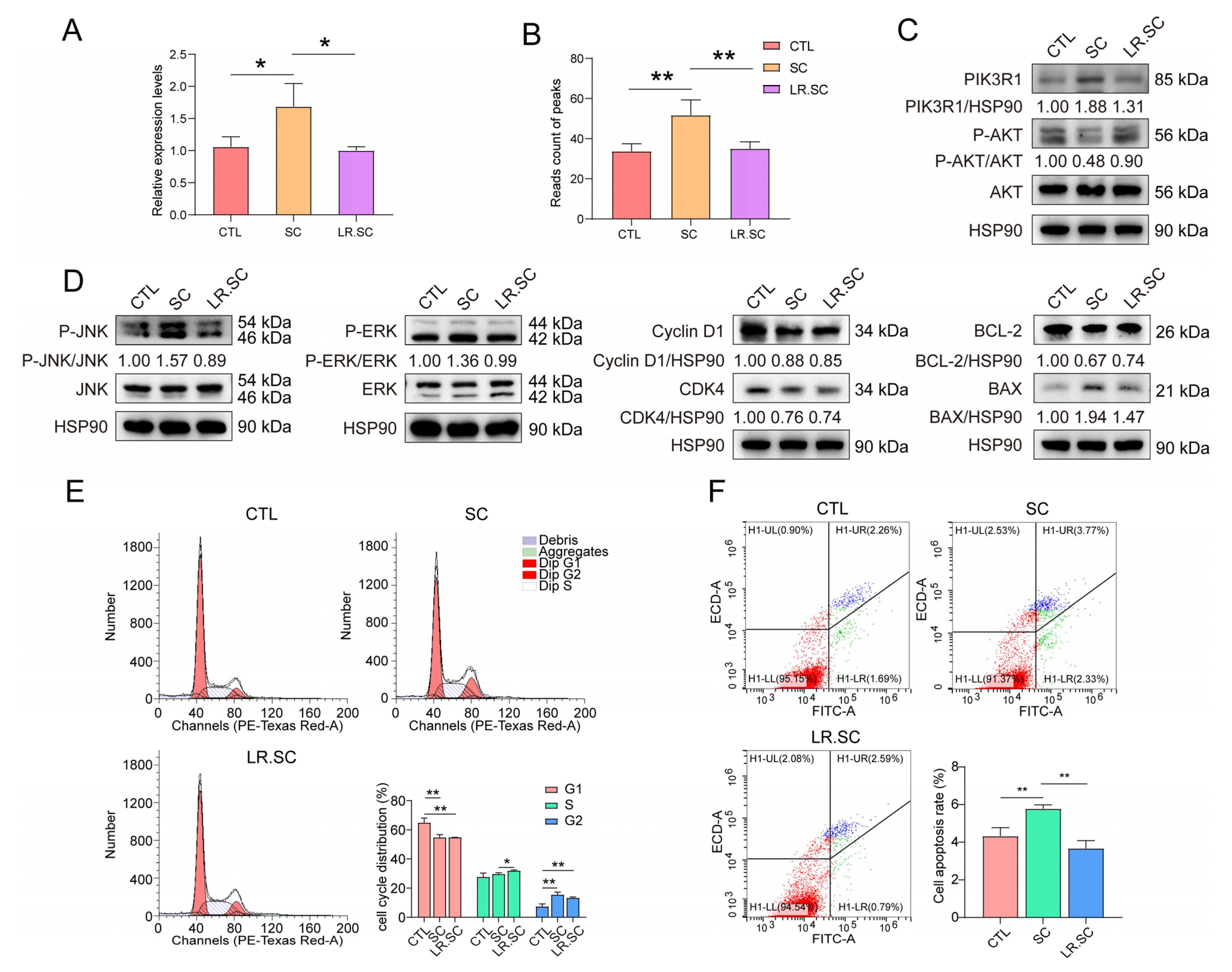
2.17. Western Blot Assay
2.18. Cell Cycle and Apoptosis Assay
2.19. Statistical Analysis and Data Availability
3. Results
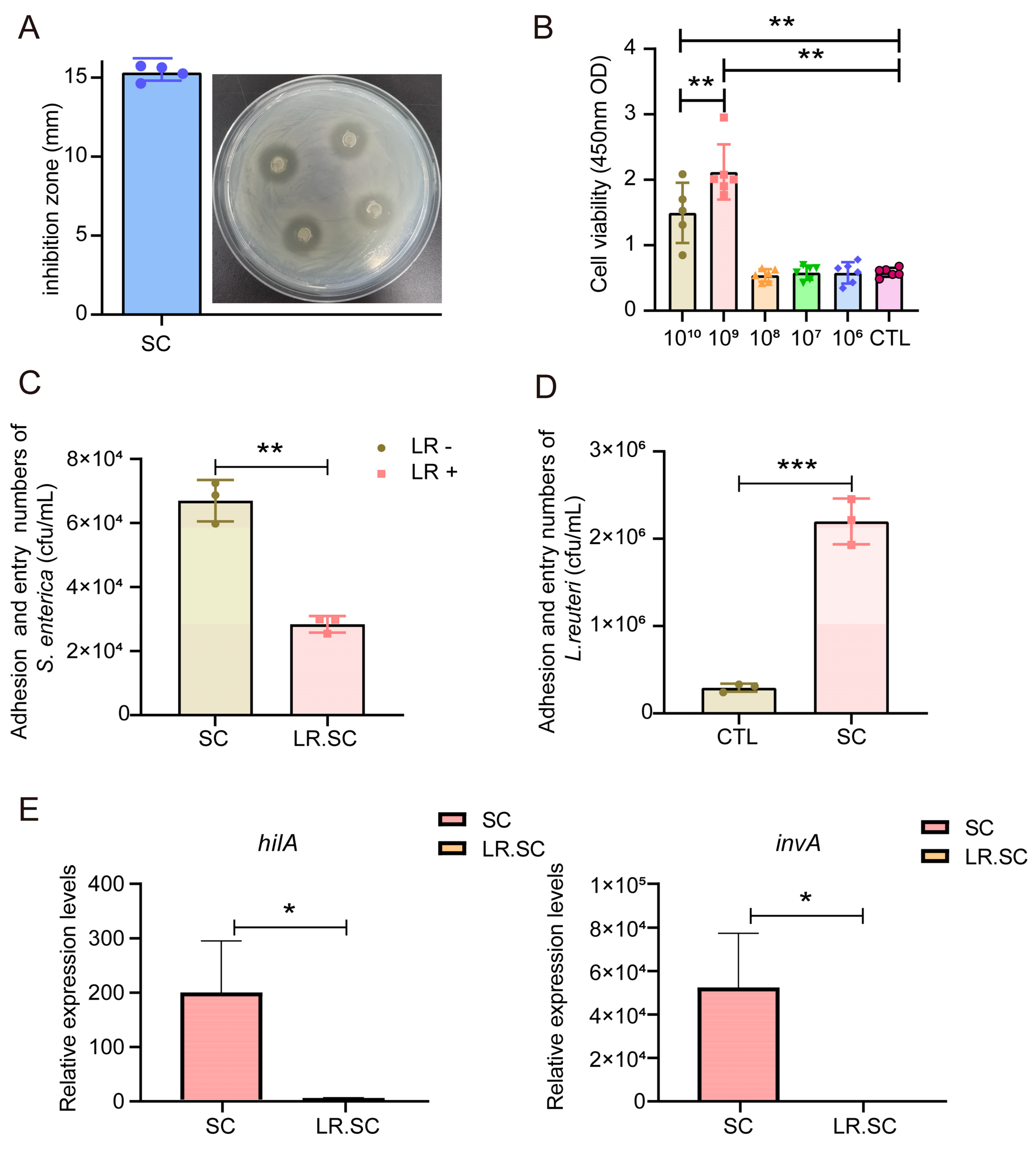
3.1. L. reuteri ATCC 53,608 Inhibits S. enterica BNCC186354 In Vitro
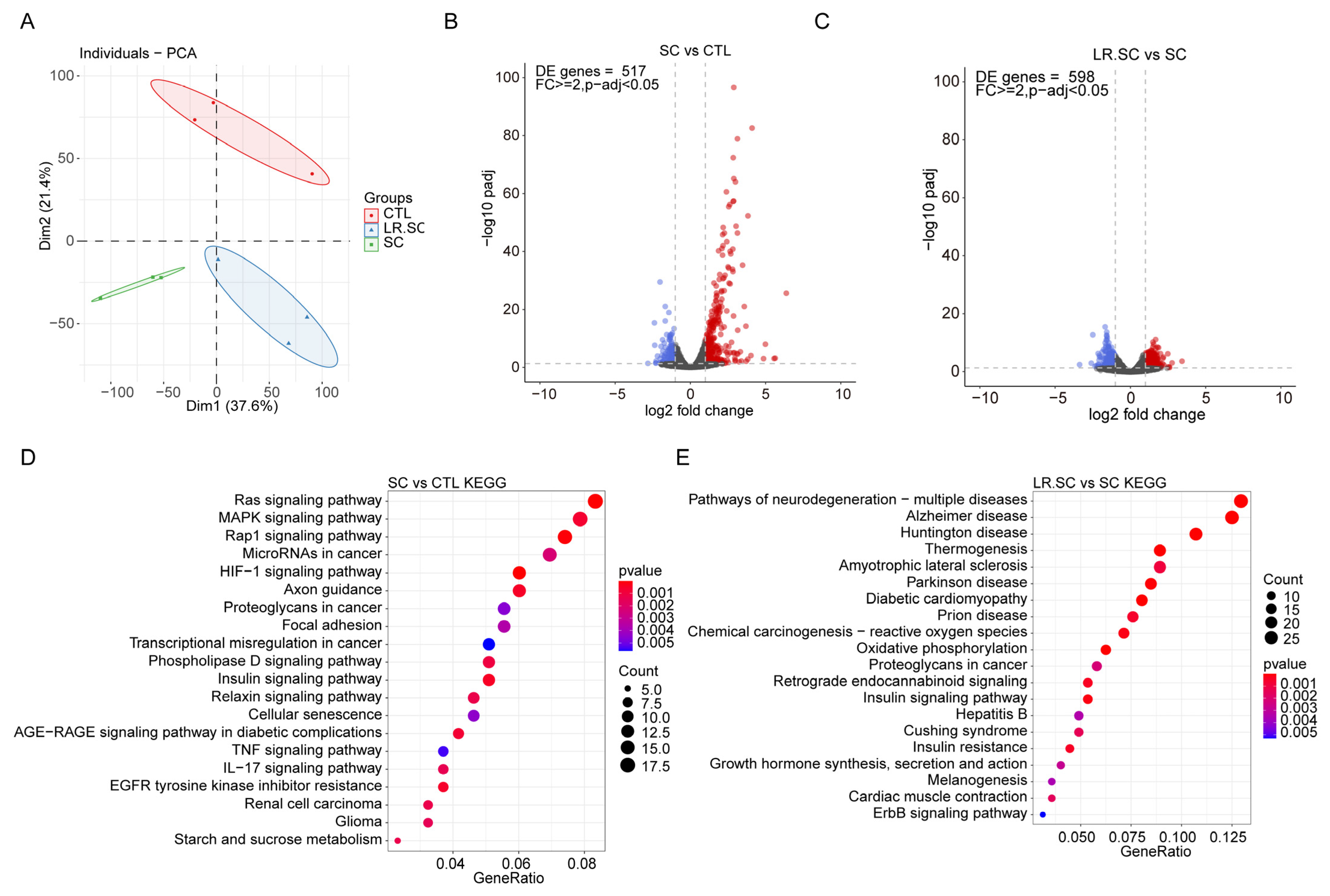
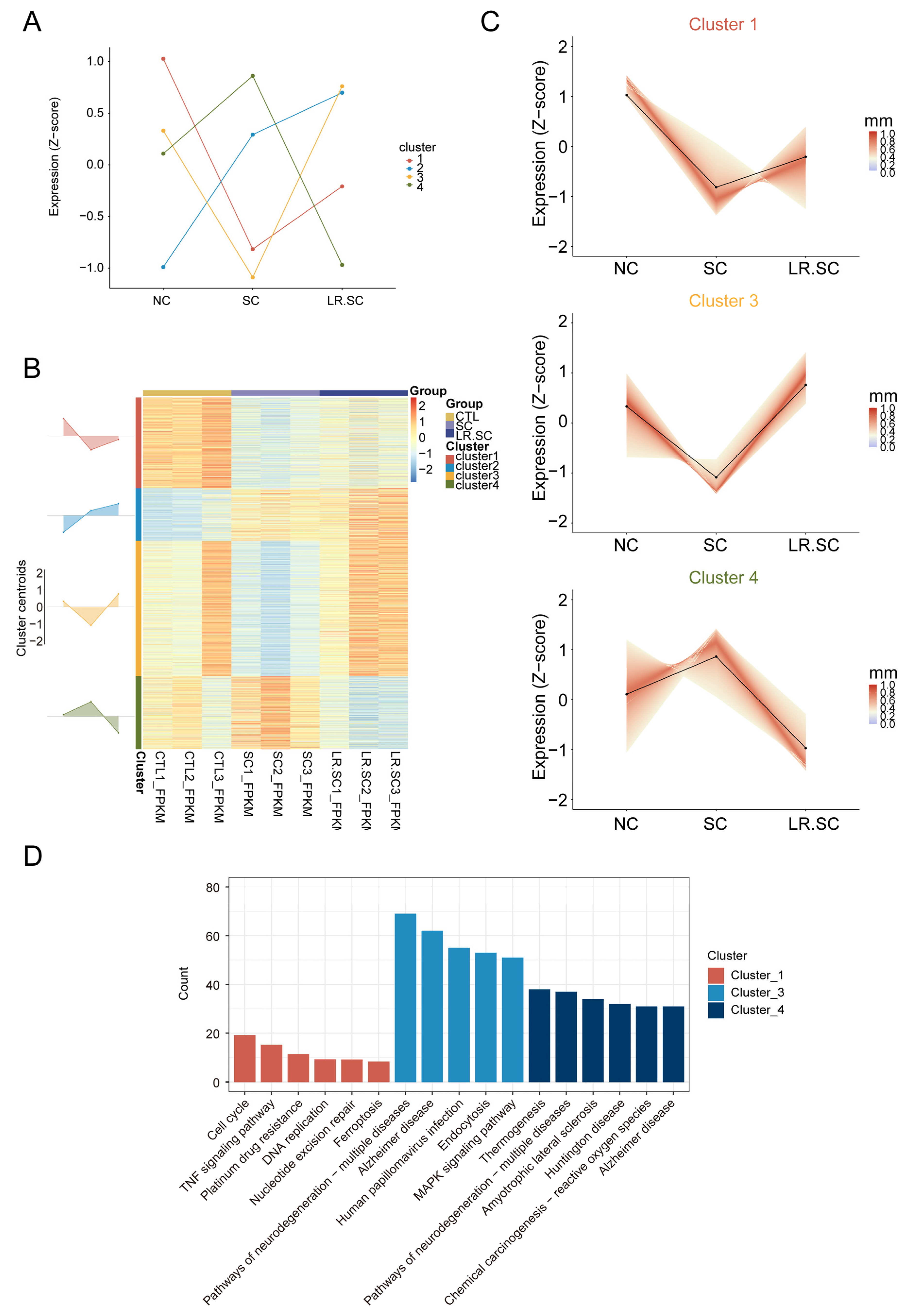
3.2. Transcriptional Landscape of IPEC-J2 Cells under the Inhibition of S. enterica BNCC186354 by L. reuteri ATCC 53608
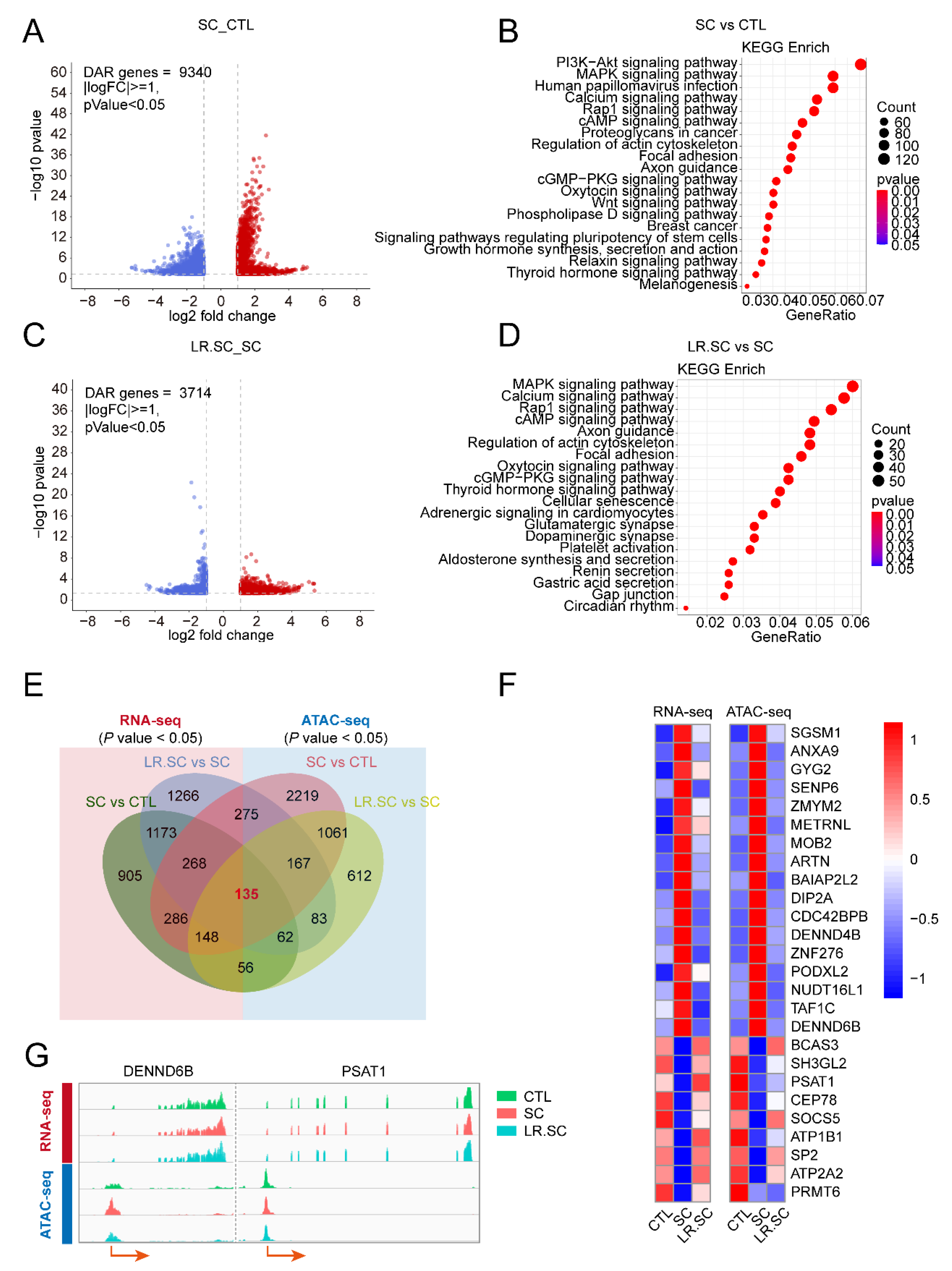
3.3. Genome-Wide Changes in Chromatin Accessibility Reveal a Pathogenic and Bacteriostatic Process
3.4. L. reuteri ATCC53608 Regulates the Apoptosis and PI3K/AKT Pathway of IPEC-J2 Cells Induced by S. enterica BNCC186354
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tian, Y.Q.; Gu, D.; Wang, F.; Liu, B.W.; Li, J.W.; Kang, X.L.; Meng, C.; Jiao, X.N.; Pan, Z.M. Prevalence and Characteristics of Salmonella spp. from a Pig Farm in Shanghai, China. Foodborne Pathog. Dis. 2021, 18, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Vinueza-Burgos, C.; Baquero, M.; Medina, J.; De Zutter, L. Occurrence, genotypes and antimicrobial susceptibility of Salmonella collected from the broiler production chain within an integrated poultry company. Int. J. Food Microbiol. 2019, 299, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Blondel, C.J.; Jiménez, J.C.; Contreras, I.; Santiviago, C.A. Comparative genomic analysis uncovers 3 novel loci encoding type six secretion systems differentially distributed in Salmonella serotypes. BMC Genom. 2009, 10, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narm, K.E.; Kalafatis, M.; Slauch, J.M. HilD, HilC, and RtsA Form Homodimers and Heterodimers to Regulate Expression of the Salmonella Pathogenicity Island I Type III Secretion System. J. Bacteriol. 2020, 202, e00012-20. [Google Scholar] [CrossRef] [PubMed]
- Beshiru, A.; Igbinosa, I.H.; Igbinosa, E.O. Prevalence of Antimicrobial Resistance and Virulence Gene Elements of Salmonella Serovars from Ready-to-Eat (RTE) Shrimps. Front. Microbiol. 2019, 10, 1613. [Google Scholar] [CrossRef] [Green Version]
- Mejia, L.; Vela, G.; Zapata, S. High Occurrence of Multiresistant Salmonella Infantis in Retail Meat in Ecuador. Foodborne Pathog. Dis. 2021, 18, 41–48. [Google Scholar] [CrossRef]
- Kumar, T.; Rajora, V.R.; Arora, N. Prevalence of Salmonella in pigs and broilers in the Tarai region of Uttarakhand, India. Indian J. Med. Microbiol. 2014, 32, 99–101. [Google Scholar] [CrossRef]
- Fairbrother, J.M.; Nadeau, E.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [Green Version]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in Human Health and Diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef] [Green Version]
- Han, S.K.; Kim, J.K.; Joo, M.K.; Lee, K.E.; Han, S.W.; Kim, D.H. Lactobacillus reuteri NK33 and Bifidobacterium adolescentis NK98 Alleviate Escherichia coli-Induced depression and Gut Dysbiosis in Mice. J. Microbiol. Biotechnol. 2020, 30, 1222–1226. [Google Scholar] [CrossRef]
- De Angelis, M.; Siragusa, S.; Berloco, M.; Caputo, L.; Settanni, L.; Alfonsi, G.; Amerio, M.; Grandi, A.; Ragni, A.; Gobbetti, M. Selection of potential probiotic lactobacilli from pig feces to be used as additives in pelleted feeding. Res. Microbiol. 2006, 157, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Casas, I.A.; Dobrogosz, W.J. Validation of the Probiotic Concept: Lactobacillus reuteri Confers Broad-spectrum Protection against Disease in Humans and Animals. Microb. Ecol. Health Dis. 2000, 12, 247–285. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Wang, F.; Wang, H.; Wu, S.; Bao, W. Exploring a Possible Link between the Fecal Microbiota and the Production Performance of Pigs. Vet. Sci. 2022, 9, 527. [Google Scholar] [CrossRef]
- Ortiz-Rivera, Y.; Sanchez-Vega, R.; Gutierrez-Mendez, N.; Leon-Felix, J.; Acosta-Muniz, C.; Sepulveda, D.R. Production of reuterin in a fermented milk product by Lactobacillus reuteri: Inhibition of pathogens, spoilage microorganisms, and lactic acid bacteria. J. Dairy Sci. 2017, 100, 4258–4268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.L.; Forsythe, P.; Bienenstock, J. Live Lactobacillus reuteri is essential for the inhibitory effect on tumor necrosis factor alpha-induced interleukin-8 expression. Infect. Immun. 2004, 72, 5308–5314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Rath, E.; Holzlwimmer, G.; Quintanilla-Martinez, L.; Loach, D.; Tannock, G.; Haller, D. Lactobacillus reuteri 100-23 transiently activates intestinal epithelial cells of mice that have a complex microbiota during early stages of colonization. J. Nutr. 2008, 138, 1684–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, K.L.; Doyle, J.S.; Jewell, L.D.; Tavernini, M.M.; Fedorak, R.N. Lactobacillus species prevents colitis in interleukin 10 gene-deficient mice. Gastroenterology 1999, 116, 1107–1114. [Google Scholar] [CrossRef]
- Pena, J.A.; Li, S.Y.; Wilson, P.H.; Thibodeau, S.A.; Szary, A.J.; Versalovic, J. Genotypic and phenotypic studies of murine intestinal lactobacilli: Species differences in mice with and without colitis. Appl. Environ. Microbiol. 2004, 70, 558–568. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.M.; Wang, H.F.; Liu, J.X.; Zhao, Y.H.; Gao, K.; Zhang, J. Adhesive ability means inhibition activities for lactobacillus against pathogens and S-layer protein plays an important role in adhesion. Anaerobe 2013, 22, 97–103. [Google Scholar] [CrossRef]
- Buddhasiri, S.; Sukjoi, C.; Kaewsakhorn, T.; Nambunmee, K.; Nakphaichit, M.; Nitisinprasert, S.; Thiennimitr, P. Anti-inflammatory Effect of Probiotic Limosilactobacillus reuteri KUB-AC5 Against Salmonella Infection in a Mouse Colitis Model. Front. Microbiol. 2021, 12, 716761. [Google Scholar] [CrossRef]
- Walsham, A.D.; MacKenzie, D.A.; Cook, V.; Wemyss-Holden, S.; Hews, C.L.; Juge, N.; Schuller, S. Lactobacillus reuteri Inhibition of Enteropathogenic Escherichia coli Adherence to Human Intestinal Epithelium. Front. Microbiol. 2016, 7, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazakevych, J.; Denizot, J.; Liebert, A.; Portovedo, M.; Mosavie, M.; Jain, P.; Stellato, C.; Fraser, C.; Corrêa, R.O.; Célestine, M.; et al. Smarcad1 mediates microbiota-induced inflammation in mouse and coordinates gene expression in the intestinal epithelium. Genome Biol. 2020, 21, 64. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.; Zhang, L.; Hu, M.Y.; Xu, Y.Y.; Hou, Y.; Li, X.Y.; Zhao, S.H.; Zhao, Y.X.; Li, C.C. Identification and Conservation Analysis of Cis-Regulatory Elements in Pig Liver. Genes 2019, 10, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattani, A.M.; Siqueira, F.M.; Guedes, R.L.M.; Schrank, I.S. Repetitive Elements in Mycoplasma hyopneumoniae Transcriptional Regulation. PLoS ONE 2016, 11, e0168626. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.S.; Zhou, S.S.; Hua, K.X.; Xiao, H.D.; Li, Z.L.; Liu, M.; Luo, R.; Bi, D.R.; Zhou, R.; Jin, H. Haemophilus parasuis infection activates chemokine RANTES in PK-15 cells. Mol. Immunol. 2015, 67, 661–666. [Google Scholar] [CrossRef]
- Thurman, R.E.; Rynes, E.; Humbert, R.; Vierstra, J.; Maurano, M.T.; Haugen, E.; Sheffield, N.C.; Stergachis, A.B.; Wang, H.; Vernot, B.; et al. The accessible chromatin landscape of the human genome. Nature 2012, 489, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Carey, M.; Workman, J.L. The role of chromatin during transcription. Cell 2007, 128, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Voss, T.C.; Hager, G.L. Dynamic regulation of transcriptional states by chromatin and transcription factors. Nat. Rev. Genet. 2014, 15, 69–81. [Google Scholar] [CrossRef]
- Buenrostro, J.D.; Giresi, P.G.; Zaba, L.C.; Chang, H.Y.; Greenleaf, W.J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat. Methods 2013, 10, 1213–1218. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-First Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2010. [Google Scholar]
- Yi, H.B.; Wang, L.; Xiong, Y.X.; Wang, Z.L.; Qiu, Y.Q.; Wen, X.L.; Jiang, Z.Y.; Yang, X.F.; Ma, X.Y. Lactobacillus reuteri LR1 Improved Expression of Genes of Tight Junction Proteins via the MLCK Pathway in IPEC-1 Cells during Infection with Enterotoxigenic Escherichia coli K88. Mediat. Inflamm. 2018, 2018, 6434910. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.B.; Hou, C.L.; Wang, G.; Jia, H.M.; Yu, H.T.; Zeng, X.F.; Thacker, P.A.; Zhang, G.L.; Qiao, S.Y. Lactobacillus reuteri I5007 Modulates Intestinal Host Defense Peptide Expression in the Model of IPEC-J2 Cells and Neonatal Piglets. Nutrients 2017, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skjolaas, K.A.; Burkey, T.E.; Dritz, S.S.; Minton, J.E. Effects of Salmonella enterica serovar Typhimurium, or serovar Choleraesuis, Lactobacillus reuteri and Bacillus licheniformis on chemokine and cytokine expression in the swine jejunal epithelial cell line, IPECJ2. Vet. Immunol. Immunopathol. 2007, 115, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.; Futschik, M.E. Mfuzz: A software package for soft clustering of microarray data. Bioinformation 2007, 2, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.J.E.J. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, F.; Dundar, F.; Diehl, S.; Gruning, B.A.; Manke, T. deepTools: A flexible platform for exploring deep-sequencing data. Nucleic Acids Res. 2014, 42, W187–W191. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; He, Q.Y. ChIPseeker: An R/Bioconductor package for ChIP peak annotation, comparison and visualization. Bioinformatics 2015, 31, 2382–2383. [Google Scholar] [CrossRef] [Green Version]
- Stark, R.; Brown, G. DiffBind: Differential binding analysis of ChIP-Seq peak data. R package Version 2011, 100, 4-3. Available online: http://bioconductor.org/packages/release/bioc/vignettes/DiffBind/inst/doc/DiffBind.pdf (accessed on 2 March 2023).
- Machanick, P.; Bailey, T.L. MEME-ChIP: Motif analysis of large DNA datasets. Bioinformatics 2011, 27, 1696–1697. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.R.; Yan, F.; Li, Q.R.; Liang, Y.P.; Yu, J.X.; Li, Z.J.; He, F.F.; Li, R.P.; Li, M. Chlorogenic Acid Promotes Autophagy and Alleviates Salmonella Typhimurium Infection Through the lncRNAGAS5/miR-23a/PTEN Axis and the p38 MAPK Pathway. Front. Cell Dev. Biol. 2020, 8, 552020. [Google Scholar] [CrossRef] [PubMed]
- Calo, E.; Wysocka, J. Modification of enhancer chromatin: What, how, and why? Mol. Cell 2013, 49, 825–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Liu, D.; Song, P.; Jiang, F.; Chi, X.; Zhang, T. Exposure to hypoxia causes stress erythropoiesis and downregulates immune response genes in spleen of mice. BMC Genom. 2021, 22, 413. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Peng, Z.; Lin, X.; Luo, H.; Song, M.; Jin, L.; Xiao, X.; Ji, H. Study on Interaction Between TATA-Box Binding Protein (TBP), TATA-Box and Multiprotein Bridging Factor 1(MBF1) in Beauveria bassiana by Graphene-Based Electrochemical Biosensors. Front. Chem. 2020, 8, 278. [Google Scholar] [CrossRef] [PubMed]
- An, W.; Yao, S.; Sun, X.; Hou, Z.; Lin, Y.; Su, L.; Liu, X. Glucocorticoid modulatory element-binding protein 1 (GMEB1) interacts with the de-ubiquitinase USP40 to stabilize CFLARL and inhibit apoptosis in human non-small cell lung cancer cells. J. Exp. Clin. Cancer Res. 2019, 38, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelman, J.A.; Luo, J.; Cantley, L.C. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat. Rev. Genet. 2006, 7, 606–619. [Google Scholar] [CrossRef]
- Alzahrani, A.S. PI3K/Akt/mTOR inhibitors in cancer: At the bench and bedside. Semin. Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef]
- Hennessy, B.T.; Smith, D.L.; Ram, P.T.; Lu, Y.L.; Mills, G.B. Exploiting the PI3K/AKT pathway for cancer drug discovery. Nat. Rev. Drug Discov. 2005, 4, 988–1004. [Google Scholar] [CrossRef]
- Kong, Y.; Li, Y.; Luo, Y.; Zhu, J.; Zheng, H.; Gao, B.; Guo, X.; Li, Z.; Chen, R.; Chen, C. circNFIB1 inhibits lymphangiogenesis and lymphatic metastasis via the miR-486-5p/PIK3R1/VEGF-C axis in pancreatic cancer. Mol. Cancer 2020, 19, 82. [Google Scholar] [CrossRef]
- Jing, Z.T.; Liu, W.; Xue, C.R.; Wu, S.X.; Chen, W.N.; Lin, X.J.; Lin, X. AKT activator SC79 protects hepatocytes from TNF-α-mediated apoptosis and alleviates d-Gal/LPS-induced liver injury. Am. J. Physiol. Gastrointest Liver Physiol. 2019, 316, G387–G396. [Google Scholar] [CrossRef]
- Arques, J.L.; Fernandez, J.; Gaya, P.; Nunez, M.; Rodriguez, E.; Medina, M. Antimicrobial activity of reuterin in combination with nisin against food-borne pathogens. Int. J. Food Microbiol. 2004, 95, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Price, A.E.; Shamardani, K.; Lugo, K.A.; Deguine, J.; Roberts, A.W.; Lee, B.L.; Barton, G.M. A Map of Toll-like Receptor Expression in the Intestinal Epithelium Reveals Distinct Spatial, Cell Type-Specific, and Temporal Patterns. Immunity 2018, 49, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faba, L.; Litjens, R.; Allaart, J.; van den Hil, P.R. Feed additive blends fed to nursery pigs challenged with Salmonella. J. Anim. Sci. 2020, 98, skz382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Liu, X.; Yu, H.; Gong, J. Lactobacillus Regulates Caenorhabditis elegans Cell Signaling to Combat Salmonella Infection. Front. Immunol. 2021, 12, 653205. [Google Scholar] [CrossRef]
- Huang, T.; Huang, X.; Shi, B.; Wang, F.; Feng, W.; Yao, M. Regulators of Salmonella-host interaction identi-fied by peripheral blood transcriptome profiling: Roles of TGFB1 and TRP53 in intracellular Salmonella replication in pigs. Vet. Res. 2018, 49, 121. [Google Scholar] [CrossRef] [Green Version]
- Dar, M.A.; Ahmad, S.M.; Bhat, B.A.; Dar, T.A.; Haq, Z.U.; Wani, B.A.; Shabir, N.; Kashoo, Z.A.; Shah, R.A.; Ganai, N.A.; et al. Comparative RNA-Seq analysis reveals insights in Salmonella disease resistance of chicken; and database development as resource for gene expression in poultry. Genomics 2022, 114, 110475. [Google Scholar] [CrossRef]
- Mambu, J.; Barilleau, E.; Fragnet-Trapp, L.; Le Vern, Y.; Olivier, M.; Sadrin, G.; Grepinet, O.; Taieb, F.; Velge, P.; Wiedemann, A. Rck of Salmonella Typhimurium Delays the Host Cell Cycle to Facilitate Bacterial Invasion. Front. Cell. Infect. Microbiol. 2020, 10, 586934. [Google Scholar] [CrossRef]
- Finn, C.E.; Chong, A.; Cooper, K.G.; Starr, T.; Steele-Mortimer, O. A second wave of Salmonella T3SS1 activity prolongs the lifespan of infected epithelial cells. PLoS Pathog. 2017, 13, e1006354. [Google Scholar] [CrossRef] [Green Version]
- Chu, B.X.; Li, Y.N.; Liu, N.; Yuan, L.X.; Chen, S.Y.; Zhu, Y.H.; Wang, J.F. Salmonella Infantis Delays the Death of Infected Epithelial Cells to Aggravate Bacterial Load by Intermittent Phosphorylation of Akt with SopB. Front. Immunol. 2021, 12, 757909. [Google Scholar] [CrossRef]
- Eun, C.S.; Kim, Y.S.; Han, D.S.; Choi, J.H.; Lee, A.R.; Park, Y.K. Lactobacillus casei prevents impaired barrier function in intestinal epithelial cells. APMIS 2011, 119, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Hu, R.; Guo, Y.; Sun, W.; Li, J.; Fan, M.; Wang, Y.; Du, H.; Tang, Z.; Chai, C. Influence of Lactobacillus reuteri SL001 on intestinal microbiota in AD model mice and C57BL/6 mice. Sheng Wu Gong Cheng Xue Bao 2020, 36, 1887–1898. [Google Scholar] [CrossRef] [PubMed]
- Kong, G.; Lê Cao, K.A.; Hannan, A.J. Alterations in the Gut Fungal Community in a Mouse Model of Huntington’s Disease. Microbiol. Spectr. 2022, 10, e0219221. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Huh, E.; Kim, N.; Kim, D.H.; Oh, M.S. High-throughput 16S rRNA gene sequencing reveals that 6-hydroxydopamine affects gut microbial environment. PLoS ONE 2019, 14, e0217194. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.C.B.; Carter, E.K.; Dammer, E.B.; Duong, D.M.; Gerasimov, E.S.; Liu, Y.; Liu, J.; Betarbet, R.; Ping, L.; Yin, L.; et al. Large-scale deep multi-layer analysis of Alzheimer’s disease brain reveals strong proteomic disease-related changes not observed at the RNA level. Nat. Neurosci. 2022, 25, 213–225. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, W.; Ren, Z.; Xu, C.; Cao, Y.-n.; Sun, M.-a.; Huang, R.; Bao, W. Chromatin Accessibility and Transcriptional Landscape during Inhibition of Salmonella enterica by Lactobacillus reuteri in IPEC-J2 Cells. Cells 2023, 12, 968. https://doi.org/10.3390/cells12060968
Qin W, Ren Z, Xu C, Cao Y-n, Sun M-a, Huang R, Bao W. Chromatin Accessibility and Transcriptional Landscape during Inhibition of Salmonella enterica by Lactobacillus reuteri in IPEC-J2 Cells. Cells. 2023; 12(6):968. https://doi.org/10.3390/cells12060968
Chicago/Turabian StyleQin, Weiyun, Zhanshi Ren, Chao Xu, Ya-nan Cao, Ming-an Sun, Ruihua Huang, and Wenbin Bao. 2023. "Chromatin Accessibility and Transcriptional Landscape during Inhibition of Salmonella enterica by Lactobacillus reuteri in IPEC-J2 Cells" Cells 12, no. 6: 968. https://doi.org/10.3390/cells12060968