
Various AAV Serotypes and Their Applications in Gene Therapy: An Overview

Abstract
:1. Introduction
2. Brief History of AAVs’ Discovery
3. Properties, Structure, and Genome Organization of AAVs
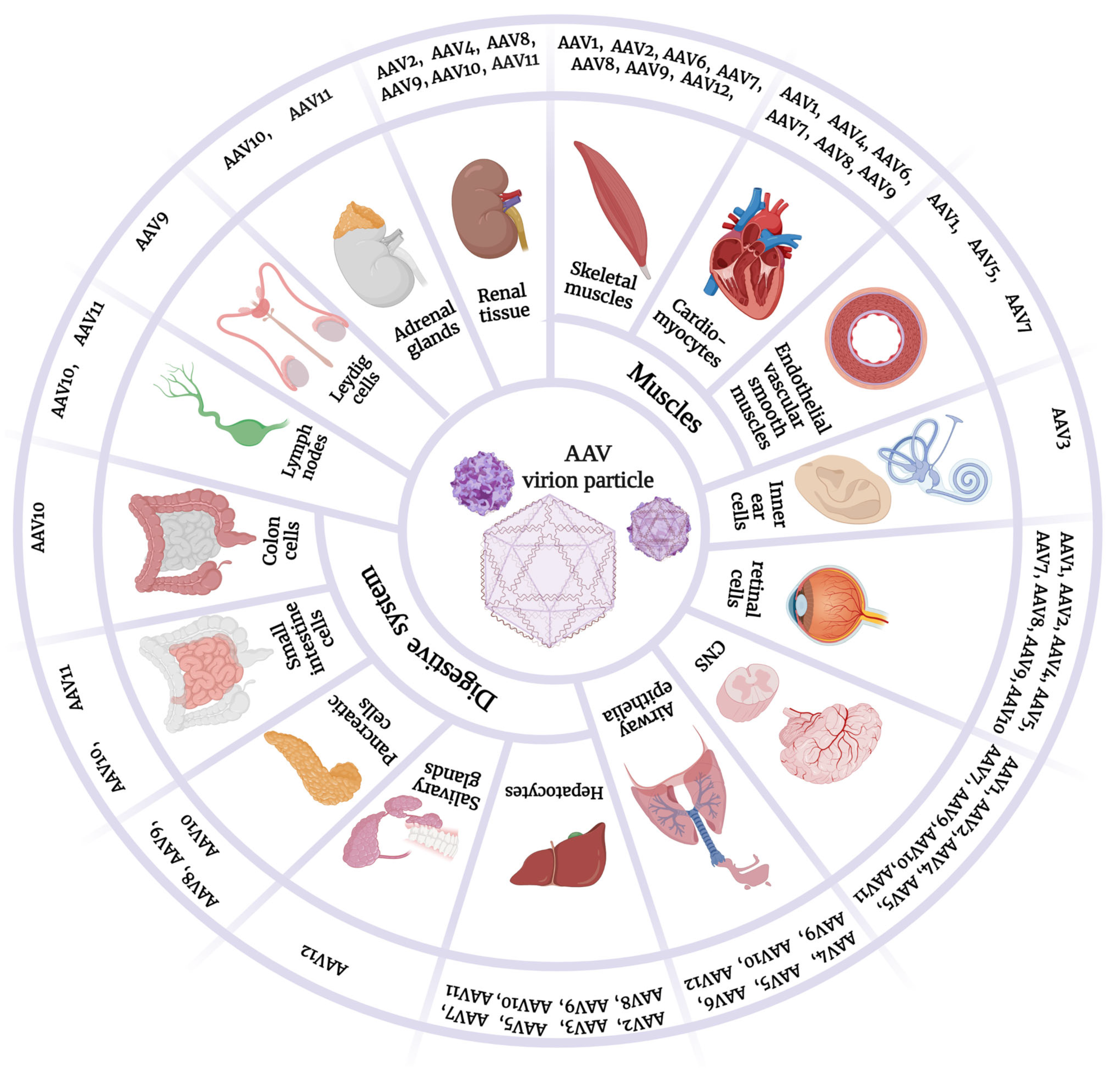
4. AAV Serotypes and Tropism
4.1. AAV1
4.2. AAV2
4.3. AAV3
4.4. AAV4
4.5. AAV5
4.6. AAV6
4.7. AAV7
4.8. AAV8
4.9. AAV9
4.10. AAV10 and AAV11
4.11. AAV12
4.12. AAV13
4.13. Novel Hybrid AAV Vectors
5. AAV as Viral Vectors for Gene Therapy Applications
5.1. AAV Viral Vectors for Gene Therapy of the CNS
5.2. AAV Viral Vectors for Gene Therapy of Respiratory Diseases
5.3. AAV Viral Vectors for Gene Therapy of Muscle Diseases
5.4. AAV Viral Vectors for Gene Therapy of Cardiovascular and Blood Diseases
5.5. AAV Viral Vectors for Gene Therapy of Liver Diseases
5.6. AAV Viral Vectors for Gene Therapy of Endocrine Disorders
5.7. AAV Viral Vectors for Gene Therapy of Cancer
6. Challenges and Limitations of AAVs for Gene Therapy AAV-Mediated Applications

{kind=link}
| AAV Serotype | Primary Receptor | Other/Coreceptors | Post Translational Modifications | Recommended Purification Method | Tropism |
|---|---|---|---|---|---|
| AAV1 | Sialic acid | AAV receptor (AAVR) | - | iodixanol gradient centrifugation, anion-exchange chromatography, and mucin column affinity chromatography | Skeletal muscles [35,43,44,45,46], heart [48,49], glial and ependymal cells in the murine brain [47], endothelial and vascular smooth muscles [50], retina [51] |
| AAV2 | HSPG | FGFR1, αVβ5 and α5β1 integrins, HGFR, LR, and CD9 | ubiquitination, phosphorylation, SUMOylation, and multiple-site-glycosylation | heparin column affinity chromatography | renal tissue [62,63,64], hepatocytes [65,66], retina [67,68,69], non-mitotic cells of central nervous system (CNS) [70,71], and skeletal muscles [45,71] |
| AAV3 | HSPG | FGFR1, LR, and HGFR | acetylation, phosphorylation, and glycosylation | iodixanol gradient centrifugation | human liver cancer cells as well as human and NHP hepatocytes [76,77], murine cochlear inner hair cells [82]. |
| AAV4 | Sialic acid | - | ubiquitination | ion-exchange chromatography, mucin column affinity chromatography | ependymal cells of mammalian CNS [90], RPE cells of the retina (canine, rodent, and NHP origins) [91], murine kidney, lung, and heart cells [92,99]. |
| AAV5 | Sialic acid | PDGFR (α and β) | ubiquitination, phosphorylation, SUMOylation, and glycosylation | ion-exchange chromatography, mucin column affinity chromatography | murine: retinal cells [38,51], mainly photoreceptors [106], airway epithelia [98,110], liver cells [92,112], vascular endothelial cells and smooth muscles [50], and neurons (murine and NHP) [107,370] |
| AAV6 | Sialic acid and HSPG | EGFR | acetylation | heparin or mucin column affinity chromatography | airway epithelia of murine and canine models [119,120], murine liver cells [92,121], skeletal muscles of murine and canine models [92,122,123,124], cardiomyocytes in murine [92,125], porcine [126], canine [127], and in sheep [128] models. |
| AAV7 | - | - | glycosylation, phosphorylation, SUMOylation, and acetylation | - | murine skeletal muscle cells [133], murine and human hepatocytes [92], murine and NHPs CNS [134,135], murine photoreceptor cells [136], murine vascular endothelial cells (limited tropism) [130], murine epicardium cells [49]. |
| AAV8 | LR | - | phosphorylation, glycosylation, and acetylation | Dual-ion-exchange chromatography, iodixanol gradient centrifugation | murine, canine, and 115 hepatocytes [92,112,141,142,143,144,145,146,147], murine skeletal and cardiac muscles [148], murine pancreatic cells [149,150], murine renal cells [151], and different cells in the murine retina [152,153,154] |
| AAV9 | terminal N-linked galactose | putative integrin, LR | glycosylation, ubiquitination, phosphorylation, SUMOylation, and acetylation | Sucrose gradient centrifugation, and ion-exchange chromatography | murine, NHP, and feline neuronal and non-neuronal cells, including astrocytes [27,159,160,161,162], murine and NHP retinal photoreceptors cells [163], murine, NHP, and porcine cardiac muscle tissue [164,165,166,167,168,169], murine hepatocytes, skeletal muscles, and pancreatic cells [81,169], photoreceptor cells [170], renal tubular epithelium cells [171,172,173], Leydig cells in the testicular interstitial tissue [174], and alveolar and nasal epithelia [175,176]. |
| AAV10 | - | - | glycosylation, ubiquitination, phosphorylation, SUMOylation, and acetylation | iodixanol gradient centrifugation | NHP intestinal cells, hepatocytes, lymph nodes, and less frequently, renal cells and adrenal glands [180], murine small intestine and colon cells [181], retinal cells (including RPE, cells in the ganglion cell layer, several cell types in the inner nuclear layer, photoreceptors, and a highly efficient transduction of horizontal cells) [152], murine liver cells, lung cells [182], renal, and pancreatic cells [183]. |
| AAV11 | - | - | - | iodixanol gradient centrifugation | NHP intestinal cells, hepatocytes, lymph nodes, and less frequently, renal cells and adrenal glands [180], murine projection neurons and astrocytes [184], and mild tropism to NHP CNS (cerebrum and spinal cord, mainly) [180]. |
| AAV12 | mannose and mannosamine have been suggested as components of a potential receptor complex | - | AVB Sepharose affinity chromatography | murine salivary glands and muscles [185], murine nasal epithelia (mainly after intranasal administration) [187]. | |
| AAV13 | - | HSPG | - | iodixanol gradient centrifugation, heparin column affinity chromatography | - |
| Gene Therapy Target | Disease ** | AAV Viral Vector | Study Type | Outcome | Ref. | |
|---|---|---|---|---|---|---|
| CNS | PD | AAV2-GAD | phase I clinical trial for advanced PD patients | safe and well-tolerated approach, providing significant improvement in motor function scores up to 12 months after unilateral subthalamic delivery | [229] | |
| double-blind, controlled, randomized clinical trial for advanced PD patients | safe and well-tolerated approach, along with improved motor function scores following bilateral subthalamic delivery | [230] | ||||
| AAV2-neurturin | open-label clinical trial for PD patients | suggested feasibility, safety, and good tolerance of the approach | [231] | |||
| rAAV2-AADC | clinical trial for moderately advanced PD patients | good tolerance, improvement on motor rating scales, however, accompanied with an increased risk of intracranial hemorrhages and headache | [232] | |||
| good tolerance and a stable expression of the transgene that lasted for the following 4 years, although higher vector doses were suggested for further studies | [233] | |||||
| AADC deficiency | rAAV2-AADC | open-label, phase I/II trial in children | good tolerance in general, with evidence for potential improvement of motor function | [234] | ||
| SMA | AA9-SMN | open-label, phase I clinical trial | significant improvement of the motor function in all 15 patients following single-dose intravenous administration, reflected by their ability to perform different activities, such as unassisted sitting and walking, oral feeding, and speaking, with no reported motor function regression at two-year follow-up. A long-term safety assessment, however, was recommended. | [236] | ||
| LSD | MPS VII | (rAAV-GUSβ) | in vivo study, murine model | stable expression of the deficient enzyme upon single administration, that was adequate for phenotype correction in the liver | [238,239] | |
| (rAAV-GUSβ) | in vivo study, neonatal murine model | therapeutic levels of the deficient enzyme in multiple organs, including ones of the CNS, with the gene expression not being affected by rapid growth and differentiation of tissues | [240] | |||
| rAAV9-GUSβ and rAAVrh10-GUSβ | in vivo study, canine model | significantly high expression levels of GUSβ in the CNS tissues, with the enzyme in brain tissue homogenates showing over 100% normal activity | [221] | |||
| MPS IIIA | AAV9-Sgsh (canine sulfamidase gene) | in vivo study, canine model | sustained and widely distributed transgene expression with no toxicity for a duration of 7 years after therapy | [241] | ||
| MLD | AAV1-ARSA | in vivo study, murine model | significant elevation of ARSA levels and activity, resulting in reduction of accumulated sulfatides | [244] | ||
| AAV5-ARSA | in vivo study, murine model | rapid, abundant, and sustained restoration of ARSA levels in the brain and brainstem up to 15 months after administration, reduction of accumulated sulfatides, and preservation of neurologic function. | [246] | |||
| AAV5-ARSA | in vivo study, NHP model | good tolerance, distribution of the transgene in the brain with elevated activity of the deficient enzyme | [370] | |||
| AAV9-ARSA | in vivo study, neonatal murine model | global expression of the transgene in the brain and spinal cord, along with muscles and heart, inhibition of sulfatide accumulation, and improvement of neurologic/motor function | [245] | |||
| AAVrh10-ARSA AAVrh10-ARSA AAVrh10-ARSA | in vivo study, murine model | correction of sulfatide accumulation following single administration, with a transduction efficacy higher than that of AAV5 as it transduced both neurons and oligodendrocytes | [243] | |||
| in vivo study, NHP model | good tolerance, neuroinflammation 3 months following the fifth dose but none after the first, detection of transgene expression after the first dose along with increased ARSA activity, and detection of the enzyme in other organs but not in gonads | [371] | ||||
| clinical trial for children with asymptomatic or early-stage MLD | significant elevation in ARSA levels in the cerebrospinal fluid (CSF) following intracerebral vector delivery, but no clinical improvement has been noticed compared to the control group. | [253] | ||||
| GM2-gangliosidose | rAAV2-HEXA + rAAV2-HEXB | in vivo study, murine model | delay of disease clinical onset, maintained motor function, good tolerance, stable and abundant levels of the deficient enzyme, resulting in a significant reduction of gangliosides’ storage in the CNS | [250] | ||
| AAVrh8-HEX | in vivo study, feline model | prevention or reduction of tremors that is characteristic of improvement in the neurologic function, and a signification elevation of the deficient enzyme levels (HEX), resulting in a significant reduction of gangliosides’ storage in different tissues of the CNS. | [252] | |||
| AAVrh8-HEXA + AAVrh8-HEXB | in vivo study, ovine model | delay of disease clinical onset and progression, improved neurologic function and clinical biomarkers. However, lifespan was not significantly higher. | [251] | |||
| AAVrh8-HEXA + AAVrh8-HEXB | first clinical trial for children with TSD | good tolerance and broad distribution of the transgene in the CNS | [248] | |||
| CD | rAAV2-ASPA | in vivo study, murine model | increased ASPA activity and, thereby, decreased NAA accumulation and white matter degeneration. However, areas remote from injection site, such as the cerebellum, were not affected | [256] | ||
| GLD | rAAV1-GALC | in vivo study, murine model | sustained expression of the deficient enzyme, improved myelination status, and prolonged lifespan. However, both treated and untreated mice died with similar symptoms, suggesting that the used approach should be initiated prior to symptoms’ onset. | [262] | ||
| rAAV2/5-GALC | in vivo study, murine model | wide dispersion of GALC transgene across the CNS reaching areas remote from the injection site, reduced loss of oligodendrocytes and Purkinje cells, along with a significant improvement of neuromotor function and a prolonged lifespan of treated mice. | [263] | |||
| AAV9-GALC AAVrh10-GALC AAVOlig001-GALC | in vivo study, murine model | All three serotypes provided wide distribution of the transgene across the CNS and liver, resulting in a significant improvement of myelination, and a prolonged lifespan, with AAV9 being the most effective when combined with bone marrow transplantation. | [265] | |||
| AAVrh10-cGALC | in vivo study, canine model | delayed symptoms onset, prolonged lifespan, correction of biochemical defects, and a positive effect on neuropathology in treated animals | [266]. | |||
| AAV9-cGALC | in vivo study, canine model | increased activity of the deficient enzyme and, therefore, normal levels of its substrate, improved myelination, and decreased inflammation both in the CNS and PNS, which, along with prevention of clinical neurological dysfunction, resulted in a significantly prolonged lifespan of treated dogs, compared to the control group. However, sufficient dosing was found to be critical, as high doses significantly extended the lifespan even for post-symptomatic subjects, and a 5-fold lower dose of the vector resulted in an attenuated form of disease | [267]. | |||
| Retinal degeneration and ocular neurovascular diseases | AAV2, 8, and 9 | in vivo study, C57BL/6 mice | transduction of retinal cells, as well as efficient transduction of ganglion cell layer by AAV8 and AAV9 | [170] | ||
| Inherited photoreceptor diseases | hybrid serotypes AAV2/7 and AAV2/8 | in vivo study, C57BL/6 mice | high transduction rates of the murine photoreceptors | [136] | ||
| Retinal blindness caused by LCA | AAV2/6 | in vivo study, wild-type 129Sv/Ev (Taconic) mice | efficient transduction of murine cone photoreceptors following subretinal injection | [268] | ||
| AAV2/4- RPE65 | clinical trial for patients with LCA | systemic and local good tolerance with no adverse effects, along with improvement of visual function presented by different parameters in different patients, including improvement of visual acuity and color vision, as well as reduction of visual fatigue or photophobia, over a follow-up period between 1 and 3 years | [269] | |||
| Retinal blindness caused by choroideremia | AAV2- REP1 | phase I/II clinical trial | improvement of retinal sensitivity in all six patients, following subretinal vector delivery, with two of them having significant increases in visual acuity, supporting further consideration of the tested therapeutic approach. | [272] | ||
| RPE65-mediated inherited retinal dystrophy | AAV2-hRPE65v2 | open-label, randomized, controlled phase III trial | good tolerance with no adverse effects, restoration of RPE65 enzymatic activity reflected by significant and sustained improvement in light perception and navigational abilities | [274] | ||
| Leber Hereditary Optic Neuropathy (LHON) | rAAV2-ND4 (gene encoding nicotinamide adenine dinucleotide dehydrogenase subunit IV) | open-label, phase I/II randomized clinical trial | good tolerance, although a mild, intraocular inflammation was detected after vector administration, but it was responsive to treatment and suggested to be overcome in further studies by systemic vector delivery instead of local. | [275] | ||
| open-label, phase I clinical trial | minor adverse events, improvement in visual activity in some but not all patients, no detection of vector DNA in patients’ blood samples | [276] | ||||
| RPGR-related X-linked retinitis pigmentosa | AAV8-RPGR | phase I/II clinical trial | no adverse effects other than steroid-responsive subretinal inflammation following administration of higher doses, sustained improvements in visual function in 6/18 patients. | [277] | ||
| X-Linked Retinoschisis | AAV8-RS1 (Retinoschisin gene) | phase I/IIa single-center, open-label, clinical trial | good tolerance in general, although steroid-responsive, dose-related inflammation was observed, and a dose-related increase of systemic antibodies against AAV8, but none against RS1 | [278,279] | ||
| CNGA3-linked achromatopsia | AAV8-CNGA3 | nonrandomized controlled clinical trial | no substantial safety issues, successful targeting of cone photoreceptors reflected by reported improvement of color vision ability, along with improvements in visual acuity and contrast sensitivity (although a cause–effect relationship was not established) | [280,281] | ||
| Hearing disorders | AAV9 | in vivo study, porcine model | persistent expression of the transgene within the mammalian inner ear following intracochlear delivery | [282] | ||
| Respiratory organs (airway epithelia) | CF | AAV2-CFTR | in vivo study-rabbits | efficient and stable gene transfer of CFTR into airway epithelium, indicating, as a result, the vector potential to be used for gene therapy | [286] | |
| phase I clinical trial for CF patients | safety, successful transduction of targeted cells, and a detected function restoration of sinuses. | [288] | ||||
| phase II, double-blind, randomized, placebo-controlled clinical trial for CF patients | safety and good tolerance, but no effective clinical treatment of disease was achieved | [289] | ||||
| phase I clinical trial for mild CF patients | safety, but no effective clinical treatment of disease was achieved | [290] | ||||
| α1AT deficiency | rAAV-hAAT | in vivo study, intravenous delivery into murine model | Serum levels of AAT that are potentially therapeutic | [297,298] | ||
| rAAV2-hAAT and rAAV5-hAAT | in vivo study, intrapleural vs. intramuscular delivery into murine model | AAT lung and serum levels higher than the ones achieved by intravenous delivery, with rAAV5 showing 10-fold higher effectiveness than rAAV2 | [299,300] | |||
| rhAAV10-hAAT | in vivo study, intrapleural delivery into murine and NHP models | safety, and persistent expression of the transgene in the chest cavity cells of both models | [303,304] | |||
| rAAV6/2-hAAT | in vivo study, intratracheal delivery into C57/Bl6 mice, and in vitro study, cultures of human airway epithelial cells | lung cells’ transduction, even more efficient than rAAV5 both in vivo and in vitro | [301] | |||
| rAAV8-hAAT | in vivo study, intratracheal delivery into C57/Bl6 mice | lung cells’ transduction superior to that of rAAV5, as it resulted in 6-fold and 2.5-fold higher AAT levels in serum and broncho-alveolar fluid, respectively. | [302] | |||
| rAAV2/8-hAAT | in vivo study, intravenous delivery into C57/Bl6 mice | high transduction rate of hepatocytes | [298,372] | |||
| rAAV1-AAT | in vivo study, intramuscular delivery into C57/Bl6 mice and rabbits | dose-dependent inflammatory infiltrates at injection sites not affecting expression of transgene, along with dose-dependent detection of vector DNA in most animals, both at injection sites and in distal organs | [46,305] | |||
| rAAV2/9-AAT | in vivo study, intratracheal delivery into C57/Bl6 mice | efficient, relatively stable transduction of alveolar and nasal epithelia, that was not affected by high levels of neutralizing antibodies upon following re-administration | [175] | |||
| rAAV6-hAAT | in vivo study, nasal delivery into C57/Bl6 mice, and intratracheal delivery into canine model | therapeutic hAAT concentrations in both studied animal models, with levels being higher in lungs than serum, accompanied with an immune response against the vector capsid in some animals despite being immunosuppressed | [120] | |||
| Allergic asthma | rAAV-IL-4 | in vivo study, intratracheal delivery into Balb/cByJ mice | significant inhibition of airway eosinophilia and mucus production along with a reduction in airway hyper-responsiveness and asthma-associated cytokine levels | [306] | ||
| SPB | rAAV6/2-SFTPB | in vivo study, intratracheal delivery into murine model | efficient transduction of airway and alveolar epithelium, good tolerance of administered vector with no adverse effects, rapid and long-term restoration of the deficient SPB protein, along with an improvement of lung function, leading, subsequently, to an extended survival | [307] | ||
| Muscles | Degenerative muscle disorders | AAV1-FS344 (follistatin gene) | in vivo study, NHP model | safety and good tolerance of administered vector, promotion of sustained improvement in muscle size and strength | [311] | |
| DMD | rAAV6-micro-dystrophin | in vivo study, dystrophin/utrophin double-knockout murine model | no serious adverse events, improvement of muscle function along with prolongation of lifespan, explained by sustained restoration of deficient protein (dystrophin) in the respiratory, cardiac, and limb muscles | [312] | ||
| rAAV1-mini-dystrophin | in vivo study, dystrophin/utrophin double knockout murine model | prolongation of lifespan, highly efficient expression of transgene, improvement in muscle histopathology and function, reflected by improved growth and motility, along with prevention of spine and extremities’ deformation | [313] | |||
| AAV2.5-mini-dystrophin | Randomized, double-blind, placebo-controlled phase I clinical trial | safety and good tolerance of the vector, and detection of transgene DNA in all patients following intramuscular administration | [320] | |||
| AAV9-micro-dystrophin | phase I/II open-label, randomized, controlled clinical trial | safety and successful expression of transgene | [319] | |||
| AAV-rh74-micro-dystrophin | phase I/II, open-label, non-randomized clinical trial | safety and successful expression of transgene, along with improvement of motor function | ||||
| LGMD | rAAV8-hδ-SG (human δ-sarcoglycan gene) | in vivo study, murine model | prolongation of lifespan, efficient and sustained transduction of cardiac and skeletal muscles along with improvement in their histopathology and function | [314] | ||
| rAAV1-hα-SG | in vivo study, murine model | efficient and sustained expression of the transgene, histological and functional improvement of musculature reflected by correction of contractile force deficits and stretch sensibility, along with increase of animal activity | [316] | |||
| rAAV-hγ-SG | in vivo study, murine model | significantly efficient transduction of muscle fibers, and general histopathological improvement, that were achieved only upon early intervention | [317] | |||
| Cardiovascular system | HF | AAV6-EGFP | in vivo study, ovine model | global expression of the transgene, that was cardiac-tropic and substantially more robust and targeted, compared to that of intramuscular or intracoronary injection. | [128] | |
| AAV1-SERCA2a | randomized phase I/II clinical trial | highest adverse effects, including death, in the placebo group, and lowest in the high-dose group, with evidence of long-term transgene expression. However, in the low-dose and mid-dose groups, adverse effects were found to be high but delayed. | [323] | |||
| Hemophilia A | rAAV6-cFVIII and rAAV8-cFVIII | in vivo study, murine and canine models | restoration of physiologic levels of the deficient factor in the canine model for three years following intravenous administration, without any toxicity or immune reactions. A similar effect in the murine model was found, however, with detection of neutralizing antibodies against cFVIII in the mice sera | [322] | ||
| Hemophilia B | rAAV8-FIX and rAAV9-FIX | in vivo study, murine model | significant increase in transgene expression and therefore in IX factor levels, with decreased proinflammatory risk following intravenous administration | [168] | ||
| Liver | WD | AAV8-mini ATP7B | in vivo study, murine model | restoration of copper homeostasis, with 20% hepatocyte transduction being sufficient for correction | [327] | |
| CNs | AAV8- UGT1A1 | ongoing clinical trial | - | [329] | ||
| in vivo study, murine model | stable correction was achieved when the vector was administered at the 28th postnatal day, coupled with a rapamycin-based immunosuppression regimen delivered intraperitoneally | [330] | ||||
| Endocrine system | T1DM | dsAAV8-mIP | in vivo study, murine model | prevention of hyperglycemia in a dose-dependent manner | [336] | |
| rAAV2-IL-10 | in vivo study, murine model | a positive effect, decreasing autoimmunity, and thereby hyperglycemia | [337] | |||
| AAV8-PDL1 | in vivo study, murine model | pancreatic β cells’ protection | [338] | |||
| AAV8-anti-VEGF fab | ongoing clinical trial for diabetic retinopathy patients | - | [339] | |||
| APS1 | AAV9-AIRE | in vivo study, murine model | high transduction efficiency, along with restoration of AIRE expression in the thymus, following intra-thymic administration, and a subsequent significant reduction of serum auto-antibodies, with relatively normal tissue morphology showing no lymphocytic infiltrations. | [341] | ||
| Cancer | laryngeal cancer cell line (HEp-2) | AAVtk/GCV system | in vitro and in vivo study, murine model | significant tumor-suppressor efficacy in human head and neck cancer xenografts, and markedly prolonged survival | [344,345,346] | |
| bile duct cancer | AAVtk/GCV system | in vivo study, murine model | increased anti-tumor effect upon simultaneous administration with 5-fluorouracil, and better survival | [347] | ||
| bladder carcinoma | AAVtk/GCV system | in vivo study, murine model | control of tumor cell growth, a strong anti-tumor efficacy | [348] | ||
| prostate cancer | AAV2-ARHP8 | in vivo study, murine model | strong androgen receptor (AR) gene silencing following intravenous administration | [350] | ||
| hepatocellular carcinoma | AAV8-miR-26a | in vivo study, murine model | Induction of tumor-specific apoptosis, resulting in a significant inhibition of cancer cell proliferation without toxicity | [351] | ||
| breast carcinoma | AAV2-VEGF Trap | in vivo study, murine model | significant suppression of tumor growth and prevention of spontaneous pulmonary metastases | [352] | ||
| colorectal peritoneal carcinomatosis | AAV2-hPEDF | in vivo study, murine model | significant tumor suppression, inhibition of metastases, and prolonged survival time | [353] | ||
| hepatocellular carcinoma | AAV-kallistatin | in vivo study, murine model | induced potent cancer cell apoptosis, and prolonged survival time | [354] | ||
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ozelo, M.C.; Mahlangu, J.; Pasi, K.J.; Giermasz, A.; Leavitt, A.D.; Laffan, M.; Symington, E.; Quon, D.V.; Wang, J.D.; Peerlinck, K.; et al. Valoctocogene Roxaparvovec Gene Therapy for Hemophilia A. N. Engl. J. Med. 2022, 386, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Zolgensma—One-time gene therapy for spinal muscular atrophy. Med. Lett. Drugs Ther. 2019, 61, 113–114.
- Prado, D.A.; Acosta-Acero, M.; Maldonado, R.S. Gene therapy beyond luxturna: A new horizon of the treatment for inherited retinal disease. Curr. Opin. Ophthalmol. 2020, 31, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Keeler, A.M.; Flotte, T.R. Recombinant Adeno-Associated Virus Gene Therapy in Light of Luxturna (and Zolgensma and Glybera): Where Are We, and How Did We Get Here? Annu. Rev. Virol. 2019, 6, 601–621. [Google Scholar] [CrossRef]
- Atchison, R.W.; Casto, B.C.; Hammon, W.M. Adenovirus-associated defective virus particles. Science 1965, 149, 754–756. [Google Scholar] [CrossRef]
- Mezzina, M.; Merten, O.W. Adeno-associated viruses. Methods Mol. Biol. 2011, 737, 211–234. [Google Scholar] [CrossRef]
- Hoggan, M.D.; Blacklow, N.R.; Rowe, W.P. Studies of small DNA viruses found in various adenovirus preparations: Physical, biological, and immunological characteristics. Proc. Natl. Acad. Sci. USA 1966, 55, 1467–1474. [Google Scholar] [CrossRef] [Green Version]
- Berns, K.I.; Giraud, C. Biology of adeno-associated virus. Curr. Top. Microbiol. Immunol. 1996, 218, 1–23. [Google Scholar] [CrossRef]
- Yalkinoglu, A.O.; Heilbronn, R.; Bürkle, A.; Schlehofer, J.R.; zur Hausen, H. DNA amplification of adeno-associated virus as a response to cellular genotoxic stress. Cancer Res. 1988, 48, 3123–3129. [Google Scholar]
- Gonçalves, M.A.F.V. Adeno-associated virus: From defective virus to effective vector. Virol. J. 2005, 2, 43. [Google Scholar] [CrossRef] [Green Version]
- Erles, K.; Sebökovà, P.; Schlehofer, J.R. Update on the prevalence of serum antibodies (IgG and IgM) to adeno-associated virus (AAV). J. Med. Virol. 1999, 59, 406–411. [Google Scholar] [CrossRef]
- Büning, H.; Perabo, L.; Coutelle, O.; Quadt-Humme, S.; Hallek, M. Recent developments in adeno-associated virus vector technology. J. Gene Med. 2008, 10, 717–733. [Google Scholar] [CrossRef]
- Tijssen, P. Handbook of Parvoviruses Vol. I and Vol. II Continued; CRC Press, Inc.: Boca Raton, FL, USA, 1990. [Google Scholar]
- Grimes, T.; King, D.; Fletcher, O. Serologic and pathogenicity studies of avian adenovirus isolated from chickens with inclusion body hepatitis. Avian Dis. 1978, 22, 177–180. [Google Scholar] [CrossRef]
- Alvarado, I.; Villegas, P.; El-Attrache, J.; Jensen, E.; Rosales, G.; Perozo, F.; Purvis, L.J. Genetic characterization, pathogenicity, and protection studies with an avian adenovirus isolate associated with inclusion body hepatitis. Avian Dis. 2007, 51, 27–32. [Google Scholar] [CrossRef]
- Bossis, I.; Chiorini, J.A. Cloning of an avian adeno-associated virus (AAAV) and generation of recombinant AAAV particles. J. Virol. 2003, 77, 6799–6810. [Google Scholar] [CrossRef] [Green Version]
- Arbetman, A.E.; Lochrie, M.; Zhou, S.; Wellman, J.; Scallan, C.; Doroudchi, M.M.; Randlev, B.; Patarroyo-White, S.; Liu, T.; Smith, P. Novel caprine adeno-associated virus (AAV) capsid (AAV-Go. 1) is closely related to the primate AAV-5 and has unique tropism and neutralization properties. J. Virol. 2005, 79, 15238–15245. [Google Scholar] [CrossRef] [Green Version]
- Olson, E.J.; Haskell, S.R.; Frank, R.K.; Lehmkuhl, H.D.; Hobbs, L.A.; Warg, J.V.; Landgraf, J.G.; Wünschmann, A. Isolation of an adenovirus and an adeno-associated virus from goat kids with enteritis. J. Vet.-Diagn. Investig. 2004, 16, 461–464. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Katano, H.; Bossis, I.; Chiorini, J.A. Cloning and characterization of a bovine adeno-associated virus. J. Virol. 2004, 78, 6509–6516. [Google Scholar] [CrossRef] [Green Version]
- Coria, M.; Lehmkuhl, H. Isolation and identification of a bovine adenovirus type 3 with an adenovirus-associated virus. J. Vet.-Diagn. Investig. 1978, 39, 1904–1906. [Google Scholar]
- Dutta, S.K. Isolation and characterization of an adenovirus and isolation of its adenovirus-associated virus in cell culture from foals with respiratory tract disease. Am. J. Vet. Res. 1975, 36, 247–250. [Google Scholar]
- Qiu, J.; Pintel, D.J. Alternative polyadenylation of adeno-associated virus type 5 RNA within an internal intron is governed by the distance between the promoter and the intron and is inhibited by U1 small nuclear RNP binding to the intervening donor. J. Biol. Chem. 2004, 279, 14889–14898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merten, O.W.; Gény-Fiamma, C.; Douar, A.M. Current issues in adeno-associated viral vector production. Gene Ther. 2005, 12 (Suppl. S1), S51–S61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, M.; Lins, B.; Mietzsch, M.; Heilbronn, R.; Van Vliet, K.; Chipman, P.; Agbandje-McKenna, M.; Cleaver, B.D.; Clément, N.; Byrne, B.J.; et al. A simplified purification protocol for recombinant adeno-associated virus vectors. Mol. Ther. Methods Clin. Dev. 2014, 1, 14034. [Google Scholar] [CrossRef] [PubMed]
- Carter, P.J.; Samulski, R.J. Adeno-associated viral vectors as gene delivery vehicles. Int. J. Mol. Med. 2000, 6, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, M.P.; McCarty, D.M. Hairpin-end conformation of adeno-associated virus genome determines interactions with DNA-repair pathways. Gene Ther. 2013, 20, 686–693. [Google Scholar] [CrossRef] [Green Version]
- Michelfelder, S.; Trepel, M. Adeno-associated viral vectors and their redirection to cell-type specific receptors. Adv. Genet. 2009, 67, 29–60. [Google Scholar] [CrossRef]
- Dubielzig, R.; King, J.A.; Weger, S.; Kern, A.; Kleinschmidt, J.A. Adeno-associated virus type 2 protein interactions: Formation of pre-encapsidation complexes. J. Virol. 1999, 73, 8989–8998. [Google Scholar] [CrossRef] [Green Version]
- Kyöstiö, S.R.; Owens, R.A.; Weitzman, M.D.; Antoni, B.A.; Chejanovsky, N.; Carter, B.J. Analysis of adeno-associated virus (AAV) wild-type and mutant Rep proteins for their abilities to negatively regulate AAV p5 and p19 mRNA levels. J. Virol. 1994, 68, 2947–2957. [Google Scholar] [CrossRef] [Green Version]
- Galibert, L.; Hyvönen, A.; Eriksson, R.A.E.; Mattola, S.; Aho, V.; Salminen, S.; Albers, J.D.; Peltola, S.K.; Weman, S.; Nieminen, T.; et al. Functional roles of the membrane-associated AAV protein MAAP. Sci. Rep. 2021, 11, 21698. [Google Scholar] [CrossRef]
- Kronenberg, S.; Kleinschmidt, J.A.; Böttcher, B. Electron cryo-microscopy and image reconstruction of adeno-associated virus type 2 empty capsids. EMBO Rep. 2001, 2, 997–1002. [Google Scholar] [CrossRef] [Green Version]
- Rabinowitz, J.E.; Samulski, R.J. Building a better vector: The manipulation of AAV virions. Virology 2000, 278, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Asokan, A.; Samulski, R.J. Adeno-associated virus serotypes: Vector toolkit for human gene therapy. Mol. Ther. J. Am. Soc. Gene Ther. 2006, 14, 316–327. [Google Scholar] [CrossRef]
- Mori, S.; Wang, L.; Takeuchi, T.; Kanda, T. Two novel adeno-associated viruses from cynomolgus monkey: Pseudotyping characterization of capsid protein. Virology 2004, 330, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Gao, G.P.; Alvira, M.R.; Wang, L.; Calcedo, R.; Johnston, J.; Wilson, J.M. Novel adeno-associated viruses from rhesus monkeys as vectors for human gene therapy. Proc. Natl. Acad. Sci. USA 2002, 99, 11854–11859. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Miller, E.; Agbandje-McKenna, M.; Samulski, R.J. Alpha2,3 and alpha2,6 N-linked sialic acids facilitate efficient binding and transduction by adeno-associated virus types 1 and 6. J. Virol. 2006, 80, 9093–9103. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Samulski, R.J. Engineering adeno-associated virus vectors for gene therapy. Nat. Rev. Genet. 2020, 21, 255–272. [Google Scholar] [CrossRef]
- Rabinowitz, J.E.; Rolling, F.; Li, C.; Conrath, H.; Xiao, W.; Xiao, X.; Samulski, R.J. Cross-packaging of a single adeno-associated virus (AAV) type 2 vector genome into multiple AAV serotypes enables transduction with broad specificity. J. Virol. 2002, 76, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Zolotukhin, S.; Potter, M.; Zolotukhin, I.; Sakai, Y.; Loiler, S.; Fraites, T.J., Jr.; Chiodo, V.A.; Phillipsberg, T.; Muzyczka, N.; Hauswirth, W.W.; et al. Production and purification of serotype 1, 2, and 5 recombinant adeno-associated viral vectors. Methods 2002, 28, 158–167. [Google Scholar] [CrossRef]
- Robert, M.A.; Chahal, P.S.; Audy, A.; Kamen, A.; Gilbert, R.; Gaillet, B. Manufacturing of recombinant adeno-associated viruses using mammalian expression platforms. Biotechnol. J. 2017, 12, 1600193. [Google Scholar] [CrossRef]
- Mary, B.; Maurya, S.; Arumugam, S.; Kumar, V.; Jayandharan, G.R. Post-translational modifications in capsid proteins of recombinant adeno-associated virus (AAV) 1-rh10 serotypes. FEBS J. 2019, 286, 4964–4981. [Google Scholar] [CrossRef]
- Huang, L.-Y.; Patel, A.; Ng, R.; Miller, E.B.; Halder, S.; McKenna, R.; Asokan, A.; Agbandje-McKenna, M.; Banks, L. Characterization of the Adeno-Associated Virus 1 and 6 Sialic Acid Binding Site. J. Virol. 2016, 90, 5219–5230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, H.; Liu, Y.; Rabinowitz, J.; Li, C.; Samulski, R.J.; Walsh, C.E. Several log increase in therapeutic transgene delivery by distinct adeno-associated viral serotype vectors. Mol. Ther. J. Am. Soc. Gene Ther. 2000, 2, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Hauck, B.; Xiao, W. Characterization of tissue tropism determinants of adeno-associated virus type 1. J. Virol. 2003, 77, 2768–2774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisowski, L.; Tay, S.S.; Alexander, I.E. Adeno-associated virus serotypes for gene therapeutics. Curr. Opin. Pharmacol. 2015, 24, 59–67. [Google Scholar] [CrossRef]
- Lu, Y.; Choi, Y.K.; Campbell-Thompson, M.; Li, C.; Tang, Q.; Crawford, J.M.; Flotte, T.R.; Song, S. Therapeutic level of functional human alpha 1 antitrypsin (hAAT) secreted from murine muscle transduced by adeno-associated virus (rAAV1) vector. J. Gene Med. 2006, 8, 730–735. [Google Scholar] [CrossRef]
- Wang, C.; Wang, C.M.; Clark, K.R.; Sferra, T.J. Recombinant AAV serotype 1 transduction efficiency and tropism in the murine brain. Gene Ther. 2003, 10, 1528–1534. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Yeghiazarians, Y.; Lee, A.; Huang, Y.; Arakawa-Hoyt, J.; Ye, J.; Orcino, G.; Grossman, W.; Kan, Y.W. AAV serotype 1 mediates more efficient gene transfer to pig myocardium than AAV serotype 2 and plasmid. J. Gene Med. 2008, 10, 33–41. [Google Scholar] [CrossRef]
- Palomeque, J.; Chemaly, E.R.; Colosi, P.; Wellman, J.A.; Zhou, S.; Del Monte, F.; Hajjar, R.J. Efficiency of eight different AAV serotypes in transducing rat myocardium in vivo. Gene Ther. 2007, 14, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Kapturczak, M.; Loiler, S.A.; Zolotukhin, S.; Glushakova, O.Y.; Madsen, K.M.; Samulski, R.J.; Hauswirth, W.W.; Campbell-Thompson, M.; Berns, K.I.; et al. Efficient transduction of vascular endothelial cells with recombinant adeno-associated virus serotype 1 and 5 vectors. Hum. Gene Ther. 2005, 16, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Auricchio, A.; Kobinger, G.; Anand, V.; Hildinger, M.; O’Connor, E.; Maguire, A.M.; Wilson, J.M.; Bennett, J. Exchange of surface proteins impacts on viral vector cellular specificity and transduction characteristics: The retina as a model. Hum. Mol. Genet. 2001, 10, 3075–3081. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A. In vivo tissue-tropism of adeno-associated viral vectors. Curr. Opin. Virol. 2016, 21, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Summerford, C.; Samulski, R.J. Membrane-Associated Heparan Sulfate Proteoglycan Is a Receptor for Adeno-Associated Virus Type 2 Virions. J. Virol. 1998, 72, 1438–1445. [Google Scholar] [CrossRef] [Green Version]
- Opie, S.R.; Warrington, K.H., Jr.; Agbandje-McKenna, M.; Zolotukhin, S.; Muzyczka, N. Identification of amino acid residues in the capsid proteins of adeno-associated virus type 2 that contribute to heparan sulfate proteoglycan binding. J. Virol. 2003, 77, 6995–7006. [Google Scholar] [CrossRef] [Green Version]
- Auricchio, A.; Sena-Esteves, M.; Gao, G. Purification of Recombinant Adeno-Associated Virus 2 (rAAV2) by Heparin Column Affinity Chromatography. Cold Spring Harb. Protoc. 2020, 2020, 095620. [Google Scholar] [CrossRef]
- Qing, K.; Mah, C.; Hansen, J.; Zhou, S.; Dwarki, V.; Srivastava, A.J. Human fibroblast growth factor receptor 1 is a co-receptor for infection by adeno-associated virus 2. Nat. Med. 1999, 5, 71–77. [Google Scholar] [CrossRef]
- Summerford, C.; Bartlett, J.S.; Samulski, R.J.J. αVβ5 integrin: A co-receptor for adeno-associated virus type 2 infection. Nat. Med. 1999, 5, 78–82. [Google Scholar] [CrossRef]
- Asokan, A.; Hamra, J.B.; Govindasamy, L.; Agbandje-McKenna, M.; Samulski, R.J.J. Adeno-associated virus type 2 contains an integrin α5β1 binding domain essential for viral cell entry. J. Virol. 2006, 80, 8961–8969. [Google Scholar] [CrossRef] [Green Version]
- Kashiwakura, Y.; Tamayose, K.; Iwabuchi, K.; Hirai, Y.; Shimada, T.; Matsumoto, K.; Nakamura, T.; Watanabe, M.; Oshimi, K.; Daida, H.J. Hepatocyte growth factor receptor is a coreceptor for adeno-associated virus type 2 infection. J. Virol. 2005, 79, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Akache, B.; Grimm, D.; Pandey, K.; Yant, S.R.; Xu, H.; Kay, M.A.J. The 37/67-kilodalton laminin receptor is a receptor for adeno-associated virus serotypes 8, 2, 3, and 9. J. Virol. 2006, 80, 9831–9836. [Google Scholar] [CrossRef] [Green Version]
- Kurzeder, C.; Koppold, B.; Sauer, G.; Pabst, S.; Kreienberg, R.; Deissler, H.J. CD9 promotes adeno-associated virus type 2 infection of mammary carcinoma cells with low cell surface expression of heparan sulphate proteoglycans. Int. J. Mol. Med. 2007, 19, 325–333. [Google Scholar] [CrossRef]
- Takeda, S.-i.; Takahashi, M.; Mizukami, H.; Kobayashi, E.; Takeuchi, K.; Hakamata, Y.; Kaneko, T.; Yamamoto, H.; Ito, C.; Ozawa, K.J. Successful gene transfer using adeno-associated virus vectors into the kidney: Comparison among adeno-associated virus serotype 1–5 vectors in vitro and in vivo. Nephron Exp. Nephrol. 2004, 96, e119–e126. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.F.; Li, Q.H.; Shenoy, V.; Zingler, M.; Jun, J.Y.; Verma, A.; Katovich, M.J.; Raizada, M.K. Comparison of the transduction efficiency of tyrosine-mutant adeno-associated virus serotype vectors in kidney. Clin. Exp. Pharmacol. Physiol. 2013, 40, 53–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddad, G.; Kölling, M.; Wegmann, U.A.; Dettling, A.; Seeger, H.; Schmitt, R.; Soerensen-Zender, I.; Haller, H.; Kistler, A.D.; Dueck, A.J. Renal AAV2-mediated overexpression of long non-coding RNA H19 attenuates ischemic acute kidney injury through sponging of microRNA-30a-5p. J. Am. Soc. Nephrol. 2021, 32, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Logan, G.J.; Dane, A.P.; Hallwirth, C.V.; Smyth, C.M.; Wilkie, E.E.; Amaya, A.K.; Zhu, E.; Khandekar, N.; Ginn, S.L.; Liao, S.H. Identification of liver-specific enhancer–promoter activity in the 3′ untranslated region of the wild-type AAV2 genome. Nat. Genet. 2017, 49, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, G.P.; Herzog, R.W.; Mount, J.; Arruda, V.R.; Tillson, D.M.; Hathcock, J.; van Ginkel, F.W.; High, K.A.; Lothrop, C.D., Jr. The Journal of the American Society of Hematology. Long-term correction of inhibitor-prone hemophilia B dogs treated with liver-directed AAV2-mediated factor IX gene therapy. Blood J. Am. Soc. Hematol. 2009, 113, 797–806. [Google Scholar]
- Petrs-Silva, H.; Dinculescu, A.; Li, Q.; Deng, W.-T.; Pang, J.-j.; Min, S.-H.; Chiodo, V.; Neeley, A.W.; Govindasamy, L.; Bennett, A.; et al. Novel Properties of Tyrosine-mutant AAV2 Vectors in the Mouse Retina. Mol. Ther. 2011, 19, 293–301. [Google Scholar] [CrossRef]
- Yin, L.; Greenberg, K.; Hunter, J.J.; Dalkara, D.; Kolstad, K.D.; Masella, B.D.; Wolfe, R.; Visel, M.; Stone, D.; Libby, R.T.; et al. Intravitreal injection of AAV2 transduces macaque inner retina. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2775–2783. [Google Scholar] [CrossRef]
- Wassmer, S.J.; Carvalho, L.S.; György, B.; Vandenberghe, L.H.; Maguire, C.A. Exosome-associated AAV2 vector mediates robust gene delivery into the murine retina upon intravitreal injection. J. Sci. Rep. 2017, 7, 45329. [Google Scholar] [CrossRef] [Green Version]
- Griffey, M.; Bible, E.; Vogler, C.; Levy, B.; Gupta, P.; Cooper, J.; Sands, M.S. Adeno-associated virus 2-mediated gene therapy decreases autofluorescent storage material and increases brain mass in a murine model of infantile neuronal ceroid lipofuscinosis. Neurobiol. Dis. 2004, 16, 360–369. [Google Scholar] [CrossRef]
- Griffey, M.A.; Wozniak, D.; Wong, M.; Bible, E.; Johnson, K.; Rothman, S.M.; Wentz, A.E.; Cooper, J.D.; Sands, M.S. CNS-directed AAV2-mediated gene therapy ameliorates functional deficits in a murine model of infantile neuronal ceroid lipofuscinosis. Mol. Ther. 2006, 13, 538–547. [Google Scholar] [CrossRef]
- Pankajakshan, D.; Makinde, T.O.; Gaurav, R.; Del Core, M.; Hatzoudis, G.; Pipinos, I.; Agrawal, D.K. Successful Transfection of Genes Using AAV-2/9 Vector in Swine Coronary and Peripheral Arteries. J. Surg. Res. 2012, 175, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Rabinowitz, J.E.; Bowles, D.E.; Faust, S.M.; Ledford, J.G.; Cunningham, S.E.; Samulski, R.J. Cross-Dressing the Virion: The Transcapsidation of Adeno-Associated Virus Serotypes Functionally Defines Subgroups. J. Virol. 2004, 78, 4421–4432. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, S.D.; Steadman, R.A.; Johnson, F.B. Attachment of adeno-associated virus type 3H to fibroblast growth factor receptor 1. Arch. Virol. 2006, 151, 617–623. [Google Scholar] [CrossRef]
- Ling, C.; Wang, Y.; Zhang, Y.; Ejjigani, A.; Yin, Z.; Lu, Y.; Wang, L.; Wang, M.; Li, J.; Hu, Z.; et al. Selective in vivo targeting of human liver tumors by optimized AAV3 vectors in a murine xenograft model. Hum. Gene Ther. 2014, 25, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Ling, C.; Yin, Z.; Li, J.; Zhang, D.; Aslanidi, G.; Srivastava, A. Strategies to generate high-titer, high-potency recombinant AAV3 serotype vectors. Mol. Ther. Methods Clin. Dev. 2016, 3, 16029. [Google Scholar] [CrossRef]
- Handa, A.; Muramatsu, S.I.; Qiu, J.; Mizukami, H.; Brown, K.E. Adeno-associated virus (AAV)-3-based vectors transduce haematopoietic cells not susceptible to transduction with AAV-2-based vectors. J. Gen. Virol. 2000, 81, 2077–2084. [Google Scholar] [CrossRef]
- Cheng, B.; Ling, C.; Dai, Y.; Lu, Y.; Glushakova, L.G.; Gee, S.W.Y.; McGoogan, K.E.; Aslanidi, G.V.; Park, M.; Stacpoole, P.W.; et al. Development of optimized AAV3 serotype vectors: Mechanism of high-efficiency transduction of human liver cancer cells. Gene Ther. 2012, 19, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Muramatsu, S.; Mizukami, H.; Young, N.S.; Brown, K.E. Nucleotide sequencing and generation of an infectious clone of adeno-associated virus 3. Virology 1996, 221, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Ling, C.; Zhong, L.; Li, M.; Su, Q.; He, R.; Tang, Q.; Greiner, D.L.; Shultz, L.D.; Brehm, M.A.; et al. Efficient and Targeted Transduction of Nonhuman Primate Liver With Systemically Delivered Optimized AAV3B Vectors. Mol. Ther. 2015, 23, 1867–1876. [Google Scholar] [CrossRef] [Green Version]
- Vercauteren, K.; Hoffman, B.E.; Zolotukhin, I.; Keeler, G.D.; Xiao, J.W.; Basner-Tschakarjan, E.; High, K.A.; Ertl, H.C.J.; Rice, C.M.; Srivastava, A.; et al. Superior In vivo Transduction of Human Hepatocytes Using Engineered AAV3 Capsid. Mol. Ther. 2016, 24, 1042–1049. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Okada, T.; Sheykholeslami, K.; Shimazaki, K.; Nomoto, T.; Muramatsu, S.; Kanazawa, T.; Takeuchi, K.; Ajalli, R.; Mizukami, H.; et al. Specific and efficient transduction of Cochlear inner hair cells with recombinant adeno-associated virus type 3 vector. Mol. Ther. J. Am. Soc. Gene Ther. 2005, 12, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Padron, E.; Bowman, V.; Kaludov, N.; Govindasamy, L.; Levy, H.; Nick, P.; McKenna, R.; Muzyczka, N.; Chiorini, J.A.; Baker, T.S.; et al. Structure of adeno-associated virus type 4. J. Virol. 2005, 79, 5047–5058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, W.P.; Boucher, D.W.; Melnick, J.L.; Taber, L.H.; Yow, M.D. Seroepidemiological and ecological studies of the adenovirus-associated satellite viruses. Infect. Immun. 1970, 2, 716–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blacklow, N.R.; Hoggan, M.D.; Rowe, W.P. Serologic evidence for human infection with adenovirus-associated viruses. J. Natl. Cancer Inst. 1968, 40, 319–327. [Google Scholar] [PubMed]
- Dreizin, R.S.; Zhuravel, T.F.; Tarasova, A.B.; Sobolev, S.G.; Kozlov, V.G. Experimental infection of green monkeys with adenoassociated virus. Vopr. Virusol. 1981, 1, 82–89. [Google Scholar]
- Chiorini, J.A.; Yang, L.; Liu, Y.; Safer, B.; Kotin, R.M. Cloning of adeno-associated virus type 4 (AAV4) and generation of recombinant AAV4 particles. J. Virol. 1997, 71, 6823–6833. [Google Scholar] [CrossRef] [Green Version]
- Kaludov, N.; Brown, K.E.; Walters, R.W.; Zabner, J.; Chiorini, J.A. Adeno-associated virus serotype 4 (AAV4) and AAV5 both require sialic acid binding for hemagglutination and efficient transduction but differ in sialic acid linkage specificity. J. Virol. 2001, 75, 6884–6893. [Google Scholar] [CrossRef] [Green Version]
- Kaludov, N.; Handelman, B.; Chiorini, J.A. Scalable purification of adeno-associated virus type 2, 4, or 5 using ion-exchange chromatography. Hum. Gene Ther. 2002, 13, 1235–1243. [Google Scholar] [CrossRef]
- Davidson, B.L.; Stein, C.S.; Heth, J.A.; Martins, I.; Kotin, R.M.; Derksen, T.A.; Zabner, J.; Ghodsi, A.; Chiorini, J.A. Recombinant adeno-associated virus type 2, 4, and 5 vectors: Transduction of variant cell types and regions in the mammalian central nervous system. Proc. Natl. Acad. Sci. USA 2000, 97, 3428–3432. [Google Scholar] [CrossRef]
- Weber, M.; Rabinowitz, J.; Provost, N.; Conrath, H.; Folliot, S.; Briot, D.; Chérel, Y.; Chenuaud, P.; Samulski, J.; Moullier, P.; et al. Recombinant adeno-associated virus serotype 4 mediates unique and exclusive long-term transduction of retinal pigmented epithelium in rat, dog, and nonhuman primate after subretinal delivery. Mol. Ther. J. Am. Soc. Gene Ther. 2003, 7, 774–781. [Google Scholar] [CrossRef]
- Zincarelli, C.; Soltys, S.; Rengo, G.; Rabinowitz, J.E. Analysis of AAV serotypes 1–9 mediated gene expression and tropism in mice after systemic injection. Mol. Ther. 2008, 16, 1073–1080. [Google Scholar] [CrossRef]
- Bantel-Schaal, U.; zur Hausen, H. Characterization of the DNA of a defective human parvovirus isolated from a genital site. Virology 1984, 134, 52–63. [Google Scholar] [CrossRef]
- Stutika, C.; Hüser, D.; Weger, S.; Rutz, N.; Heßler, M.; Heilbronn, R. Definition of herpes simplex virus helper functions for the replication of adeno-associated virus type 5. J. Gen. Virol. 2015, 96, 840–850. [Google Scholar] [CrossRef]
- Bantel-Schaal, U.; Delius, H.; Schmidt, R.; Hausen, H.Z. Human Adeno-Associated Virus Type 5 Is Only Distantly Related to Other Known Primate Helper-Dependent Parvoviruses. J. Virol. 1999, 73, 939–947. [Google Scholar] [CrossRef] [Green Version]
- Chiorini, J.A.; Kim, F.; Yang, L.; Kotin, R.M. Cloning and characterization of adeno-associated virus type 5. J. Virol. 1999, 73, 1309–1319. [Google Scholar] [CrossRef] [Green Version]
- Bantel-Schaal, U.; Hub, B.; Kartenbeck, J. Endocytosis of adeno-associated virus type 5 leads to accumulation of virus particles in the Golgi compartment. J. Virol. 2002, 76, 2340–2349. [Google Scholar] [CrossRef] [Green Version]
- Zabner, J.; Seiler, M.; Walters, R.; Kotin, R.M.; Fulgeras, W.; Davidson, B.L.; Chiorini, J.A. Adeno-associated virus type 5 (AAV5) but not AAV2 binds to the apical surfaces of airway epithelia and facilitates gene transfer. J. Virol. 2000, 74, 3852–3858. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D.; Kay, M.A. From virus evolution to vector revolution: Use of naturally occurring serotypes of adeno-associated virus (AAV) as novel vectors for human gene therapy. Curr. Gene Ther. 2003, 3, 281–304. [Google Scholar] [CrossRef]
- Walters, R.W.; Yi, S.M.; Keshavjee, S.; Brown, K.E.; Welsh, M.J.; Chiorini, J.A.; Zabner, J. Binding of adeno-associated virus type 5 to 2,3-linked sialic acid is required for gene transfer. J. Biol. Chem. 2001, 276, 20610–20616. [Google Scholar] [CrossRef] [Green Version]
- Henckaerts, E.; Linden, R.M. Adeno-associated virus: A key to the human genome? Future Virol. 2010, 5, 555–574. [Google Scholar] [CrossRef] [Green Version]
- Di Pasquale, G.; Davidson, B.L.; Stein, C.S.; Martins, I.; Scudiero, D.; Monks, A.; Chiorini, J.A. Identification of PDGFR as a receptor for AAV-5 transduction. Nat. Med. 2003, 9, 1306–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonnenmacher, M.; Weber, T. Intracellular transport of recombinant adeno-associated virus vectors. Gene Ther. 2012, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Auricchio, A.; O’Connor, E.; Hildinger, M.; Wilson, J.M. A Single-Step Affinity Column for Purification of Serotype-5 Based Adeno-associated Viral Vectors. Mol. Ther. 2001, 4, 372–374. [Google Scholar] [CrossRef] [PubMed]
- Walters, R.W.; Pilewski, J.M.; Chiorini, J.A.; Zabner, J. Secreted and Transmembrane Mucins Inhibit Gene Transfer with AAV4 More Efficiently than AAV5*. J. Biol. Chem. 2002, 277, 23709–23713. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.S.; Schmidt, M.; Yan, Z.; Lindbloom, J.D.; Harding, T.C.; Donahue, B.A.; Engelhardt, J.F.; Kotin, R.; Davidson, B.L. Virus-mediated transduction of murine retina with adeno-associated virus: Effects of viral capsid and genome size. J. Virol. 2002, 76, 7651–7660. [Google Scholar] [CrossRef] [Green Version]
- Alisky, J.M.; Hughes, S.M.; Sauter, S.L.; Jolly, D.; Dubensky, T.W., Jr.; Staber, P.D.; Chiorini, J.A.; Davidson, B.L. Transduction of murine cerebellar neurons with recombinant FIV and AAV5 vectors. Neuroreport 2000, 11, 2669–2673. [Google Scholar] [CrossRef]
- Burger, C.; Gorbatyuk, O.S.; Velardo, M.J.; Peden, C.S.; Williams, P.; Zolotukhin, S.; Reier, P.J.; Mandel, R.J.; Muzyczka, N. Recombinant AAV viral vectors pseudotyped with viral capsids from serotypes 1, 2, and 5 display differential efficiency and cell tropism after delivery to different regions of the central nervous system. Mol. Ther. J. Am. Soc. Gene Ther. 2004, 10, 302–317. [Google Scholar] [CrossRef]
- Ding, W.; Yan, Z.; Zak, R.; Saavedra, M.; Rodman, D.M.; Engelhardt, J.F. Second-strand genome conversion of adeno-associated virus type 2 (AAV-2) and AAV-5 is not rate limiting following apical infection of polarized human airway epithelia. J. Virol. 2003, 77, 7361–7366. [Google Scholar] [CrossRef] [Green Version]
- Auricchio, A.; O’Connor, E.; Weiner, D.; Gao, G.P.; Hildinger, M.; Wang, L.; Calcedo, R.; Wilson, J.M. Noninvasive gene transfer to the lung for systemic delivery of therapeutic proteins. J. Clin. Investig. 2002, 110, 499–504. [Google Scholar] [CrossRef]
- Seiler, M.P.; Miller, A.D.; Zabner, J.; Halbert, C.L. Adeno-associated virus types 5 and 6 use distinct receptors for cell entry. Hum. Gene Ther. 2006, 17, 10–19. [Google Scholar] [CrossRef]
- Pañeda, A.; Vanrell, L.; Mauleon, I.; Crettaz, J.S.; Berraondo, P.; Timmermans, E.J.; Beattie, S.G.; Twisk, J.; van Deventer, S.; Prieto, J.; et al. Effect of adeno-associated virus serotype and genomic structure on liver transduction and biodistribution in mice of both genders. Hum. Gene Ther. 2009, 20, 908–917. [Google Scholar] [CrossRef]
- Xiao, W.; Chirmule, N.; Berta, S.C.; McCullough, B.; Gao, G.; Wilson, J.M. Gene Therapy Vectors Based on Adeno-Associated Virus Type 1. J. Virol. 1999, 73, 3994–4003. [Google Scholar] [CrossRef] [Green Version]
- Rutledge, E.A.; Halbert, C.L.; Russell, D.W. Infectious Clones and Vectors Derived from Adeno-Associated Virus (AAV) Serotypes Other Than AAV Type 2. J. Virol. 1998, 72, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Korneyenkov, M.A.; Zamyatnin, A.A., Jr. Next Step in Gene Delivery: Modern Approaches and Further Perspectives of AAV Tropism Modification. Pharmaceutics 2021, 13, 750. [Google Scholar] [CrossRef]
- Ng, R.; Govindasamy, L.; Gurda, B.L.; McKenna, R.; Kozyreva, O.G.; Samulski, R.J.; Parent, K.N.; Baker, T.S.; Agbandje-McKenna, M. Structural characterization of the dual glycan binding adeno-associated virus serotype 6. J. Virol. 2010, 84, 12945–12957. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Asokan, A.; Grieger, J.C.; Govindasamy, L.; Agbandje-McKenna, M.; Samulski, R.J. Single amino acid changes can influence titer, heparin binding, and tissue tropism in different adeno-associated virus serotypes. J. Virol. 2006, 80, 11393–11397. [Google Scholar] [CrossRef] [Green Version]
- Weller, M.L.; Amornphimoltham, P.; Schmidt, M.; Wilson, P.A.; Gutkind, J.S.; Chiorini, J.A. Epidermal growth factor receptor is a co-receptor for adeno-associated virus serotype 6. Nat. Med. 2010, 16, 662–664. [Google Scholar] [CrossRef] [Green Version]
- Halbert, C.L.; Allen, J.M.; Miller, A.D. Adeno-associated virus type 6 (AAV6) vectors mediate efficient transduction of airway epithelial cells in mouse lungs compared to that of AAV2 vectors. J. Virol. 2001, 75, 6615–6624. [Google Scholar] [CrossRef] [Green Version]
- Halbert, C.L.; Madtes, D.K.; Vaughan, A.E.; Wang, Z.; Storb, R.; Tapscott, S.J.; Miller, A.D. Expression of human alpha1-antitrypsin in mice and dogs following AAV6 vector-mediated gene transfer to the lungs. Mol. Ther. J. Am. Soc. Gene Ther. 2010, 18, 1165–1172. [Google Scholar] [CrossRef]
- Grimm, D.; Zhou, S.; Nakai, H.; Thomas, C.E.; Storm, T.A.; Fuess, S.; Matsushita, T.; Allen, J.; Surosky, R.; Lochrie, M.; et al. Preclinical in vivo evaluation of pseudotyped adeno-associated virus vectors for liver gene therapy. Blood 2003, 102, 2412–2419. [Google Scholar] [CrossRef]
- Blankinship, M.J.; Gregorevic, P.; Allen, J.M.; Harper, S.Q.; Harper, H.; Halbert, C.L.; Miller, A.D.; Chamberlain, J.S. Efficient transduction of skeletal muscle using vectors based on adeno-associated virus serotype 6. Mol. Ther. J. Am. Soc. Gene Ther. 2004, 10, 671–678. [Google Scholar] [CrossRef]
- Wang, Z.; Kuhr, C.S.; Allen, J.M.; Blankinship, M.; Gregorevic, P.; Chamberlain, J.S.; Tapscott, S.J.; Storb, R. Sustained AAV-mediated dystrophin expression in a canine model of Duchenne muscular dystrophy with a brief course of immunosuppression. Mol. Ther. J. Am. Soc. Gene Ther. 2007, 15, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Qiao, C.; Zhang, W.; Yuan, Z.; Shin, J.H.; Li, J.; Jayandharan, G.R.; Zhong, L.; Srivastava, A.; Xiao, X.; Duan, D. Adeno-associated virus serotype 6 capsid tyrosine-to-phenylalanine mutations improve gene transfer to skeletal muscle. Hum. Gene Ther. 2010, 21, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Zincarelli, C.; Soltys, S.; Rengo, G.; Koch, W.J.; Rabinowitz, J.E. Comparative cardiac gene delivery of adeno-associated virus serotypes 1–9 reveals that AAV6 mediates the most efficient transduction in mouse heart. Clin. Transl. Sci. 2010, 3, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Raake, P.W.; Hinkel, R.; Müller, S.; Delker, S.; Kreuzpointner, R.; Kupatt, C.; Katus, H.A.; Kleinschmidt, J.A.; Boekstegers, P.; Müller, O.J. Cardio-specific long-term gene expression in a porcine model after selective pressure-regulated retroinfusion of adeno-associated viral (AAV) vectors. Gene Ther. 2008, 15, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Bish, L.T.; Sleeper, M.M.; Reynolds, C.; Gazzara, J.; Withnall, E.; Singletary, G.E.; Buchlis, G.; Hui, D.; High, K.A.; Gao, G.; et al. Cardiac gene transfer of short hairpin RNA directed against phospholamban effectively knocks down gene expression but causes cellular toxicity in canines. Hum. Gene Ther. 2011, 22, 969–977. [Google Scholar] [CrossRef] [Green Version]
- White, J.D.; Thesier, D.M.; Swain, J.B.; Katz, M.G.; Tomasulo, C.; Henderson, A.; Wang, L.; Yarnall, C.; Fargnoli, A.; Sumaroka, M.; et al. Myocardial gene delivery using molecular cardiac surgery with recombinant adeno-associated virus vectors in vivo. Gene Ther. 2011, 18, 546–552. [Google Scholar] [CrossRef]
- Gao, G.; Vandenberghe, L.H.; Alvira, M.R.; Lu, Y.; Calcedo, R.; Zhou, X.; Wilson, J.M. Clades of Adeno-associated viruses are widely disseminated in human tissues. J. Virol. 2004, 78, 6381–6388. [Google Scholar] [CrossRef] [Green Version]
- Denby, L.; Nicklin, S.A.; Baker, A.H. Adeno-associated virus (AAV)-7 and -8 poorly transduce vascular endothelial cells and are sensitive to proteasomal degradation. Gene Ther. 2005, 12, 1534–1538. [Google Scholar] [CrossRef]
- Mietzsch, M.; Broecker, F.; Reinhardt, A.; Seeberger, P.H.; Heilbronn, R. Differential Adeno-Associated Virus Serotype-Specific Interaction Patterns with Synthetic Heparins and Other Glycans. J. Virol. 2014, 88, 2991–3003. [Google Scholar] [CrossRef] [Green Version]
- Calcedo, R.; Vandenberghe, L.H.; Gao, G.; Lin, J.; Wilson, J.M. Worldwide Epidemiology of Neutralizing Antibodies to Adeno-Associated Viruses. J. Infect. Dis. 2009, 199, 381–390. [Google Scholar] [CrossRef]
- Louboutin, J.-P.; Wang, L.; Wilson, J.M. Gene transfer into skeletal muscle using novel AAV serotypes. J. Gene Med. A Cross-Discip. J. Res. Sci. Gene Transf. Clin. Appl. 2005, 7, 442–451. [Google Scholar] [CrossRef]
- Samaranch, L.; Salegio, E.A.; San Sebastian, W.; Kells, A.P.; Bringas, J.R.; Forsayeth, J.; Bankiewicz, K.S. Strong cortical and spinal cord transduction after AAV7 and AAV9 delivery into the cerebrospinal fluid of nonhuman primates. Hum. Gene Ther. 2013, 24, 526–532. [Google Scholar] [CrossRef] [Green Version]
- Taymans, J.M.; Vandenberghe, L.H.; Haute, C.V.; Thiry, I.; Deroose, C.M.; Mortelmans, L.; Wilson, J.M.; Debyser, Z.; Baekelandt, V. Comparative analysis of adeno-associated viral vector serotypes 1, 2, 5, 7, and 8 in mouse brain. Hum. Gene Ther. 2007, 18, 195–206. [Google Scholar] [CrossRef]
- Allocca, M.; Mussolino, C.; Garcia-Hoyos, M.; Sanges, D.; Iodice, C.; Petrillo, M.; Vandenberghe, L.H.; Wilson, J.M.; Marigo, V.; Surace, E.M.; et al. Novel adeno-associated virus serotypes efficiently transduce murine photoreceptors. J. Virol. 2007, 81, 11372–11380. [Google Scholar] [CrossRef] [Green Version]
- Okada, T.; Nonaka-Sarukawa, M.; Uchibori, R.; Kinoshita, K.; Hayashita-Kinoh, H.; Nitahara-Kasahara, Y.; Takeda, S.i.; Ozawa, K. Scalable purification of adeno-associated virus serotype 1 (AAV1) and AAV8 vectors, using dual ion-exchange adsorptive membranes. Hum. Gene Ther. 2009, 20, 1013–1021. [Google Scholar] [CrossRef]
- Davidoff, A.M.; Ng, C.Y.; Sleep, S.; Gray, J.; Azam, S.; Zhao, Y.; McIntosh, J.H.; Karimipoor, M.; Nathwani, A.C. Purification of recombinant adeno-associated virus type 8 vectors by ion exchange chromatography generates clinical grade vector stock. J. Virol. Methods 2004, 121, 209–215. [Google Scholar] [CrossRef]
- Lane, M.D.; Nam, H.-J.; Padron, E.; Gurda-Whitaker, B.; Kohlbrenner, E.; Aslanidi, G.; Byrne, B.; McKenna, R.; Muzyczka, N.; Zolotukhin, S.; et al. Production, purification, crystallization and preliminary X-ray analysis of adeno-associated virus serotype 8. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2005, 61, 558–561. [Google Scholar] [CrossRef] [Green Version]
- Miyake, K.; Miyake, N.; Yamazaki, Y.; Shimada, T.; Hirai, Y.J. Serotype-independent method of recombinant adeno-associated virus (AAV) vector production and purification. J. Nippon. Med. Sch. 2012, 79, 394–402. [Google Scholar] [CrossRef] [Green Version]
- Nam, H.-J.; Lane, M.D.; Padron, E.; Gurda, B.; McKenna, R.; Kohlbrenner, E.; Aslanidi, G.; Byrne, B.; Muzyczka, N.; Zolotukhin, S.; et al. Structure of Adeno-Associated Virus Serotype 8, a Gene Therapy Vector. J. Virol. 2007, 81, 12260–12271. [Google Scholar] [CrossRef] [Green Version]
- Cabanes-Creus, M.; Navarro, R.G.; Zhu, E.; Baltazar, G.; Liao, S.H.Y.; Drouyer, M.; Amaya, A.K.; Scott, S.; Nguyen, L.H.; Westhaus, A.; et al. Novel human liver-tropic AAV variants define transferable domains that markedly enhance the human tropism of AAV7 and AAV8. Mol. Ther. Methods Clin. Dev. 2022, 24, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Nakai, H.; Fuess, S.; Storm, T.A.; Muramatsu, S.; Nara, Y.; Kay, M.A. Unrestricted hepatocyte transduction with adeno-associated virus serotype 8 vectors in mice. J. Virol. 2005, 79, 214–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.E.; Storm, T.A.; Huang, Z.; Kay, M.A. Rapid uncoating of vector genomes is the key to efficient liver transduction with pseudotyped adeno-associated virus vectors. J. Virol. 2004, 78, 3110–3122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, H.-J.; Gurda, B.L.; McKenna, R.; Potter, M.; Byrne, B.; Salganik, M.; Muzyczka, N.; Agbandje-McKenna, M. Structural Studies of Adeno-Associated Virus Serotype 8 Capsid Transitions Associated with Endosomal Trafficking. J. Virol. 2011, 85, 11791–11799. [Google Scholar] [CrossRef] [Green Version]
- Monahan, P.E.; Lothrop, C.D.; Sun, J.; Hirsch, M.L.; Kafri, T.; Kantor, B.; Sarkar, R.; Tillson, D.M.; Elia, J.R.; Samulski, R.J. Proteasome inhibitors enhance gene delivery by AAV virus vectors expressing large genomes in hemophilia mouse and dog models: A strategy for broad clinical application. Mol. Ther. J. Am. Soc. Gene Ther. 2010, 18, 1907–1916. [Google Scholar] [CrossRef]
- Jiang, H.; Couto, L.B.; Patarroyo-White, S.; Liu, T.; Nagy, D.; Vargas, J.A.; Zhou, S.; Scallan, C.D.; Sommer, J.; Vijay, S.; et al. Effects of transient immunosuppression on adenoassociated, virus-mediated, liver-directed gene transfer in rhesus macaques and implications for human gene therapy. Blood 2006, 108, 3321–3328. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zhu, T.; Qiao, C.; Zhou, L.; Wang, B.; Zhang, J.; Chen, C.; Li, J.; Xiao, X. Adeno-associated virus serotype 8 efficiently delivers genes to muscle and heart. Nat. Biotechnol. 2005, 23, 321–328. [Google Scholar] [CrossRef]
- Wang, A.Y.; Peng, P.D.; Ehrhardt, A.; Storm, T.A.; Kay, M.A. Comparison of adenoviral and adeno-associated viral vectors for pancreatic gene delivery in vivo. Hum. Gene Ther. 2004, 15, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Loiler, S.A.; Tang, Q.; Clarke, T.; Campbell-Thompson, M.L.; Chiodo, V.; Hauswirth, W.; Cruz, P.; Perret-Gentil, M.; Atkinson, M.A.; Ramiya, V.K.; et al. Localized gene expression following administration of adeno-associated viral vectors via pancreatic ducts. Mol. Ther. J. Am. Soc. Gene Ther. 2005, 12, 519–527. [Google Scholar] [CrossRef]
- Rubin, J.D.; Nguyen, T.V.; Allen, K.L.; Ayasoufi, K.; Barry, M.A. Comparison of gene delivery to the kidney by adenovirus, adeno-associated virus, and lentiviral vectors after intravenous and direct kidney injections. Hum. Gene Ther. 2019, 30, 1559–1571. [Google Scholar] [CrossRef]
- Giove, T.J.; Khankhel, Z.S.; Sena-Esteves, M.; Eldred, W.D. Cellular Tropism of Aav8, Aav9 and Aav10 in Mouse Retina. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3013. [Google Scholar]
- Ding, K.; Shen, J.; Hafiz, Z.; Hackett, S.F.; Silva, R.L.E.; Khan, M.; Lorenc, V.E.; Chen, D.; Chadha, R.; Zhang, M.; et al. AAV8-vectored suprachoroidal gene transfer produces widespread ocular transgene expression. J. Clin. Investig. 2019, 129, 4901–4911. [Google Scholar] [CrossRef]
- Igarashi, T.; Miyake, K.; Asakawa, N.; Miyake, N.; Shimada, T.; Takahashi, H. Direct Comparison of Administration Routes for AAV8-mediated Ocular Gene Therapy. Curr. Eye Res. 2013, 38, 569–577. [Google Scholar] [CrossRef]
- Gao, G.; Vandenberghe, L.H.; Wilson, J.M. New recombinant serotypes of AAV vectors. Curr. Gene Ther. 2005, 5, 285–297. [Google Scholar] [CrossRef]
- Shen, S.; Bryant, K.D.; Brown, S.M.; Randell, S.H.; Asokan, A. Terminal N-Linked Galactose Is the Primary Receptor for Adeno-associated Virus 9*. J. Biol. Chem. 2011, 286, 13532–13540. [Google Scholar] [CrossRef] [Green Version]
- Bell, C.L.; Gurda, B.L.; Van Vliet, K.; Agbandje-McKenna, M.; Wilson, J.M. Identification of the galactose binding domain of the adeno- associated virus serotype 9 capsid. J. Virol. 2012, 86, 7326–7333. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, M.; Nam, H.-J.; Carter, A.; McCall, A.; Rence, C.; Bennett, A.; Gurda, B.; McKenna, R.; Porter, M.; Sakai, Y.; et al. Production, purification and preliminary X-ray crystallographic studies of adeno-associated virus serotype 9. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2009, 65, 715–718. [Google Scholar] [CrossRef] [Green Version]
- Foust, K.D.; Nurre, E.; Montgomery, C.L.; Hernandez, A.; Chan, C.M.; Kaspar, B.K. Intravascular AAV9 preferentially targets neonatal neurons and adult astrocytes. Nat. Biotechnol. 2009, 27, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.J.; Matagne, V.; Bachaboina, L.; Yadav, S.; Ojeda, S.R.; Samulski, R.J. Preclinical Differences of Intravascular AAV9 Delivery to Neurons and Glia: A Comparative Study of Adult Mice and Nonhuman Primates. Mol. Ther. 2011, 19, 1058–1069. [Google Scholar] [CrossRef]
- Bevan, A.K.; Duque, S.; Foust, K.D.; Morales, P.R.; Braun, L.; Schmelzer, L.; Chan, C.M.; McCrate, M.; Chicoine, L.G.; Coley, B.D.; et al. Systemic Gene Delivery in Large Species for Targeting Spinal Cord, Brain, and Peripheral Tissues for Pediatric Disorders. Mol. Ther. 2011, 19, 1971–1980. [Google Scholar] [CrossRef] [Green Version]
- Duque, S.; Joussemet, B.; Riviere, C.; Marais, T.; Dubreil, L.; Douar, A.M.; Fyfe, J.; Moullier, P.; Colle, M.A.; Barkats, M. Intravenous administration of self-complementary AAV9 enables transgene delivery to adult motor neurons. Mol. Ther. J. Am. Soc. Gene Ther. 2009, 17, 1187–1196. [Google Scholar] [CrossRef]
- Vandenberghe, L.H.; Bell, P.; Maguire, A.M.; Xiao, R.; Hopkins, T.B.; Grant, R.; Bennett, J.; Wilson, J.M.; Xiao, X. AAV9 targets cone photoreceptors in the nonhuman primate retina. PLoS ONE 2013, 8, e53463. [Google Scholar] [CrossRef]
- Pacak, C.A.; Mah, C.S.; Thattaliyath, B.D.; Conlon, T.J.; Lewis, M.A.; Cloutier, D.E.; Zolotukhin, I.; Tarantal, A.F.; Byrne, B.J. Recombinant adeno-associated virus serotype 9 leads to preferential cardiac transduction in vivo. Circ. Res. 2006, 99, e3–e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asokan, A.; Schaffer, D.V.; Samulski, R.J. The AAV vector toolkit: Poised at the clinical crossroads. Mol. Ther. 2012, 20, 699–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pleger, S.T.; Shan, C.; Ksienzyk, J.; Bekeredjian, R.; Boekstegers, P.; Hinkel, R.; Schinkel, S.; Leuchs, B.; Ludwig, J.; Qiu, G. Cardiac AAV9-S100A1 gene therapy rescues post-ischemic heart failure in a preclinical large animal model. Sci. Transl. Med. 2011, 3, 92ra64. [Google Scholar] [CrossRef] [Green Version]
- Bish, L.T.; Morine, K.; Sleeper, M.M.; Sanmiguel, J.; Wu, D.; Gao, G.; Wilson, J.M.; Sweeney, H.L. Adeno-associated virus (AAV) serotype 9 provides global cardiac gene transfer superior to AAV1, AAV6, AAV7, and AAV8 in the mouse and rat. Hum. Gene Ther. 2008, 19, 1359–1368. [Google Scholar] [CrossRef]
- Vandendriessche, T.; Thorrez, L.; Acosta-Sanchez, A.; Petrus, I.; Wang, L.; Ma, L.; De Waele, L.; Iwasaki, Y.; Gillijns, V.; Wilson, J.M.; et al. Efficacy and safety of adeno-associated viral vectors based on serotype 8 and 9 vs. lentiviral vectors for hemophilia B gene therapy. J. Thromb. Haemost. JTH 2007, 5, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, K.; Fuess, S.; Storm, T.A.; Gibson, G.A.; McTiernan, C.F.; Kay, M.A.; Nakai, H. Robust systemic transduction with AAV9 vectors in mice: Efficient global cardiac gene transfer superior to that of AAV8. Mol. Ther. J. Am. Soc. Gene Ther. 2006, 14, 45–53. [Google Scholar] [CrossRef]
- Petrs-Silva, H.; Dinculescu, A.; Li, Q.; Min, S.-H.; Chiodo, V.; Pang, J.-J.; Zhong, L.; Zolotukhin, S.; Srivastava, A.; Lewin, A.S.; et al. High-efficiency Transduction of the Mouse Retina by Tyrosine-mutant AAV Serotype Vectors. Mol. Ther. 2009, 17, 463–471. [Google Scholar] [CrossRef]
- Shen, X.; Xu, Y.; Bai, Z.; Ma, D.; Niu, Q.; Meng, J.; Fan, S.; Zhang, L.; Hao, Z.; Zhang, X.J. Transparenchymal renal pelvis injection of recombinant adeno-associated virus serotype 9 vectors is a practical approach for gene delivery in the kidney. Hum. Gene Ther. Methods 2018, 29, 251–258. [Google Scholar] [CrossRef]
- Konkalmatt, P.; Asico, L.; Feranil, J.; Jose, P.; Armando, I. Efficient in vivo gene transfer to murine renal cells using AAV9 vectors (912.7). FASEB J. 2014, 28, 917. [Google Scholar] [CrossRef]
- Schievenbusch, S.; Strack, I.; Scheffler, M.; Nischt, R.; Coutelle, O.; Hösel, M.; Hallek, M.; Fries, J.W.U.; Dienes, H.-P.; Odenthal, M.; et al. Combined Paracrine and Endocrine AAV9 mediated Expression of Hepatocyte Growth Factor for the Treatment of Renal Fibrosis. Mol. Ther. 2010, 18, 1302–1309. [Google Scholar] [CrossRef]
- Darbey, A.; Rebourcet, D.; Curley, M.; Kilcoyne, K.; Jeffery, N.; Reed, N.; Milne, L.; Roesl, C.; Brown, P.; Smith, L.B. A comparison of in vivo viral targeting systems identifies adeno-associated virus serotype 9 (AAV9) as an effective vector for genetic manipulation of Leydig cells in adult mice. Andrology 2021, 9, 460–473. [Google Scholar] [CrossRef]
- Limberis, M.P.; Wilson, J.M. Adeno-associated virus serotype 9 vectors transduce murine alveolar and nasal epithelia and can be readministered. Proc. Natl. Acad. Sci. USA 2006, 103, 12993–12998. [Google Scholar] [CrossRef] [Green Version]
- Bell, C.L.; Vandenberghe, L.H.; Bell, P.; Limberis, M.P.; Gao, G.-P.; Van Vliet, K.; Agbandje-McKenna, M.; Wilson, J.M. The AAV9 receptor and its modification to improve in vivo lung gene transfer in mice. J. Clin. Investig. 2011, 121, 2427–2435. [Google Scholar] [CrossRef] [Green Version]
- Vance, M.A.; Mitchell, A.; Samulski, R.J. AAV Biology, Infectivity and Therapeutic Use from Bench to Clinic. In Gene Therapy—Principles and Challenges; IntechOpen: London, UK, 2015. [Google Scholar] [CrossRef] [Green Version]
- Vandenberghe, L.H.; Miller, J.R.; Gao, G.-P.; Calcedo, R.; Wilson, J.M. 116. The Proposed AAV Serotypes 10 and 11 Serologically Cross-React with AAV8 and AAV4. Mol. Ther. 2006, 13, S47. [Google Scholar] [CrossRef]
- Klein, R.L.; Dayton, R.D.; Tatom, J.B.; Henderson, K.M.; Henning, P.P. AAV8, 9, Rh10, Rh43 Vector Gene Transfer in the Rat Brain: Effects of Serotype, Promoter and Purification Method. Mol. Ther. 2008, 16, 89–96. [Google Scholar] [CrossRef]
- Mori, S.; Takeuchi, T.; Enomoto, Y.; Kondo, K.; Sato, K.; Ono, F.; Sata, T.; Kanda, T. Tissue distribution of cynomolgus adeno-associated viruses AAV10, AAV11, and AAVcy.7 in naturally infected monkeys. Arch. Virol. 2008, 153, 375–380. [Google Scholar] [CrossRef]
- Polyak, S.; Mach, A.; Porvasnik, S.; Dixon, L.; Conlon, T.; Erger, K.E.; Acosta, A.; Wright, A.J.; Campbell-Thompson, M.; Zolotukhin, I.; et al. Identification of adeno-associated viral vectors suitable for intestinal gene delivery and modulation of experimental colitis. Am. J. Physiol.-Gastrointest. Liver Physiol. 2012, 302, G296–G308. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.K.; Uehara, H.; Zhang, X.; Vashist, D.; Carroll, L.; Archer, B.; Huang, Y.; Ambati, B. Systemic Distribution of AAV10.COMPAng1 in the Ins2Akita mouse. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5056. [Google Scholar]
- Tian, M.; Carroll, L.S.; Tang, L.; Uehara, H.; Westenfelder, C.; Ambati, B.K.; Huang, Y. Systemic AAV10. COMP-Ang1 rescues renal glomeruli and pancreatic islets in type 2 diabetic mice. BMJ Open Diabetes Res. Care 2020, 8, e000882. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Luo, N.; Kou, J.; Li, L.; Ma, W.; Peng, S.; Xu, Z.; Zhang, W.; Qiu, Y.; Wu, Y. AAV11 permits efficient retrograde targeting of projection neurons. bioRxiv 2022. [Google Scholar] [CrossRef]
- Schmidt, M.; Voutetakis, A.; Afione, S.; Zheng, C.; Mandikian, D.; Chiorini, J.A. Adeno-Associated Virus Type 12 (AAV12): A Novel AAV Serotype with Sialic Acid- and Heparan Sulfate Proteoglycan-Independent Transduction Activity. J. Virol. 2008, 82, 1399–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.; Voutetakis, A.; Afione, S.; Zheng, C.; Chiorini, J.A. 745. AAV12, Isolated from Vervet Monkey, Has Unique Tropism and Biological as Well as Neutralization Properties. Mol. Ther. 2006, 13, S288. [Google Scholar] [CrossRef]
- Quinn, K.; Quirion, M.R.; Lo, C.-Y.; Misplon, J.A.; Epstein, S.L.; Chiorini, J.A. Intranasal Administration of Adeno-associated Virus Type 12 (AAV12) Leads to Transduction of the Nasal Epithelia and Can Initiate Transgene-specific Immune Response. Mol. Ther. 2011, 19, 1990–1998. [Google Scholar] [CrossRef]
- Mietzsch, M.; Jose, A.; Chipman, P.; Bhattacharya, N.; Daneshparvar, N.; McKenna, R.; Agbandje-McKenna, M. Completion of the AAV Structural Atlas: Serotype Capsid Structures Reveals Clade-Specific Features. Viruses 2021, 13, 101. [Google Scholar] [CrossRef]
- Schmidt, M.; Govindasamy, L.; Afione, S.; Kaludov, N.; Agbandje-McKenna, M.; Chiorini, J.A. Molecular characterization of the heparin-dependent transduction domain on the capsid of a novel adeno-associated virus isolate, AAV(VR-942). J. Virol. 2008, 82, 8911–8916. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.-Y.; Halder, S.; Agbandje-McKenna, M. Parvovirus glycan interactions. Curr. Opin. Virol. 2014, 7, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.J.C.; Insights, G.T. Advances and challenges in the use of recombinant AAV vectors for human gene therapy. Cell Gene Ther. Insights 2016, 2, 553–575. [Google Scholar] [CrossRef]
- Muzyczka, N.; Warrington, K.H., Jr. Custom adeno-associated virus capsids: The next generation of recombinant vectors with novel tropism. Hum. Gene Ther. 2005, 16, 408–416. [Google Scholar] [CrossRef]
- Zhang, L.; Rossi, A.; Lange, L.; Meumann, N.; Koitzsch, U.; Christie, K.; Nesbit, M.A.; Moore, C.B.T.; Hacker, U.T.; Morgan, M.; et al. Capsid Engineering Overcomes Barriers Toward Adeno-Associated Virus Vector-Mediated Transduction of Endothelial Cells. Hum. Gene Ther. 2019, 30, 1284–1296. [Google Scholar] [CrossRef]
- Ward, P.; Walsh, C.E. Chimeric AAV Cap sequences alter gene transduction. Virology 2009, 386, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Jiang, J.; Drouin, L.M.; Agbandje-McKenna, M.; Chen, C.; Qiao, C.; Pu, D.; Hu, X.; Wang, D.Z.; Li, J.; et al. A myocardium tropic adeno-associated virus (AAV) evolved by DNA shuffling and in vivo selection. Proc. Natl. Acad. Sci. USA 2009, 106, 3946–3951. [Google Scholar] [CrossRef] [Green Version]
- White, A.F.; Mazur, M.; Sorscher, E.J.; Zinn, K.R.; Ponnazhagan, S. Genetic modification of adeno-associated viral vector type 2 capsid enhances gene transfer efficiency in polarized human airway epithelial cells. Hum. Gene Ther. 2008, 19, 1407–1414. [Google Scholar] [CrossRef]
- Zhang, C.; Freistaedter, A.; Schmelas, C.; Gunkel, M.; Dao Thi, V.L.; Grimm, D. An RNA Interference/Adeno-Associated Virus Vector-Based Combinatorial Gene Therapy Approach Against Hepatitis E Virus. Hepatol. Commun. 2022, 6, 878–888. [Google Scholar] [CrossRef]
- Van Vliet, K.M.; Blouin, V.; Brument, N.; Agbandje-McKenna, M.; Snyder, R.O. The role of the adeno-associated virus capsid in gene transfer. Methods Mol. Biol. 2008, 437, 51–91. [Google Scholar] [CrossRef]
- Perabo, L.; Endell, J.; King, S.; Lux, K.; Goldnau, D.; Hallek, M.; Büning, H. Combinatorial engineering of a gene therapy vector: Directed evolution of adeno-associated virus. J. Gene Med. 2006, 8, 155–162. [Google Scholar] [CrossRef]
- Maheshri, N.; Koerber, J.T.; Kaspar, B.K.; Schaffer, D.V. Directed evolution of adeno-associated virus yields enhanced gene delivery vectors. Nat. Biotechnol. 2006, 24, 198–204. [Google Scholar] [CrossRef]
- Xiao, X.; deVlaminck, W.; Monahan, J. Adeno-associated virus (AAV) vectors for gene transfer. Adv. Drug Deliv. Rev. 1993, 12, 201–215. [Google Scholar] [CrossRef]
- Han, Z.; Zhong, L.; Maina, N.; Hu, Z.; Li, X.; Chouthai, N.S.; Bischof, D.; Weigel-Van Aken, K.A.; Slayton, W.B.; Yoder, M.C. Stable integration of recombinant adeno-associated virus vector genomes after transduction of murine hematopoietic stem cells. Hum. Gene Ther. 2008, 19, 267–278. [Google Scholar] [CrossRef]
- Tratschin, J.-D.; Miller, I.; Smith, M.; Carter, B.J.M.; Biology, C. Adeno-associated virus vector for high-frequency integration, expression, and rescue of genes in mammalian cells. Mol. Cell. Biol. 1985, 5, 3251–3260. [Google Scholar] [PubMed] [Green Version]
- Samulski, R.; Zhu, X.; Xiao, X.; Brook, J.; Housman, D.; Epstein, N.A.; Hunter, L. Targeted integration of adeno-associated virus (AAV) into human chromosome 19. EMBO J. 1991, 10, 3941–3950. [Google Scholar] [CrossRef]
- McCarty, D.M.; Young, S.M., Jr.; Samulski, R.J. Integration of adeno-associated virus (AAV) and recombinant AAV vectors. Annu. Rev. Genet. 2004, 38, 819–845. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.E.; Liu, J.M.; Xiao, X.; Young, N.S.; Nienhuis, A.W.; Samulski, R.J. Regulated high level expression of a human gamma-globin gene introduced into erythroid cells by an adeno-associated virus vector. Proc. Natl. Acad. Sci. USA 1992, 89, 7257–7261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winocour, E.; Callaham, M.; Huberman, E. Perturbation of the cell cycle by adeno-associated virus. Virology 1988, 167, 393–399. [Google Scholar] [CrossRef]
- Bantel-Schaal, U.; Stöhr, M.J. Influence of adeno-associated virus on adherence and growth properties of normal cells. J. Virol. 1992, 66, 773–779. [Google Scholar] [CrossRef] [Green Version]
- Samulski, R.J. Adeno-associated virus: Integration at a specific chromosomal locus. Curr. Opin. Genet. Dev. 1993, 3, 74–80. [Google Scholar] [CrossRef]
- Bijlani, S.; Pang, K.M.; Sivanandam, V.; Singh, A.; Chatterjee, S. The Role of Recombinant AAV in Precise Genome Editing. Front. Genome Ed. 2021, 3, 799722. [Google Scholar] [CrossRef]
- Wang, D.; Tai, P.W.; Gao, G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat. Rev. Drug Discov. 2019, 18, 358–378. [Google Scholar] [CrossRef]
- Calcedo, R.; Wilson, J.M. Humoral immune response to AAV. Front. Immunol. 2013, 4, 341. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, M.L.; Samulski, R.J. AAV-mediated gene editing via double-strand break repair. In Gene Correction; Springer: Berlin/Heidelberg, Germany, 2014; pp. 291–307. [Google Scholar]
- Gray, S.J.; Nagabhushan Kalburgi, S.; McCown, T.J.; Jude Samulski, R. Global CNS gene delivery and evasion of anti-AAV-neutralizing antibodies by intrathecal AAV administration in non-human primates. Gene Ther. 2013, 20, 450–459. [Google Scholar] [CrossRef] [Green Version]
- Ge, L.-J.; Yang, F.-H.; Li, W.; Wang, T.; Lin, Y.; Feng, J.; Chen, N.-H.; Jiang, M.; Wang, J.-H.; Hu, X.-T. In vivo neuroregeneration to treat ischemic stroke through NeuroD1 AAV-based gene therapy in adult non-human primates. Front. Cell Dev. Biol. 2020, 8, 590008. [Google Scholar] [CrossRef]
- Hu, C.; Lipshutz, G.S. AAV-based neonatal gene therapy for hemophilia A: Long-term correction and avoidance of immune responses in mice. Gene Ther. 2012, 19, 1166–1176. [Google Scholar] [CrossRef]
- Suzuki, J.; Hashimoto, K.; Xiao, R.; Vandenberghe, L.H.; Liberman, M.C. Cochlear gene therapy with ancestral AAV in adult mice: Complete transduction of inner hair cells without cochlear dysfunction. Sci. Rep. 2017, 7, 45524. [Google Scholar] [CrossRef] [Green Version]
- Akil, O.; Dyka, F.; Calvet, C.; Emptoz, A.; Lahlou, G.; Nouaille, S.; Boutet de Monvel, J.; Hardelin, J.-P.; Hauswirth, W.W.; Avan, P. Dual AAV-mediated gene therapy restores hearing in a DFNB9 mouse model. Proc. Natl. Acad. Sci. USA 2019, 116, 4496–4501. [Google Scholar] [CrossRef] [Green Version]
- Acland, G.M.; Aguirre, G.D.; Ray, J.; Zhang, Q.; Aleman, T.S.; Cideciyan, A.V.; Pearce-Kelling, S.E.; Anand, V.; Zeng, Y.; Maguire, A.M. Gene therapy restores vision in a canine model of childhood blindness. Nat. Genet. 2001, 28, 92–95. [Google Scholar] [CrossRef]
- Kodippili, K.; Hakim, C.H.; Pan, X.; Yang, H.T.; Yue, Y.; Zhang, Y.; Shin, J.-H.; Yang, N.N.; Duan, D. Dual AAV gene therapy for Duchenne muscular dystrophy with a 7-kb mini-dystrophin gene in the canine model. Hum. Gene Ther. 2018, 29, 299–311. [Google Scholar] [CrossRef]
- Gurda, B.L.; De Guilhem De Lataillade, A.; Bell, P.; Zhu, Y.; Yu, H.; Wang, P.; Bagel, J.; Vite, C.H.; Sikora, T.; Hinderer, C.; et al. Evaluation of AAV-mediated Gene Therapy for Central Nervous System Disease in Canine Mucopolysaccharidosis VII. Mol. Ther. J. Am. Soc. Gene Ther. 2016, 24, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Gray-Edwards, H.L.; Maguire, A.S.; Salibi, N.; Ellis, L.E.; Voss, T.L.; Diffie, E.B.; Koehler, J.; Randle, A.N.; Taylor, A.R.; Brunson, B.L.; et al. 7T MRI predicts amelioration of neurodegeneration in the brain after AAV gene therapy. Mol. Ther.-Methods Clin. Dev. 2020, 17, 258–270. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, S.R.; Fitzpatrick, Z.; Harris, A.F.; Maitland, S.A.; Ferreira, J.S.; Zhang, Y.; Ma, S.; Sharma, R.B.; Gray-Edwards, H.L.; Johnson, J.A. In vivo selection yields AAV-B1 capsid for central nervous system and muscle gene therapy. Mol. Ther. 2016, 24, 1247–1257. [Google Scholar] [CrossRef] [Green Version]
- McCown, T.J. Adeno-associated virus (AAV) vectors in the CNS. Curr. Gene Ther. 2005, 5, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Kaplitt, M.G.; Leone, P.; Samulski, R.J.; Xiao, X.; Pfaff, D.W.; O’Malley, K.L.; During, M.J. Long-term gene expression and phenotypic correction using adeno-associated virus vectors in the mammalian brain. Nat. Genet. 1994, 8, 148–154. [Google Scholar] [CrossRef] [PubMed]
- McElroy, A.; Sena-Esteves, M.; Arjomandnejad, M.; Keeler, A.M.; Gray-Edwards, H.L. Redosing Adeno-Associated Virus Gene Therapy to the Central Nervous System. Hum. Gene Ther. 2022, 33, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Bartus, R.T.; Weinberg, M.S.; Samulski, R.J. Parkinson’s Disease Gene Therapy: Success by Design Meets Failure by Efficacy. Mol. Ther. 2014, 22, 487–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marks, W.J.; Bartus, R.T.; Siffert, J.; Davis, C.S.; Lozano, A.; Boulis, N.; Vitek, J.; Stacy, M.; Turner, D.; Verhagen, L.; et al. Gene delivery of AAV2-neurturin for Parkinson’s disease: A double-blind, randomised, controlled trial. Lancet Neurol. 2010, 9, 1164–1172. [Google Scholar] [CrossRef]
- Kaplitt, M.G.; Feigin, A.; Tang, C.; Fitzsimons, H.L.; Mattis, P.; Lawlor, P.A.; Bland, R.J.; Young, D.; Strybing, K.; Eidelberg, D.; et al. Safety and tolerability of gene therapy with an adeno-associated virus (AAV) borne GAD gene for Parkinson’s disease: An open label, phase I trial. Lancet 2007, 369, 2097–2105. [Google Scholar] [CrossRef]
- LeWitt, P.A.; Rezai, A.R.; Leehey, M.A.; Ojemann, S.G.; Flaherty, A.W.; Eskandar, E.N.; Kostyk, S.K.; Thomas, K.; Sarkar, A.; Siddiqui, M.S.; et al. AAV2-GAD gene therapy for advanced Parkinson’s disease: A double-blind, sham-surgery controlled, randomised trial. Lancet Neurol. 2011, 10, 309–319. [Google Scholar] [CrossRef]
- Bartus, R.T.; Baumann, T.L.; Siffert, J.; Herzog, C.D.; Alterman, R.; Boulis, N.; Turner, D.A.; Stacy, M.; Lang, A.E.; Lozano, A.M.; et al. Safety/feasibility of targeting the substantia nigra with AAV2-neurturin in Parkinson patients. Neurology 2013, 80, 1698–1701. [Google Scholar] [CrossRef] [Green Version]
- Christine, C.W.; Starr, P.A.; Larson, P.S.; Eberling, J.L.; Jagust, W.J.; Hawkins, R.A.; VanBrocklin, H.F.; Wright, J.F.; Bankiewicz, K.S.; Aminoff, M.J. Safety and tolerability of putaminal AADC gene therapy for Parkinson disease. Neurology 2009, 73, 1662–1669. [Google Scholar] [CrossRef] [Green Version]
- Mittermeyer, G.; Christine, C.W.; Rosenbluth, K.H.; Baker, S.L.; Starr, P.; Larson, P.; Kaplan, P.L.; Forsayeth, J.; Aminoff, M.J.; Bankiewicz, K.S. Long-term evaluation of a phase 1 study of AADC gene therapy for Parkinson’s disease. Hum. Gene Ther. 2012, 23, 377–381. [Google Scholar] [CrossRef] [Green Version]
- Chien, Y.H.; Lee, N.C.; Tseng, S.H.; Tai, C.H.; Muramatsu, S.I.; Byrne, B.J.; Hwu, W.L. Efficacy and safety of AAV2 gene therapy in children with aromatic L-amino acid decarboxylase deficiency: An open-label, phase 1/2 trial. Lancet Child Adolesc. Health 2017, 1, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Kolb, S.J.; Kissel, J.T. Spinal muscular atrophy: A timely review. Arch. Neurol. 2011, 68, 979–984. [Google Scholar] [CrossRef] [Green Version]
- Aguti, S.; Malerba, A.; Zhou, H. The progress of AAV-mediated gene therapy in neuromuscular disorders. Expert Opin. Biol. Ther. 2018, 18, 681–693. [Google Scholar] [CrossRef]
- Spahiu, L.; Behluli, E.; Peterlin, B.; Nefic, H.; Hadziselimovic, R.; Liehr, T.; Temaj, G. Mucopolysaccharidosis III: Molecular basis and treatment. Pediatr. Endocrinol. Diabetes Metab. 2021, 27, 201–208. [Google Scholar] [CrossRef]
- Watson, G.L.; Sayles, J.N.; Chen, C.; Elliger, S.S.; Elliger, C.A.; Raju, N.R.; Kurtzman, G.J.; Podsakoff, G.M. Treatment of lysosomal storage disease in MPS VII mice using a recombinant adeno-associated virus. Gene Ther. 1998, 5, 1642–1649. [Google Scholar] [CrossRef] [Green Version]
- Skorupa, A.F.; Fisher, K.J.; Wilson, J.M.; Parente, M.K.; Wolfe, J.H. Sustained production of beta-glucuronidase from localized sites after AAV vector gene transfer results in widespread distribution of enzyme and reversal of lysosomal storage lesions in a large volume of brain in mucopolysaccharidosis VII mice. Exp. Neurol. 1999, 160, 17–27. [Google Scholar] [CrossRef]
- Daly, T.M.; Vogler, C.; Levy, B.; Haskins, M.E.; Sands, M.S. Neonatal gene transfer leads to widespread correction of pathology in a murine model of lysosomal storage disease. Proc. Natl. Acad. Sci. USA 1999, 96, 2296–2300. [Google Scholar] [CrossRef] [Green Version]
- Marcó, S.; Haurigot, V.; Jaén, M.L.; Ribera, A.; Sánchez, V.; Molas, M.; Garcia, M.; León, X.; Roca, C.; Sánchez, X.; et al. Seven-year follow-up of durability and safety of AAV CNS gene therapy for a lysosomal storage disorder in a large animal. Mol. Ther.-Methods Clin. Dev. 2021, 23, 370–389. [Google Scholar] [CrossRef]
- Shaimardanova, A.A.; Chulpanova, D.S.; Solovyeva, V.V.; Mullagulova, A.I.; Kitaeva, K.V.; Allegrucci, C.; Rizvanov, A.A. Metachromatic Leukodystrophy: Diagnosis, Modeling, and Treatment Approaches. Front. Med. 2020, 7, 576221. [Google Scholar] [CrossRef]
- Piguet, F.; Sondhi, D.; Piraud, M.; Fouquet, F.; Hackett, N.R.; Ahouansou, O.; Vanier, M.-T.; Bieche, I.; Aubourg, P.; Crystal, R.G. Correction of brain oligodendrocytes by AAVrh. 10 intracerebral gene therapy in metachromatic leukodystrophy mice. Hum. Gene Ther. 2012, 23, 903–914. [Google Scholar] [CrossRef] [Green Version]
- Kurai, T.; Hisayasu, S.; Kitagawa, R.; Migita, M.; Suzuki, H.; Hirai, Y.; Shimada, T. AAV1 mediated co-expression of formylglycine-generating enzyme and arylsulfatase a efficiently corrects sulfatide storage in a mouse model of metachromatic leukodystrophy. Mol. Ther. 2007, 15, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Miyake, N.; Miyake, K.; Asakawa, N.; Yamamoto, M.; Shimada, T. Long-term correction of biochemical and neurological abnormalities in MLD mice model by neonatal systemic injection of an AAV serotype 9 vector. Gene Ther. 2014, 21, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevin, C.; Benraiss, A.; Van Dam, D.; Bonnin, D.; Nagels, G.; Verot, L.; Laurendeau, I.; Vidaud, M.; Gieselmann, V.; Vanier, M. Intracerebral adeno-associated virus-mediated gene transfer in rapidly progressive forms of metachromatic leukodystrophy. Hum. Mol. Genet. 2006, 15, 53–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaimardanova, A.A.; Chulpanova, D.S.; Solovyeva, V.V.; Aimaletdinov, A.M.; Rizvanov, A.A. Functionality of a bicistronic construction containing HEXA and HEXB genes encoding β-hexosaminidase A for cell-mediated therapy of GM2 gangliosidoses. Neural Regen. Res. 2022, 17, 122–129. [Google Scholar] [CrossRef]
- Taghian, T.; Marosfoi, M.G.; Puri, A.S.; Cataltepe, O.I.; King, R.M.; Diffie, E.B.; Maguire, A.S.; Martin, D.R.; Fernau, D.; Batista, A.R.; et al. A Safe and Reliable Technique for CNS Delivery of AAV Vectors in the Cisterna Magna. Mol. Ther. J. Am. Soc. Gene Ther. 2020, 28, 411–421. [Google Scholar] [CrossRef]
- Solovyeva, V.V.; Shaimardanova, A.A.; Chulpanova, D.S.; Kitaeva, K.V.; Chakrabarti, L.; Rizvanov, A.A. New Approaches to Tay-Sachs Disease Therapy. Front. Physiol. 2018, 9, 1663. [Google Scholar] [CrossRef] [Green Version]
- Cachón-González, M.B.; Wang, S.Z.; Lynch, A.; Ziegler, R.; Cheng, S.H.; Cox, T.M. Effective gene therapy in an authentic model of Tay-Sachs-related diseases. Proc. Natl. Acad. Sci. USA 2006, 103, 10373–10378. [Google Scholar] [CrossRef] [Green Version]
- Gray-Edwards, H.L.; Randle, A.N.; Maitland, S.A.; Benatti, H.R.; Hubbard, S.M.; Canning, P.F.; Vogel, M.B.; Brunson, B.L.; Hwang, M.; Ellis, L.E.; et al. Adeno-Associated Virus Gene Therapy in a Sheep Model of Tay-Sachs Disease. Hum. Gene Ther. 2018, 29, 312–326. [Google Scholar] [CrossRef]
- Rockwell, H.E.; McCurdy, V.J.; Eaton, S.C.; Wilson, D.U.; Johnson, A.K.; Randle, A.N.; Bradbury, A.M.; Gray-Edwards, H.L.; Baker, H.J.; Hudson, J.A.; et al. AAV-mediated gene delivery in a feline model of Sandhoff disease corrects lysosomal storage in the central nervous system. ASN Neuro 2015, 7, 1759091415569908. [Google Scholar] [CrossRef]
- Sevin, C.; Roujeau, T.; Cartier, N.; Baugnon, T.; Adamsbaum, C.; Piraud, M.; Martino, S.; Mouiller, P.; Couzinié, C.; Bellesme, C. Intracerebral Gene Therapy in Children with Metachromatic Leukodystrophy: Results of a Phase I/II Trial. Mol. Genet. Metab. 2018, 123, S129. [Google Scholar] [CrossRef]
- Tenenbaum, L.; Chtarto, A.; Lehtonen, E.; Velu, T.; Brotchi, J.; Levivier, M. Recombinant AAV-mediated gene delivery to the central nervous system. J. Gene Med. 2004, 6, S212–S222. [Google Scholar] [CrossRef]
- Surendran, S.; Michals-Matalon, K.; Quast, M.J.; Tyring, S.K.; Wei, J.; Ezell, E.L.; Matalon, R. Canavan disease: A monogenic trait with complex genomic interaction. Mol. Genet. Metab. 2003, 80, 74–80. [Google Scholar] [CrossRef]
- Matalon, R.; Surendran, S.; Rady, P.L.; Quast, M.J.; Campbell, G.A.; Matalon, K.M.; Tyring, S.K.; Wei, J.; Peden, C.S.; Ezell, E.L.; et al. Adeno-associated virus-mediated aspartoacylase gene transfer to the brain of knockout mouse for canavan disease. Mol. Ther. J. Am. Soc. Gene Ther. 2003, 7, 580–587. [Google Scholar] [CrossRef]
- Janson, C.; McPhee, S.; Bilaniuk, L.; Haselgrove, J.; Testaiuti, M.; Freese, A.; Wang, D.J.; Shera, D.; Hurh, P.; Rupin, J.; et al. Clinical protocol. Gene therapy of Canavan disease: AAV-2 vector for neurosurgical delivery of aspartoacylase gene (ASPA) to the human brain. Hum. Gene Ther. 2002, 13, 1391–1412. [Google Scholar] [CrossRef] [Green Version]
- rAAV-Olig001-ASPA Gene Therapy for Treatment of Children With Typical Canavan Disease. Available online: https://ClinicalTrials.gov/show/NCT04833907 (accessed on 1 December 2022).
- Canavan-Single Patient IND. Available online: https://ClinicalTrials.gov/show/NCT05317780 (accessed on 1 December 2022).
- A Study of AAV9 Gene Therapy in Participants With Canavan Disease. Available online: https://ClinicalTrials.gov/show/NCT04998396 (accessed on 1 December 2022).
- Nasir, G.; Chopra, R.; Elwood, F.; Ahmed, S.S. Krabbe Disease: Prospects of Finding a Cure Using AAV Gene Therapy. Front. Med. 2021, 8, 760236. [Google Scholar] [CrossRef]
- Rafi, M.A.; Rao, H.Z.; Passini, M.A.; Curtis, M.; Vanier, M.T.; Zaka, M.; Luzi, P.; Wolfe, J.H.; Wenger, D.A. AAV-mediated expression of galactocerebrosidase in brain results in attenuated symptoms and extended life span in murine models of globoid cell leukodystrophy. Mol. Ther. 2005, 11, 734–744. [Google Scholar] [CrossRef]
- Lin, D.-S.; Hsiao, C.-D.; Liau, I.; Lin, S.-P.; Chiang, M.-F.; Chuang, C.-K.; Wang, T.-J.; Wu, T.-Y.; Jian, Y.-R.; Huang, S.-F.; et al. CNS-targeted AAV5 gene transfer results in global dispersal of vector and prevention of morphological and function deterioration in CNS of globoid cell leukodystrophy mouse model. Mol. Genet. Metab. 2011, 103, 367–377. [Google Scholar] [CrossRef]
- Lin, D.; Fantz, C.R.; Levy, B.; Rafi, M.A.; Vogler, C.; Wenger, D.A.; Sands, M.S. AAV2/5 vector expressing galactocerebrosidase ameliorates CNS disease in the murine model of globoid-cell leukodystrophy more efficiently than AAV2. Mol. Ther. 2005, 12, 422–430. [Google Scholar] [CrossRef]
- Karumuthil-Melethil, S.; Marshall, M.S.; Heindel, C.; Jakubauskas, B.; Bongarzone, E.R.; Gray, S.J. Intrathecal administration of AAV/GALC vectors in 10–11-day-old twitcher mice improves survival and is enhanced by bone marrow transplant. J. Neurosci. Res. 2016, 94, 1138–1151. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, A.M.; Rafi, M.A.; Bagel, J.H.; Brisson, B.K.; Marshall, M.S.; Pesayco Salvador, J.; Jiang, X.; Swain, G.P.; Prociuk, M.L.; ODonnell, P.A. AAVrh10 gene therapy ameliorates central and peripheral nervous system disease in canine globoid cell leukodystrophy (Krabbe disease). Hum. Gene Ther. 2018, 29, 785–801. [Google Scholar] [CrossRef]
- Bradbury, A.M.; Bagel, J.H.; Nguyen, D.; Lykken, E.A.; Salvador, J.P.; Jiang, X.; Swain, G.P.; Assenmacher, C.A.; Hendricks, I.J.; Miyadera, K. Krabbe disease successfully treated via monotherapy of intrathecal gene therapy. J. Clin. Investig. 2020, 130, 4906–4920. [Google Scholar] [CrossRef] [PubMed]
- Hori, T.; Fukutome, M.; Maejima, C.; Matsushima, H.; Kobayashi, K.; Kitazawa, S.; Kitahara, R.; Kitano, K.; Kobayashi, K.; Moritoh, S.; et al. Gene delivery to cone photoreceptors by subretinal injection of rAAV2/6 in the mouse retina. Biochem. Biophys. Res. Commun. 2019, 515, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Le Meur, G.; Lebranchu, P.; Billaud, F.; Adjali, O.; Schmitt, S.; Bézieau, S.; Péréon, Y.; Valabregue, R.; Ivan, C.; Darmon, C.; et al. Safety and Long-Term Efficacy of AAV4 Gene Therapy in Patients with RPE65 Leber Congenital Amaurosis. Mol. Ther. J. Am. Soc. Gene Ther. 2018, 26, 256–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Meur, G.; Lebranchu, P.; Pereon, Y.; Billaud, F.; Ivan, C.; Chauveau, C.; Rolling, F.; Weber, M. Efficacy and safety of gene therapy with AAV4 in childhood blindness due to rpe65 mutations. Acta Ophthalmol. 2012, 90. [Google Scholar] [CrossRef]
- Trapani, I.; Auricchio, A. Seeing the Light after 25 Years of Retinal Gene Therapy. Trends Mol. Med. 2018, 24, 669–681. [Google Scholar] [CrossRef]
- MacLaren, R.E.; Groppe, M.; Barnard, A.R.; Cottriall, C.L.; Tolmachova, T.; Seymour, L.; Clark, K.R.; During, M.J.; Cremers, F.P.M.; Black, G.C.M.; et al. Retinal gene therapy in patients with choroideremia: Initial findings from a phase 1/2 clinical trial. Lancet 2014, 383, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Faynus, M.A.; Clegg, D.O. Chapter 7—Modeling inherited retinal dystrophies using induced pluripotent stem cells. In Current Progress in iPSC Disease Modeling; Birbrair, A., Ed.; Academic Press: Cambridge, MS, USA, 2022; pp. 157–184. [Google Scholar] [CrossRef]
- Russell, S.; Bennett, J.; Wellman, J.A.; Chung, D.C.; Yu, Z.-F.; Tillman, A.; Wittes, J.; Pappas, J.; Elci, O.; McCague, S.; et al. Efficacy and safety of voretigene neparvovec (AAV2-hRPE65v2) in patients with RPE65-mediated inherited retinal dystrophy: A randomised, controlled, open-label, phase 3 trial. Lancet 2017, 390, 849–860. [Google Scholar] [CrossRef]
- Bouquet, C.; Vignal Clermont, C.; Galy, A.; Fitoussi, S.; Blouin, L.; Munk, M.R.; Valero, S.; Meunier, S.; Katz, B.; Sahel, J.A.; et al. Immune Response and Intraocular Inflammation in Patients With Leber Hereditary Optic Neuropathy Treated With Intravitreal Injection of Recombinant Adeno-Associated Virus 2 Carrying the ND4 Gene: A Secondary Analysis of a Phase 1/2 Clinical Trial. JAMA Ophthalmol. 2019, 137, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Feuer, W.J.; Schiffman, J.C.; Davis, J.L.; Porciatti, V.; Gonzalez, P.; Koilkonda, R.D.; Yuan, H.; Lalwani, A.; Lam, B.L.; Guy, J. Gene Therapy for Leber Hereditary Optic Neuropathy: Initial Results. Ophthalmology 2016, 123, 558–570. [Google Scholar] [CrossRef] [Green Version]
- Cehajic-Kapetanovic, J.; Xue, K.; Martinez-Fernandez de la Camara, C.; Nanda, A.; Davies, A.; Wood, L.J.; Salvetti, A.P.; Fischer, M.D.; Aylward, J.W.; Barnard, A.R.; et al. Initial results from a first-in-human gene therapy trial on X-linked retinitis pigmentosa caused by mutations in RPGR. Nat. Med. 2020, 26, 354–359. [Google Scholar] [CrossRef]
- Cukras, C.; Wiley, H.E.; Jeffrey, B.G.; Sen, H.N.; Turriff, A.; Zeng, Y.; Vijayasarathy, C.; Marangoni, D.; Ziccardi, L.; Kjellstrom, S.; et al. Retinal AAV8-RS1 Gene Therapy for X-Linked Retinoschisis: Initial Findings from a Phase I/IIa Trial by Intravitreal Delivery. Mol. Ther. 2018, 26, 2282–2294. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Vijayasarathy, C.; Cukras, C.A.; Wiley, H.E.; Sen, H.N.; Zeng, Y.; Wei, L.L.; Sieving, P.A. Immune function in X-linked retinoschisis subjects in an AAV8-RS1 phase I/IIa gene therapy trial. Mol. Ther. 2021, 29, 2030–2040. [Google Scholar] [CrossRef]
- Fischer, M.D.; Michalakis, S.; Wilhelm, B.; Zobor, D.; Muehlfriedel, R.; Kohl, S.; Weisschuh, N.; Ochakovski, G.A.; Klein, R.; Schoen, C.; et al. Safety and Vision Outcomes of Subretinal Gene Therapy Targeting Cone Photoreceptors in Achromatopsia: A Nonrandomized Controlled Trial. JAMA Ophthalmol. 2020, 138, 643–651. [Google Scholar] [CrossRef]
- Kahle, N.A.; Peters, T.; Zobor, D.; Kuehlewein, L.; Kohl, S.; Zhour, A.; Werner, A.; Seitz, I.P.; Sothilingam, V.; Michalakis, S.; et al. Development of Methodology and Study Protocol: Safety and Efficacy of a Single Subretinal Injection of rAAV.hCNGA3 in Patients with CNGA3-Linked Achromatopsia Investigated in an Exploratory Dose-Escalation Trial. Hum. Gene Ther. Clin. Dev. 2018, 29, 121–131. [Google Scholar] [CrossRef]
- Lalwani, A.K.; Walsh, B.J.; Reilly, P.G.; Muzyczka, N.; Mhatre, A.N. Development of in vivo gene therapy for hearing disorders: Introduction of adeno-associated virus into the cochlea of the guinea pig. Gene Ther. 1996, 3, 588–592. [Google Scholar]
- Sondhi, D.; Stiles, K.M.; De, B.P.; Crystal, R.G. Genetic Modification of the Lung Directed Toward Treatment of Human Disease. Hum. Gene Ther. 2017, 28, 3–84. [Google Scholar] [CrossRef]
- McLachlan, G.; Alton, E.; Boyd, A.C.; Clarke, N.K.; Davies, J.C.; Gill, D.R.; Griesenbach, U.; Hickmott, J.W.; Hyde, S.C.; Miah, K.M.; et al. Progress in Respiratory Gene Therapy. Hum. Gene Ther. 2022, 33, 893–912. [Google Scholar] [CrossRef]
- Burney, T.J.; Davies, J.C. Gene therapy for the treatment of cystic fibrosis. Appl. Clin. Genet. 2012, 5, 29. [Google Scholar]
- Flotte, T.R.; Afione, S.A.; Conrad, C.; McGrath, S.A.; Solow, R.; Oka, H.; Zeitlin, P.L.; Guggino, W.B.; Carter, B.J. Stable in vivo expression of the cystic fibrosis transmembrane conductance regulator with an adeno-associated virus vector. Proc. Natl. Acad. Sci. USA 1993, 90, 10613–10617. [Google Scholar] [CrossRef] [Green Version]
- Guggino, W.B.; Cebotaru, L. Gene therapy for cystic fibrosis paved the way for the use of adeno-associated virus in gene therapy. Hum. Gene Ther. 2020, 31, 538–541. [Google Scholar] [CrossRef]
- Wagner, J.A.; Reynolds, T.; Moran, M.L.; Moss, R.B.; Wine, J.J.; Flotte, T.R.; Gardner, P. Efficient and persistent gene transfer of AAV-CFTR in maxillary sinus. Lancet 1998, 351, 1702–1703. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.A.; Nepomuceno, I.B.; Messner, A.H.; Moran, M.L.; Batson, E.P.; Dimiceli, S.; Brown, B.W.; Desch, J.K.; Norbash, A.M.; Conrad, C.K. A phase II, double-blind, randomized, placebo-controlled clinical trial of tgAAVCF using maxillary sinus delivery in patients with cystic fibrosis with antrostomies. Hum. Gene Ther. 2002, 13, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Aitken, M.; Moss, R.; Waltz, D.; Dovey, M.; Tonelli, M.; McNamara, S.; Gibson, R.; Ramsey, B.; Carter, B.; Reynolds, T. A phase I study of aerosolized administration of tgAAVCF to cystic fibrosis subjects with mild lung disease. Hum. Gene Ther. 2001, 12, 1907–1916. [Google Scholar] [CrossRef] [PubMed]
- Flotte, T.R.; Zeitlin, P.L.; Reynolds, T.C.; Heald, A.E.; Pedersen, P.; Beck, S.; Conrad, C.K.; Brass-Ernst, L.; Humphries, M.; Sullivan, K. Phase I trial of intranasal and endobronchial administration of a recombinant adeno-associated virus serotype 2 (rAAV2)-CFTR vector in adult cystic fibrosis patients: A two-part clinical study. Hum. Gene Ther. 2003, 14, 1079–1088. [Google Scholar] [CrossRef]
- Moss, R.B.; Rodman, D.; Spencer, L.T.; Aitken, M.L.; Zeitlin, P.L.; Waltz, D.; Milla, C.; Brody, A.S.; Clancy, J.P.; Ramsey, B. Repeated adeno-associated virus serotype 2 aerosol-mediated cystic fibrosis transmembrane regulator gene transfer to the lungs of patients with cystic fibrosis: A multicenter, double-blind, placebo-controlled trial. Chest 2004, 125, 509–521. [Google Scholar] [CrossRef] [Green Version]
- Moss, R.B.; Milla, C.; Colombo, J.; Accurso, F.; Zeitlin, P.L.; Clancy, J.P.; Spencer, L.T.; Pilewski, J.; Waltz, D.A.; Dorkin, H.L. Repeated aerosolized AAV-CFTR for treatment of cystic fibrosis: A randomized placebo-controlled phase 2B trial. Hum. Gene Ther. 2007, 18, 726–732. [Google Scholar] [CrossRef]
- Kurosaki, F.; Uchibori, R.; Mato, N.; Sehara, Y.; Saga, Y.; Urabe, M.; Mizukami, H.; Sugiyama, Y.; Kume, A. Optimization of adeno-associated virus vector-mediated gene transfer to the respiratory tract. Gene Ther. 2017, 24, 290–297. [Google Scholar] [CrossRef]
- Cooney, A.L.; McCray, P.B.; Sinn, P.L. Cystic Fibrosis Gene Therapy: Looking Back, Looking Forward. Genes 2018, 9, 538. [Google Scholar] [CrossRef] [Green Version]
- Stiles, K.M.; Sondhi, D.; Kaminsky, S.M.; De, B.P.; Rosenberg, J.B.; Crystal, R.G. Intrapleural Gene Therapy for Alpha-1 Antitrypsin Deficiency-Related Lung Disease. Chronic Obstr. Pulm. Dis. 2018, 5, 244–257. [Google Scholar] [CrossRef]
- Song, S.; Embury, J.; Laipis, P.; Berns, K.; Crawford, J.; Flotte, T. Stable therapeutic serum levels of human alpha-1 antitrypsin (AAT) after portal vein injection of recombinant adeno-associated virus (rAAV) vectors. Gene Ther. 2001, 8, 1299–1306. [Google Scholar] [CrossRef] [Green Version]
- Conlon, T.J.; Cossette, T.; Erger, K.; Choi, Y.-K.; Clarke, T.; Scott-Jorgensen, M.; Song, S.; Campbell-Thompson, M.; Crawford, J.; Flotte, T.R. Efficient Hepatic Delivery and Expression from a Recombinant Adeno-associated Virus 8 Pseudotyped α1-Antitrypsin Vector. Mol. Ther. 2005, 12, 867–875. [Google Scholar] [CrossRef]
- De, B.; Heguy, A.; Leopold, P.L.; Wasif, N.; Korst, R.J.; Hackett, N.R.; Crystal, R.G. Intrapleural administration of a serotype 5 adeno-associated virus coding for α1-antitrypsin mediates persistent, high lung and serum levels of α1-antitrypsin. Mol. Ther. 2004, 10, 1003–1010. [Google Scholar] [CrossRef]
- Chulay, J.D.; Ye, G.J.; Thomas, D.L.; Knop, D.R.; Benson, J.M.; Hutt, J.A.; Wang, G.; Humphries, M.; Flotte, T.R. Preclinical evaluation of a recombinant adeno-associated virus vector expressing human alpha-1 antitrypsin made using a recombinant herpes simplex virus production method. Hum. Gene Ther. 2011, 22, 155–165. [Google Scholar] [CrossRef]
- Limberis, M.P.; Vandenberghe, L.H.; Zhang, L.; Pickles, R.J.; Wilson, J.M. Transduction Efficiencies of Novel AAV Vectors in Mouse Airway Epithelium In Vivo and Human Ciliated Airway Epithelium In Vitro. Mol. Ther. 2009, 17, 294–301. [Google Scholar] [CrossRef]
- Liqun Wang, R.; McLaughlin, T.; Cossette, T.; Tang, Q.; Foust, K.; Campbell-Thompson, M.; Martino, A.; Cruz, P.; Loiler, S.; Mueller, C.; et al. Recombinant AAV Serotype and Capsid Mutant Comparison for Pulmonary Gene Transfer of α-1-Antitrypsin Using Invasive and Noninvasive Delivery. Mol. Ther. 2009, 17, 81–87. [Google Scholar] [CrossRef]
- Chiuchiolo, M.J.; Kaminsky, S.M.; Sondhi, D.; Hackett, N.R.; Rosenberg, J.B.; Frenk, E.Z.; Hwang, Y.; Van de Graaf, B.G.; Hutt, J.A.; Wang, G.; et al. Intrapleural administration of an AAVrh.10 vector coding for human α1-antitrypsin for the treatment of α1-antitrypsin deficiency. Hum. Gene Ther. Clin. Dev. 2013, 24, 161–173. [Google Scholar] [CrossRef] [Green Version]
- De, B.P.; Heguy, A.; Hackett, N.R.; Ferris, B.; Leopold, P.L.; Lee, J.; Pierre, L.; Gao, G.; Wilson, J.M.; Crystal, R.G. High Levels of Persistent Expression of α1-Antitrypsin Mediated by the Nonhuman Primate Serotype rh.10 Adeno-associated Virus Despite Preexisting Immunity to Common Human Adeno-associated Viruses. Mol. Ther. 2006, 13, 67–76. [Google Scholar] [CrossRef]
- Flotte, T.R.; Conlon, T.J.; Poirier, A.; Campbell-Thompson, M.; Byrne, B.J. Preclinical characterization of a recombinant adeno-associated virus type 1-pseudotyped vector demonstrates dose-dependent injection site inflammation and dissemination of vector genomes to distant sites. Hum. Gene Ther. 2007, 18, 245–256. [Google Scholar] [CrossRef]
- Zavorotinskaya, T.; Tomkinson, A.; Murphy, J.E. Treatment of experimental asthma by long-term gene therapy directed against IL-4 and IL-13. Mol. Ther. 2003, 7, 155–162. [Google Scholar] [CrossRef]
- Kang, M.H.; van Lieshout, L.P.; Xu, L.; Domm, J.M.; Vadivel, A.; Renesme, L.; Mühlfeld, C.; Hurskainen, M.; Mižíková, I.; Pei, Y. A lung tropic AAV vector improves survival in a mouse model of surfactant B deficiency. Nat. Commun. 2020, 11, 3929. [Google Scholar] [CrossRef]
- Cooney, A.L.; Wambach, J.A.; Sinn, P.L.; McCray, P.B., Jr. Gene therapy potential for genetic disorders of surfactant dysfunction. Front. Genome Ed. 2021, 3, 43. [Google Scholar] [CrossRef] [PubMed]
- Guse, K.; Suzuki, M.; Sule, G.; Bertin, T.K.; Tyynismaa, H.; Ahola-Erkkilä, S.; Palmer, D.; Suomalainen, A.; Ng, P.; Cerullo, V.; et al. Capsid-modified adenoviral vectors for improved muscle-directed gene therapy. Hum. Gene Ther. 2012, 23, 1065–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Cummins, J.; Huard, J.; Wang, B. AAV-directed muscular dystrophy gene therapy. Expert Opin. Biol. Ther. 2010, 10, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Kota, J.; Handy, C.R.; Haidet, A.M.; Montgomery, C.L.; Eagle, A.; Rodino-Klapac, L.R.; Tucker, D.; Shilling, C.J.; Therlfall, W.R.; Walker, C.M.; et al. Follistatin gene delivery enhances muscle growth and strength in nonhuman primates. Sci. Transl. Med. 2009, 1, 6ra15. [Google Scholar] [CrossRef] [Green Version]
- Gregorevic, P.; Allen, J.M.; Minami, E.; Blankinship, M.J.; Haraguchi, M.; Meuse, L.; Finn, E.; Adams, M.E.; Froehner, S.C.; Murry, C.E.; et al. rAAV6-microdystrophin preserves muscle function and extends lifespan in severely dystrophic mice. Nat. Med. 2006, 12, 787–789. [Google Scholar] [CrossRef]
- Wang, B.; Li, J.; Fu, F.H.; Xiao, X. Systemic human minidystrophin gene transfer improves functions and life span of dystrophin and dystrophin/utrophin-deficient mice. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2009, 27, 421–426. [Google Scholar] [CrossRef]
- Zhu, T.; Zhou, L.; Mori, S.; Wang, Z.; McTiernan, C.F.; Qiao, C.; Chen, C.; Wang, D.W.; Li, J.; Xiao, X. Sustained Whole-Body Functional Rescue in Congestive Heart Failure and Muscular Dystrophy Hamsters by Systemic Gene Transfer. Circulation 2005, 112, 2650–2659. [Google Scholar] [CrossRef] [Green Version]
- Pacak, C.A.; Walter, G.A.; Gaidosh, G.; Bryant, N.; Lewis, M.A.; Germain, S.; Mah, C.S.; Campbell, K.P.; Byrne, B.J. Long-term skeletal muscle protection after gene transfer in a mouse model of LGMD-2D. Mol. Ther. J. Am. Soc. Gene Ther. 2007, 15, 1775–1781. [Google Scholar] [CrossRef]
- Fougerousse, F.; Bartoli, M.; Poupiot, J.; Arandel, L.; Durand, M.; Guerchet, N.; Gicquel, E.; Danos, O.; Richard, I. Phenotypic Correction of α-Sarcoglycan Deficiency by Intra-arterial Injection of a Muscle-specific Serotype 1 rAAV Vector. Mol. Ther. J. Am. Soc. Gene Ther. 2007, 15, 53–61. [Google Scholar] [CrossRef]
- Cordier, L.; Hack, A.A.; Scott, M.O.; Barton-Davis, E.R.; Gao, G.; Wilson, J.M.; McNally, E.M.; Sweeney, H.L. Rescue of skeletal muscles of gamma-sarcoglycan-deficient mice with adeno-associated virus-mediated gene transfer. Mol. Ther. J. Am. Soc. Gene Ther. 2000, 1, 119–129. [Google Scholar] [CrossRef]
- Bartoli, M.; Roudaut, C.; Martin, S.; Fougerousse, F.; Suel, L.; Poupiot, J.; Gicquel, E.; Noulet, F.; Danos, O.; Richard, I. Safety and efficacy of AAV-mediated calpain 3 gene transfer in a mouse model of limb-girdle muscular dystrophy type 2A. Mol. Ther. J. Am. Soc. Gene Ther. 2006, 13, 250–259. [Google Scholar] [CrossRef]
- Duan, D. Systemic AAV Micro-dystrophin Gene Therapy for Duchenne Muscular Dystrophy. Mol. Ther. 2018, 26, 2337–2356. [Google Scholar] [CrossRef] [Green Version]
- Bowles, D.E.; McPhee, S.W.J.; Li, C.; Gray, S.J.; Samulski, J.J.; Camp, A.S.; Li, J.; Wang, B.; Monahan, P.E.; Rabinowitz, J.E.; et al. Phase 1 Gene Therapy for Duchenne Muscular Dystrophy Using a Translational Optimized AAV Vector. Mol. Ther. 2012, 20, 443–455. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Cardiovascular Diseases (CVDs). Available online: https://www.who.int/en/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 1 December 2022).
- Jiang, H.; Lillicrap, D.; Patarroyo-White, S.; Liu, T.; Qian, X.; Scallan, C.D.; Powell, S.; Keller, T.; McMurray, M.; Labelle, A.; et al. Multiyear therapeutic benefit of AAV serotypes 2, 6, and 8 delivering factor VIII to hemophilia A mice and dogs. Blood 2006, 108, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Zsebo, K.; Yaroshinsky, A.; Rudy, J.J.; Wagner, K.; Greenberg, B.; Jessup, M.; Hajjar, R.J. Long-Term Effects of AAV1/SERCA2a Gene Transfer in Patients With Severe Heart Failure. Circ. Res. 2014, 114, 101–108. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Ferry, N. Liver gene therapy: Advances and hurdles. Gene Ther. 2004, 11, S76–S84. [Google Scholar] [CrossRef] [Green Version]
- Członkowska, A.; Litwin, T.; Dusek, P.; Ferenci, P.; Lutsenko, S.; Medici, V.; Rybakowski, J.K.; Weiss, K.H.; Schilsky, M.L. Wilson disease. Nat. Rev. Dis. Prim. 2018, 4, 21. [Google Scholar] [CrossRef]
- Murillo, O.; Collantes, M.; Gazquez, C.; Moreno, D.; Hernandez-Alcoceba, R.; Barberia, M.; Ecay, M.; Tamarit, B.; Douar, A.; Ferrer, V.; et al. High value of (64)Cu as a tool to evaluate the restoration of physiological copper excretion after gene therapy in Wilson’s disease. Mol. Ther. Methods Clin. Dev. 2022, 26, 98–106. [Google Scholar] [CrossRef]
- Murillo, O.; Moreno, D.; Gazquez, C.; Barberia, M.; Cenzano, I.; Navarro, I.; Uriarte, I.; Sebastian, V.; Arruebo, M.; Ferrer, V.; et al. Liver Expression of a MiniATP7B Gene Results in Long-Term Restoration of Copper Homeostasis in a Wilson Disease Model in Mice. Hepatology 2019, 70, 108–126. [Google Scholar] [CrossRef]
- Crigler, J.F., Jr.; Najjar, V.A. Congenital familial nonhemolytic jaundice with kernicterus. Pediatrics 1952, 10, 169–180. [Google Scholar]
- Gene Therapy for Severe Crigler Najjar Syndrome. Available online: https://ClinicalTrials.gov/show/NCT03466463 (accessed on 1 December 2022).
- Shi, X.; Aronson, S.J.; Ten Bloemendaal, L.; Duijst, S.; Bakker, R.S.; de Waart, D.R.; Bortolussi, G.; Collaud, F.; Oude Elferink, R.P.; Muro, A.F.; et al. Efficacy of AAV8-hUGT1A1 with Rapamycin in neonatal, suckling, and juvenile rats to model treatment in pediatric CNs patients. Mol. Ther. Methods Clin. Dev. 2021, 20, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Golden, S.H.; Robinson, K.A.; Saldanha, I.; Anton, B.; Ladenson, P.W. Clinical review: Prevalence and incidence of endocrine and metabolic disorders in the United States: A comprehensive review. J. Clin. Endocrinol. Metab. 2009, 94, 1853–1878. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.D. Prospects for research for disorders of the endocrine system. JAMA 2001, 285, 624–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barzon, L.; Bonaguro, R.; Palù, G.; Boscaro, M. New perspectives for gene therapy in endocrinology. Eur. J. Endocrinol. 2000, 143, 447–466. [Google Scholar] [CrossRef] [Green Version]
- Chellappan, D.K.; Sivam, N.S.; Teoh, K.X.; Leong, W.P.; Fui, T.Z.; Chooi, K.; Khoo, N.; Yi, F.J.; Chellian, J.; Cheng, L.L.; et al. Gene therapy and type 1 diabetes mellitus. Biomed. Pharmacother. 2018, 108, 1188–1200. [Google Scholar] [CrossRef]
- Atkinson, M.A. Chapter 32—Type 1 Diabetes Mellitus. In Williams Textbook of Endocrinology, 3rd ed.; Melmed, S., Polonsky, K.S., Larsen, P.R., Kronenberg, H.M., Eds.; Elsevier: Philadelphia, PA, USA, 2016; pp. 1451–1483. [Google Scholar] [CrossRef]
- Flores, R.R.; Zhou, L.; Robbins, P.D. Expression of IL-2 in β cells by AAV8 gene transfer in pre-diabetic NOD mice prevents diabetes through activation of FoxP3-positive regulatory T cells. Gene Ther. 2014, 21, 715–722. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Pileggi, A.; Agarwal, A.; Molano, R.D.; Powers, M.; Brusko, T.; Wasserfall, C.; Goudy, K.; Zahr, E.; Poggioli, R.; et al. Adeno-Associated Virus-Mediated IL-10 Gene Therapy Inhibits Diabetes Recurrence in Syngeneic Islet Cell Transplantation of NOD Mice. Diabetes 2003, 52, 708–716. [Google Scholar] [CrossRef] [Green Version]
- El Khatib, M.; Sakuma, T.; Tonne, J.M.; Mohamed, M.S.; Holditch, S.J.; Lu, B.; Kudva, Y.C.; Ikeda, Y. β-Cell-targeted blockage of PD1 and CTLA4 pathways prevents development of autoimmune diabetes and acute allogeneic islets rejection. Gene Ther. 2015, 22, 430–438. [Google Scholar] [CrossRef] [Green Version]
- RGX-314 Gene Therapy Administered in the Suprachoroidal Space for Participants With Diabetic Retinopathy (DR) without Center Involved-Diabetic Macular Edema (CI-DME). Available online: https://ClinicalTrials.gov/show/NCT04567550 (accessed on 1 December 2022).
- Borden, K.L. Pondering the promyelocytic leukemia protein (PML) puzzle: Possible functions for PML nuclear bodies. Mol. Cell. Biol. 2002, 22, 5259–5269. [Google Scholar] [CrossRef] [Green Version]
- Almaghrabi, S.; Azzouz, M.; Tazi Ahnini, R. AAV9-mediated AIRE gene delivery clears circulating antibodies and tissue T-cell infiltration in a mouse model of autoimmune polyglandular syndrome type-1. Clin. Transl. Immunol. 2020, 9, e1166. [Google Scholar] [CrossRef]
- Luo, J.; Luo, Y.; Sun, J.; Zhou, Y.; Zhang, Y.; Yang, X. Adeno-associated virus-mediated cancer gene therapy: Current status. Cancer Lett. 2015, 356, 347–356. [Google Scholar] [CrossRef] [Green Version]
- WHO. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 1 December 2022).
- Kanazawa, T.; Mizukami, H.; Okada, T.; Hanazono, Y.; Kume, A.; Nishino, H.; Takeuchi, K.; Kitamura, K.; Ichimura, K.; Ozawa, K. Suicide gene therapy using AAV-HSVtk/ganciclovir in combination with irradiation results in regression of human head and neck cancer xenografts in nude mice. Gene Ther. 2003, 10, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Kim, J.H.; Khim, M.; Lee, H.S.; Jung, J.H.; Moon, D.H.; Jeong, S.; Lee, H. Persistent anti-tumor effects via recombinant adeno-associated virus encoding herpes thymidine kinase gene monitored by PET-imaging. Oncol. Rep. 2011, 25, 1263–1269. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, M.; Yoshida, J.; Colosi, P.; Kurtzman, G. Adeno-associated Virus Vector Containing the Herpes Simplex Virus Thymidine Kinase Gene Causes Complete Regression of Intracerebrally Implanted Human Gliomas in Mice, in Conjunction with Ganciclovir Administration. Jpn. J. Cancer Res. 1998, 89, 76–80. [Google Scholar] [CrossRef]
- Kojima, Y.; Honda, K.; Hamada, H.; Kobayashi, N. Oncolytic gene therapy combined with double suicide genes for human bile duct cancer in nude mouse models. J. Surg. Res. 2009, 157, e63–e70. [Google Scholar] [CrossRef]
- Pan, J.G.; Zhou, X.; Luo, R.; Han, R.F. The adeno-associated virus-mediated HSV-TK/GCV suicide system: A potential strategy for the treatment of bladder carcinoma. Med. Oncol. 2012, 29, 1938–1947. [Google Scholar] [CrossRef]
- Borel, F.; Kay, M.A.; Mueller, C. Recombinant AAV as a platform for translating the therapeutic potential of RNA interference. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22, 692–701. [Google Scholar] [CrossRef] [Green Version]
- Sun, A.; Tang, J.; Terranova, P.F.; Zhang, X.; Thrasher, J.B.; Li, B. Adeno-associated virus-delivered short hairpin-structured RNA for androgen receptor gene silencing induces tumor eradication of prostate cancer xenografts in nude mice: A preclinical study. Int. J. Cancer 2010, 126, 764–774. [Google Scholar] [CrossRef]
- Kota, J.; Chivukula, R.R.; O’Donnell, K.A.; Wentzel, E.A.; Montgomery, C.L.; Hwang, H.W.; Chang, T.C.; Vivekanandan, P.; Torbenson, M.; Clark, K.R.; et al. Therapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer model. Cell 2009, 137, 1005–1017. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Luo, S.T.; Shi, H.S.; Li, M.; Zhang, H.L.; He, S.S.; Liu, Y.; Pan, Y.; Yang, L. AAV2-mediated gene transfer of VEGF-Trap with potent suppression of primary breast tumor growth and spontaneous pulmonary metastases by long-term expression. Oncol. Rep. 2012, 28, 1332–1338. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.J.; Gong, C.Y.; Luo, S.T.; Zhang, D.M.; Zhang, S.; Shi, H.S.; Lu, L.; Yan, H.X.; He, S.S.; Li, D.D.; et al. AAV-mediated human PEDF inhibits tumor growth and metastasis in murine colorectal peritoneal carcinomatosis model. BMC Cancer 2012, 12, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tse, L.Y.; Sun, X.; Jiang, H.; Dong, X.; Fung, P.W.; Farzaneh, F.; Xu, R. Adeno-associated virus-mediated expression of kallistatin suppresses local and remote hepatocellular carcinomas. J. Gene Med. 2008, 10, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Hunter, J.E.; Ramos, L.; Wolfe, J.H. Viral Vectors in the CNS. In Reference Module in Neuroscience and Biobehavioral Psychology; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Hermonat, P.L.; Quirk, J.G.; Bishop, B.M.; Han, L. The packaging capacity of adeno-associated virus (AAV) and the potential for wild-type-plus AAV gene therapy vectors. FEBS Lett. 1997, 407, 78–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flotte, T.R.; Carter, B.J. Adeno-Associated Viral Vectors. In Gene Therapy Technologies, Applications and Regulations; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 1999; pp. 109–125. [Google Scholar] [CrossRef]
- Verdera, H.C.; Kuranda, K.; Mingozzi, F. AAV Vector Immunogenicity in Humans: A Long Journey to Successful Gene Transfer. Mol. Ther. J. Am. Soc. Gene Ther. 2020, 28, 723–746. [Google Scholar] [CrossRef]
- Ertl, H.C.J. Immunogenicity and toxicity of AAV gene therapy. Front. Immunol. 2022, 13, 975803. [Google Scholar] [CrossRef]
- Fitzpatrick, Z.; Leborgne, C.; Barbon, E.; Masat, E.; Ronzitti, G.; van Wittenberghe, L.; Vignaud, A.; Collaud, F.; Charles, S.; Simon Sola, M.; et al. Influence of Pre-existing Anti-capsid Neutralizing and Binding Antibodies on AAV Vector Transduction. Mol. Ther. Methods Clin. Dev. 2018, 9, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Denard, J.; Marolleau, B.; Jenny, C.; Rao, T.N.; Fehling, H.J.; Voit, T.; Svinartchouk, F. C-reactive protein (CRP) is essential for efficient systemic transduction of recombinant adeno-associated virus vector 1 (rAAV-1) and rAAV-6 in mice. J. Virol. 2013, 87, 10784–10791. [Google Scholar] [CrossRef] [Green Version]
- Murphy, S.L.; Li, H.; Mingozzi, F.; Sabatino, D.E.; Hui, D.J.; Edmonson, S.A.; High, K.A. Diverse IgG subclass responses to adeno-associated virus infection and vector administration. J. Med. Virol. 2009, 81, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Mingozzi, F.; Meulenberg, J.J.; Hui, D.J.; Basner-Tschakarjan, E.; Hasbrouck, N.C.; Edmonson, S.A.; Hutnick, N.A.; Betts, M.R.; Kastelein, J.J.; Stroes, E.S.; et al. AAV-1-mediated gene transfer to skeletal muscle in humans results in dose-dependent activation of capsid-specific T cells. Blood 2009, 114, 2077–2086. [Google Scholar] [CrossRef]
- Louis Jeune, V.; Joergensen, J.A.; Hajjar, R.J.; Weber, T. Pre-existing anti-adeno-associated virus antibodies as a challenge in AAV gene therapy. Hum. Gene Ther. Methods 2013, 24, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Calcedo, R.; Bell, P.; Lin, J.; Grant, R.L.; Siegel, D.L.; Wilson, J.M. Impact of pre-existing immunity on gene transfer to nonhuman primate liver with adeno-associated virus 8 vectors. Hum. Gene Ther. 2011, 22, 1389–1401. [Google Scholar] [CrossRef] [Green Version]
- Herzog, R.W.; Biswas, M. Neutralizing the Neutralizers in AAV Gene Therapy. Mol. Ther. 2020, 28, 1741–1742. [Google Scholar] [CrossRef]
- Zengel, J.; Carette, J.E. Structural and cellular biology of adeno-associated virus attachment and entry. Adv. Virus Res. 2020, 106, 39–84. [Google Scholar] [CrossRef]
- Chowdhury, E.A.; Meno-Tetang, G.; Chang, H.Y.; Wu, S.; Huang, H.W.; Jamier, T.; Chandran, J.; Shah, D.K. Current progress and limitations of AAV mediated delivery of protein therapeutic genes and the importance of developing quantitative pharmacokinetic/pharmacodynamic (PK/PD) models. Adv. Drug Deliv. Rev. 2021, 170, 214–237. [Google Scholar] [CrossRef]
- Kang, L.; Jin, S.; Wang, J.; Lv, Z.; Xin, C.; Tan, C.; Zhao, M.; Wang, L.; Liu, J. AAV vectors applied to the treatment of CNS disorders: Clinical status and challenges. J. Control. Release 2023, 355, 458–473. [Google Scholar] [CrossRef]
- Colle, M.A.; Piguet, F.; Bertrand, L.; Raoul, S.; Bieche, I.; Dubreil, L.; Sloothaak, D.; Bouquet, C.; Moullier, P.; Aubourg, P.; et al. Efficient intracerebral delivery of AAV5 vector encoding human ARSA in non-human primate. Hum. Mol. Genet. 2010, 19, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Zerah, M.; Piguet, F.; Colle, M.A.; Raoul, S.; Deschamps, J.Y.; Deniaud, J.; Gautier, B.; Toulgoat, F.; Bieche, I.; Laurendeau, I.; et al. Intracerebral Gene Therapy Using AAVrh.10-hARSA Recombinant Vector to Treat Patients with Early-Onset Forms of Metachromatic Leukodystrophy: Preclinical Feasibility and Safety Assessments in Nonhuman Primates. Hum. Gene Ther. Clin. Dev. 2015, 26, 113–124. [Google Scholar] [CrossRef]
- Zieger, M.; Borel, F.; Greer, C.; Gernoux, G.; Blackwood, M.; Flotte, T.R.; Mueller, C. Liver-directed SERPINA1 gene therapy attenuates progression of spontaneous and tobacco smoke-induced emphysema in α1-antitrypsin null mice. Mol. Ther. Methods Clin. Dev. 2022, 25, 425–438. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Issa, S.S.; Shaimardanova, A.A.; Solovyeva, V.V.; Rizvanov, A.A. Various AAV Serotypes and Their Applications in Gene Therapy: An Overview. Cells 2023, 12, 785. https://doi.org/10.3390/cells12050785
Issa SS, Shaimardanova AA, Solovyeva VV, Rizvanov AA. Various AAV Serotypes and Their Applications in Gene Therapy: An Overview. Cells. 2023; 12(5):785. https://doi.org/10.3390/cells12050785
Chicago/Turabian StyleIssa, Shaza S., Alisa A. Shaimardanova, Valeriya V. Solovyeva, and Albert A. Rizvanov. 2023. "Various AAV Serotypes and Their Applications in Gene Therapy: An Overview" Cells 12, no. 5: 785. https://doi.org/10.3390/cells12050785