
Diallyl Trisulfide, a Biologically Active Component of Garlic Essential Oil, Decreases Male Fertility in Sitotroga cerealella by Impairing Dimorphic Spermatogenesis, Sperm Motility and Lipid Homeostasis

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tested Insects and DAT Fumigation
2.2. Transmission Electron Microscopy of Testes
2.3. Determination of Lipid Droplets
2.4. Morphological Observation and Quantity Statistics of Sperm
2.5. Sperm-Survival Assay
2.6. Determination of Total Sperm Motility
2.7. Motile-Sperm-Image Acquisition, Processing and Measurement
2.8. Determination of Adenosine Triphosphate (ATP) Contents in Sperm
2.9. Measurement of Triacylglycerol and Cholesterol Contents
2.10. Statistical Analysis
3. Results
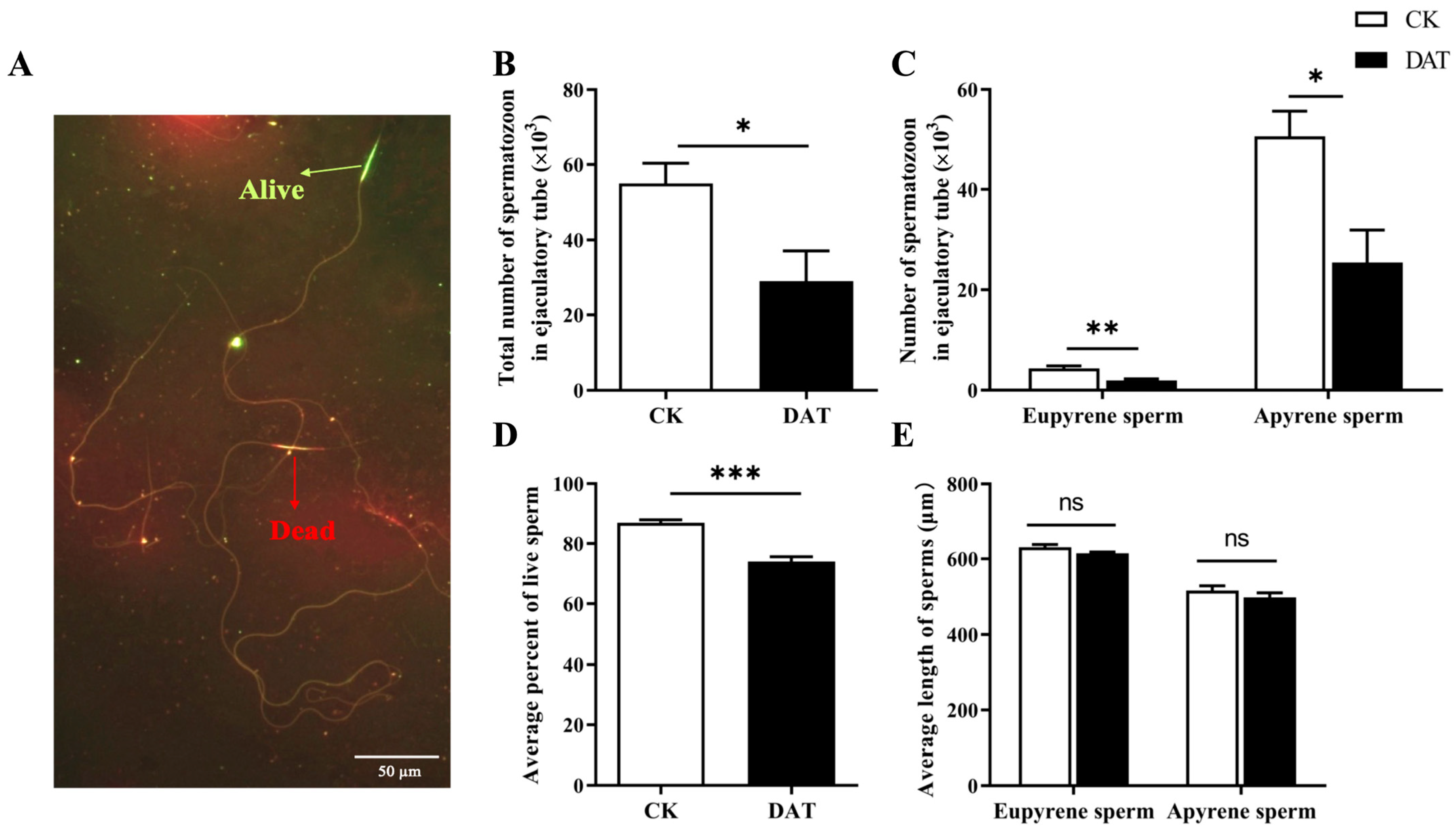
3.1. DAT Reduced the Number and Survival of S. cerealella Spermatozoa
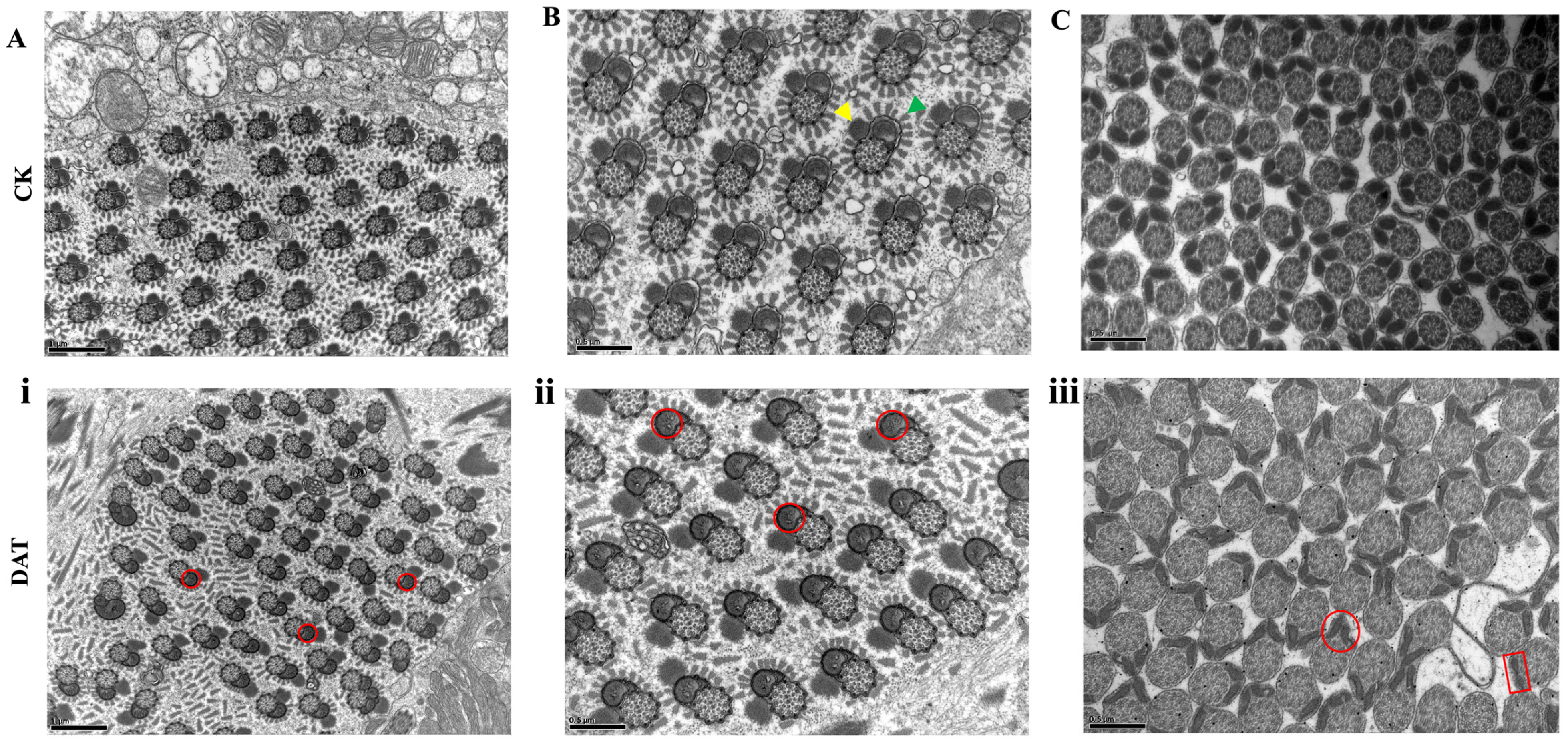
3.2. DAT Altered Ultrastructure of Apyrene and Eupyrene Sperm of S. cerealella
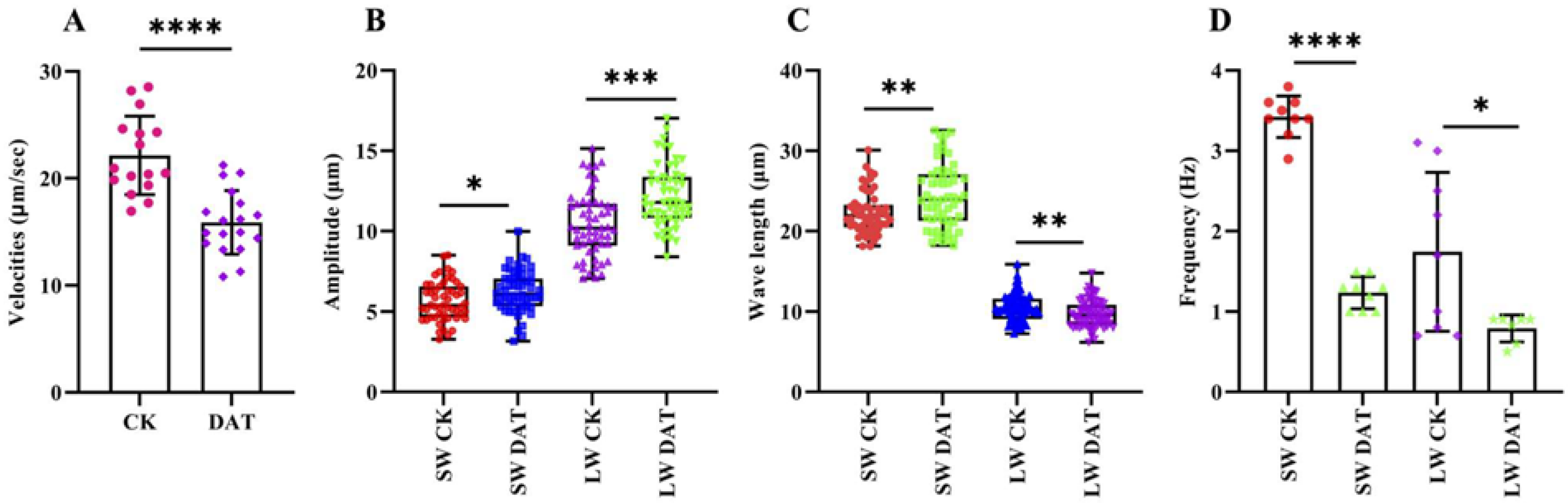
3.3. DAT Decreased the Sperm Motility of S. cerealella
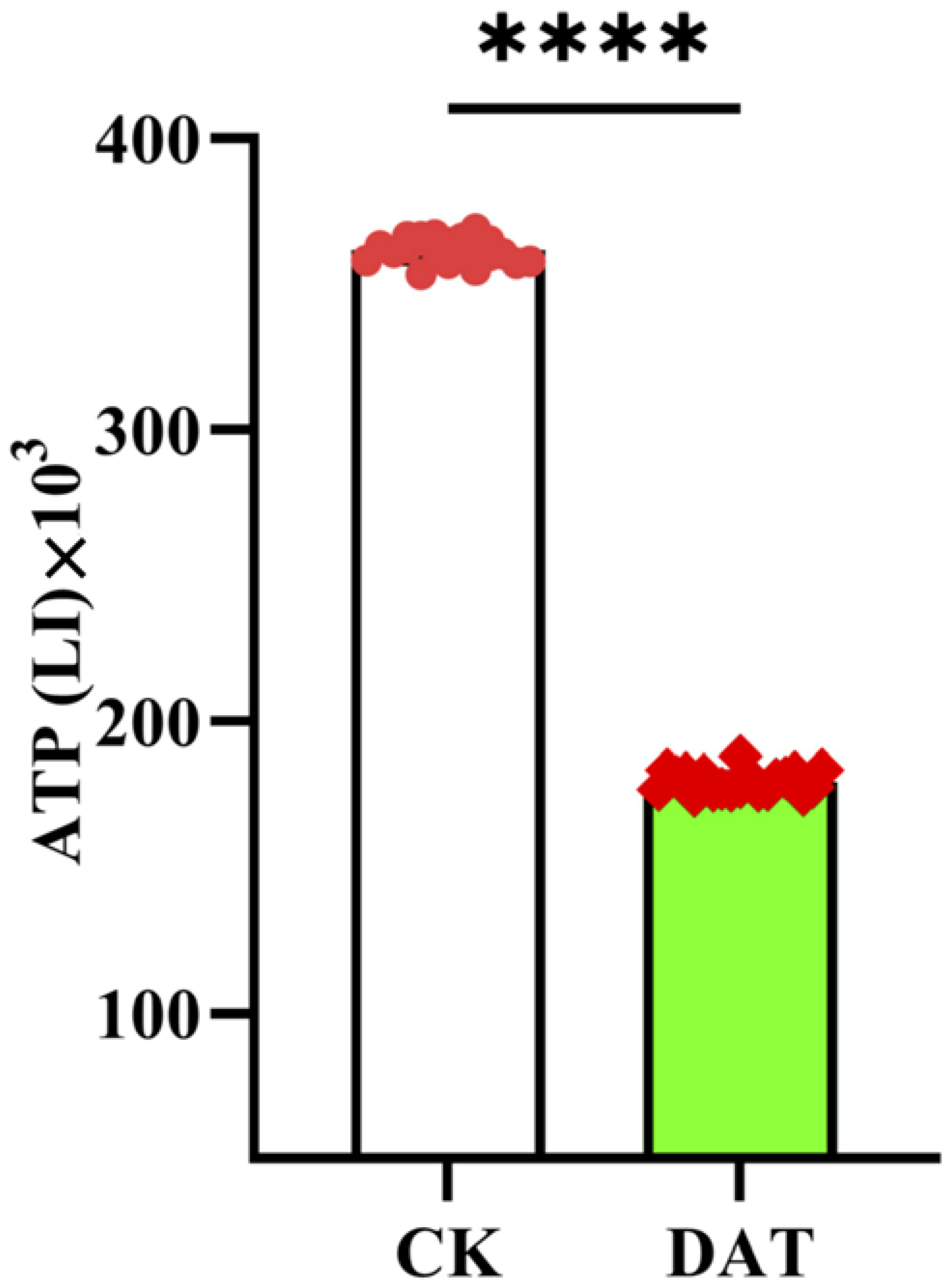
3.4. DAT Decreased Sperm ATP Contents of S. cerealella
3.5. DAT Decreased Lipid Homeostasis of S. cerealella
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ilić, D.P.; Nikolić, V.D.; Nikolić, L.B.; Stanković, M.Z.; Stanojević, L.P.; Cakić, M.D. Allicin and related compounds: Biosynthesis, synthesis and pharmacological activity. Facta Univ. Phys. Chem. Technol. 2011, 9, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Thuy, B.T.P.; My, T.T.A.; Hai, N.T.T.; Hieu, L.T.; Hoa, T.T.; Thi Phuong Loan, H.; Triet, N.T.; Van Anh, T.T.; Quy, P.T.; Van Tat, P. Investigation into SARS-CoV-2 resistance of compounds in garlic essential oil. ACS Omega 2020, 5, 8312–8320. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Hosono, T.; Hosono-Fukao, T.; Inada, K.; Tanaka, R.; Ogihara, J.; Ariga, T. Anticancer effects of diallyl trisulfide derived from garlic. Asia Pac. J. Clin. Nutr. 2008, 17, 249–252. [Google Scholar]
- Predmore, B.L.; Kondo, K.; Bhushan, S.; Zlatopolsky, M.A.; King, A.L.; Aragon, J.P.; Grinsfelder, D.B.; Condit, M.E.; Lefer, D.J. The polysulfide diallyl trisulfide protects the ischemic myocardium by preservation of endogenous hydrogen sulfide and increasing nitric oxide bioavailability. Am. J. Physiol. Circ. Physiol. 2012, 302, H2410–H2418. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.M.; Shah, S.; Wu, M.Y.; Zhang, S.S.; Wu, G.; Yang, F.L. Effect of diallyl trisulfide on the reproductive behavior of the grain moth, Sitotroga cerealella (Lepidoptera: Gelechiidae). Insects 2020, 11, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Chen, S.X.; Ho, S.H. Bioactivities of methyl allyl disulfide and diallyl trisulfide from essential oil of garlic to two species of stored-product pests, Sitophilus zeamais (Coleoptera: Curculionidae) and Tribolium castaneum (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2000, 93, 537–543. [Google Scholar] [CrossRef]
- Hodgson, A.N. Paraspermatogenesis in gastropod molluscs. Invertebr. Reprod. Dev. 1997, 31, 31–38. [Google Scholar] [CrossRef]
- Koehler, J.K.; Birky, C.W. An electron microscope study of the dimorphic spermatozoa of Asplanchna (Rotifera). Z. Für Zellforsch. Und Mikrosk. Anat. 1966, 70, 303–321. [Google Scholar] [CrossRef]
- Alberti, G. Double spermatogenesis in Chelicerata. J. Morphol. 2005, 266, 281–297. [Google Scholar] [CrossRef]
- Chen, S.; Liu, Y.; Yang, X.; Liu, Z.; Luo, X.; Xu, J.; Huang, Y. Dysfunction of dimorphic sperm impairs male fertility in the silkworm. Cell Discov. 2020, 6, 60. [Google Scholar] [CrossRef]
- Sakai, H.; Oshima, H.; Yuri, K.; Gotoh, H.; Daimon, T.; Yaginuma, T.; Sahara, K.; Niimi, T. Dimorphic sperm formation by Sex-lethal. Proc. Natl. Acad. Sci. USA 2019, 116, 10412–10417. [Google Scholar] [CrossRef] [Green Version]
- Pitnick, S.; Wolfner, M.F.; Dorus, S. Post-ejaculatory modifications to sperm (PEMS). Biol. Rev. 2020, 95, 365–392. [Google Scholar] [CrossRef]
- Shah, S.; Hafeez, M.; Wu, M.-Y.; Zhang, S.-S.; Ilyas, M.; Wu, G.; Yang, F.-L. Downregulation of chitin synthase A gene by diallyl trisulfide, an active substance from garlic essential oil, inhibits oviposition and alters the morphology of adult Sitotroga cerealella. J. Pest Sci. 2020, 93, 1097–1106. [Google Scholar] [CrossRef]
- Yan, W.; Wu, M.-Y.; Shah, S.; Yao, Y.-C.; Elgizawy, K.K.; Tang, N.; Wu, G.; Yang, F.-L. Silencing the Triacylglycerol Lipase (TGL) Gene Decreases the Number of Apyrene Sperm and Inhibits Oviposition in Sitotroga Cerealella. Cell. Mol. Life Sci. 2021, 79, 44. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Zhang, S.-S.; Elgizawy, K.K.; Yan, W.-H.; Tang, N.; Wu, G.; Yang, F.-L. Diallyl trisulfide reduced the reproductive capacity of male Sitotroga cerealella via the regulation of juvenile and ecdysone hormones. Ecotoxicol. Environ. Saf. 2022, 248, 114304. [Google Scholar] [CrossRef] [PubMed]
- De Los Santos, C.G. Analysis of Transitions in Sperm Motility; University of California: Riverside, CA, USA, 2015; ISBN 1339183234. [Google Scholar]
- Omar, S.H.; Al-Wabel, N.A. Organosulfur compounds and possible mechanism of garlic in cancer. Saudi Pharm. J. 2010, 18, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef] [Green Version]
- Ying, Y.Y. Regulation of Diallyl Trisulfide on Post-Mating Effect of Sitotroga cerealella at Sublethal Concentration. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2019. [Google Scholar] [CrossRef]
- Werner, M.; Simmons, L.W. Insect Sperm Motility. Biol. Rev. Camb. Philos. Soc. 2008, 83, 191–208. [Google Scholar] [CrossRef]
- Tremoen, N.H.; Gaustad, A.H.; Andersen-Ranberg, I.; van Son, M.; Zeremichael, T.T.; Frydenlund, K.; Grindflek, E.; Våge, D.I.; Myromslien, F.D. Relationship between sperm motility characteristics and ATP concentrations, and association with fertility in two different pig breeds. Anim. Reprod. Sci. 2018, 193, 226–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toprak, U.; Hegedus, D.; Doğan, C.; Güney, G. A journey into the world of insect lipid metabolism. Arch. Insect Biochem. Physiol. 2020, 104, e21682. [Google Scholar] [CrossRef]
- Frühbeck, G.; Méndez-Giménez, L.; Fernández-Formoso, J.-A.; Fernández, S.; Rodriguez, A. Regulation of adipocyte lipolysis. Nutr. Res. Rev. 2014, 27, 63–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baer, B.; Eubel, H.; Taylor, N.L.; O’Toole, N.; Millar, A.H. Insights into female sperm storage from the spermathecal fluid proteome of the honeybee Apis mellifera. Genome Biol. 2009, 10, R67. [Google Scholar] [CrossRef] [Green Version]
- Prokupek, A.M.; Kachman, S.D.; Ladunga, I.; Harshman, L.G. Transcriptional profiling of the sperm storage organs of Drosophila melanogaster. Insect Mol. Biol. 2009, 18, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Das, C.C.; Kaufmann, B.P.; Gay, H. Histone-protein transition in Drosophila melanogaster: I. Changes during spermatogenesis. Exp. Cell Res. 1964, 35, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, V.K.; Kumar, K. Effect of the chemosterilant bisazir on the testes of the spotted bollworm Earias fabia Stoll. Toxicology 1984, 31, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Ghost, M.R.; Sahu, C.R. Effects of insecticides on protein histochemistry of the testis of Diacrisia obliqua Walker (Lepidoptera: Arctiidae). Environ. Ecol. 1991, 9, 167–169. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % Motile Sperm | 0% | 20% | 40% | 60% | 80% | 95% |
|---|---|---|---|---|---|---|
| Scale | 0 | 1 | 2 | 3 | 4 | 5 |
| CK | - | - | - | - | 8 | 42 |
| DAT | 10 | 25 | 13 | 2 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, S.; Elgizawy, K.K.; Shi, C.-M.; Yao, H.; Yan, W.-H.; Li, Y.; Wang, X.-P.; Wu, G.; Yang, F.-L. Diallyl Trisulfide, a Biologically Active Component of Garlic Essential Oil, Decreases Male Fertility in Sitotroga cerealella by Impairing Dimorphic Spermatogenesis, Sperm Motility and Lipid Homeostasis. Cells 2023, 12, 669. https://doi.org/10.3390/cells12040669
Shah S, Elgizawy KK, Shi C-M, Yao H, Yan W-H, Li Y, Wang X-P, Wu G, Yang F-L. Diallyl Trisulfide, a Biologically Active Component of Garlic Essential Oil, Decreases Male Fertility in Sitotroga cerealella by Impairing Dimorphic Spermatogenesis, Sperm Motility and Lipid Homeostasis. Cells. 2023; 12(4):669. https://doi.org/10.3390/cells12040669
Chicago/Turabian StyleShah, Sakhawat, Karam Khamis Elgizawy, Chun-Mei Shi, Hucheng Yao, Wen-Han Yan, Yu Li, Xiao-Ping Wang, Gang Wu, and Feng-Lian Yang. 2023. "Diallyl Trisulfide, a Biologically Active Component of Garlic Essential Oil, Decreases Male Fertility in Sitotroga cerealella by Impairing Dimorphic Spermatogenesis, Sperm Motility and Lipid Homeostasis" Cells 12, no. 4: 669. https://doi.org/10.3390/cells12040669