
Androgens and NGF Mediate the Neurite-Outgrowth through Inactivation of RhoA
 , , and
, , and 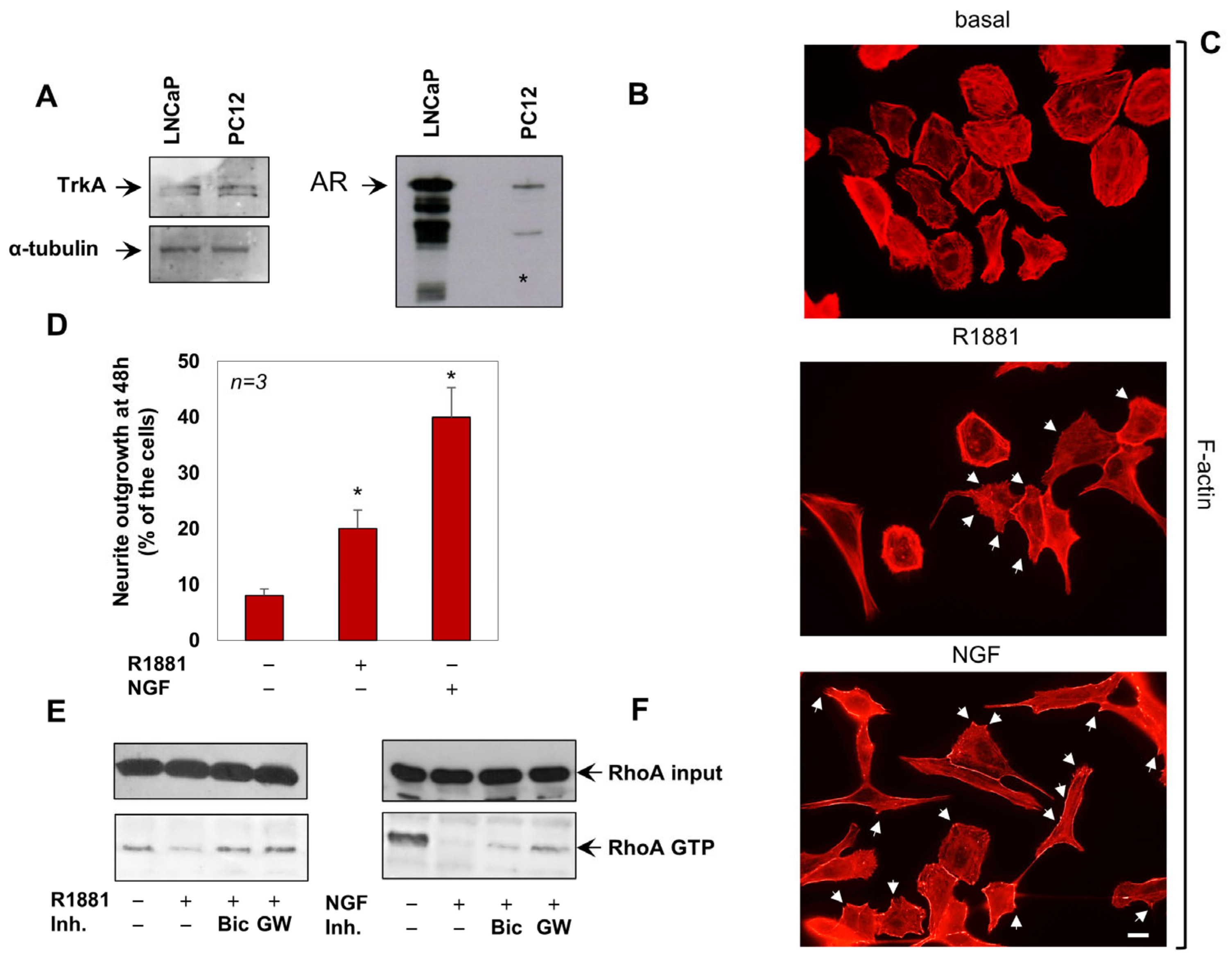{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
2.2. Cell Culture and Transfection
2.3. Primary Mouse Hippocampal Neurons
2.4. Neurite Outgrowth Assay, Contrast Phase and Immunofluorescence (IF) Microscopy
2.5. Lysates, RhoA Pull down Assay and Western Blot
2.6. Statistical Analysis and Data Availability Statement
3. Results
3.1. Androgens and NGF Promote the Neurite-Outgrowth through the RhoA Inactivation
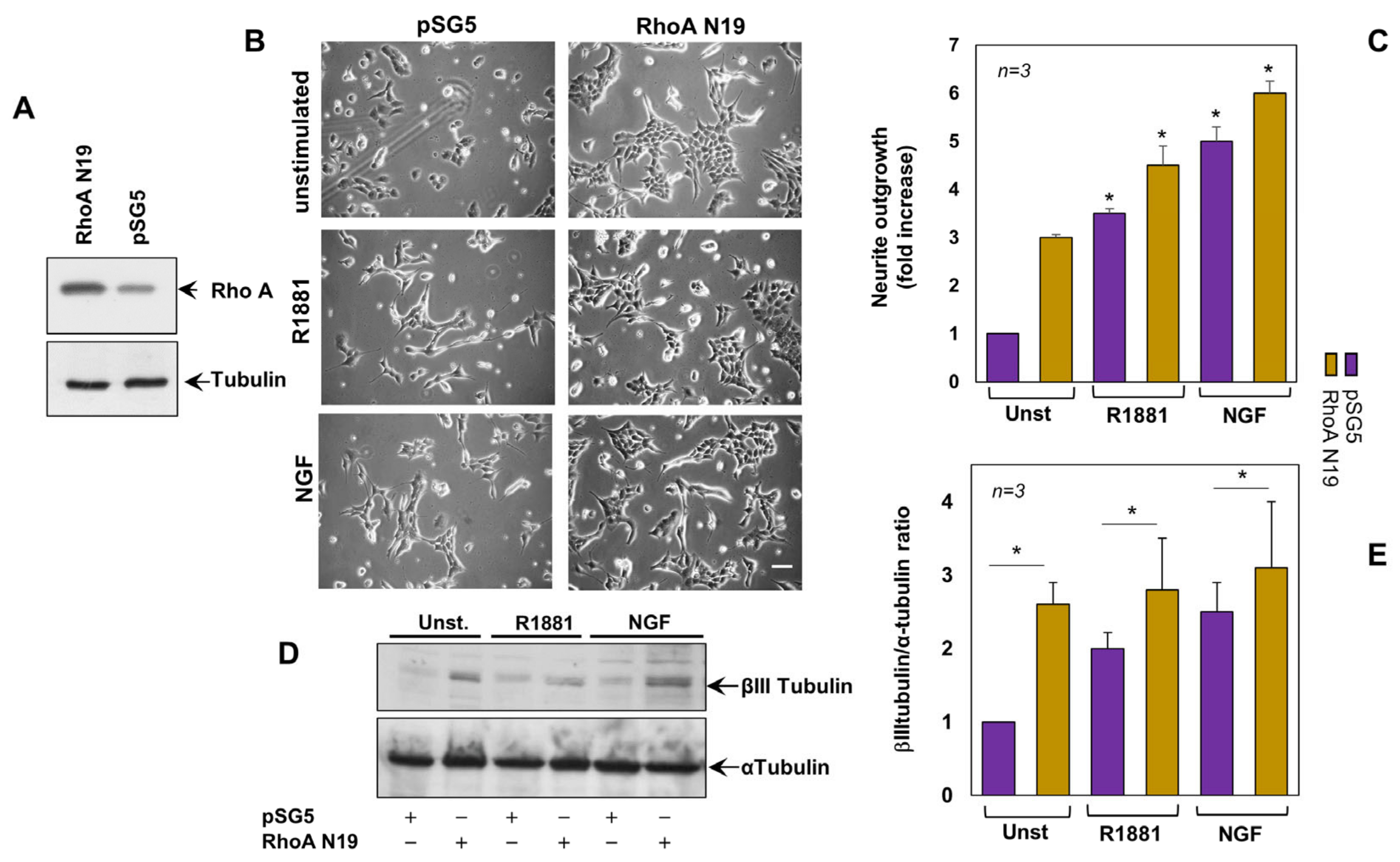
3.2. Inactivation of RhoA Is Implicated in Androgen- and NG- Induced Neurite Outgrowth of PC12 Cells
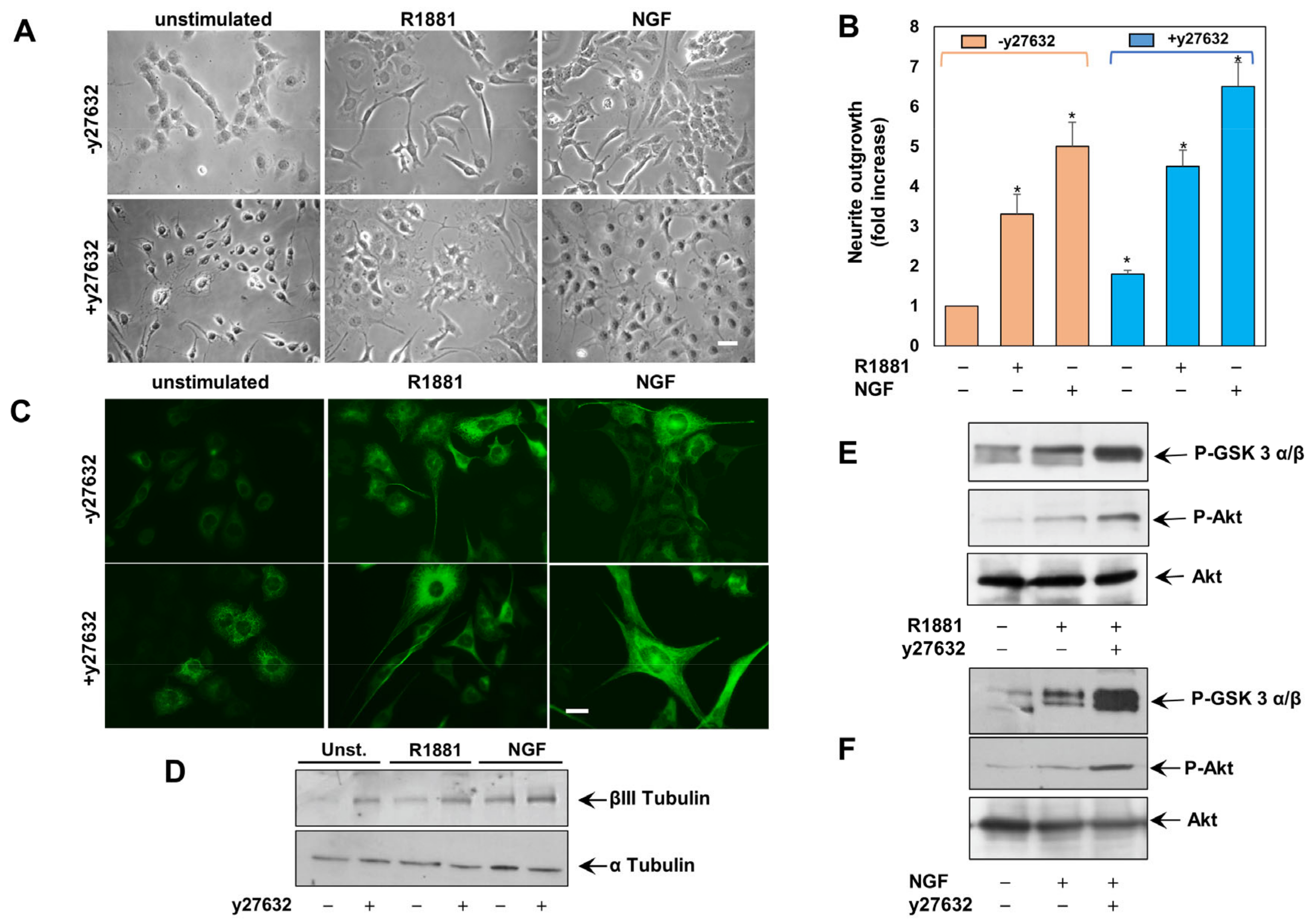
3.3. The Inhibition of ROCK Enhances Neuritogenesis in Androgen- and NGF-Challanged PC12 Cells
3.4. AR, TRKa and ROCK Are Involved in Neuritogenesis of Hippocampal Neurons in Primary Cultures
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vegeto, E.; Villa, A.; Della Torre, S.; Crippa, V.; Rusmini, P.; Cristofani, R.; Galbiati, M.; Maggi, A.; Poletti, A. The Role of Sex and Sex Hormones in Neurodegenerative Diseases. Endocr. Rev. 2019, 41, 273–319. [Google Scholar] [CrossRef] [PubMed]
- García-Ovejero, D.; Veiga, S.; García-Segura, L.M.; Doncarlos, L.L. Glial Expression of Estrogen and Androgen Receptors after Rat Brain Injury. J. Comp. Neurol. 2002, 450, 256–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsen, J.; Chen, S.; Irwin, R.W.; Iwamoto, S.; Brinton, R.D. Estrogen Protects Neuronal Cells from Amyloid Beta-Induced Apoptosis via Regulation of Mitochondrial Proteins and Function. BMC Neurosci. 2006, 7, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pike, C.J. Sex and the Development of Alzheimer’s Disease. J. Neurosci. Res. 2017, 95, 671–680. [Google Scholar] [CrossRef] [Green Version]
- Aloe, L.; Rocco, M.L.; Balzamino, B.O.; Micera, A. Nerve Growth Factor: Role in Growth, Differentiation and Controlling Cancer Cell Development. J. Exp. Clin. Cancer Res. 2016, 35, 116. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, V.E.; Rizzi, L.; Bresciani, E.; Omeljaniuk, R.J.; Torsello, A. Androgen Therapy in Neurodegenerative Diseases. J. Endocr. Soc. 2020, 4, bvaa120. [Google Scholar] [CrossRef]
- Lei, Y.; Renyuan, Z. Effects of Androgens on the Amyloid-β Protein in Alzheimer’s Disease. Endocrinology 2018, 159, 3885–3894. [Google Scholar] [CrossRef]
- Abi-Ghanem, C.; Robison, L.S.; Zuloaga, K.L. Androgens’ Effects on Cerebrovascular Function in Health and Disease. Biol. Sex Differ. 2020, 11, 35. [Google Scholar] [CrossRef]
- Castoria, G.; Auricchio, F.; Migliaccio, A. Extranuclear Partners of Androgen Receptor: At the Crossroads of Proliferation, Migration, and Neuritogenesis. FASEB J. 2017, 31, 1289–1300. [Google Scholar] [CrossRef] [Green Version]
- Cornil, C.A.; Ball, G.F.; Balthazart, J. Functional Significance of the Rapid Regulation of Brain Estrogens: Where Do the Estrogens Come From? Brain Res. 2006, 1126, 2–26. [Google Scholar] [CrossRef]
- Di Donato, M.; Bilancio, A.; D’Amato, L.; Claudiani, P.; Oliviero, M.A.; Barone, M.V.; Auricchio, A.; Appella, E.; Migliaccio, A.; Auricchio, F.; et al. Cross-Talk between Androgen Receptor/Filamin A and TrkA Regulates Neurite Outgrowth in PC12 Cells. Mol. Biol. Cell 2015, 26, 2858–2872. [Google Scholar] [CrossRef] [PubMed]
- La Spada, A.R.; Wilson, E.M.; Lubahn, D.B.; Harding, A.E.; Fischbeck, K.H. Androgen Receptor Gene Mutations in X-Linked Spinal and Bulbar Muscular Atrophy. Nature 1991, 352, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.F.; Forouhan, M.; Roberts, T.C.; Dabney, J.; Ellerington, R.; Speciale, A.A.; Manzano, R.; Lieto, M.; Sangha, G.; Banerjee, S.; et al. Gene Therapy with AR Isoform 2 Rescues Spinal and Bulbar Muscular Atrophy Phenotype by Modulating AR Transcriptional Activity. Sci. Adv. 2021, 7, eabi6896. [Google Scholar] [CrossRef] [PubMed]
- Duong, P.; Tenkorang, M.A.A.; Trieu, J.; McCuiston, C.; Rybalchenko, N.; Cunningham, R.L. Neuroprotective and Neurotoxic Outcomes of Androgens and Estrogens in an Oxidative Stress Environment. Biol. Sex Differ. 2020, 11, 12. [Google Scholar] [CrossRef]
- Garza-Contreras, J.; Duong, P.; Snyder, B.D.; Schreihofer, D.A.; Cunningham, R.L. Presence of Androgen Receptor Variant in Neuronal Lipid Rafts. eNeuro 2017, 4, ENEURO.0109-17.2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foradori, C.D.; Weiser, M.J.; Handa, R.J. Non-Genomic Actions of Androgens. Front. Neuroendocrinol. 2008, 29, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.W.; Lamperti, E.D.; Ekşioğlu, Y.Z.; Hong, S.E.; Feng, Y.; Graham, D.A.; Scheffer, I.E.; Dobyns, W.B.; Hirsch, B.A.; Radtke, R.A.; et al. Mutations in Filamin 1 Prevent Migration of Cerebral Cortical Neurons in Human Periventricular Heterotopia. Neuron 1998, 21, 1315–1325. [Google Scholar] [CrossRef] [Green Version]
- Lian, G.; Lu, J.; Hu, J.; Zhang, J.; Cross, S.H.; Ferland, R.J.; Sheen, V.L. Filamin a Regulates Neural Progenitor Proliferation and Cortical Size through Wee1-Dependent Cdk1 Phosphorylation. J. Neurosci. 2012, 32, 7672–7684. [Google Scholar] [CrossRef] [Green Version]
- Stossel, T.P.; Condeelis, J.; Cooley, L.; Hartwig, J.H.; Noegel, A.; Schleicher, M.; Shapiro, S.S. Filamins as Integrators of Cell Mechanics and Signalling. Nat. Rev. Mol. Cell Biol. 2001, 2, 138–145. [Google Scholar] [CrossRef]
- Burridge, K.; Wennerberg, K. Rho and Rac Take Center Stage. Cell 2004, 116, 167–179. [Google Scholar] [CrossRef]
- Costa, J.F.; Dines, M.; Lamprecht, R. The Role of Rac GTPase in Dendritic Spine Morphogenesis and Memory. Front. Synaptic Neurosci. 2020, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, A.; Minden, A.; Yuste, R. Regulation of Dendritic Spine Morphology by the Rho Family of Small GTPases: Antagonistic Roles of Rac and Rho. Cereb. Cortex 2000, 10, 927–938. [Google Scholar] [CrossRef] [Green Version]
- Cosker, K.E.; Shadan, S.; Van Diepen, M.; Morgan, C.; Li, M.; Allen-Baume, V.; Hobbs, C.; Doherty, P.; Cockcroft, S.; Eickholt, B.J. Regulation of PI3K Signalling by the Phosphatidylinositol Transfer Protein PITPα during Axonal Extension in Hippocampal Neurons. J Cell Sci. 2008, 121, 796–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Palmero, I.; Simon-Areces, J.; Garcia-Segura, L.M.; Arevalo, M.-A. Notch/Neurogenin 3 Signalling Is Involved in the Neuritogenic Actions of Oestradiol in Developing Hippocampal Neurones. J. Neuroendocrinol. 2011, 23, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fath, T.; Ke, Y.D.; Gunning, P.; Götz, J.; Ittner, L.M. Primary Support Cultures of Hippocampal and Substantia Nigra Neurons. Nat. Protoc. 2009, 4, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Kaech, S.; Banker, G. Culturing Hippocampal Neurons. Nat. Protoc. 2006, 1, 2406–2415. [Google Scholar] [CrossRef]
- Bithell, A.; Finch, S.E.; Hornby, M.F.; Williams, B.P. Fibroblast Growth Factor 2 Maintains the Neurogenic Capacity of Embryonic Neural Progenitor Cells in Vitro but Changes Their Neuronal Subtype Specification. Stem Cells 2008, 26, 1565–1574. [Google Scholar] [CrossRef]
- Castoria, G.; Giovannelli, P.; Di Donato, M.; Ciociola, A.; Hayashi, R.; Bernal, F.; Appella, E.; Auricchio, F.; Migliaccio, A. Role of Non-Genomic Androgen Signalling in Suppressing Proliferation of Fibroblasts and Fibrosarcoma Cells. Cell Death Dis. 2014, 5, e1548. [Google Scholar] [CrossRef] [Green Version]
- Di Donato, M.; Zamagni, A.; Galasso, G.; Di Zazzo, E.; Giovannelli, P.; Barone, M.V.; Zanoni, M.; Gunelli, R.; Costantini, M.; Auricchio, F.; et al. The Androgen Receptor/Filamin A Complex as a Target in Prostate Cancer Microenvironment. Cell Death Dis. 2021, 12, 127. [Google Scholar] [CrossRef]
- Di Donato, M.; Giovannelli, P.; Migliaccio, A.; Bilancio, A. Inhibition of Vps34 and P110δ PI3K Impairs Migration, Invasion and Three-Dimensional Spheroid Growth in Breast Cancer Cells. Int. J. Mol. Sci. 2022, 23, 9008. [Google Scholar] [CrossRef]
- Greene, L.A.; Tischler, A.S. Establishment of a Noradrenergic Clonal Line of Rat Adrenal Pheochromocytoma Cells Which Respond to Nerve Growth Factor. Proc. Natl. Acad. Sci. USA 1976, 73, 2424–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, C.J. Specificity of Receptor Tyrosine Kinase Signaling: Transient versus Sustained Extracellular Signal-Regulated Kinase Activation. Cell 1995, 80, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Wood, E.R.; Kuyper, L.; Petrov, K.G.; Hunter, R.N.; Harris, P.A.; Lackey, K. Discovery and in Vitro Evaluation of Potent TrkA Kinase Inhibitors: Oxindole and Aza-Oxindoles. Bioorg. Med. Chem. Lett. 2004, 14, 953–957. [Google Scholar] [CrossRef] [PubMed]
- Di Donato, M.; Cernera, G.; Auricchio, F.; Migliaccio, A.; Castoria, G. Cross-Talk between Androgen Receptor and Nerve Growth Factor Receptor in Prostate Cancer Cells: Implications for a New Therapeutic Approach. Cell Death Discov. 2018, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobes, C.D.; Hall, A. Rho GTPases Control Polarity, Protrusion, and Adhesion during Cell Movement. J Cell Biol. 1999, 144, 1235–1244. [Google Scholar] [CrossRef] [Green Version]
- Moody, S.A.; Quigg, M.S.; Frankfurter, A. Development of the Peripheral Trigeminal System in the Chick Revealed by an Isotype-Specific Anti-Beta-Tubulin Monoclonal Antibody. J. Comp. Neurol. 1989, 279, 567–580. [Google Scholar] [CrossRef]
- Dennis, K.; Uittenbogaard, M.; Chiaramello, A.; Moody, S.A. Cloning and Characterization of the 5’-Flanking Region of the Rat Neuron-Specific Class III Beta-Tubulin Gene. Gene 2002, 294, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Julian, L.; Olson, M.F. Rho-Associated Coiled-Coil Containing Kinases (ROCK). Small GTPases 2014, 5, e29846. [Google Scholar] [CrossRef]
- Narumiya, S.; Tanji, M.; Ishizaki, T. Rho Signaling, ROCK and MDia1, in Transformation, Metastasis and Invasion. Cancer Metastasis Rev. 2009, 28, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Ye, F.; Wang, Y.; Feng, D. ROCK Inhibition Enhances Neurite Outgrowth in Neural Stem Cells by Upregulating YAP Expression in Vitro. Neural Regen. Res. 2016, 11, 983–987. [Google Scholar] [CrossRef]
- Fujita, Y.; Yamashita, T. Axon Growth Inhibition by RhoA/ROCK in the Central Nervous System. Front. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Narumiya, S.; Ishizaki, T.; Ufhata, M. Use and Properties of ROCK-Specific Inhibitor Y-27632. In Meth Enzymol; Regulators and Effectors of Small GTPases—Part D: Rho Family; Balch, W.E., Der, C.J., Hall, A., Eds.; Academic Press: Cambridge, MA, USA, 2000; Volume 325, pp. 273–284. [Google Scholar] [CrossRef]
- Read, D.; Gorman, A. Involvement of Akt in Neurite Outgrowth. Cell. Mol. Life Sci. 2009, 66, 2975–2984. [Google Scholar] [CrossRef]
- Ruiz-Palmero, I.; Ortiz-Rodriguez, A.; Melcangi, R.C.; Caruso, D.; Garcia-Segura, L.M.; Rune, G.M.; Arevalo, M.-A. Oestradiol Synthesized by Female Neurons Generates Sex Differences in Neuritogenesis. Sci. Rep. 2016, 6, 31891. [Google Scholar] [CrossRef] [Green Version]
- Fetler, L.; Amigorena, S. Neuroscience. Brain under Surveillance: The Microglia Patrol. Science 2005, 309, 392–393. [Google Scholar] [CrossRef]
- Argente-Arizón, P.; Guerra-Cantera, S.; Garcia-Segura, L.M.; Argente, J.; Chowen, J.A. Glial Cells and Energy Balance. J. Mol. Endocrinol. 2017, 58, R59–R71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, K.J.; Serrano, A.; Lacaille, J.-C.; Robitaille, R. Glial Cells in Synaptic Plasticity. J. Physiol. Paris 2006, 99, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Silver, J.; Miller, J.H. Regeneration beyond the Glial Scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef] [Green Version]
- Do-Rego, J.-L.; Seong, J.Y.; Burel, D.; LEPRINCE, J.; Vaudry, D.; Luu-The, V.; Tonon, M.-C.; Tsutsui, K.; Pelletier, G.; Vaudry, H. Regulation of Neurosteroid Biosynthesis by Neurotransmitters and Neuropeptides. Front. Endocrinol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, C.G.; Sicard, F.; Lattke, P.; Bornstein, S.R.; Ehrhart-Bornstein, M.; Krug, A.W. Dehydroepiandrosterone Induces a Neuroendocrine Phenotype in Nerve Growth Factor-Stimulated Chromaffin Pheochromocytoma PC12 Cells. Endocrinology 2008, 149, 320–328. [Google Scholar] [CrossRef]
- Charalampopoulos, I.; Alexaki, V.-I.; Lazaridis, I.; Dermitzaki, E.; Avlonitis, N.; Tsatsanis, C.; Calogeropoulou, T.; Margioris, A.N.; Castanas, E.; Gravanis, A. G Protein-Associated, Specific Membrane Binding Sites Mediate the Neuroprotective Effect of Dehydroepiandrosterone. FASEB J. 2006, 20, 577–579. [Google Scholar] [CrossRef]
- Di Donato, M.; Cernera, G.; Migliaccio, A.; Castoria, G. Nerve Growth Factor Induces Proliferation and Aggressiveness in Prostate Cancer Cells. Cancers 2019, 11, 784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukai, H.; Takata, N.; Ishii, H.-t.; Tanabe, N.; Hojo, Y.; Furukawa, A.; Kimoto, T.; Kawato, S. Hippocampal Synthesis of Estrogens and Androgens Which Are Paracrine Modulators of Synaptic Plasticity: Synaptocrinology. Neuroscience 2006, 138, 757–764. [Google Scholar] [CrossRef]
- Lauber, M.E.; Lichtensteiger, W. Ontogeny of 5 Alpha-Reductase (Type 1) Messenger Ribonucleic Acid Expression in Rat Brain: Early Presence in Germinal Zones. Endocrinology 1996, 137, 2718–2730. [Google Scholar] [CrossRef] [Green Version]
- Poletti, A.; Negri-Cesi, P.; Rabuffetti, M.; Colciago, A.; Celotti, F.; Martini, L. Transient Expression of the 5alpha-Reductase Type 2 Isozyme in the Rat Brain in Late Fetal and Early Postnatal Life. Endocrinology 1998, 139, 2171–2178. [Google Scholar] [CrossRef]
- Poletti, A.; Celotti, F.; Rumio, C.; Rabuffetti, M.; Martini, L. Identification of Type 1 5α-Reductase in Myelin Membranes of Male and Female Rat Brain. Mol. Cell. Endocrinol. 1997, 129, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Melcangi, R.C.; Poletti, A.; Cavarretta, I.; Celotti, F.; Colciago, A.; Magnaghi, V.; Motta, M.; Negri-Cesi, P.; Martini, L. The 5a-Reductase in the Central Nervous System: Expression and Modes of Control. J. Steroid Biochem. Mol. Biol. 1998, 65, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.M.; Ortega, E. Differential Regulation of Steroid 5α-Reductase Isozymes Expression by Androgens in the Adult Rat Brain. FASEB J. 2003, 17, 1428–1433. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, D.; Castrén, E.; Berzaghi, M.; Blöchl, A.; Thoenen, H. Activity-Dependent and Hormonal Regulation of Neurotrophin MRNA Levels in the Brain--Implications for Neuronal Plasticity. J. Neurobiol. 1994, 25, 1362–1372. [Google Scholar] [CrossRef]
- Gibbs, R.B. Estrogen and Nerve Growth Factor-Related Systems in Brain. Effects on Basal Forebrain Cholinergic Neurons and Implications for Learning and Memory Processes and Aging. Ann. N. Y. Acad. Sci. 1994, 743, 165–196, discussion 197–199. [Google Scholar] [CrossRef]
- Scully, J.L.; Otten, U. NGF: Not Just for Neurons. Cell Biol. Int. 1995, 19, 459–469. [Google Scholar] [CrossRef]
- Toran-Allerand, C.D. The Estrogen/Neurotrophin Connection during Neural Development: Is Co-Localization of Estrogen Receptors with the Neurotrophins and Their Receptors Biologically Relevant? Dev. Neurosci. 1996, 18, 36–48. [Google Scholar] [CrossRef] [PubMed]
- De Nicola, A.F.; Ferrini, M.; Gonzalez, S.L.; Gonzalez Deniselle, M.C.; Grillo, C.A.; Piroli, G.; Saravia, F.; De Kloet, E.R. Regulation of Gene Expression by Corticoid Hormones in the Brain and Spinal Cord. J. Steroid Biochem. Mol. Biol. 1998, 65, 253–272. [Google Scholar] [CrossRef] [PubMed]
- Baker, L.D.; Asthana, S.; Cholerton, B.A.; Wilkinson, C.W.; Plymate, S.R.; Green, P.S.; Merriam, G.R.; Fishel, M.A.; Watson, G.S.; Cherrier, M.M.; et al. Cognitive Response to Estradiol in Postmenopausal Women Is Modified by High Cortisol. Neurobiol. Aging 2012, 33, 829.e9–829.e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, A.M.; Nagi, K.; Thillaiappan, N.B.; Sukumaran, V.; Akhtar, S. Vitamin D and Its Potential Interplay With Pain Signaling Pathways. Front. Immunol. 2020, 11, 820. [Google Scholar] [CrossRef]
- Ohta, Y.; Suzuki, N.; Nakamura, S.; Hartwig, J.H.; Stossel, T.P. The Small GTPase RalA Targets Filamin to Induce Filopodia. Proc. Natl. Acad. Sci. USA 1999, 96, 2122–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castoria, G.; Lombardi, M.; Barone, M.V.; Bilancio, A.; Di Domenico, M.; Bottero, D.; Vitale, F.; Migliaccio, A.; Auricchio, F. Androgen-Stimulated DNA Synthesis and Cytoskeletal Changes in Fibroblasts by a Nontranscriptional Receptor Action. J. Cell Biol. 2003, 161, 547–556. [Google Scholar] [CrossRef]
- Castoria, G.; D’Amato, L.; Ciociola, A.; Giovannelli, P.; Giraldi, T.; Sepe, L.; Paolella, G.; Barone, M.V.; Migliaccio, A.; Auricchio, F. Androgen-Induced Cell Migration: Role of Androgen Receptor/Filamin A Association. PLoS ONE 2011, 6, e17218. [Google Scholar] [CrossRef] [Green Version]
- Giovannelli, P.; Di Donato, M.; Auricchio, F.; Castoria, G.; Migliaccio, A. Androgens Induce Invasiveness of Triple Negative Breast Cancer Cells Through AR/Src/PI3-K Complex Assembly. Sci. Rep. 2019, 9, 4490. [Google Scholar] [CrossRef] [Green Version]
- Simoncini, T.; Genazzani, A.R. Non-Genomic Actions of Sex Steroid Hormones. Eur. J. Endocrinol. 2003, 48, 281–292. [Google Scholar] [CrossRef]
- Giretti, M.S.; Fu, X.-D.; Rosa, G.D.; Sarotto, I.; Baldacci, C.; Garibaldi, S.; Mannella, P.; Biglia, N.; Sismondi, P.; Genazzani, A.R.; et al. Extra-Nuclear Signalling of Estrogen Receptor to Breast Cancer Cytoskeletal Remodelling, Migration and Invasion. PLoS ONE 2008, 3, e2238. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Henske, E.P. Estrogen-Induced Activation of Mammalian Target of Rapamycin Is Mediated via Tuberin and the Small GTPase Ras Homologue Enriched in Brain. Cancer Res. 2006, 66, 9461–9466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, C.-M.; Ai, N.; Ming-Yuen Lee, S. ROCK in CNS: Different Roles of Isoforms and Therapeutic Target for Neurodegenerative Disorders. Curr. Drug Targets 2017, 18, 455–462. [Google Scholar] [CrossRef]
- Tajouri, L.; Mellick, A.; Tourtellotte, A.; Nagra, R.M.; Griffiths, L.R. An Examination of MS Candidate Genes Identified as Differentially Regulated in Multiple Sclerosis Plaque Tissue, Using Absolute and Comparative Real-Time Q-PCR Analysis. Brain Res. Protoc. 2005, 15, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Qiao, L.; Tian, J.; Liu, A.; Wu, J.; Huang, J.; Shen, M.; Lai, X. Effect of Statins on Parkinson’s Disease: A Systematic Review and Meta-Analysis. Medicine 2019, 98, e14852. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.A.; Küderle, A.; Gaßner, H.; Klucken, J.; Eskofier, B.M.; Kluge, F. Inertial Sensor-Based Gait Parameters Reflect Patient-Reported Fatigue in Multiple Sclerosis. J. Neuroeng. Rehabil. 2020, 17, 165. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, Y.; Ito, T.; Mizutari, K.; Kakehata, S. Regenerative Effect of a ROCK Inhibitor, Y-27632, on Excitotoxic Trauma in an Organotypic Culture of the Cochlea. Front. Cell. Neurosci. 2020, 14, 572434. [Google Scholar] [CrossRef]
- Mueller, B.; Mack, H.; Teusch, N. Rho Kinase, a Promising Drug Target for Neurological Disorders. Nat. Rev. Drug Discov. 2005, 4, 387–398. [Google Scholar] [CrossRef]
- Wiatrak, B.; Kubis-Kubiak, A.; Piwowar, A.; Barg, E. PC12 Cell Line: Cell Types, Coating of Culture Vessels, Differentiation and Other Culture Conditions. Cells 2020, 9, 958. [Google Scholar] [CrossRef]
- Castelli, M.P.; Casti, A.; Casu, A.; Frau, R.; Bortolato, M.; Spiga, S.; Ennas, M.G. Regional Distribution of 5α-Reductase Type 2 in the Adult Rat Brain: An Immunohistochemical Analysis. Psychoneuroendocrinology 2013, 38, 281–293. [Google Scholar] [CrossRef]
- Fester, L.; Brandt, N.; Windhorst, S.; Pröls, F.; Bläute, C.; Rune, G.M. Control of Aromatase in Hippocampal Neurons. J. Steroid Biochem. Mol. Biol. 2016, 160, 9–14. [Google Scholar] [CrossRef]
- Hojo, Y.; Hattori, T.; Enami, T.; Furukawa, A.; Suzuki, K.; Ishii, H.; Mukai, H.; Morrison, J.H.; Janssen, W.G.M.; Kominami, S.; et al. Adult Male Rat Hippocampus Synthesizes Estradiol from Pregnenolone by Cytochromes P45017α and P450 Aromatase Localized in Neurons. Proc. Nat. Acad. Sci. USA 2004, 101, 865–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prange-Kiel, J.; Schmutterer, T.; Fester, L.; Zhou, L.; Imholz, P.; Brandt, N.; Vierk, R.; Jarry, H.; Rune, G.M. Endocrine Regulation of Estrogen Synthesis in the Hippocampus? Prog. Histochem. Cytochem. 2013, 48, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Hojo, Y.; Inoue, K.; Matsui, T.; Kawato, S.; McEwen, B.S.; Soya, H. Mild Exercise Increases Dihydrotestosterone in Hippocampus Providing Evidence for Androgenic Mediation of Neurogenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 13100–13105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, N.; Rune, G.M. Sex-Dependency of Oestrogen-Induced Structural Synaptic Plasticity: Inhibition of Aromatase versus Application of Estradiol in Rodents. Eur. J. Neurosci. 2020, 52, 2548–2559. [Google Scholar] [CrossRef] [PubMed]
- Leranth, C.; Prange-Kiel, J.; Frick, K.M.; Horvath, T.L. Low CA1 Spine Synapse Density Is Further Reduced by Castration in Male Non-Human Primates. Cereb. Cortex 2004, 14, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Leranth, C.; Hajszan, T.; MacLusky, N.J. Androgens Increase Spine Synapse Density in the CA1 Hippocampal Subfield of Ovariectomized Female Rats. J. Neurosci. 2004, 24, 495–499. [Google Scholar] [CrossRef] [Green Version]
- Do Rego, J.L.; Seong, J.Y.; Burel, D.; Leprince, J.; Luu-The, V.; Tsutsui, K.; Tonon, M.-C.; Pelletier, G.; Vaudry, H. Neurosteroid Biosynthesis: Enzymatic Pathways and Neuroendocrine Regulation by Neurotransmitters and Neuropeptides. Front. Neuroendocrinol. 2009, 30, 259–301. [Google Scholar] [CrossRef]
- Pelletier, G. Steroidogenic Enzymes in the Brain: Morphological Aspects. Prog. Brain Res. 2010, 181, 193–207. [Google Scholar] [CrossRef]
- Hojo, Y.; Higo, S.; Ishii, H.; Ooishi, Y.; Mukai, H.; Murakami, G.; Kominami, T.; Kimoto, T.; Honma, S.; Poirier, D.; et al. Comparison between Hippocampus-Synthesized and Circulation-Derived Sex Steroids in the Hippocampus. Endocrinology 2009, 150, 5106–5112. [Google Scholar] [CrossRef]
- Brandt, N.; Fester, L.; Rune, G.M. Neural Sex Steroids and Hippocampal Synaptic Plasticity. Vitam. Horm. 2020, 114, 125–143. [Google Scholar] [CrossRef]
- Kretz, O.; Fester, L.; Wehrenberg, U.; Zhou, L.; Brauckmann, S.; Zhao, S.; Prange-Kiel, J.; Naumann, T.; Jarry, H.; Frotscher, M.; et al. Hippocampal Synapses Depend on Hippocampal Estrogen Synthesis. J. Neurosci. 2004, 24, 5913–5921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migliaccio, A.; Castoria, G.; Bilancio, A.; Giovannelli, P.; Di Donato, M.; Auricchio, F. Non-genomic action of steroid hormones: More questions than answers. In Advances in Rapid Sex-Steroid Action: New Challenges and New Chances in Breast and Prostate Cancers; Castoria, G., Migliaccio, A., Eds.; Springer: New York, NY, USA, 2012; pp. 1–15. ISBN 978-1-4614-1764-4. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Donato, M.; Bilancio, A.; Auricchio, F.; Castoria, G.; Migliaccio, A. Androgens and NGF Mediate the Neurite-Outgrowth through Inactivation of RhoA. Cells 2023, 12, 373. https://doi.org/10.3390/cells12030373
Di Donato M, Bilancio A, Auricchio F, Castoria G, Migliaccio A. Androgens and NGF Mediate the Neurite-Outgrowth through Inactivation of RhoA. Cells. 2023; 12(3):373. https://doi.org/10.3390/cells12030373
Chicago/Turabian StyleDi Donato, Marzia, Antonio Bilancio, Ferdinando Auricchio, Gabriella Castoria, and Antimo Migliaccio. 2023. "Androgens and NGF Mediate the Neurite-Outgrowth through Inactivation of RhoA" Cells 12, no. 3: 373. https://doi.org/10.3390/cells12030373