
Epigenetic Perspective of Immunotherapy for Cancers

Abstract
:1. Introduction
2. Epigenetic Modifiers
3. Epigenetic Modifiers in T Cells
3.1. Epigenetic Modifiers in Immune Checkpoint Therapy
3.2. Epigenetic Modifiers in Antigen Processing and Presentation
3.3. Epigenetic Modifiers in Tumor-Infiltrating Immunosuppressive Cells
3.4. Epigenetic Modifiers in Inflammatory Cytokines and Chemokines
3.5. Epigenetic Modifiers in Natural Killer Cells
3.6. Epigenetic Modifiers in CAR-T Therapy
4. Transcription Factor Circuitry in ICT Resistance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buchbinder, E.I.; Desai, A. CTLA-4 and PD-1 Pathways: Similarities, Differences, and Implications of Their Inhibition. Am. J. Clin. Oncol. 2016, 39, 98–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, G.J.; Long, A.J.; Iwai, Y.; Bourque, K.; Chernova, T.; Nishimura, H.; Fitz, L.J.; Malenkovich, N.; Okazaki, T.; Byrne, M.C.; et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J. Exp. Med. 2000, 192, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidel, J.A.; Otsuka, A.; Kabashima, K. Anti-PD-1 and Anti-CTLA-4 Therapies in Cancer: Mechanisms of Action, Efficacy, and Limitations. Front. Oncol. 2018, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C. Tim-3, a negative regulator of anti-tumor immunity. Curr. Opin. Immunol. 2012, 24, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C. Tim-3: An emerging target in the cancer immunotherapy landscape. Cancer Immunol. Res. 2014, 2, 393–398. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Cao, J.; Zhao, C.; Li, X.; Zhou, C.; Hirsch, F.R. TIM-3, a promising target for cancer immunotherapy. Onco Targets Ther. 2018, 11, 7005–7009. [Google Scholar] [CrossRef] [Green Version]
- Huard, B.; Prigent, P.; Tournier, M.; Bruniquel, D.; Triebel, F. CD4/major histocompatibility complex class II interaction analyzed with CD4- and lymphocyte activation gene-3 (LAG-3)-Ig fusion proteins. Eur. J. Immunol. 1995, 25, 2718–2721. [Google Scholar] [CrossRef]
- Triebel, F.; Jitsukawa, S.; Baixeras, E.; Roman-Roman, S.; Genevee, C.; Viegas-Pequignot, E.; Hercend, T. LAG-3, a novel lymphocyte activation gene closely related to CD4. J. Exp. Med. 1990, 171, 1393–1405. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Harden, K.; Gonzalez, L.C.; Francesco, M.; Chiang, E.; Irving, B.; Tom, I.; Ivelja, S.; Refino, C.J.; Clark, H.; et al. The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells. Nat. Immunol. 2009, 10, 48–57. [Google Scholar] [CrossRef]
- Johnston, R.J.; Comps-Agrar, L.; Hackney, J.; Yu, X.; Huseni, M.; Yang, Y.; Park, S.; Javinal, V.; Chiu, H.; Irving, B.; et al. The immunoreceptor TIGIT regulates antitumor and antiviral CD8(+) T cell effector function. Cancer Cell 2014, 26, 923–937. [Google Scholar] [CrossRef]
- Abbas, H.A.; Hao, D.; Tomczak, K.; Barrodia, P.; Im, J.S.; Reville, P.K.; Alaniz, Z.; Wang, W.; Wang, R.; Wang, F.; et al. Single cell T cell landscape and T cell receptor repertoire profiling of AML in context of PD-1 blockade therapy. Nat. Commun. 2021, 12, 6071. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Liu, C.; Wang, Z.; Wu, W.; Zhang, N.; Zhang, L.; Hu, J.; Luo, P.; Zhang, J.; et al. Immune checkpoint modulators in cancer immunotherapy: Recent advances and emerging concepts. J. Hematol Oncol 2022, 15, 111. [Google Scholar] [CrossRef] [PubMed]
- Rohaan, M.W.; Wilgenhof, S.; Haanen, J. Adoptive cellular therapies: The current landscape. Virchows Arch. 2019, 474, 449–461. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Cao, Y.J. Adoptive Cell Therapy Targeting Neoantigens: A Frontier for Cancer Research. Front. Immunol. 2020, 11, 176. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, S.A.; Restifo, N.P.; Yang, J.C.; Morgan, R.A.; Dudley, M.E. Adoptive cell transfer: A clinical path to effective cancer immunotherapy. Nat. Rev. Cancer 2008, 8, 299–308. [Google Scholar] [CrossRef]
- Miller, J.S.; Soignier, Y.; Panoskaltsis-Mortari, A.; McNearney, S.A.; Yun, G.H.; Fautsch, S.K.; McKenna, D.; Le, C.; Defor, T.E.; Burns, L.J.; et al. Successful adoptive transfer and in vivo expansion of human haploidentical NK cells in patients with cancer. Blood 2005, 105, 3051–3057. [Google Scholar] [CrossRef] [Green Version]
- Qu, C.; Zhang, H.; Cao, H.; Tang, L.; Mo, H.; Liu, F.; Zhang, L.; Yi, Z.; Long, L.; Yan, L.; et al. Tumor buster—Where will the CAR-T cell therapy ‘missile’ go? Mol. Cancer 2022, 21, 201. [Google Scholar] [CrossRef]
- Donninger, H.; Li, C.; Eaton, J.W.; Yaddanapudi, K. Cancer Vaccines: Promising Therapeutics or an Unattainable Dream. Vaccines 2021, 9, 668. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.G.; Sang, Y.B.; Lee, J.H.; Chon, H.J. Combining Cancer Vaccines with Immunotherapy: Establishing a New Immunological Approach. Int. J. Mol. Sci. 2021, 22, 8035. [Google Scholar] [CrossRef]
- Corral, L.G.; Haslett, P.A.; Muller, G.W.; Chen, R.; Wong, L.M.; Ocampo, C.J.; Patterson, R.T.; Stirling, D.I.; Kaplan, G. Differential cytokine modulation and T cell activation by two distinct classes of thalidomide analogues that are potent inhibitors of TNF-alpha. J. Immunol. 1999, 163, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Davies, F.E.; Raje, N.; Hideshima, T.; Lentzsch, S.; Young, G.; Tai, Y.T.; Lin, B.; Podar, K.; Gupta, D.; Chauhan, D.; et al. Thalidomide and immunomodulatory derivatives augment natural killer cell cytotoxicity in multiple myeloma. Blood 2001, 98, 210–216. [Google Scholar] [CrossRef]
- Haslett, P.A.; Klausner, J.D.; Makonkawkeyoon, S.; Moreira, A.; Metatratip, P.; Boyle, B.; Kunachiwa, W.; Maneekarn, N.; Vongchan, P.; Corral, L.G.; et al. Thalidomide stimulates T cell responses and interleukin 12 production in HIV-infected patients. AIDS Res. Hum. Retroviruses 1999, 15, 1169–1179. [Google Scholar] [CrossRef] [Green Version]
- Singhal, S.; Mehta, J.; Desikan, R.; Ayers, D.; Roberson, P.; Eddlemon, P.; Munshi, N.; Anaissie, E.; Wilson, C.; Dhodapkar, M.; et al. Antitumor activity of thalidomide in refractory multiple myeloma. N. Engl. J. Med. 1999, 341, 1565–1571. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, L.B.; Salama, A.K.S. A review of cancer immunotherapy toxicity. CA Cancer J. Clin. 2020, 70, 86–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michot, J.M.; Bigenwald, C.; Champiat, S.; Collins, M.; Carbonnel, F.; Postel-Vinay, S.; Berdelou, A.; Varga, A.; Bahleda, R.; Hollebecque, A.; et al. Immune-related adverse events with immune checkpoint blockade: A comprehensive review. Eur J. Cancer 2016, 54, 139–148. [Google Scholar] [CrossRef]
- Postow, M.A.; Sidlow, R.; Hellmann, M.D. Immune-Related Adverse Events Associated with Immune Checkpoint Blockade. N. Engl. J. Med. 2018, 378, 158–168. [Google Scholar] [CrossRef]
- Ramos-Casals, M.; Brahmer, J.R.; Callahan, M.K.; Flores-Chavez, A.; Keegan, N.; Khamashta, M.A.; Lambotte, O.; Mariette, X.; Prat, A.; Suarez-Almazor, M.E. Immune-related adverse events of checkpoint inhibitors. Nat. Rev. Dis. Primers 2020, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Beyes, S.; Bediaga, N.G.; Zippo, A. An Epigenetic Perspective on Intra-Tumour Heterogeneity: Novel Insights and New Challenges from Multiple Fields. Cancers 2021, 13, 4969. [Google Scholar] [CrossRef]
- Easwaran, H.; Tsai, H.C.; Baylin, S.B. Cancer epigenetics: Tumor heterogeneity, plasticity of stem-like states, and drug resistance. Mol. Cell 2014, 54, 716–727. [Google Scholar] [CrossRef] [Green Version]
- Darwiche, N. Epigenetic mechanisms and the hallmarks of cancer: An intimate affair. Am. J. Cancer Res. 2020, 10, 1954–1978. [Google Scholar] [PubMed]
- Kugel, S.; Feldman, J.L.; Klein, M.A.; Silberman, D.M.; Sebastian, C.; Mermel, C.; Dobersch, S.; Clark, A.R.; Getz, G.; Denu, J.M.; et al. Identification of and Molecular Basis for SIRT6 Loss-of-Function Point Mutations in Cancer. Cell Rep. 2015, 13, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Peng, D.; Kryczek, I.; Nagarsheth, N.; Zhao, L.; Wei, S.; Wang, W.; Sun, Y.; Zhao, E.; Vatan, L.; Szeliga, W.; et al. Epigenetic silencing of TH1-type chemokines shapes tumour immunity and immunotherapy. Nature 2015, 527, 249–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topper, M.J.; Vaz, M.; Marrone, K.A.; Brahmer, J.R.; Baylin, S.B. The emerging role of epigenetic therapeutics in immuno-oncology. Nat. Rev. Clin. Oncol. 2020, 17, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Callahan, S.C.; Divenko, M.; Barrodia, P.; Singh, A.K.; Arslan, E.; Liu, Z.; Yang, J.; Anvar, N.; Amit, M.; Xie, T.; et al. KMT2D Loss Promotes Head and Neck Squamous Cell Carcinoma Through Enhancer Reprogramming and Modulation of Immune Microenvironment. Biorxiv 2021. [Google Scholar] [CrossRef]
- Kim, Y.D.; Park, S.M.; Ha, H.C.; Lee, A.R.; Won, H.; Cha, H.; Cho, S.; Cho, J.M. HDAC Inhibitor, CG-745, Enhances the Anti-Cancer Effect of Anti-PD-1 Immune Checkpoint Inhibitor by Modulation of the Immune Microenvironment. J. Cancer 2020, 11, 4059–4072. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Bengsch, F.; Svoronos, N.; Rutkowski, M.R.; Bitler, B.G.; Allegrezza, M.J.; Yokoyama, Y.; Kossenkov, A.V.; Bradner, J.E.; Conejo-Garcia, J.R.; et al. BET Bromodomain Inhibition Promotes Anti-tumor Immunity by Suppressing PD-L1 Expression. Cell Rep. 2016, 16, 2829–2837. [Google Scholar] [CrossRef] [Green Version]
- Naik, R.R.; Singh, A.K.; Mali, A.M.; Khirade, M.F.; Bapat, S.A. A tumor deconstruction platform identifies definitive end points in the evaluation of drug responses. Oncogene 2016, 35, 727–737. [Google Scholar] [CrossRef]
- Orouji, E.; Raman, A.T.; Singh, A.K.; Sorokin, A.; Arslan, E.; Ghosh, A.K.; Schulz, J.; Terranova, C.; Jiang, S.; Tang, M.; et al. Chromatin state dynamics confers specific therapeutic strategies in enhancer subtypes of colorectal cancer. Gut 2022, 71, 938–949. [Google Scholar] [CrossRef]
- Singh, A.K.; Chandra, N.; Bapat, S.A. Evaluation of Epigenetic Drug Targeting of Heterogenous Tumor Cell Fractions Using Potential Biomarkers of Response in Ovarian Cancer. Clin. Cancer Res. 2015, 21, 5151–5163. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.B., Jr.; Anand, K.S.; Bapat, S.; Clements, A.J. Transforming the future of treatment for ovarian cancer. Clin. Exp. Pharmacol. 2014, 4, 3. [Google Scholar]
- O’Donnell, J.S.; Long, G.V.; Scolyer, R.A.; Teng, M.W.; Smyth, M.J. Resistance to PD1/PDL1 checkpoint inhibition. Cancer Treat. Rev. 2017, 52, 71–81. [Google Scholar] [CrossRef]
- Pitt, J.M.; Vetizou, M.; Daillere, R.; Roberti, M.P.; Yamazaki, T.; Routy, B.; Lepage, P.; Boneca, I.G.; Chamaillard, M.; Kroemer, G.; et al. Resistance Mechanisms to Immune-Checkpoint Blockade in Cancer: Tumor-Intrinsic and -Extrinsic Factors. Immunity 2016, 44, 1255–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Hu-Lieskovan, S.; Wargo, J.A.; Ribas, A. Primary, Adaptive, and Acquired Resistance to Cancer Immunotherapy. Cell 2017, 168, 707–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual-Garcia, M.; Bonfill-Teixidor, E.; Planas-Rigol, E.; Rubio-Perez, C.; Iurlaro, R.; Arias, A.; Cuartas, I.; Sala-Hojman, A.; Escudero, L.; Martinez-Ricarte, F.; et al. LIF regulates CXCL9 in tumor-associated macrophages and prevents CD8(+) T cell tumor-infiltration impairing anti-PD1 therapy. Nat. Commun. 2019, 10, 2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.A.; Takai, D. The role of DNA methylation in mammalian epigenetics. Science 2001, 293, 1068–1070. [Google Scholar] [CrossRef] [PubMed]
- Terranova, C.J.; Tang, M.; Maitituoheti, M.; Raman, A.T.; Ghosh, A.K.; Schulz, J.; Amin, S.B.; Orouji, E.; Tomczak, K.; Sarkar, S.; et al. Reprogramming of bivalent chromatin states in NRAS mutant melanoma suggests PRC2 inhibition as a therapeutic strategy. Cell Rep. 2021, 36, 109410. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Audia, J.E.; Campbell, R.M. Histone Modifications and Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, a019521. [Google Scholar] [CrossRef]
- Dawson, M.A.; Kouzarides, T. Cancer epigenetics: From mechanism to therapy. Cell 2012, 150, 12–27. [Google Scholar] [CrossRef] [Green Version]
- McCaw, T.R.; Goel, N.; Brooke, D.J.; Katre, A.A.; Londono, A.I.; Smith, H.J.; Randall, T.D.; Arend, R.C. Class I histone deacetylase inhibition promotes CD8 T cell activation in ovarian cancer. Cancer Med. 2021, 10, 709–717. [Google Scholar] [CrossRef]
- Preglej, T.; Hamminger, P.; Luu, M.; Bulat, T.; Andersen, L.; Goschl, L.; Stolz, V.; Rica, R.; Sandner, L.; Waltenberger, D.; et al. Histone deacetylases 1 and 2 restrain CD4+ cytotoxic T lymphocyte differentiation. JCI Insight 2020, 5, e133393. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.C.; Belmonte, P.J.; Constans, M.M.; Chen, M.W.; McWilliams, D.C.; Hiebert, S.W.; Shapiro, V.S. Histone Deacetylase 3 Is Required for T Cell Maturation. J. Immunol. 2015, 195, 1578–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolz, J.C.; Harty, J.T. IL-15 regulates memory CD8+ T cell O-glycan synthesis and affects trafficking. J. Clin. Investig. 2014, 124, 1013–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheer, S.; Runting, J.; Bramhall, M.; Russ, B.; Zaini, A.; Ellemor, J.; Rodrigues, G.; Ng, J.; Zaph, C. The Methyltransferase DOT1L Controls Activation and Lineage Integrity in CD4(+) T Cells during Infection and Inflammation. Cell Rep. 2020, 33, 108505. [Google Scholar] [CrossRef]
- Onodera, A.; Kiuchi, M.; Kokubo, K.; Kato, M.; Ogino, T.; Horiuchi, S.; Kanai, U.; Hirahara, K.; Nakayama, T. Menin Controls the Memory Th2 Cell Function by Maintaining the Epigenetic Integrity of Th2 Cells. J. Immunol. 2017, 199, 1153–1162. [Google Scholar] [CrossRef] [Green Version]
- Cribbs, A.P.; Terlecki-Zaniewicz, S.; Philpott, M.; Baardman, J.; Ahern, D.; Lindow, M.; Obad, S.; Oerum, H.; Sampey, B.; Mander, P.K.; et al. Histone H3K27me3 demethylases regulate human Th17 cell development and effector functions by impacting on metabolism. Proc. Natl. Acad. Sci. USA 2020, 117, 6056–6066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
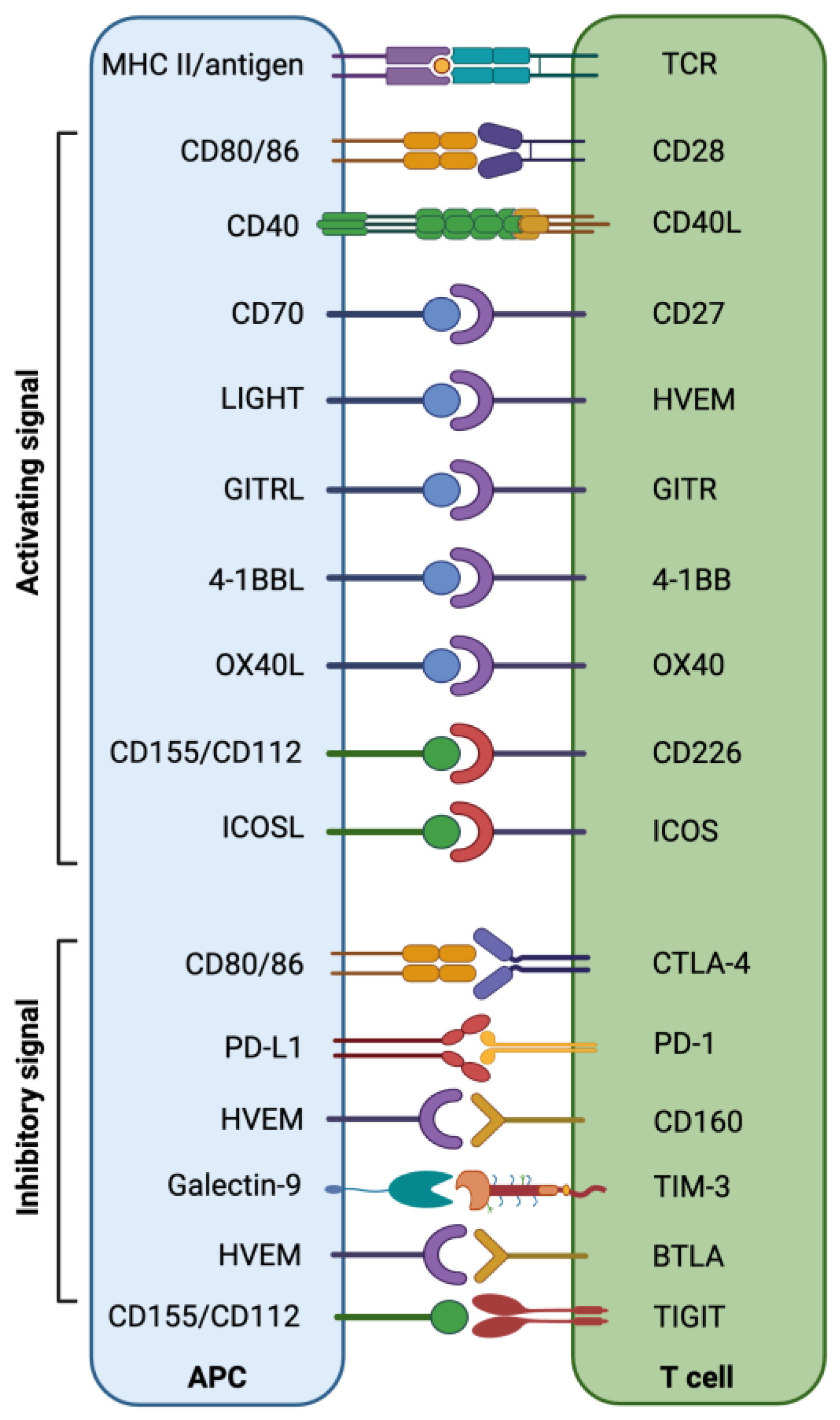
- O’Neill, R.E.; Cao, X. Co-stimulatory and co-inhibitory pathways in cancer immunotherapy. Adv. Cancer Res. 2019, 143, 145–194. [Google Scholar] [CrossRef]
- Maitituoheti, M.A.S.; Tang, M.; Ho, L.-L.; Terranova, C.; Galani, K.; Keung, E.Z.; Creasy, C.A.; Wu, M.; Chen, J.; Chen, N.; et al. Enhancer Reprogramming in Melanoma Immune Checkpoint Therapy Resistance. bioRxiv 2022. [Google Scholar]
- Ni, L.; Dong, C. New B7 Family Checkpoints in Human Cancers. Mol. Cancer Ther. 2017, 16, 1203–1211. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Feng, H.; Cheng, X.; Liu, K.; Cai, D.; Zhao, R. Potential Therapeutic Targets of B7 Family in Colorectal Cancer. Front. Immunol. 2020, 11, 681. [Google Scholar] [CrossRef]
- Saleh, R.; Toor, S.M.; Sasidharan Nair, V.; Elkord, E. Role of Epigenetic Modifications in Inhibitory Immune Checkpoints in Cancer Development and Progression. Front. Immunol. 2020, 11, 1469. [Google Scholar] [CrossRef] [PubMed]
- Buttler, C.A.; Chuong, E.B. Emerging roles for endogenous retroviruses in immune epigenetic regulation. Immunol. Rev. 2022, 305, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Liu, X.; Zeng, Y.; Liu, J.; Wu, F. DNA methyltransferase inhibitors combination therapy for the treatment of solid tumor: Mechanism and clinical application. Clin. Epigenetics 2021, 13, 166. [Google Scholar] [CrossRef] [PubMed]
- Nijhof, I.S.; Casneuf, T.; van Velzen, J.; van Kessel, B.; Axel, A.E.; Syed, K.; Groen, R.W.; van Duin, M.; Sonneveld, P.; Minnema, M.C.; et al. CD38 expression and complement inhibitors affect response and resistance to daratumumab therapy in myeloma. Blood 2016, 128, 959–970. [Google Scholar] [CrossRef] [Green Version]
- Woods, D.M.; Sodre, A.L.; Villagra, A.; Sarnaik, A.; Sotomayor, E.M.; Weber, J. HDAC Inhibition Upregulates PD-1 Ligands in Melanoma and Augments Immunotherapy with PD-1 Blockade. Cancer Immunol. Res. 2015, 3, 1375–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, D.A.; Hou, Y.; Bakouny, Z.; Ficial, M.; Sant’ Angelo, M.; Forman, J.; Ross-Macdonald, P.; Berger, A.C.; Jegede, O.A.; Elagina, L.; et al. Interplay of somatic alterations and immune infiltration modulates response to PD-1 blockade in advanced clear cell renal cell carcinoma. Nat. Med. 2020, 26, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Griffin, G.K.; Wu, J.; Iracheta-Vellve, A.; Patti, J.C.; Hsu, J.; Davis, T.; Dele-Oni, D.; Du, P.P.; Halawi, A.G.; Ishizuka, J.J.; et al. Epigenetic silencing by SETDB1 suppresses tumour intrinsic immunogenicity. Nature 2021, 595, 309–314. [Google Scholar] [CrossRef]
- Harjes, U. SETDB1, a new target for immunotherapy. Nat. Rev. Cancer 2021, 21, 412. [Google Scholar] [CrossRef]
- Dutta, R.; Khalil, R.; Mayilsamy, K.; Green, R.; Howell, M.; Bharadwaj, S.; Mohapatra, S.S.; Mohapatra, S. Combination Therapy of Mithramycin A and Immune Checkpoint Inhibitor for the Treatment of Colorectal Cancer in an Orthotopic Murine Model. Front. Immunol. 2021, 12, 706133. [Google Scholar] [CrossRef]
- Andrieu, G.P.; Shafran, J.S.; Smith, C.L.; Belkina, A.C.; Casey, A.N.; Jafari, N.; Denis, G.V. BET protein targeting suppresses the PD-1/PD-L1 pathway in triple-negative breast cancer and elicits anti-tumor immune response. Cancer Lett. 2019, 465, 45–58. [Google Scholar] [CrossRef]
- Adeegbe, D.O.; Liu, S.; Hattersley, M.M.; Bowden, M.; Zhou, C.W.; Li, S.; Vlahos, R.; Grondine, M.; Dolgalev, I.; Ivanova, E.V.; et al. BET Bromodomain Inhibition Cooperates with PD-1 Blockade to Facilitate Antitumor Response in Kras-Mutant Non-Small Cell Lung Cancer. Cancer Immunol. Res. 2018, 6, 1234–1245. [Google Scholar] [CrossRef] [Green Version]
- Klumper, N.; Ralser, D.J.; Bawden, E.G.; Landsberg, J.; Zarbl, R.; Kristiansen, G.; Toma, M.; Ritter, M.; Holzel, M.; Ellinger, J.; et al. LAG3 (LAG-3, CD223) DNA methylation correlates with LAG3 expression by tumor and immune cells, immune cell infiltration, and overall survival in clear cell renal cell carcinoma. J. Immunother. Cancer 2020, 8, e000552. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, J.; Meng, J.; Jiang, H.; Zhao, J.; Qian, H.; Chen, T. Epigenetic Modification Mediates the Increase of LAG-3(+) T Cells in Chronic Osteomyelitis. Inflammation 2017, 40, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hu, W.; Zheng, X.; Zhang, C.; Du, P.; Zheng, Z.; Yang, Y.; Wu, J.; Ji, M.; Jiang, J.; et al. Emerging immune checkpoints for cancer therapy. Acta Oncol. 2015, 54, 1706–1713. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, K.; Kukita, A.; Li, Y.J.; Uehara, N.; Zhang, J.Q.; Takahashi, I.; Kukita, T. Regulation of osteoclastogenesis through Tim-3: Possible involvement of the Tim-3/galectin-9 system in the modulation of inflammatory bone destruction. Lab. Invest. 2014, 94, 1200–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Song, Y.; Yang, H.; Liu, Z.; Gao, L.; Liang, X.; Ma, C. Tumor cell-intrinsic Tim-3 promotes liver cancer via NF-kappaB/IL-6/STAT3 axis. Oncogene 2018, 37, 2456–2468. [Google Scholar] [CrossRef]
- Zhang, L.; Tian, S.; Pei, M.; Zhao, M.; Wang, L.; Jiang, Y.; Yang, T.; Zhao, J.; Song, L.; Yang, X. Crosstalk between histone modification and DNA methylation orchestrates the epigenetic regulation of the costimulatory factors, Tim3 and galectin9, in cervical cancer. Oncol. Rep. 2019, 42, 2655–2669. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tian, S.; Zhao, M.; Yang, T.; Quan, S.; Yang, Q.; Song, L.; Yang, X. SUV39H1-DNMT3A-mediated epigenetic regulation of Tim-3 and galectin-9 in the cervical cancer. Cancer Cell Int. 2020, 20, 325. [Google Scholar] [CrossRef]
- Niebel, D.; Frohlich, A.; Zarbl, R.; Fietz, S.; de Vos, L.; Vogt, T.J.; Dietrich, J.; Sirokay, J.; Kuster, P.; Saavedra, G.; et al. DNA methylation regulates TIGIT expression within the melanoma microenvironment, is prognostic for overall survival, and predicts progression-free survival in patients treated with anti-PD-1 immunotherapy. Clin. Epigenetics 2022, 14, 50. [Google Scholar] [CrossRef]
- Gibbs, Z.A.; Whitehurst, A.W. Emerging Contributions of Cancer/Testis Antigens to Neoplastic Behaviors. Trends Cancer 2018, 4, 701–712. [Google Scholar] [CrossRef]
- Wang, C.; Gu, Y.; Zhang, K.; Xie, K.; Zhu, M.; Dai, N.; Jiang, Y.; Guo, X.; Liu, M.; Dai, J.; et al. Systematic identification of genes with a cancer-testis expression pattern in 19 cancer types. Nat. Commun. 2016, 7, 10499. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, E.A.; Srivastava, P.; Matsuzaki, J.; Brumberger, Z.; Wang, E.S.; Kocent, J.; Miller, A.; Roloff, G.W.; Wong, H.Y.; Paluch, B.E.; et al. NY-ESO-1 Vaccination in Combination with Decitabine Induces Antigen-Specific T-lymphocyte Responses in Patients with Myelodysplastic Syndrome. Clin. Cancer Res. 2018, 24, 1019–1029. [Google Scholar] [CrossRef]
- Ishihara, M.; Kitano, S.; Kageyama, S.; Miyahara, Y.; Yamamoto, N.; Kato, H.; Mishima, H.; Hattori, H.; Funakoshi, T.; Kojima, T.; et al. NY-ESO-1-specific redirected T cells with endogenous TCR knockdown mediate tumor response and cytokine release syndrome. J. Immunother. Cancer 2022, 10, e003811. [Google Scholar] [CrossRef]
- Grunewald, C.M.; Schulz, W.A.; Skowron, M.A.; Hoffmann, M.J.; Niegisch, G. Tumor immunotherapy—The potential of epigenetic drugs to overcome resistance. Transl. Cancer Res. 2018, 7, 1151–1160. [Google Scholar] [CrossRef]
- Khan, A.N.; Gregorie, C.J.; Tomasi, T.B. Histone deacetylase inhibitors induce TAP, LMP, Tapasin genes and MHC class I antigen presentation by melanoma cells. Cancer Immunol. Immunother. 2008, 57, 647–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magner, W.J.; Kazim, A.L.; Stewart, C.; Romano, M.A.; Catalano, G.; Grande, C.; Keiser, N.; Santaniello, F.; Tomasi, T.B. Activation of MHC class I, II, and CD40 gene expression by histone deacetylase inhibitors. J. Immunol. 2000, 165, 7017–7024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setiadi, A.F.; Omilusik, K.; David, M.D.; Seipp, R.P.; Hartikainen, J.; Gopaul, R.; Choi, K.B.; Jefferies, W.A. Epigenetic enhancement of antigen processing and presentation promotes immune recognition of tumors. Cancer Res. 2008, 68, 9601–9607. [Google Scholar] [CrossRef] [Green Version]
- Maeda, T.; Towatari, M.; Kosugi, H.; Saito, H. Up-regulation of costimulatory/adhesion molecules by histone deacetylase inhibitors in acute myeloid leukemia cells. Blood 2000, 96, 3847–3856. [Google Scholar] [CrossRef]
- Wang, L.X.; Mei, Z.Y.; Zhou, J.H.; Yao, Y.S.; Li, Y.H.; Xu, Y.H.; Li, J.X.; Gao, X.N.; Zhou, M.H.; Jiang, M.M.; et al. Low dose decitabine treatment induces CD80 expression in cancer cells and stimulates tumor specific cytotoxic T lymphocyte responses. PLoS ONE 2013, 8, e62924. [Google Scholar] [CrossRef] [PubMed]
- Federico, A.; Steinfass, T.; Larribere, L.; Novak, D.; Moris, F.; Nunez, L.E.; Umansky, V.; Utikal, J. Mithramycin A and Mithralog EC-8042 Inhibit SETDB1 Expression and Its Oncogenic Activity in Malignant Melanoma. Mol. Ther. Oncolytics 2020, 18, 83–99. [Google Scholar] [CrossRef]
- Lin, J.; Guo, D.; Liu, H.; Zhou, W.; Wang, C.; Muller, I.; Kossenkov, A.V.; Drapkin, R.; Bitler, B.G.; Helin, K.; et al. The SETDB1-TRIM28 Complex Suppresses Antitumor Immunity. Cancer Immunol. Res. 2021, 9, 1413–1424. [Google Scholar] [CrossRef]
- Truong, A.S.; Zhou, M.; Krishnan, B.; Utsumi, T.; Manocha, U.; Stewart, K.G.; Beck, W.; Rose, T.L.; Milowsky, M.I.; He, X.; et al. Entinostat induces antitumor immune responses through immune editing of tumor neoantigens. J. Clin. Invest. 2021, 131, e138560. [Google Scholar] [CrossRef]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef]
- Xie, Z.; Ago, Y.; Okada, N.; Tachibana, M. Valproic acid attenuates immunosuppressive function of myeloid-derived suppressor cells. J. Pharmacol. Sci. 2018, 137, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Ikegami, T.; Ago, Y.; Okada, N.; Tachibana, M. Valproic acid attenuates CCR2-dependent tumor infiltration of monocytic myeloid-derived suppressor cells, limiting tumor progression. Oncoimmunology 2020, 9, 1734268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.F.; Ning, F.; Liu, Z.C.; Wu, L.; Li, Z.Q.; Qi, Y.F.; Zhang, G.; Wang, H.S.; Cai, S.H.; Du, J. Histone deacetylase inhibitors deplete myeloid-derived suppressor cells induced by 4T1 mammary tumors in vivo and in vitro. Cancer Immunol. Immunother. 2017, 66, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Fontenot, J.D.; Rasmussen, J.P.; Williams, L.M.; Dooley, J.L.; Farr, A.G.; Rudensky, A.Y. Regulatory T cell lineage specification by the forkhead transcription factor foxp3. Immunity 2005, 22, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Marie, J.C.; Letterio, J.J.; Gavin, M.; Rudensky, A.Y. TGF-beta1 maintains suppressor function and Foxp3 expression in CD4+CD25+ regulatory T cells. J. Exp. Med. 2005, 201, 1061–1067. [Google Scholar] [CrossRef]
- Burchill, M.A.; Yang, J.; Vogtenhuber, C.; Blazar, B.R.; Farrar, M.A. IL-2 receptor beta-dependent STAT5 activation is required for the development of Foxp3+ regulatory T cells. J. Immunol. 2007, 178, 280–290. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.P.; Leonard, W.J. CREB/ATF-dependent T cell receptor-induced FoxP3 gene expression: A role for DNA methylation. J. Exp. Med. 2007, 204, 1543–1551. [Google Scholar] [CrossRef]
- Lal, G.; Zhang, N.; van der Touw, W.; Ding, Y.; Ju, W.; Bottinger, E.P.; Reid, S.P.; Levy, D.E.; Bromberg, J.S. Epigenetic regulation of Foxp3 expression in regulatory T cells by DNA methylation. J. Immunol. 2009, 182, 259–273. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Jin, W.; Hardegen, N.; Lei, K.J.; Li, L.; Marinos, N.; McGrady, G.; Wahl, S.M. Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J. Exp. Med. 2003, 198, 1875–1886. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; de Zoeten, E.F.; Ozkaynak, E.; Chen, C.; Wang, L.; Porrett, P.M.; Li, B.; Turka, L.A.; Olson, E.N.; Greene, M.I.; et al. Deacetylase inhibition promotes the generation and function of regulatory T cells. Nat. Med. 2007, 13, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xiao, Y.; Zhu, Z.; Li, B.; Greene, M.I. Immune regulation by histone deacetylases: A focus on the alteration of FOXP3 activity. Immunol. Cell Biol. 2012, 90, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Ohkura, N.; Hamaguchi, M.; Morikawa, H.; Sugimura, K.; Tanaka, A.; Ito, Y.; Osaki, M.; Tanaka, Y.; Yamashita, R.; Nakano, N.; et al. T cell receptor stimulation-induced epigenetic changes and Foxp3 expression are independent and complementary events required for Treg cell development. Immunity 2012, 37, 785–799. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Quiros, J.; Mahuron, K.; Pai, C.C.; Ranzani, V.; Young, A.; Silveria, S.; Harwin, T.; Abnousian, A.; Pagani, M.; et al. Targeting EZH2 Reprograms Intratumoral Regulatory T Cells to Enhance Cancer Immunity. Cell Rep. 2018, 23, 3262–3274. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, J.M.; Glenn, J.D.; Calabresi, P.A.; Whartenby, K.A. KLF4 modulates expression of IL-6 in dendritic cells via both promoter activation and epigenetic modification. J. Biol. Chem. 2013, 288, 23868–23874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; He, Z.; Li, H.; Yao, C.; Zhang, Y.; He, L.; Li, S.; Huang, J.; Guo, Z. Distinct functional patterns of gene promoter hypomethylation and hypermethylation in cancer genomes. PLoS ONE 2012, 7, e44822. [Google Scholar] [CrossRef]
- Ma, D.; Jiang, C.; Hu, X.; Liu, H.; Li, Q.; Li, T.; Yang, Y.; Li, O. Methylation patterns of the IFN-gamma gene in cervical cancer tissues. Sci. Rep. 2014, 4, 6331. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Xu, J.; Zhu, Q.; Qin, X.; Cao, Y.; Lou, J.; Xu, Y.; Ke, X.; Li, Q.; Xie, E.; et al. Downregulation of IFNG in CD4(+) T cells in lung cancer through hypermethylation: A possible mechanism of tumor-induced immunosuppression. PLoS ONE 2013, 8, e79064. [Google Scholar] [CrossRef]
- Rincon-Orozco, B.; Halec, G.; Rosenberger, S.; Muschik, D.; Nindl, I.; Bachmann, A.; Ritter, T.M.; Dondog, B.; Ly, R.; Bosch, F.X.; et al. Epigenetic silencing of interferon-kappa in human papillomavirus type 16-positive cells. Cancer Res. 2009, 69, 8718–8725. [Google Scholar] [CrossRef] [Green Version]
- Schoenborn, J.R.; Dorschner, M.O.; Sekimata, M.; Santer, D.M.; Shnyreva, M.; Fitzpatrick, D.R.; Stamatoyannopoulos, J.A.; Wilson, C.B. Comprehensive epigenetic profiling identifies multiple distal regulatory elements directing transcription of the gene encoding interferon-gamma. Nat. Immunol. 2007, 8, 732–742. [Google Scholar] [CrossRef]
- Santangelo, S.; Cousins, D.J.; Winkelmann, N.; Triantaphyllopoulos, K.; Staynov, D.Z. Chromatin structure and DNA methylation of the IL-4 gene in human T(H)2 cells. Chromosome Res. 2009, 17, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Starnes, T.; Rasila, K.K.; Robertson, M.J.; Brahmi, Z.; Dahl, R.; Christopherson, K.; Hromas, R. The chemokine CXCL14 (BRAK) stimulates activated NK cell migration: Implications for the downregulation of CXCL14 in malignancy. Exp. Hematol. 2006, 34, 1101–1105. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Lin, F.; Zhu, G.; Xue, X.; Ding, Y.; Zhao, Z.; Zhang, L.; Shen, X. Abnormal hypermethylation of promoter region downregulates chemokine CXC ligand 14 expression in gastric cancer. Int. J. Oncol. 2013, 43, 1487–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, E.Y.; Shurin, M.R.; Tourkova, I.L.; Gutkin, D.W.; Shurin, G.V. Epigenetic mechanisms of promigratory chemokine CXCL14 regulation in human prostate cancer cells. Cancer Res. 2010, 70, 4394–4401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, E.A.; Camargo, A.A.; Braun, K.; Slowik, R.; Cavalli, I.J.; Ribeiro, E.M.; Pedrosa Fde, O.; de Souza, E.M.; Costa, F.F.; Klassen, G. Simultaneous CXCL12 and ESR1 CpG island hypermethylation correlates with poor prognosis in sporadic breast cancer. BMC Cancer 2010, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- Tekpli, X.; Landvik, N.E.; Anmarkud, K.H.; Skaug, V.; Haugen, A.; Zienolddiny, S. DNA methylation at promoter regions of interleukin 1B, interleukin 6, and interleukin 8 in non-small cell lung cancer. Cancer Immunol. Immunother. 2013, 62, 337–345. [Google Scholar] [CrossRef]
- Baird, A.M.; Leonard, J.; Naicker, K.M.; Kilmartin, L.; O’Byrne, K.J.; Gray, S.G. IL-23 is pro-proliferative, epigenetically regulated and modulated by chemotherapy in non-small cell lung cancer. Lung Cancer 2013, 79, 83–90. [Google Scholar] [CrossRef]
- Suzuki, M.; Iizasa, T.; Nakajima, T.; Kubo, R.; Iyoda, A.; Hiroshima, K.; Nakatani, Y.; Fujisawa, T. Aberrant methylation of IL-12Rbeta2 gene in lung adenocarcinoma cells is associated with unfavorable prognosis. Ann. Surg. Oncol. 2007, 14, 2636–2642. [Google Scholar] [CrossRef]
- Suzuki, M.; Wada, H.; Yoshino, M.; Tian, L.; Shigematsu, H.; Suzuki, H.; Alaa, M.; Tamura, H.; Fujiwara, T.; Nagato, K.; et al. Molecular characterization of chronic obstructive pulmonary disease-related non-small cell lung cancer through aberrant methylation and alterations of EGFR signaling. Ann. Surg. Oncol. 2010, 17, 878–888. [Google Scholar] [CrossRef]
- Conte, M.; De Palma, R.; Altucci, L. HDAC inhibitors as epigenetic regulators for cancer immunotherapy. Int. J. Biochem. Cell Biol. 2018, 98, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Maio, M.; Covre, A.; Fratta, E.; Di Giacomo, A.M.; Taverna, P.; Natali, P.G.; Coral, S.; Sigalotti, L. Molecular Pathways: At the Crossroads of Cancer Epigenetics and Immunotherapy. Clin. Cancer Res. 2015, 21, 4040–4047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Thomas, S.; Munster, P.N. Epigenetic modulation with histone deacetylase inhibitors in combination with immunotherapy. Epigenomics 2015, 7, 641–652. [Google Scholar] [CrossRef]
- Fraietta, J.A.; Nobles, C.L.; Sammons, M.A.; Lundh, S.; Carty, S.A.; Reich, T.J.; Cogdill, A.P.; Morrissette, J.J.D.; DeNizio, J.E.; Reddy, S.; et al. Disruption of TET2 promotes the therapeutic efficacy of CD19-targeted T cells. Nature 2018, 558, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Foskolou, I.P.; Barbieri, L.; Vernet, A.; Bargiela, D.; Cunha, P.P.; Velica, P.; Suh, E.; Pietsch, S.; Matuleviciute, R.; Rundqvist, H.; et al. The S enantiomer of 2-hydroxyglutarate increases central memory CD8 populations and improves CAR-T therapy outcome. Blood Adv. 2020, 4, 4483–4493. [Google Scholar] [CrossRef]
- Li, S.; Xue, L.; Wang, M.; Qiang, P.; Xu, H.; Zhang, X.; Kang, W.; You, F.; Xu, H.; Wang, Y.; et al. Decitabine enhances cytotoxic effect of T cells with an anti-CD19 chimeric antigen receptor in treatment of lymphoma. Onco Targets Ther. 2019, 12, 5627–5638. [Google Scholar] [CrossRef] [Green Version]
- Zou, F.; Lu, L.; Liu, J.; Xia, B.; Zhang, W.; Hu, Q.; Liu, W.; Zhang, Y.; Lin, Y.; Jing, S.; et al. Engineered triple inhibitory receptor resistance improves anti-tumor CAR-T cell performance via CD56. Nat. Commun. 2019, 10, 4109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kailayangiri, S.; Altvater, B.; Lesch, S.; Balbach, S.; Gottlich, C.; Kuhnemundt, J.; Mikesch, J.H.; Schelhaas, S.; Jamitzky, S.; Meltzer, J.; et al. EZH2 Inhibition in Ewing Sarcoma Upregulates G(D2) Expression for Targeting with Gene-Modified T Cells. Mol. Ther. 2019, 27, 933–946. [Google Scholar] [CrossRef]
- Seo, W.; Jerin, C.; Nishikawa, H. Transcriptional regulatory network for the establishment of CD8(+) T cell exhaustion. Exp. Mol. Med. 2021, 53, 202–209. [Google Scholar] [CrossRef]
- Yang, R.; Cheng, S.; Luo, N.; Gao, R.; Yu, K.; Kang, B.; Wang, L.; Zhang, Q.; Fang, Q.; Zhang, L.; et al. Distinct epigenetic features of tumor-reactive CD8+ T cells in colorectal cancer patients revealed by genome-wide DNA methylation analysis. Genome Biol. 2019, 21, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; He, J.; Xu, S.; Wang, Q.; Cheng, J. NR4A1 promotes LEF1 expression in the pathogenesis of papillary thyroid cancer. Cell Death Discov. 2022, 8, 46. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.A.; Weinmann, A.S. Molecular mechanisms by which T-bet regulates T-helper cell commitment. Immunol. Rev. 2010, 238, 233–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Istaces, N.; Splittgerber, M.; Lima Silva, V.; Nguyen, M.; Thomas, S.; Le, A.; Achouri, Y.; Calonne, E.; Defrance, M.; Fuks, F.; et al. EOMES interacts with RUNX3 and BRG1 to promote innate memory cell formation through epigenetic reprogramming. Nat. Commun. 2019, 10, 3306. [Google Scholar] [CrossRef] [Green Version]
- Itahashi, K.; Irie, T.; Yuda, J.; Kumagai, S.; Tanegashima, T.; Lin, Y.T.; Watanabe, S.; Goto, Y.; Suzuki, J.; Aokage, K.; et al. BATF epigenetically and transcriptionally controls the activation program of regulatory T cells in human tumors. Sci. Immunol. 2022, 7, eabk0957. [Google Scholar] [CrossRef]
- Chang, S.; Collins, P.L.; Aune, T.M. T-bet dependent removal of Sin3A-histone deacetylase complexes at the Ifng locus drives Th1 differentiation. J. Immunol. 2008, 181, 8372–8381. [Google Scholar] [CrossRef] [Green Version]
- Tumes, D.J.; Onodera, A.; Suzuki, A.; Shinoda, K.; Endo, Y.; Iwamura, C.; Hosokawa, H.; Koseki, H.; Tokoyoda, K.; Suzuki, Y.; et al. The polycomb protein Ezh2 regulates differentiation and plasticity of CD4(+) T helper type 1 and type 2 cells. Immunity 2013, 39, 819–832. [Google Scholar] [CrossRef] [Green Version]
- Kurachi, M.; Barnitz, R.A.; Yosef, N.; Odorizzi, P.M.; DiIorio, M.A.; Lemieux, M.E.; Yates, K.; Godec, J.; Klatt, M.G.; Regev, A.; et al. The transcription factor BATF operates as an essential differentiation checkpoint in early effector CD8+ T cells. Nat. Immunol. 2014, 15, 373–383. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| NCT Identifier | Malignant Conditions | Therapeutics (Single or Combined) | Start Year | Status |
|---|---|---|---|---|
| NCT05089370 | •Malignant Melanoma | •Combination Product: Oral Decitabine/Cedazuridine (DNMT inhibitor) in Combination with Nivolumab (PD-1 inhibitor) | 2022 | Recruiting |
| NCT04705818 | •Advanced Solid Tumor •Advanced Colorectal Carcinoma •Advanced Soft tissue Sarcoma •Advanced Pancreatic Adenocarcinoma •Adult Solid Tumor | •Drug: Durvalumab (PD-L1 inhibitor) •Drug: Tazemetostat (EZH2 inhibitor) | 2021 | Recruiting |
| NCT04648826 | •Sarcomas •Melanomas •Germ Cell Tumors •Epithelial Malignancies (Excluding Lung and Renal Cell Carcinomas) •Pulmonary Metastases | •Drug: Bintrafusp alfa (bifunctional fusion protein composed of the extracellular domain of the TGF-receptor II fused to an IgG1 antibody blocking PD-L1) •Drug: Azacytidine (DNMT1 inhibitor) | 2021 | Withdrawn |
| NCT04190056 | •Anatomic Stage IV Breast Cancer AJCC v8 •Prognostic Stage IV Breast Cancer AJCC v8 | •Biological: Pembrolizumab (PD-1 inhibitor) •Drug: Tamoxifen (antiestrogen) •Drug: Vorinostat (HDAC inhibitor) | 2021 | Recruiting |
| NCT04471974 | •Castration-Resistant Prostate Carcinoma •Metastatic Prostate Adenocarcinoma •Metastatic Prostate Small Cell Carcinoma •Stage IV Prostate Cancer AJCC v8 •Stage IVA Prostate Cancer AJCC v8 •Stage IVB Prostate Cancer AJCC v8 | •Drug: ZEN-3694 (BET bromodomain inhibitor) •Drug: Enzalutamide (nonsteroidal antiandrogen (NSAA) medication) •Biological: Pembrolizumab (PD-1 inhibitor) | 2021 | Recruiting |
| NCT04708470 | •Cancer •Solid Tumor •Metastatic Checkpoint Refractory HPV-Associated Malignancies •Microsatellite Stable Colon Cancer (MSS) | •Drug: Bintrafusp Alfa (bifunctional fusion protein composed of the extracellular domain of the TGF-receptor II fused to an IgG1 antibody blocking PD-L1) •Drug: NHS-IL12 (tumor-targeting immunocytokine) •Drug: Entinostat (HDAC inhibitor) | 2021 | Recruiting |
| NCT04257448 | •Pancreas Cancer •Pancreatic Adenocarcinoma •Pancreatic Ductal Adenocarcinoma | •Drug: Romidepsin (HDAC inhibitor) •Drug: Azacitidine (DNMT inhibitor) •Drug: Nab-Paclitaxel (stops cancer cells from separating into two new cells) •Drug: Gemcitabine (induces interferon signaling) •Drug: Durvalumab (PD-L1 inhibitor) •Drug: Lenalidomide capsule (potent molecular analog of thalidomide) | 2020 | Recruiting |
| NCT04611711 | •Patients With Digestive System Tumors Resistant to PD-1 Inhibitors | •Drug: Decitabine (DNMT inhibitor) + TQB2450 injection (PD-1 inhibitor) •Drug: Decitabine (DNMT inhibitor) + TQB2450 injection (PD-1 inhibitor) + Anlotinib (VEGFR inhibitor) | 2020 | Not yet recruiting |
| NCT04553393 | •Refractory or Relapsed Aggressive r/r BNHL With Huge Tumor Burden | •Drug: Chidamide (HDAC inhibitor) •Drug: Decitabine (DNMT inhibitor) •Biological: Decitabine-Primed Tandem CAR19/20-Engineered T Cells | 2020 | Recruiting |
| NCT04407741 | •Solid Tumor •Lymphoma | •Drug: SHR2554 (EZH2 inhibitor) •Drug: SHR1701 (PD-1 and TGF-β inhibitor) | 2020 | Recruiting |
| NCT04414969 | •Immune Checkpoint Inhibitor •Chemotherapy Effect •Epigenetic Disorder •NK/T-Cell Lymphoma of Nasal Cavity | •Drug: Anti-PD-1 antibody + Peg-Asparaginase + Chidamide (HDAC inhibitor) | 2020 | Recruiting |
| NCT04250246 | •Melanoma •Non-Small Cell Lung Cancer | •Drug: Ipilimumab (CTLA-4 antibody) •Biological: Nivolumab (PD-1 inhibitor) •Drug: Guadecitabine (DNMT inhibitor) | 2020 | Not yet recruiting |
| NCT04277442 | •Acute Myeloid Leukemia | •Drug: Decitabine (DNMT inhibitor) •Biological: Nivolumab (PD-1 inhibitor) •Drug: Venetoclax (Bcl-2 inhibitor) | 2020 | Suspended |
| NCT03812796 | •Cancer •GI Cancer | •Drug: Domatinostat (HDAC inhibitor) •Drug: Avelumab (PD-1 inhibitor) | 2019 | Unknown status |
| NCT03765229 | •Melanoma | •Drug: Entinostat (HDAC inhibitor) •Drug: Pembrolizumab (PD-1 inhibitor) | 2019 | Recruiting |
| NCT03854474 | •Locally Advanced Urothelial Carcinoma •Metastatic Urothelial Carcinoma •Stage III Bladder Cancer AJCC v8 •Stage IIIA Bladder Cancer AJCC v8 •Stage IIIB Bladder Cancer AJCC v8 •Stage IV Bladder Cancer AJCC v8 •Stage IVA Bladder Cancer AJCC v8 •Stage IVB Bladder Cancer AJCC v8 | •Biological: Pembrolizumab (PD-1 inhibitor) •Drug: Tazemetostat (EZH2 inhibitor) | 2019 | Recruiting |
| NCT03233724 | •Non-Small-Cell Lung Carcinoma •Lung Cancer •Non-Small Cell Lung Cancer •Esophageal Carcinoma •Malignant Pleural Mesotheliomas | •Drug: Decitabine (DNMT inhibitor) •Drug: Tetrahydrouridine (inhibitor of cytidine deaminase) •Drug: Pembrolizumab (PD-1 inhibitor) | 2018 | Recruiting |
| NCT03445858 | •Childhood Solid Tumor •Childhood Lymphoma •Relapsed Cancer •Refractory Cancer •Adult Solid Tumor •Adult Lymphoma | •Drug: Pembrolizumab (PD-1 inhibitor) •Drug: Decitabine (DNMT inhibitor) | 2018 | Active, not recruiting |
| NCT03161223 | •T-Cell Lymphoma | •Drug: Durvalumab (PD-L1 inhibitor) •Drug: Pralatrexate (dihydrofolate reductase inhibitor) •Drug: Romidepsin (HDAC inhibitor) •Drug: 5-Azacitidine (Methyltransferase inhibitor) | 2018 | Recruiting |
| NCT02664181 | •Lung Cancer •Non-Small Cell Lung Cancer | •Drug: Nivolumab (PD-1 inhibitor) •Drug: Oral decitabine (DNMT inhibitor) •Drug: Tetrahydrouridine (inhibitor of cytidine deaminase) | 2017 | Active, not recruiting |
| NCT03206047 | •Platinum-Resistant Fallopian Tube Carcinoma •Platinum-Resistant Ovarian Carcinoma •Platinum-Resistant Primary Peritoneal Carcinoma •Recurrent Fallopian Tube Carcinoma •Recurrent Ovarian Carcinoma •Recurrent Primary Peritoneal Carcinoma | •Drug: Atezolizumab (PD-L1 inhibitor) •Biological: DEC-205/NYESO-1 Fusion Protein CDX-1401 (vaccine that may help the immune system specifically target and kill cancer cells) •Drug: Guadecitabine (DNMT inhibitor) •Drug: Poly ICLC (induces immunohematopoietic cells) | 2017 | Active, not recruiting |
| NCT03250273 | •Metastatic Cholangiocarcinoma •Cholangiocarcinoma •Pancreatic Cancer •Metastatic Pancreatic Cancer •Unresectable Pancreatic Cancer •Unresectable Cholangiocarcinoma | •Drug: Entinostat (HDAC inhibitor) •Drug: Nivolumab (PD-1 inhibitor) | 2017 | Completed |
| NCT02915523 | •Epithelial Ovarian Cancer •Peritoneal Cancer •Fallopian Tube Cancer | •Drug: Entinostat (HDAC inhibitor) •Drug: Avelumab (PD-1 inhibitor) | 2017 | Unknown status |
| NCT03024437 | •Metastatic Cancer •Renal Cancer | •Drug: Atezolizumab (PD-L1 inhibitor) •Drug: Bevacizumab (VEGF inhibitor) •Drug: Entinostat (HDAC inhibitor) | 2017 | Suspended |
| NCT02437136 | Non-Small Cell Lung Cancer •Melanoma •Mismatch Repair-Proficient Colorectal Cancer | •Drug: Entinostat (HDAC inhibitor) •Drug: Pembrolizumab (PD-1 inhibitor) | 2017 | Active, not recruiting |
| NCT02959437 | •Solid Tumors •Advanced Malignancies •Metastatic Cancer | •Drug: Azacitidine (DNMT inhibitor) •Drug: Pembrolizumab (PD-1 inhibitor) •Drug: Epacadostat (indoleamine2,3-dioxygenase inhibitor) •Drug: INCB057643 (BET inhibitor) •Drug: INCB059872 (LSD1 inhibitor) | 2017 | Terminated |
| NCT02816021 | •Melanoma and Other Malignant Neoplasms of Skin •Metastatic Melanoma | •Drug: Azacitidine (DNMT inhibitor) •Drug: Pembrolizumab (PD-1 inhibitor) | 2017 | Active, not recruiting |
| NCT02890329 | •Previously Treated Myelodysplastic Syndrome •Recurrent Acute Myeloid Leukemia •Recurrent Acute Myeloid Leukemia with Myelodysplasia Related Changes •Recurrent Myelodysplastic Syndrome •Refractory Acute Myeloid Leukemia •Refractory Myelodysplastic Syndrome •Secondary Acute Myeloid Leukemia •Secondary Myelodysplastic Syndrome | •Drug: Decitabine (DNMT inhibitor) •Biological: Ipilimumab (CTLA-4 antibody) | 2017 | Active, not recruiting |
| NCT03019003 | •Head and Neck Cancer | •Drug: Oral Decitabine (DNMT inhibitor) •Drug: Durvalumab (PD-L1 inhibitor) | 2017 | Active, not recruiting |
| NCT03066648 | •Leukemia •Myeloid Leukemia •Acute Myeloid Leukemia •Myelodysplastic Syndromes •Preleukemia •Bone Marrow Diseases •Hematologic Diseases •Chronic Myelomonocytic Leukemia | •Drug: Decitabine (DNMT inhibitor) •Drug: PDR001 (PD-1 antibody) •Drug: MBG453 (Tim3 antibody) •Drug: Azacitidine (DNMT inhibitor) | 2017 | Active, not recruiting |
| NCT02951156 | •Diffuse Large B-cell Lymphoma | •Biological: Avelumab (PD-1 inhibitor) •Biological: Utomilumab (binds to CD-137 protein receptor and stimulates/increases the number of immune cells) •Biological: Rituximab (chimeric monoclonal antibody against the protein CD20) •Other: Azacitidine (DNMT inhibitor) •Drug: Bendamustine (chemotherapy medication) •Drug: Gemcitabine (induces interferon signaling) •Drug: Oxaliplatin (inhibits the synthesis of deoxyribonucleic acid (DNA)) | 2016 | Terminated |
| NCT02900560 | •Epithelial Ovarian Cancer | •Drug: CC-486 (hypomethylation of DNA) •Biological: Pembrolizumab (PD-1 inhibitor) | 2016 | Terminated |
| NCT02512172 | •Colorectal Cancer | •Drug: Oral CC-486 (hypomethylation of DNA) •Drug: Romidepsin (HDAC inhibitor) •Drug: MK-3475 (PD-1 inhibitor) | 2016 | Completed |
| NCT02395627 | Breast Neoplasms | •Drug: Tamoxifen (antiestrogens) •Drug: Vorinostat (HDAC inhibitor) •Drug: Pembrolizumab (PD-1 inhibitor) | 2015 | Terminated |
| NCT02608437 | •Metastatic Melanoma | •Drug: SGI-110 (DNA methylation inhibitor) •Drug: Ipilimumab (CTLA-4 antibody) | 2015 | Unknown status |
| NCT02453620 | •Breast Adenocarcinoma •Invasive Breast Carcinoma •Metastatic Breast Carcinoma •Metastatic Malignant Solid Neoplasm •Stage III Breast Cancer AJCC v7 •Stage IIIA Breast Cancer AJCC v7 •Stage IIIB Breast Cancer AJCC v7 •Stage IIIC Breast Cancer AJCC v7 •Stage IV Breast Cancer AJCC v6 and v7 •Unresectable Solid Neoplasm | •Drug: Entinostat (HDAC inhibitor) •Biological: Ipilimumab (CTLA-4 antibody) •Biological: Nivolumab (PD-1 inhibitor) | 2015 | Active, not recruiting |
| NCT02546986 | •Non-Small-Cell Lung Carcinoma | •Drug: CC-486 (hypomethylation of DNA) •Drug: Pembrolizumab (PD-1 inhibitor) | 2015 | Active, not recruiting |
| NCT02397720 | •Acute Bilineal Leukemia •Acute Biphenotypic Leukemia •Acute Myeloid Leukemia Arising from Previous Myelodysplastic Syndrome •Chronic Myelomonocytic Leukemia •Myelodysplastic Syndrome •Recurrent Acute Myeloid Leukemia •Refractory Acute Myeloid Leukemia •Secondary Acute Myeloid Leukemia •Therapy-Related Acute Myeloid Leukemia | •Drug: Azacitidine (DNMT inhibitor) •Biological: Ipilimumab (CTLA-4 antibody) •Biological: Nivolumab (PD-1 inhibitor) | 2015 | Recruiting |
| NCT02608268 | •Advanced Malignancies | •Drug: MBG453 (Tim3 antibody) •Drug: PDR001 (PD-1 antibody) •Drug: Decitabine (DNMT inhibitor) | 2015 | Active, not recruiting |
| NCT01834248 | •Acute Myeloid Leukemia •Alkylating Agent-Related Acute Myeloid Leukemia •Chronic Myelomonocytic Leukemia •Myelodysplastic Syndrome •Refractory Anemia with Excess Blasts | •Biological: DEC-205/NYESO-1 Fusion Protein CDX-1401 (vaccine that may help the immune system specifically target and kill cancer cells) •Drug: Decitabine (DNMT inhibitor) •Drug: Poly ICLC (induces immunohematopoietic cells) | 2013 | Completed |
| NCT01928576 | •Non-Small Cell Lung Cancer •Epigenetic Therapy | •Drug: Azacitidine (DNMT inhibitor) •Drug: Entinostat (HDAC inhibitor) •Drug: Nivolumab (PD-1 inhibitor) | 2013 | Recruiting |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keshari, S.; Barrodia, P.; Singh, A.K. Epigenetic Perspective of Immunotherapy for Cancers. Cells 2023, 12, 365. https://doi.org/10.3390/cells12030365
Keshari S, Barrodia P, Singh AK. Epigenetic Perspective of Immunotherapy for Cancers. Cells. 2023; 12(3):365. https://doi.org/10.3390/cells12030365
Chicago/Turabian StyleKeshari, Sunita, Praveen Barrodia, and Anand Kamal Singh. 2023. "Epigenetic Perspective of Immunotherapy for Cancers" Cells 12, no. 3: 365. https://doi.org/10.3390/cells12030365