

Cellular and Subcellular Characteristics of Neuromuscular Junctions in Muscles with Disparate Duty Cycles and Myofiber Profiles
Abstract
:1. Introduction
2. Experimental Procedures
2.1. Animal Care and Muscle Selection
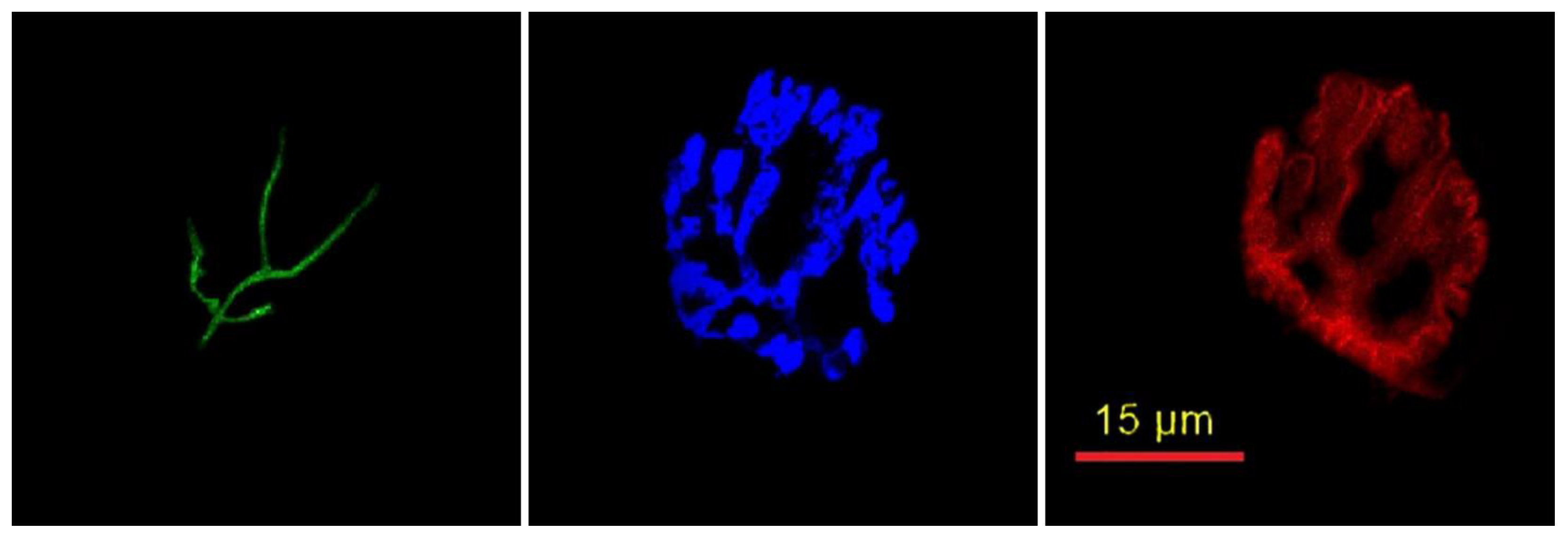
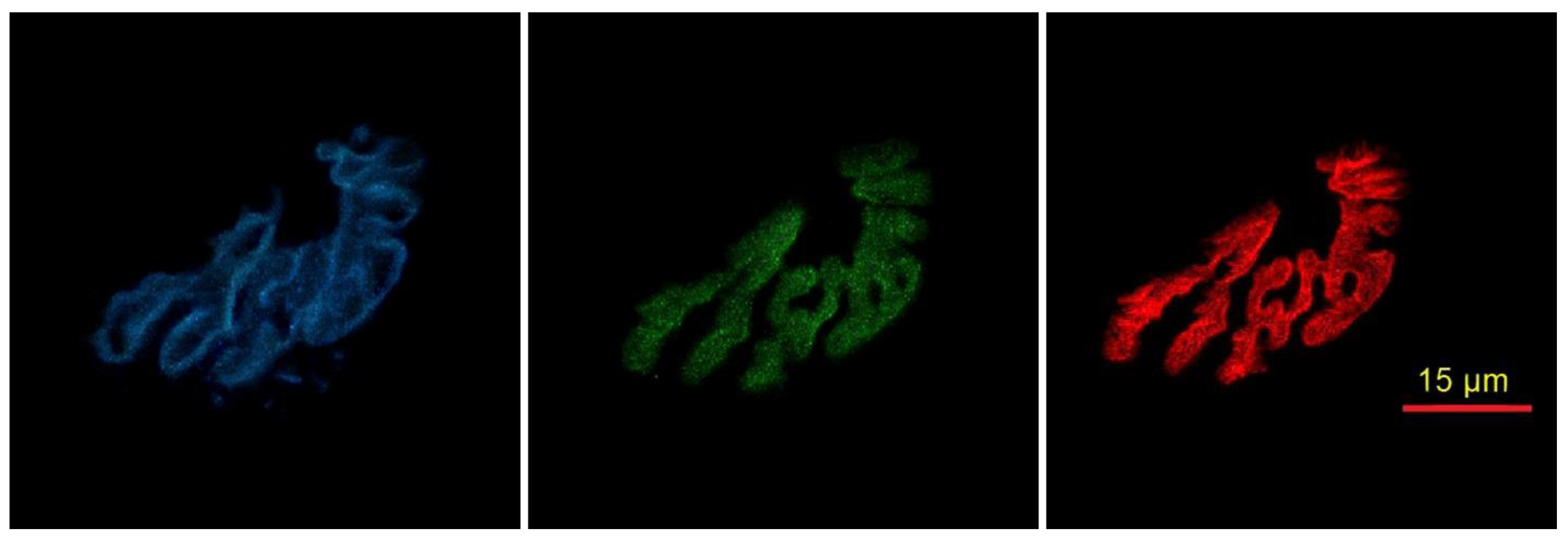
2.2. Cytofluorescent Staining of Cellular Components of the NMJ
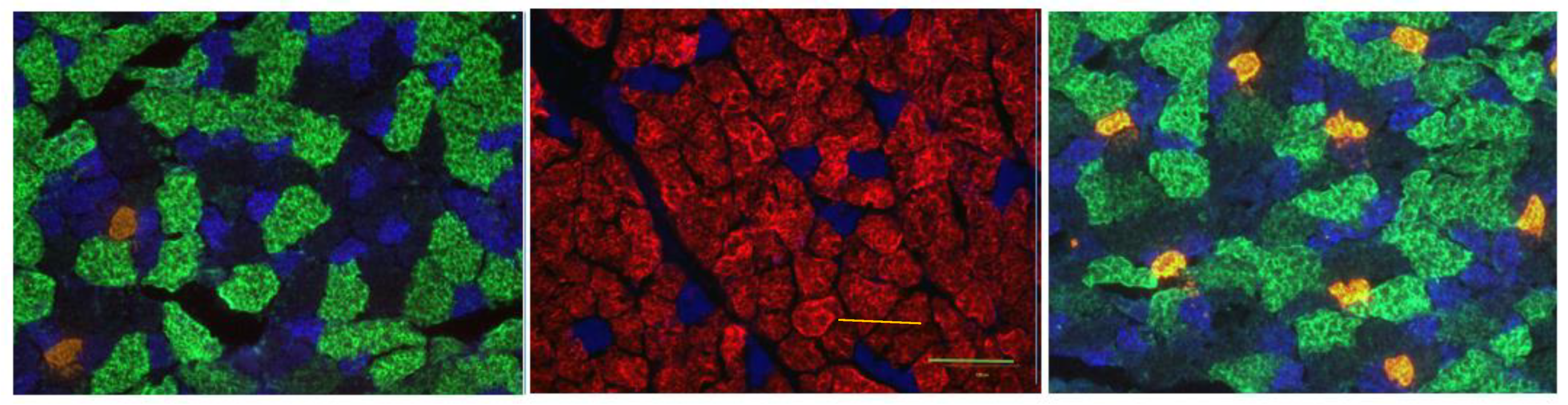
2.3. Immunofluorescent Staining of Myofibers
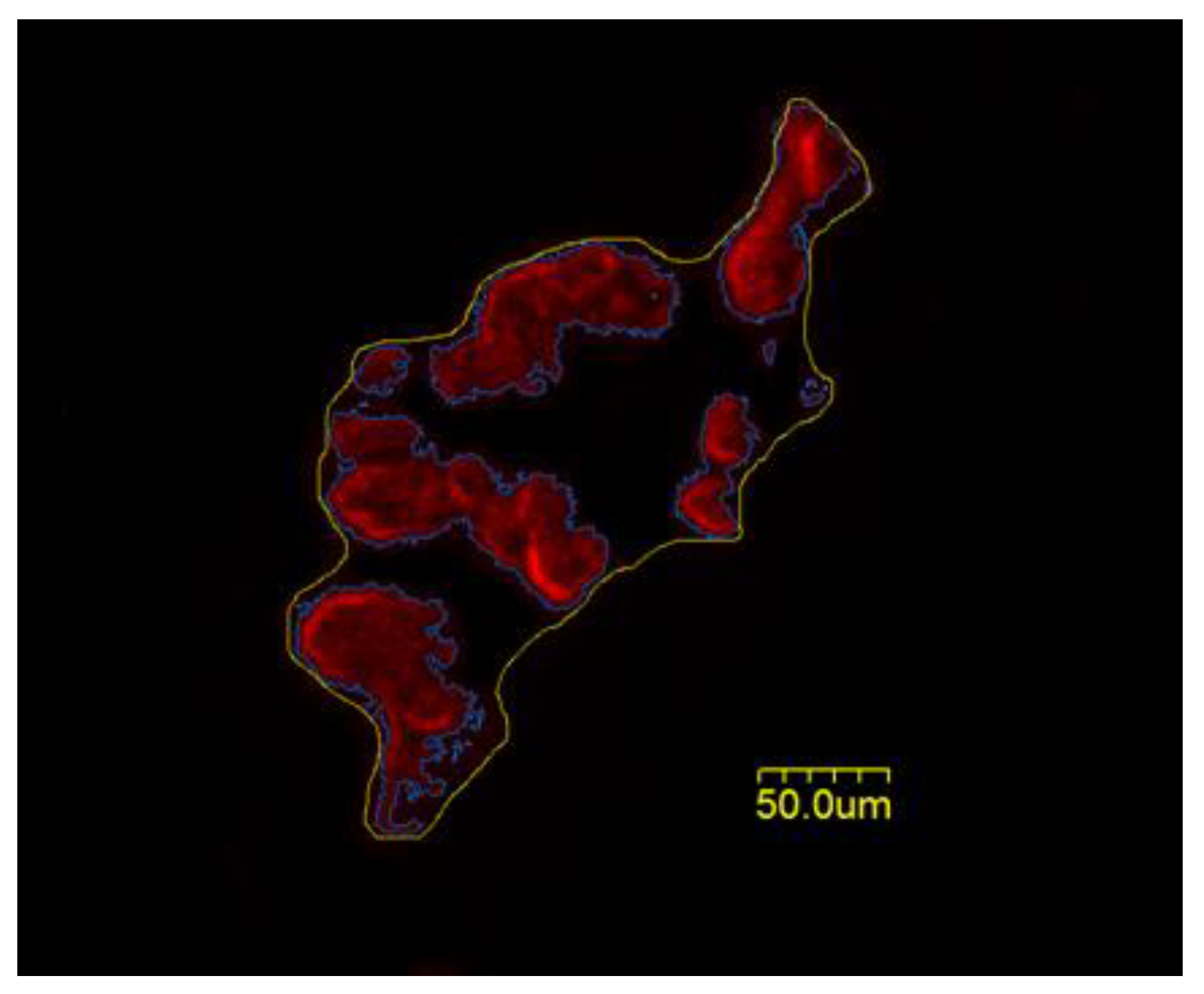
2.4. Microscopy
2.5. Statistical Analysis
3. Results
Cellular NMJ Results
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ginebaugh, S.P.; Badawi, Y.; Tarr, T.B.; Meriney, S.D. Neuromuscular Active Zone Structure and Function in Healthy and Lambert-Eaton Myasthenic Syndrome States. Biomolecules 2022, 12, 740. [Google Scholar] [CrossRef]
- Herrera, A.A.; Grinnell, A.D. Contralateral denervation causes enhanced transmitter release from frog motor nerve terminals. Nature 1981, 291, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Deschenes, M.R.; Tufts, H.L.; Oh, J.; Li, S.; Noronha, A.L.; Adan, M.A. Effects of exercise training on neuromuscular junctions and their active zones in young and aged muscles. Neurobiol. Aging 2020, 95, 1–8. [Google Scholar] [CrossRef]
- Deschenes, M.R.; Patek, L.G.; Trebelhorn, A.M.; High, M.C.; Flannery, R.E. Juvenile Neuromuscular Systems Show Amplified Disturbance to Muscle Unloading. Front. Physiol. 2021, 12, 754052. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, M.H.; Korthuis, R.J.; Sexton, W.L.; Armstrong, R.B. Regional muscle blood flow capacity and exercise hyperemia in high-intensity trained rats. J. Appl. Physiol. 1988, 64, 2420–2427. [Google Scholar] [CrossRef]
- Dum RP, Kennedy TT. Physiological and histochemical characteristics of motor units in cat tibialis anterior and extensor digi-torum longus muscles. J. Neurophysiol. 1980, 43, 1615–1630. [Google Scholar] [CrossRef] [Green Version]
- Roy, R.R.; Hutchison, D.L.; Pierotti, D.J.; Hodgson, J.A.; Edgerton, V.R. EMG patterns of rat ankle extensors and flexors during treadmill locomotion and swimming. J. Appl. Physiol. 1991, 70, 2522–2529. [Google Scholar] [CrossRef] [PubMed]
- Delp, M.; Duan, C. Composition and size of type I, IIA, IID/X, and IIB fibers and citrate synthase activity of rat muscle. J. Appl. Physiol. 1996, 80, 261–270. [Google Scholar] [CrossRef]
- Pearson, J.; Sabarra, A. A Method for Obtaining Longitudinal Cryostat Sections of Living Muscle Without Contraction Artifacts. Stain. Technol. 1974, 49, 143–146. [Google Scholar] [CrossRef]
- Anderton, B.H.; Breinburg, D.; Downes, M.J.; Green, P.J.; Tomlinson, B.E.; Ulrich, J.; Wood, J.N.; Kahn, J. Monoclonal antibodies show that neurofibrillary tangles and neurofilaments share antigenic determinants. Nature 1982, 298, 84–86. [Google Scholar] [CrossRef]
- Kwon, S.E.; Chapman, E.R. Synaptophysin Regulates the Kinetics of Synaptic Vesicle Endocytosis in Central Neurons. Neuron 2011, 70, 847–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossio-Bolanos, M.; Gomez Campos, R.; Vargas Vitoria, R.; Hochmuller Fogaca, R.T.; de Arruda, M. Reference curves for assessing the physical growth of male Wistar rats. Nutr. Hosp. 2013, 28, 2151–2156. [Google Scholar] [PubMed]
- Armstrong, R.B.; Laughlin, M.H. Rat muscle blood flows during high-speed locomotion. J. Appl. Physiol. 1985, 59, 1322–1328. [Google Scholar] [CrossRef]
- Dudley, G.A.; Abraham, W.M.; Terjung, R.L. Influence of exercise intensity and duration on biochemical adaptations in skeletal muscle. J. Appl. Physiol. 1982, 53, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Silverthorn, D.U. Homeostasis nd Control: Muscles. In Human Physiology: An Integrated Approach; Pearson: New York, NY, USA, 2013; pp. 398–440. [Google Scholar]
- Lomo, T.; Rosenthal, J. Control of ACh sensitivity by muscle activity in the rat. J. Physiol. 1972, 221, 493–513. [Google Scholar] [CrossRef] [Green Version]
- Pagala, M.K.D.; Namba, T.; Grob, D. Failure of neuromuscular transmission and contractility during muscle fatigue. Muscle Nerve 1984, 7, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Fahim, M.A. Endurance exercise modulates neuromuscular junction of C57BL/6NNia aging mice. J. Appl. Physiol. 1997, 83, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Grinnell, A.D.; Herrera, A. Physiological regulation of synaptic effectiveness at frog neuromuscular junctions. J. Physiol. 1980, 307, 301–317. [Google Scholar] [CrossRef]
- Prakash, Y.; Sieck, G.C. Age-related remodeling of neuromuscular junctions on type-identified diaphragm fibers. Muscle Nerve 1998, 21, 887–895. [Google Scholar] [CrossRef]
- Katz, B.; Miledi, R. Estimates of quantal content during ’chemical potentiation’ of transmitter release. Proc. R. Soc. Lond. Ser. B Boil. Sci. 1979, 205, 369–378. [Google Scholar] [CrossRef]
- Nishimune, H.; Sanes, J.R.; Carlson, S.S. A synaptic laminin-calcium channel interaction organizes active zones in motor nerve terminals. Nature 2004, 432, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Nishimune, H.; Badawi, Y.; Mori, S.; Shigemoto, K. Dual-color STED microscopy reveals a sandwich structure of Bassoon and Piccolo in active zones of adult and aged mice. Sci. Rep. 2016, 6, 27935. [Google Scholar] [CrossRef] [Green Version]
- Laghaei, R.; Meriney, S.D. Microphysiological Modeling of the Structure and Function of Neuromuscular Transmitter Release Sites. Front. Synaptic Neurosci. 2022, 14, 917285. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chakkalakal, J.V. The Composition, Development, and Regeneration of Neuromuscular Junctions. Curr. Top. Dev. Biol. 2018, 126, 99–124. [Google Scholar] [CrossRef] [PubMed]
- Bao, Z.; Cui, C.; Chow, S.K.-H.; Qin, L.; Wong, R.M.Y.; Cheung, W.-H. AChRs Degeneration at NMJ in Aging-Associated Sarcopenia–A Systematic Review. Front. Aging Neurosci. 2020, 12, 597811. [Google Scholar] [CrossRef]
- Chen, J.; Billings, S.E.; Nishimune, H. Calcium channels link the muscle-derived synapse organizer laminin beta2 to Bassoon and CAST/Erc2 to organize presynaptic active zones. J. Neurosci. 2011, 31, 512–525. [Google Scholar] [CrossRef] [Green Version]
- Pierotti, D.J.; Roy, R.R.; Gregor, R.J.; Edgerton, V.R. Electromyographic activity of cat hindlimb flexors and extensors during locomotion at varying speeds and inclines. Brain Res. 1989, 481, 57–66. [Google Scholar] [CrossRef]
- Steinbach, J.H.; Sine, S.M. Function of nicotinic acetylcholine receptors. Soc. Gen. Physiol. Ser. 1987, 41, 19–42. [Google Scholar]
- Glavinovic, M.I. Postsynaptic potentiation and desensitization in frog neuromuscular junction. Can. J. Physiol. Pharmacol. 1988, 66, 624–629. [Google Scholar] [CrossRef]
- Burden, S.J.; Huijbers, M.G.; Remedio, L. Fundamental Molecules and Mechanisms for Forming and Maintaining Neuromuscular Synapses. Int. J. Mol. Sci. 2018, 19, 490. [Google Scholar] [CrossRef] [Green Version]
- Oda, K. The relationship between motor endplate size and musxcle fiber diameter in different muslce groups of the rat. Jpn. J. Physiol. 1985, 35, 1091–1095. [Google Scholar] [CrossRef] [PubMed]
- Balice-Gordon, R.J.; Lichtman, J.W. In vivo visualization of the growth of pre- and postsynaptic elements of neuromuscular junc-tions in the mouse. J. Neurosci. 1990, 10, 894–908. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | EDL | Soleus | Plantaris |
|---|---|---|---|
| Nerve terminal branch number | 5.9 ± 0.3 | 7.6 ± 0.3 * | 6.0 ± 0.6 |
| Total branch length (µm) | 129.8 ± 1.0 | 145.6 ± 0.6 | 100.4 ± 5.9 * |
| Average branch length (µm2) | 22.9 ± 29.4 * | 19.5 ± 0.6 | 18.9 ± 1.4 |
| Branching complexity | 8.5 ± 0.8 | 11.7 ± 0.8 * | 7.1 ± 1.3 |
| Vesicle/Branch length | 0.667 ± 0.08 | 0.959 ± 0.07 † | 0.884 ± 0.09 |
| Total length around vesicles (µm) | 140.8 ± 5.2 | 163.5 ± 10.8 | 143.7 ± 9.5 |
| Stained vesicle perimeter (µm) | 180.1 ± 14.1 | 239.5 ± 18.4 * | 160.3 ± 8.0 |
| Total vesicle area (µm2) | 421.9 ± 52.1 | 510.9 ± 33.7 ‡ | 351.5 ± 21.5 |
| Stained vesicle area (µm2) | 79.7 ± 9.1 | 138.3 ± 12.5 * | 77.1 ± 6.0 |
| Vesicle dispersion (%) | 23.0 ± 2.6 | 27.8 ± 1.5 | 24.8 ± 2.7 |
| Total BTX perimeter (µm) | 95.3 ± 6.8 | 319.0 ± 17.7 * | 90.6 ± 5.2 |
| Stained BTX perimeter (µm) | 224.5 ± 16.1 | 315.9 ± 19.9 * | 237.5 ± 25.3 |
| Total BTX area (µm2) | 372.5 ± 29.4 | 401.7 ± 24.8 | 354.8 ± 30.4 |
| Stained BTX area (µm2) | 191.2 ± 35.1 | 272.8 ± 17.1 * | 145.7 ± 15.9 |
| BTX dispersion (%) | 49.1 ± 5.9 | 67.8 ± 1.6 * | 42.1 ± 2.8 |
| Variable | EDL | Soleus | Plantaris |
|---|---|---|---|
| Total Bassoon perimeter (µm) | 91.3 ± 7.5 | 282.8 ± 16.9 * | 86.9 ± 4.2 |
| Stained Bassoon perimeter (µm) | 328.6 ± 49.9 | 359.8 ± 23.4 | 605.1 ± 45.8 * |
| Total Bassoon area (µm2) | 362.4 ± 38.8 | 357.9 ± 24.4 | 358.4 ± 29.6 |
| Stained Bassoon area (µm2) | 180.4 ± 39.6 | 189.8 ± 13.8 | 104.9 ± 11.4 * |
| Bassoon dispersion (%) | 46.6 ± 6.3 | 53.3 ± 2.3 | 27.9 ± 1.5 * |
| Total P/Q perimeter (µm) | 91.4 ± 4.9 | 221.8 ± 17.7 * | 90.6 ± 5.2 |
| Stained P/Q perimeter (µm) | 230.0 ± 11.1 * | 319.0 ± 22.4 | 306.4 ± 30.3 |
| Total P/Q area (µm2) | 377.5 ± 30.7 | 369.5 ± 47.8 | 409.9 ± 43.4 |
| Stained P/Q area (µm2) | 172.9 ± 35.2 | 274.9 ± 27.2 * | 151.8 ± 16.3 |
| P/Q dispersion (%) | 42.6 ± 5.5 | 80.6 ± 7.8 * | 38.6 ± 2.1 |
| BTX stained area/Bassoon stained area | 0.92 ± 0.10 * | 1.45 ± 0.05 | 1.43 ± 0.13 |
| P/Q stained area/Bassoon stained area | 1.00 ± 0.07 * | 1.48 ± 0.13 | 1.46 ± 0.10 |
| BTX stained area/P/Q stained area | 1.25 ± 0.21 | 1.03 ± 0.06 | 0.97 ± 0.05 |
| Variable | EDL | Soleus | Plantaris |
|---|---|---|---|
| Cross-sectional area (µm2) | |||
| Types combined * | 2181 ± 155 | 2862 ± 112 | 3720 ± 223 |
| Type I * | 1284 ± 61 | 3001 ± 121 | 2006 ± 170 |
| Type IIA | 1238 ± 56 ‡ | 2246 ± 94 | 2141 ± 155 |
| Type IIX | 2030 ± 125 | 2340 ± § | 3776 ± 331 ‡# |
| Type IIB * | 3063 ± 244 | N/A | 5289 ± 683 |
| Fiber type composition (%) | |||
| Type I * | 6.0 ± 0.8 | 80.7 ± 1.3 | 10.2 ± 1.1 |
| Type IIA | 27.8 ± 3.0 | 18.1 ± 1.2 ‡ | 27.7 ± 2.8 |
| Type IIX | 30.5 ± 2.1 | 1.3 ± § ‡ | 29.3 ± 2.9 |
| Type IIB | 35.6 ± 4.5 | N/A | 32.7 ± 3.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deschenes, M.R.; Mifsud, M.K.; Patek, L.G.; Flannery, R.E. Cellular and Subcellular Characteristics of Neuromuscular Junctions in Muscles with Disparate Duty Cycles and Myofiber Profiles. Cells 2023, 12, 361. https://doi.org/10.3390/cells12030361
Deschenes MR, Mifsud MK, Patek LG, Flannery RE. Cellular and Subcellular Characteristics of Neuromuscular Junctions in Muscles with Disparate Duty Cycles and Myofiber Profiles. Cells. 2023; 12(3):361. https://doi.org/10.3390/cells12030361
Chicago/Turabian StyleDeschenes, Michael R., Mia K. Mifsud, Leah G. Patek, and Rachel E. Flannery. 2023. "Cellular and Subcellular Characteristics of Neuromuscular Junctions in Muscles with Disparate Duty Cycles and Myofiber Profiles" Cells 12, no. 3: 361. https://doi.org/10.3390/cells12030361