

Neuroprotective Action of Tacrolimus before and after Onset of Neonatal Hypoxic–Ischaemic Brain Injury in Rats
 , ,
, ,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Animal Handling
2.3. Experimental Groups
2.4. Animal Surgery
2.5. Post-Mortem and Tissue Processing
2.6. Cytokine Analysis
2.7. Histology
2.8. Immunohistochemistry
2.9. Statistics
3. Results
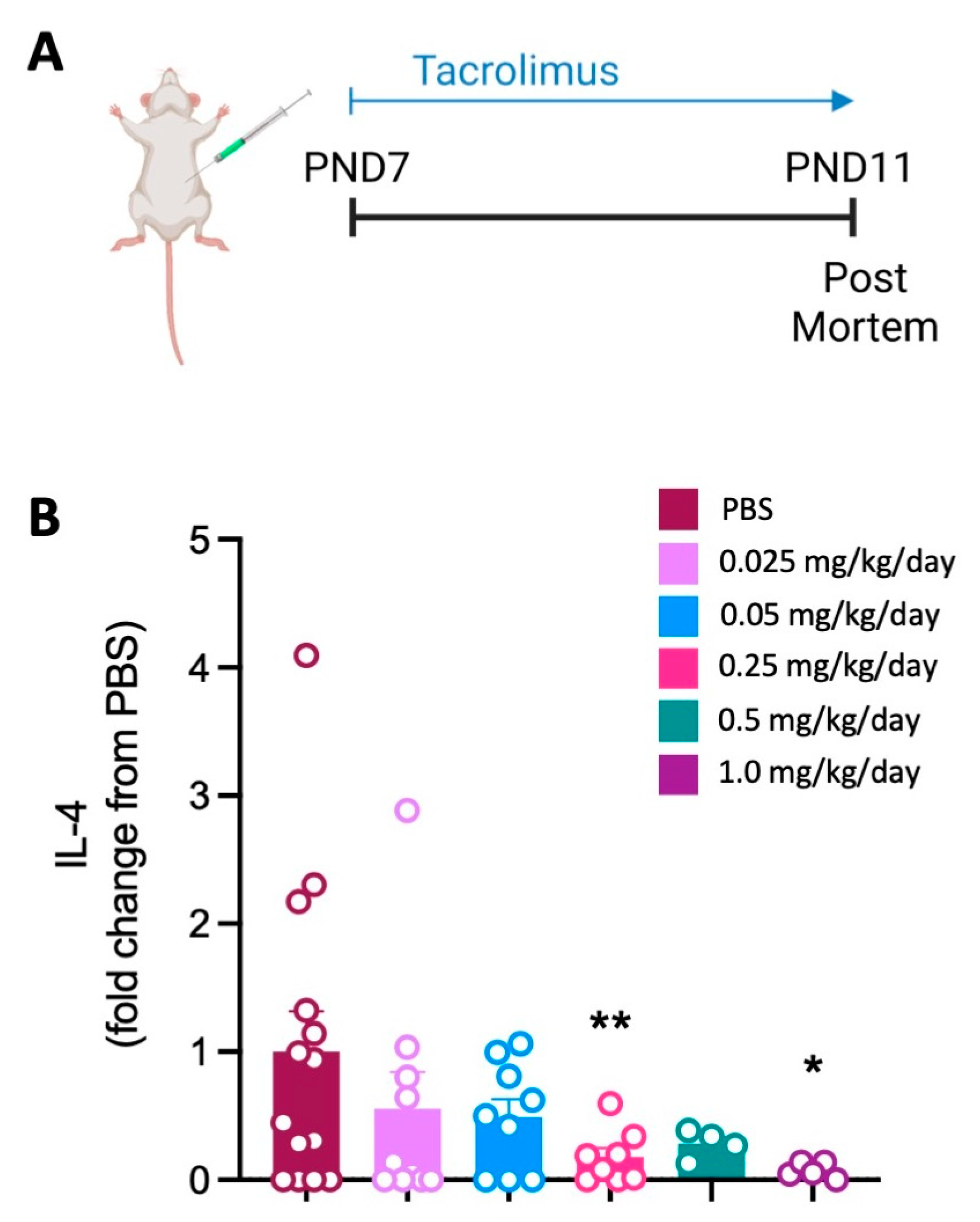
3.1. Tacrolimus Reduces the Spleen Cytokine Response at High Doses within Three Days of Initiating Treatment
3.2. Long-Term Rat Pup Survival and Weight Decreased with Increasing Tacrolimus Concentration
3.3. Tacrolimus Administration Prior to Hypoxic–Ischaemic Brain Injury Is Neuroprotective
3.3.1. Brain Tissue Loss, Neuronal Cell Loss and Apoptosis
3.3.2. Neuroinflammation
3.4. Tacrolimus Administration after Hypoxic–Ischaemic Brain Injury Is Neuroprotective at High Doses
3.4.1. Brain Tissue Loss, Neuronal Cell Loss and Apoptosis
3.4.2. Neuroinflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thornton, C.; Rousset, C.I.; Kichev, A.; Miyakuni, Y.; Vontell, R.; Baburamani, A.A.; Fleiss, B.; Gressens, P.; Hagberg, H. Molecular mechanisms of neonatal brain injury. Neurol. Res. Int. 2012, 2012, 506320. [Google Scholar] [CrossRef]
- Eklind, S.; Mallard, C.; Leverin, A.L.; Gilland, E.; Blomgren, K.; Mattsby-Baltzer, I.; Hagberg, H. Bacterial endotoxin sensitizes the immature brain to hypoxic—Ischaemic injury. Eur. J. Neurosci. 2001, 13, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L.; Huppi, P.S.; Mallard, C. The consequences of fetal growth restriction on brain structure and neurodevelopmental outcome. J. Physiol. 2016, 594, 807–823. [Google Scholar] [CrossRef]
- Nelson, K.B.; Bingham, P.; Edwards, E.M.; Horbar, J.D.; Kenny, M.J.; Inder, T.; Pfister, R.H.; Raju, T.; Soll, R.F. Antecedents of neonatal encephalopathy in the Vermont Oxford Network Encephalopathy Registry. Pediatrics 2012, 130, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.W.; Colford, J.M., Jr. Chorioamnionitis as a risk factor for cerebral palsy: A meta-analysis. JAMA 2000, 284, 1417–1424. [Google Scholar] [CrossRef]
- Hagberg, H.; Mallard, C.; Ferriero, D.M.; Vannucci, S.J.; Levison, S.W.; Vexler, Z.S.; Gressens, P. The role of inflammation in perinatal brain injury. Nat. Rev. Neurol. 2015, 11, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Herz, J.; Bendix, I.; Felderhoff-Müser, U. Peripheral immune cells and perinatal brain injury: A double-edged sword? Pediatr. Res. 2022, 91, 392–403. [Google Scholar] [CrossRef]
- Benjelloun, N.; Renolleau, S.; Represa, A.; Ben-Ari, Y.; Charriaut-Marlangue, C. Inflammatory responses in the cerebral cortex after ischemia in the P7 neonatal Rat. Stroke 1999, 30, 1916–1923; discussion 1923–1924. [Google Scholar] [CrossRef]
- Winerdal, M.; Winerdal, M.E.; Kinn, J.; Urmaliya, V.; Winqvist, O.; Adén, U. Long lasting local and systemic inflammation after cerebral hypoxic ischemia in newborn mice. PLoS ONE 2012, 7, e36422. [Google Scholar] [CrossRef]
- McDonald, C.A.; Penny, T.R.; Paton, M.C.B.; Sutherland, A.E.; Nekkanti, L.; Yawno, T.; Castillo-Melendez, M.; Fahey, M.C.; Jones, N.M.; Jenkin, G.; et al. Effects of umbilical cord blood cells, and subtypes, to reduce neuroinflammation following perinatal hypoxic-ischemic brain injury. J. Neuroinflamm. 2018, 15, 47. [Google Scholar] [CrossRef]
- Nazmi, A.; Albertsson, A.M.; Rocha-Ferreira, E.; Zhang, X.; Vontell, R.; Zelco, A.; Rutherford, M.; Zhu, C.; Nilsson, G.; Mallard, C.; et al. Lymphocytes Contribute to the Pathophysiology of Neonatal Brain Injury. Front. Neurol. 2018, 9, 159. [Google Scholar] [CrossRef] [PubMed]
- Rudd, B.D. Neonatal T Cells: A Reinterpretation. Annu. Rev. Immunol. 2020, 38, 229–247. [Google Scholar] [CrossRef] [PubMed]
- Starzl, T.E.; Todo, S.; Fung, J.; Demetris, A.J.; Venkataramman, R.; Jain, A. FK 506 for liver, kidney, and pancreas transplantation. Lancet 1989, 2, 1000–1004. [Google Scholar] [CrossRef]
- Jain, J.; McCaffrey, P.G.; Miner, Z.; Kerppola, T.K.; Lambert, J.N.; Verdine, G.L.; Curran, T.; Rao, A. The T-cell transcription factor NFATp is a substrate for calcineurin and interacts with Fos and Jun. Nature 1993, 365, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Saffari, T.M.; Bedar, M.; Zuidam, J.M.; Shin, A.Y.; Baan, C.C.; Hesselink, D.A.; Hundepool, C.A. Exploring the neuroregenerative potential of tacrolimus. Expert Rev. Clin. Pharmacol. 2019, 12, 1047–1057. [Google Scholar] [CrossRef]
- Wakita, H.; Tomimoto, H.; Akiguchi, I.; Kimura, J. Dose-dependent, protective effect of FK506 against white matter changes in the rat brain after chronic cerebral ischemia. Brain Res. 1998, 792, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Madsen, J.R.; MacDonald, P.; Irwin, N.; Goldberg, D.E.; Yao, G.L.; Meiri, K.F.; Rimm, I.J.; Stieg, P.E.; Benowitz, L.I. Tacrolimus (FK506) increases neuronal expression of GAP-43 and improves functional recovery after spinal cord injury in rats. Exp. Neurol. 1998, 154, 673–683. [Google Scholar] [CrossRef]
- Setkowicz, Z.; Caryk, M.; Szafraniec, M.; Zmudzińska, A.; Janeczko, K. Tacrolimus (FK506) and cyclosporin A reduce macrophage recruitment to the rat brain injured at perinatal and early postnatal periods. Neurol. Res. 2009, 31, 1060–1067. [Google Scholar] [CrossRef]
- McDonald, C.A.; Payne, N.L.; Sun, G.; Moussa, L.; Siatskas, C.; Lim, R.; Wallace, E.M.; Jenkin, G.; Bernard, C.C. Immunosuppressive potential of human amnion epithelial cells in the treatment of experimental autoimmune encephalomyelitis. J. Neuroinflamm. 2015, 12, 112. [Google Scholar] [CrossRef]
- Wright, A.K.; Miller, C.; Williams, M.; Arbuthnott, G. Microglial activation is not prevented by tacrolimus but dopamine neuron damage is reduced in a rat model of Parkinson’s disease progression. Brain Res. 2008, 1216, 78–86. [Google Scholar] [CrossRef]
- Brecht, S.; Waetzig, V.; Hidding, U.; Hanisch, U.K.; Walther, M.; Herdegen, T.; Neiss, W.F. FK506 protects against various immune responses and secondary degeneration following cerebral ischemia. Anat. Rec. 2009, 292, 1993–2001. [Google Scholar] [CrossRef] [PubMed]
- Brecht, S.; Schwarze, K.; Waetzig, V.; Christner, C.; Heiland, S.; Fischer, G.; Sartor, K.; Herdegen, T. Changes in peptidyl-prolyl cis/trans isomerase activity and FK506 binding protein expression following neuroprotection by FK506 in the ischemic rat brain. Neuroscience 2003, 120, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Furuichi, Y.; Maeda, M.; Moriguchi, A.; Sawamoto, T.; Kawamura, A.; Matsuoka, N.; Mutoh, S.; Yanagihara, T. Tacrolimus, a potential neuroprotective agent, ameliorates ischemic brain damage and neurologic deficits after focal cerebral ischemia in nonhuman primates. J. Cereb. Blood Flow. Metab. 2003, 23, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Furuichi, Y.; Noto, T.; Matsuoka, N.; Mutoh, S.; Yoneda, Y. Tacrolimus (FK506) suppresses rt-PA-induced hemorrhagic transformation in a rat thrombotic ischemia stroke model. Brain Res. 2009, 1254, 99–108. [Google Scholar] [CrossRef]
- Martin-Villalba, A.; Herr, I.; Jeremias, I.; Hahne, M.; Brandt, R.; Vogel, J.; Schenkel, J.; Herdegen, T.; Debatin, K.M. CD95 ligand (Fas-L/APO-1L) and tumor necrosis factor-related apoptosis-inducing ligand mediate ischemia-induced apoptosis in neurons. J. Neurosci. 1999, 19, 3809–3817. [Google Scholar] [CrossRef]
- Sharkey, J.; Butcher, S.P. Immunophilins mediate the neuroprotective effects of FK506 in focal cerebral ischaemia. Nature 1994, 371, 336–339. [Google Scholar] [CrossRef]
- Noto, T.; Ishiye, M.; Furuich, Y.; Keida, Y.; Katsuta, K.; Moriguchi, A.; Matsuoka, N.; Aramori, I.; Goto, T.; Yanagihara, T. Neuroprotective effect of tacrolimus (FK506) on ischemic brain damage following permanent focal cerebral ischemia in the rat. Brain Res. Mol. Brain Res. 2004, 128, 30–38. [Google Scholar] [CrossRef]
- Ankarcrona, M.; Dypbukt, J.M.; Orrenius, S.; Nicotera, P. Calcineurin and mitochondrial function in glutamate-induced neuronal cell death. FEBS Lett. 1996, 394, 321–324. [Google Scholar] [CrossRef]
- Wang, H.G.; Pathan, N.; Ethell, I.M.; Krajewski, S.; Yamaguchi, Y.; Shibasaki, F.; McKeon, F.; Bobo, T.; Franke, T.F.; Reed, J.C. Ca2+-induced apoptosis through calcineurin dephosphorylation of BAD. Science 1999, 284, 339–343. [Google Scholar] [CrossRef]
- Dawson, T.M.; Steiner, J.P.; Dawson, V.L.; Dinerman, J.L.; Uhl, G.R.; Snyder, S.H. Immunosuppressant FK506 enhances phosphorylation of nitric oxide synthase and protects against glutamate neurotoxicity. Proc. Natl. Acad. Sci. USA 1993, 90, 9808–9812. [Google Scholar] [CrossRef]
- Kaminska, B.; Gaweda-Walerych, K.; Zawadzka, M. Molecular mechanisms of neuroprotective action of immunosuppressants—Facts and hypotheses. J. Cell. Mol. Med. 2004, 8, 45–58. [Google Scholar] [CrossRef]
- Dumont, F.J.; Koprak, S.; Staruch, M.J.; Talento, A.; Koo, G.; DaSilva, C.; Sinclair, P.J.; Wong, F.; Woods, J.; Barker, J.; et al. A tacrolimus-related immunosuppressant with reduced toxicity. Transplantation 1998, 65, 18–26. [Google Scholar] [CrossRef]
- Saganová, K.; Gálik, J.; Blaško, J.; Korimová, A.; Račeková, E.; Vanický, I. Immunosuppressant FK506: Focusing on neuroprotective effects following brain and spinal cord injury. Life Sci. 2012, 91, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Bechstein, W.O. Neurotoxicity of calcineurin inhibitors: Impact and clinical management. Transpl. Int. 2000, 13, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Anghel, D.; Tanasescu, R.; Campeanu, A.; Lupescu, I.; Podda, G.; Bajenaru, O. Neurotoxicity of immunosuppressive therapies in organ transplantation. Maedica 2013, 8, 170–175. [Google Scholar] [PubMed]
- Dumont, F.J.; Staruch, M.J.; Koprak, S.L.; Siekierka, J.J.; Lin, C.S.; Harrison, R.; Sewell, T.; Kindt, V.M.; Beattie, T.R.; Wyvratt, M. The immunosuppressive and toxic effects of FK-506 are mechanistically related: Pharmacology of a novel antagonist of FK-506 and rapamycin. J. Exp. Med. 1992, 176, 751–760. [Google Scholar] [CrossRef]
- Aly, H.; Khashaba, M.T.; El-Ayouty, M.; El-Sayed, O.; Hasanein, B.M. IL-1beta, IL-6 and TNF-alpha and outcomes of neonatal hypoxic ischemic encephalopathy. Brain Dev. 2006, 28, 178–182. [Google Scholar] [CrossRef]
- Cai, Z.; Lin, S.; Pang, Y.; Rhodes, P.G. Brain injury induced by intracerebral injection of interleukin-1beta and tumor necrosis factor-alpha in the neonatal rat. Pediatr. Res. 2004, 56, 377–384. [Google Scholar] [CrossRef]
- Liu, J.; Feng, Z.C. Increased umbilical cord plasma interleukin-1 beta levels was correlated with adverse outcomes of neonatal hypoxic-ischemic encephalopathy. J. Trop. Pediatr. 2010, 56, 178–182. [Google Scholar] [CrossRef]
- Liu, X.; Quan, N. Microglia and CNS Interleukin-1: Beyond Immunological Concepts. Front. Neurol. 2018, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Fleiss, B.; Gressens, P. Tertiary mechanisms of brain damage: A new hope for treatment of cerebral palsy? Lancet Neurol. 2012, 11, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, H.; Gressens, P.; Mallard, C. Inflammation during fetal and neonatal life: Implications for neurologic and neuropsychiatric disease in children and adults. Ann. Neurol. 2012, 71, 444–457. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, M.J.; Penny, T.; Pham, Y.; Sutherland, A.E.; Jenkin, G.; Fahey, M.C.; Paton, M.C.B.; Finch-Edmondson, M.; Miller, S.L.; McDonald, C.A. Neuroprotective Action of Tacrolimus before and after Onset of Neonatal Hypoxic–Ischaemic Brain Injury in Rats. Cells 2023, 12, 2659. https://doi.org/10.3390/cells12222659
Smith MJ, Penny T, Pham Y, Sutherland AE, Jenkin G, Fahey MC, Paton MCB, Finch-Edmondson M, Miller SL, McDonald CA. Neuroprotective Action of Tacrolimus before and after Onset of Neonatal Hypoxic–Ischaemic Brain Injury in Rats. Cells. 2023; 12(22):2659. https://doi.org/10.3390/cells12222659
Chicago/Turabian StyleSmith, Madeleine J., Tayla Penny, Yen Pham, Amy E. Sutherland, Graham Jenkin, Michael C. Fahey, Madison C. B. Paton, Megan Finch-Edmondson, Suzanne L. Miller, and Courtney A. McDonald. 2023. "Neuroprotective Action of Tacrolimus before and after Onset of Neonatal Hypoxic–Ischaemic Brain Injury in Rats" Cells 12, no. 22: 2659. https://doi.org/10.3390/cells12222659