

How to Differentiate between Resistant and Susceptible Wheat Cultivars for Leaf Rust Fungi Using Antioxidant Enzymes and Histological and Molecular Studies?

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Evaluation of Eight Wheat Cultivars at Seedling and Adult Stages of Leaf Rust
2.2. he Laboratory Studies
2.2.1. Histochemical Analysis of Reactive Oxygen Species (ROS)
2.2.2. Activities of Antioxidant Enzymes
2.2.3. Electrolyte Leakage
2.2.4. Histological Studies
2.2.5. Detection of Lr50 in Eight Wheat Cultivars
2.2.6. Statistical Analysis
3. Results
3.1. Evaluation of Eight Wheat Cultivars at Seedling and Adult Stages after Infection with Leaf Rust
3.2. Correlation Analysis
3.3. Activity of Antioxidant Enzymes
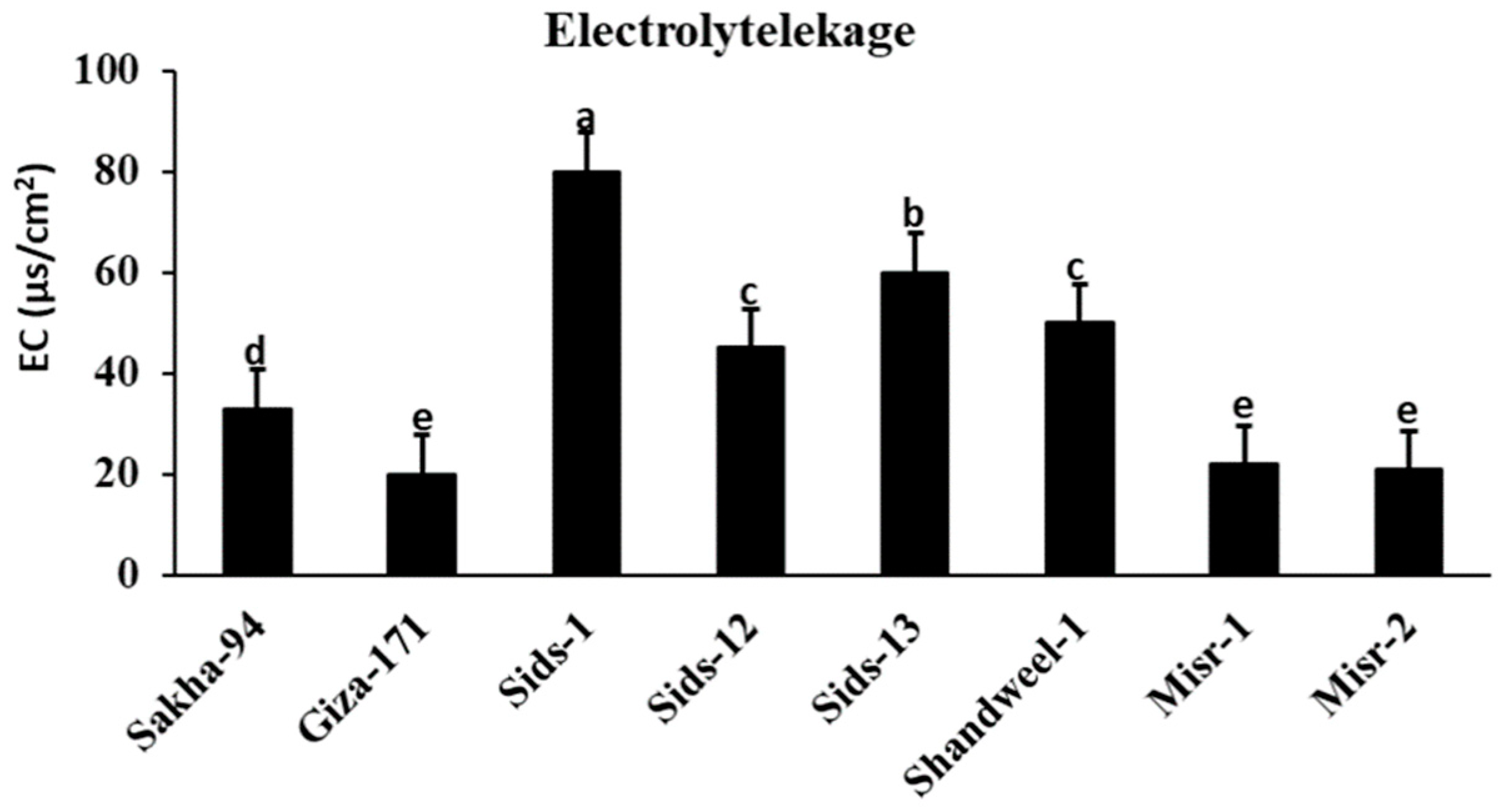
3.4. Electrolyte Leakage
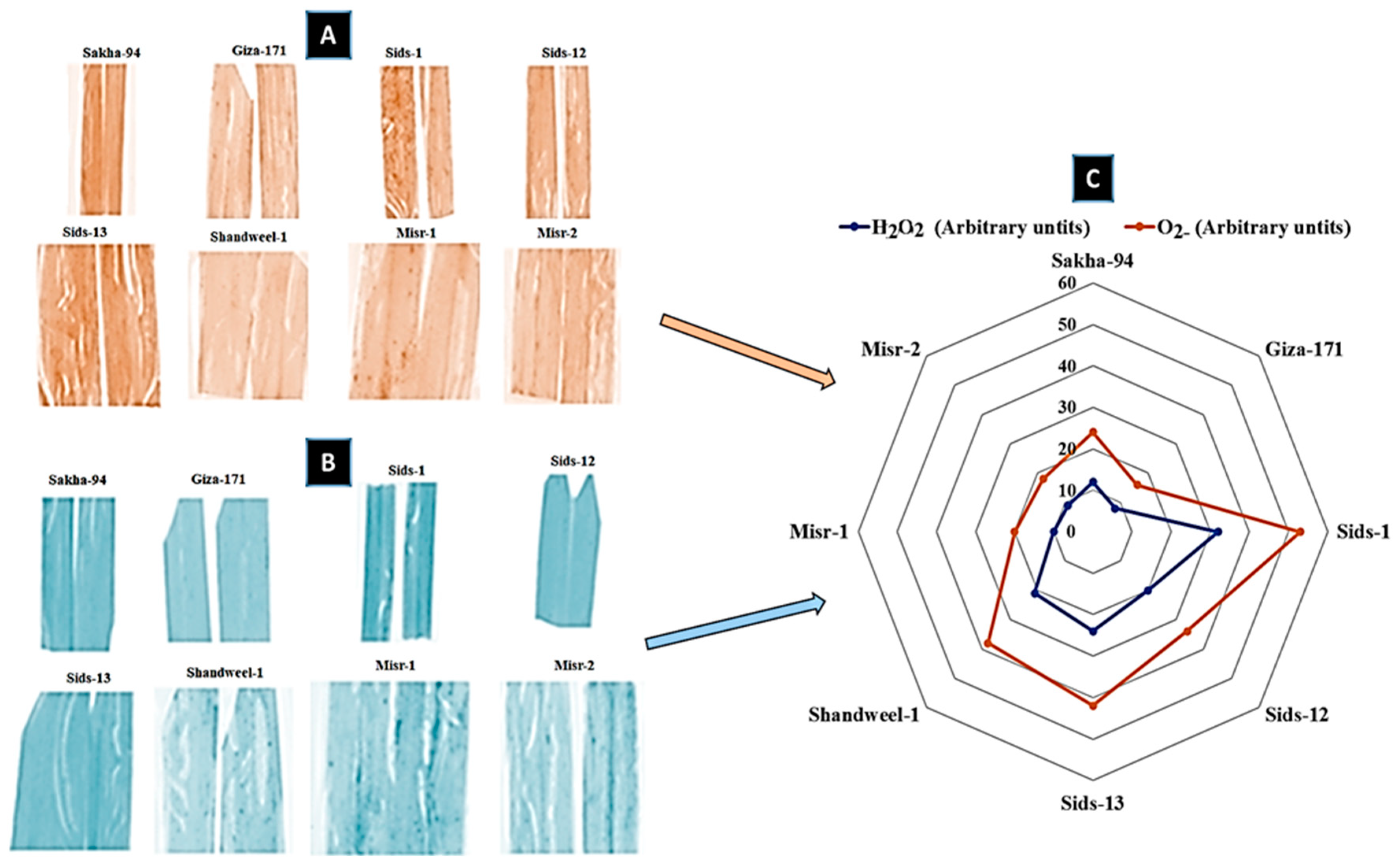
3.5. Histochemical Analysis of Reactive Oxygen Species (ROS)
3.6. Detection of Lr50 in Eight Wheat Cultivars
3.7. Histological Studies of Eight Wheat Cultivars Infected with P. triticina
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sofy, M.R.; Elhawat, N.; Alshaal, T. Glycine betaine counters salinity stress by maintaining a high K+/Na+ ratio and antioxidant defense via limiting Na+ uptake in common bean (Phaseolus vulgaris L.). Ecotoxicol. Environ. Saf. 2020, 200, 110732. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, B.; Li, L.; Zeng, F.; Li, X. Negative effects of long-term exposure to salinity, drought, and combined stresses on halophyte Halogeton glomeratus. Physiologia Plantarum. 2021, 173, 2307–2322. [Google Scholar] [CrossRef] [PubMed]
- Syeed, S.; Sehar, Z.; Masood, A.; Anjum, N.A.; Khan, N.A. Control of elevated ion accumulation, oxidative stress, and lipid peroxidation with salicylic acid-induced accumulation of glycine betaine in salinity-exposed Vigna radiata L. Appl. Biochem. Biotechnol. 2021, 193, 3301–3320. [Google Scholar] [PubMed]
- Abdelaal, K.; Mazrou, Y.; Mohamed, A.A.; Ghazy, M.; Barakat, M.; Hafez, Y.; Gaballah, M. The different responses of rice genotypes to heat stress associated with morphological, chlorophyll and yield characteristics. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12550. [Google Scholar] [CrossRef]
- Ibrahim, A.E.; Abd El Mageed, T.; Abohamid, Y.; Abdallah, H.; El-Saadony, M.; AbuQamar, S.; El-Tarabily, K.; Abdou, N. Exogenously applied proline enhances morph-physiological responses and yield of drought-stressed maize plants grown under different irrigation systems. Front. Plant Sci. 2022, 14, 897027. [Google Scholar] [CrossRef]
- Khedr, R.; Aboukhadrah, S.; El-Hag, D.; El-Mohamady, E.; Abdelaal, K. Ameliorative effects of nano silica and some growth stimulants on water relations, biochemical and productivity of wheat under saline soil conditions. Fresenius Environ. Bull. 2003, 32, 375–384. [Google Scholar]
- El-Orabey, W.M.; Mabrouk, O.I.; Abd-Allah, M.G.; Esmail, M.S. Inheritance and detection of leaf rust resistance genes in some Egyptian wheat cultivars. Int. J. Genet. Genom. 2020, 8, 1–10. [Google Scholar] [CrossRef]
- Saudy, H.; Hamed, M.F.; Abd El-Momen, W.R.; Hussein, H. Nitrogen use Rationalization and Boosting Wheat Productivity by Applying Packages of Humic, Amino Acids, and Microorganisms. Commun. Soil Sci. Plant Anal. 2020, 51, 1036–1047. [Google Scholar] [CrossRef]
- Saudy, H.S.; Mubarak, M. Mitigating the Detrimental Impacts of Nitrogen Deficit and Fenoxaprop-p-ethyl Herbicide on Wheat using Silicon. Commun. Soil Sci. Plant Anal. 2015, 46, 897–907. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Mert, Z.; Akan, K.; Demir, L.; Unsal, R.; Sermet, C.; Keser, M.; Akin, B.; Morgounov, A. Virulence of Puccinia triticina in Turkey and leaf rust resistance in Turkish wheat cultivars. Eur. J. Plant Pathol. 2013, 135, 703–716. [Google Scholar] [CrossRef]
- Hasan, M.A.; Abu Aly, A.A.; El-Shehawy, A.E.A. Losses in grain yield due to leaf rust, caused by Puccinia triticina Eriks. J. Plant Protec. Pathol. Mansoura Univ. 2012, 3, 959–966. [Google Scholar] [CrossRef]
- Draz, I.S.; Abou-Elseoud, M.S.; Kamara, A.M.; Alaa-Eldein Omaima, A.; EL-Bebany, A.F. Screening of wheat genotypes for leaf rust resistance along with grain yield. Annals Agric. Sci. 2015, 60, 29–39. [Google Scholar] [CrossRef]
- Nazim, M.; El-Shehidi, A.A.; Abdou, Y.A.; El-Daoudi, Y.H. Yield loss caused by leaf rust on four wheat cultivars under epiphytotic levels. In Proceedings of the 4th Conference of Microbiology, Cairo, Egypt, 24–28 December 1980; pp. 17–27. [Google Scholar]
- Hafez, Y.M.; Abdelaal, K. Investigation of susceptibility and resistance mechanisms of some Egyptian wheat cultivars (Triticum aestivum L.) inoculated with Blumeria graminis f.sp. tritici using certain biochemical, molecular characterization and SEM. J. Plant Prot. Pathol. Mansoura Univ. 2015, 6, 431–454. [Google Scholar] [CrossRef]
- Omara, R.I.; Abdelaal, K. Biochemical, histopathological and genetic analysis associated with leaf rust infection in wheat plants (Triticum aestivum L.). Physiol. Mol. Plant Pathol. 2018, 104, 48–57. [Google Scholar] [CrossRef]
- Omara, R.I.; El-Kot, G.A.; Fadel, F.M.; Abdelaal, K.; Saleh, E.M. Efficacy of certain bioagents on patho-physiological characters of wheat plants under wheat leaf rust stress. Physiol. Mol. Plant Pathol. 2019, 106, 102–108. [Google Scholar] [CrossRef]
- Al-Shammari, W.B.; Altamimi, H.R.; Abdelaal, K. Improvement in Physiobiochemical and Yield Characteristics of Pea Plants with Nano Silica and Melatonin under Salinity Stress Conditions. Horticulturae 2023, 9, 711. [Google Scholar] [CrossRef]
- Esmail, S.M.; Omara, R.I.; Abdelaal, K.; Hafez, Y.M. Histological and biochemical aspects of compatible and incompatible wheat-Puccinia striiformis interactions. Physiol. Mol. Plant Pathol. 2019, 106, 120–128. [Google Scholar] [CrossRef]
- Stakman, E.C.; Stewari, D.M.; Loegering, W.Q. Identification of Physiological Races of Puccinia graminis var. tritici; US Department of Agricultural Publications E617; USDA: Washington, DC, USA, 1962; 53p.
- Huckelhoven, R.; Fodor, J.; Preis, C.; Kogel, K.H. Hypersensitive cell death and papilla formation in barley attacked by the powdery mildew fungus are associated with hydrogen peroxide but not with salicylic acid accumulation. Plant Physiol. 1999, 119, 1251–1260. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Malik, C.P.; Singh, M.B. Plant Emynology and Histoenzymology; Indian and printed in Navin; Kalyani Publishers: Shanndara, Delhi, India, 1980; pp. 54–56. [Google Scholar]
- Hammerschmidt, R.; Nuckles, E.M.; Kuc, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Szalai, G.; Janda, T.; Szigeti, E.Z. Role of light in post chilling symptoms in maize. J. Plant Physiol. 1996, 148, 378–383. [Google Scholar] [CrossRef]
- Nassar, M.A.; El-Sahhar, K.F. Botanical Preparations and Microscopy (Microtechnique); Academic Bookshop: Giza, Egypt, 1998; p. 219. (In Arabic) [Google Scholar]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley and Sons, Inc.: New York, NY, USA, 1984; p. 680. [Google Scholar]
- Duncan, D.B. Multiple range and multiple f test. Biometrics 1955, 11, 1–24. [Google Scholar] [CrossRef]
- Gomma, A.S.A. Report of the Egyptian Wheat Varieties and Their Development; Egyptian Society of Genetics Science: Cairo, Egypt, 1978. [Google Scholar]
- Abdelbacki, A.M.M.; Omara, R.I.; Najeeb, M.A.; Soliman, N.E. Identification of leaf rust resistant genes Lr9, Lr25, Lr28, Lr29 and Lr67 in ten Egyptian wheat cultivars using molecular markers. Int. J. Biotechnol. Res. 2014, 2, 89–96. [Google Scholar]
- Mabrouk, O.I.A. Durable Resistance to Leaf Rust in Some Egyptian Wheat Cultivars. Ph.D. Thesis, Faculty of Agriculture Cairo University, Giza, Egypt, 2016. [Google Scholar]
- Kolmer, J.A.; Singh, R.P.; Garvin, D.F.; Viccars, L.; William, H.M.; Huerta-Espino, J.H.; Obonnaya, F.C.; Raman, H.; Orford, S.; Bariana, H.S.; et al. Analysis of the Lr34/Yr18 rust resistance region in wheat germplasm. Crop Sci. 2008, 48, 1841–1852. [Google Scholar] [CrossRef]
- Singh, R.P.; Huerta-Espino, J.; William, H.M. Genetics and breeding for durable resistance to leaf and stripe rusts in wheat. Turk. J. Agric. 2005, 29, 121–127. [Google Scholar]
- Boulot, O.A. Durable resistance for leaf rust in twelve Egyptian wheat varieties. Egypt. J. Appl. Sci. 2007, 22, 40–60. [Google Scholar]
- Gechev, T.; Gadjev, I.; Breusegem, F.; Inzé, D.; Dukiandjiev, S.; Toneva, V. and Minkov, I. Hydrogen peroxide protects tobacco from oxidative stress by inducing a set of antioxidant enzymes. Cell. Mol. Life Sci. 2002, 59, 708–714. [Google Scholar] [CrossRef]
- Naz, R.; Batool, S.; Shahid, M.; Keyani, R.; Yasmin, H.; Nosheen, A.; Hassan, M.N.; Mumtaz, S.; Siddiqui, M.H. Exogenous silicon and hydrogen sulfide alleviates the simultaneously occurring drought stress and leaf rust infection in wheat. Plant Physiol. Biochem. 2021, 166, 558–571. [Google Scholar] [CrossRef]
- Zakaria, W.G.E.; Atia, M.M.; Ali, A.Z.; Abbas, E.E.A.; Salim, B.M.A.; Marey, S.A.; Hatamleh, A.A.; Elnahal, A.S.M. Assessing the Effectiveness of Eco-Friendly Management Approaches for Controlling Wheat Yellow Rust and Their Impact on Antioxidant Enzymes. Plants 2023, 12, 2954. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Tiwari, K.; Prakasha, T.; Prasad, S.S. Use of Host Resistance for Management Wheat Rusts. In Innovative Approaches in Diagnosis and Management of Crop Diseases; Apple Academic Press: New York, NY, USA, 2021; pp. 269–301. [Google Scholar]
- Dannies, M.; Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A.; Terefe, T.; Mhlongo, M.I. Metabolomic evaluation of PGPR defence priming in wheat (Triticum aestivum L.) cultivars infected with Puccinia striiformis f. sp. tritici (stripe rust). Front. Plant Sci. 2023, 14, 1261. [Google Scholar]
- Atia, M.A.; El-Khateeb, E.A.; Abd El-Maksoud, R.M.; Abou-Zeid, M.A.; Salah, A.; Abdel-Hamid, A.M.E. Mining of leaf rust resistance genes content in Egyptian bread wheat collection. Plants 2021, 10, 1378. [Google Scholar] [CrossRef]
- Esmail, S.M.; Omar, G.E.; Mourad, A.M. In-Depth Understanding of the Genetic Control of Stripe Rust Resistance (Puccinia striiformis f. Sp. tritici) Induced in Wheat (Triticum aestivum) by Trichoderma Asperellum T34. Plant Dis. 2023, 107, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, C.; Cheng, Y.; Chen, X.; Han, Q.; Huang, L.; Wei, G.; Kang, Z. Histological and cytological characterization of adult plant resistance to wheat stripe rust. Plant Cell Rep. 2012, 31, 2121–2137. [Google Scholar] [CrossRef]
- Pereyra, M.A.; Colabelli, M.N.; Barassi, C.A.; Cerus, C.M. A better water status in wheat seedling induced by azospirillum under osmotic stress is related to morphological changes in xylem vessels of the coleoptile. Appl. Soil Ecol. 2012, 53, 94–97. [Google Scholar] [CrossRef]
- Rehan, M.; Ahmad, A.T. Alleviating the adverse effects of plant pathogens, drought and salinity stress factors using plant growth promoting bacteria. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12911. [Google Scholar]
- Mandal, K.; Saravanan, R.; Maiti, S.; Kothari, I.L. Effect of downy mildew disease on photosynthesis and chlorophyll fluorescence in Plantago ovata Forsk. J. Plant Dis. Prot. 2009, 116, 164–168. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Pedigree |
|---|---|
| Sakha94 | OPATA/RAYON//KAUZ CMBW9043180-OTOPM-3Y-010M… |
| Giza171 | Sakha93/Gemmeiza9 S 6-1GZ-2GZ-2GZ-0S |
| Sids1 | HD2172/Pavon“S”//1158.57/Maya74“S”. SD46-4Sd-2SD-1SD-0SD |
| Sids12 | Buc//7c/ald/5/maya74/on//1160-147/3/bb/gll/4/chat“s′” |
| Sids13 | AMAZ19=KAUZ“S”//TSI/SNB“S” |
| Shandweel1 | SITE/MO/4/NAC/TH.AC//3*PVN/3/MIRLO/BUC |
| Misr1 | OASIS/SKAUZ//4*BCN/3/2*PASTOR |
| Misr2 | SKAUZ/BAV92 |
| Wheat Cultivars | Thickness of Blade (µ) | Thickness of Upper Epidermis (µ) | Thickness of Lower Epidermis (µ) | Thickness of Mesophyll Tissue (µ) | Bundle Length (µ) | Bundle Width (µ) |
|---|---|---|---|---|---|---|
| Sakha-94 | 217.35 | 11.81 | 12.71 | 124.23 | 86.72 | 85.68 |
| Giza-171 | 220.23 | 13.23 | 13.98 | 128.76 | 88.76 | 90.32 |
| Sids-1 | 202.15 | 8.54 | 8.98 | 104.21 | 73.74 | 70.23 |
| Sids-12 | 210.23 | 10.98 | 11.04 | 112.34 | 82.47 | 80.46 |
| Sids-13 | 208.13 | 9.87 | 9.87 | 110.34 | 79.23 | 76.34 |
| Shandweel-1 | 211.68 | 10.23 | 10.65 | 118.34 | 81.43 | 79.68 |
| Misr-1 | 218.23 | 13.79 | 13.22 | 129.75 | 89.2 | 87.22 |
| Misr-2 | 219.21 | 13.48 | 13.79 | 128.65 | 86.67 | 89.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omara, R.I.; Alkhateeb, O.A.; Abdou, A.H.; El-Kot, G.A.; Shahin, A.A.; Saad-El-Din, H.I.; Abdelghany, R.; AL-Shammari, W.B.; Albadrani, M.; Hafez, Y.; et al. How to Differentiate between Resistant and Susceptible Wheat Cultivars for Leaf Rust Fungi Using Antioxidant Enzymes and Histological and Molecular Studies? Cells 2023, 12, 2643. https://doi.org/10.3390/cells12222643
Omara RI, Alkhateeb OA, Abdou AH, El-Kot GA, Shahin AA, Saad-El-Din HI, Abdelghany R, AL-Shammari WB, Albadrani M, Hafez Y, et al. How to Differentiate between Resistant and Susceptible Wheat Cultivars for Leaf Rust Fungi Using Antioxidant Enzymes and Histological and Molecular Studies? Cells. 2023; 12(22):2643. https://doi.org/10.3390/cells12222643
Chicago/Turabian StyleOmara, Reda I., Omar Abdullah Alkhateeb, Ahmed Hassan Abdou, Gabr A. El-Kot, Atef A. Shahin, Heba I. Saad-El-Din, Rady Abdelghany, Wasimah B. AL-Shammari, Muayad Albadrani, Yaser Hafez, and et al. 2023. "How to Differentiate between Resistant and Susceptible Wheat Cultivars for Leaf Rust Fungi Using Antioxidant Enzymes and Histological and Molecular Studies?" Cells 12, no. 22: 2643. https://doi.org/10.3390/cells12222643