

Cissus quadrangularis (Hadjod) Inhibits RANKL-Induced Osteoclastogenesis and Augments Bone Health in an Estrogen-Deficient Preclinical Model of Osteoporosis Via Modulating the Host Osteoimmune System
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
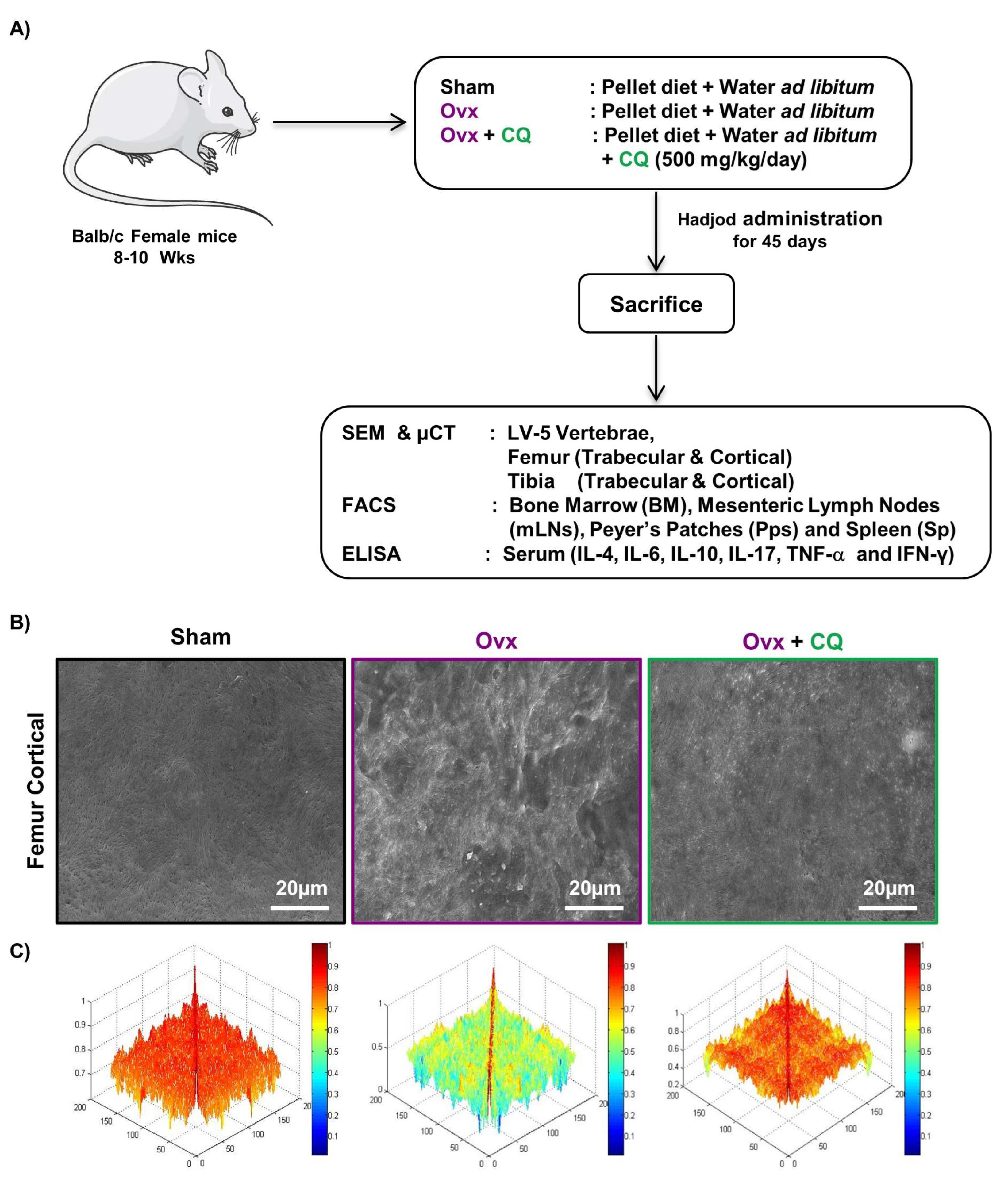
2.1. Animals and Treatments
2.2. Collection of Plant Material
2.3. Preparation of CQ Extract
2.4. Reagents and Antibodies
2.5. Osteoclasts Differentiation and TRAP Staining
2.6. F-Actin Ring Polymerization Assay
2.7. Cell Viability or Metabolic Activity Assay
2.8. Scanning Electron Microscopy (SEM)
2.9. Micro-Computed Tomography (µ-CT)
2.10. Flow Cytometry
2.11. Enzyme-Linked Immunosorbent Assay (ELISA)
2.12. Histologic Analysis
2.13. Statistical Analysis of Data
3. Results
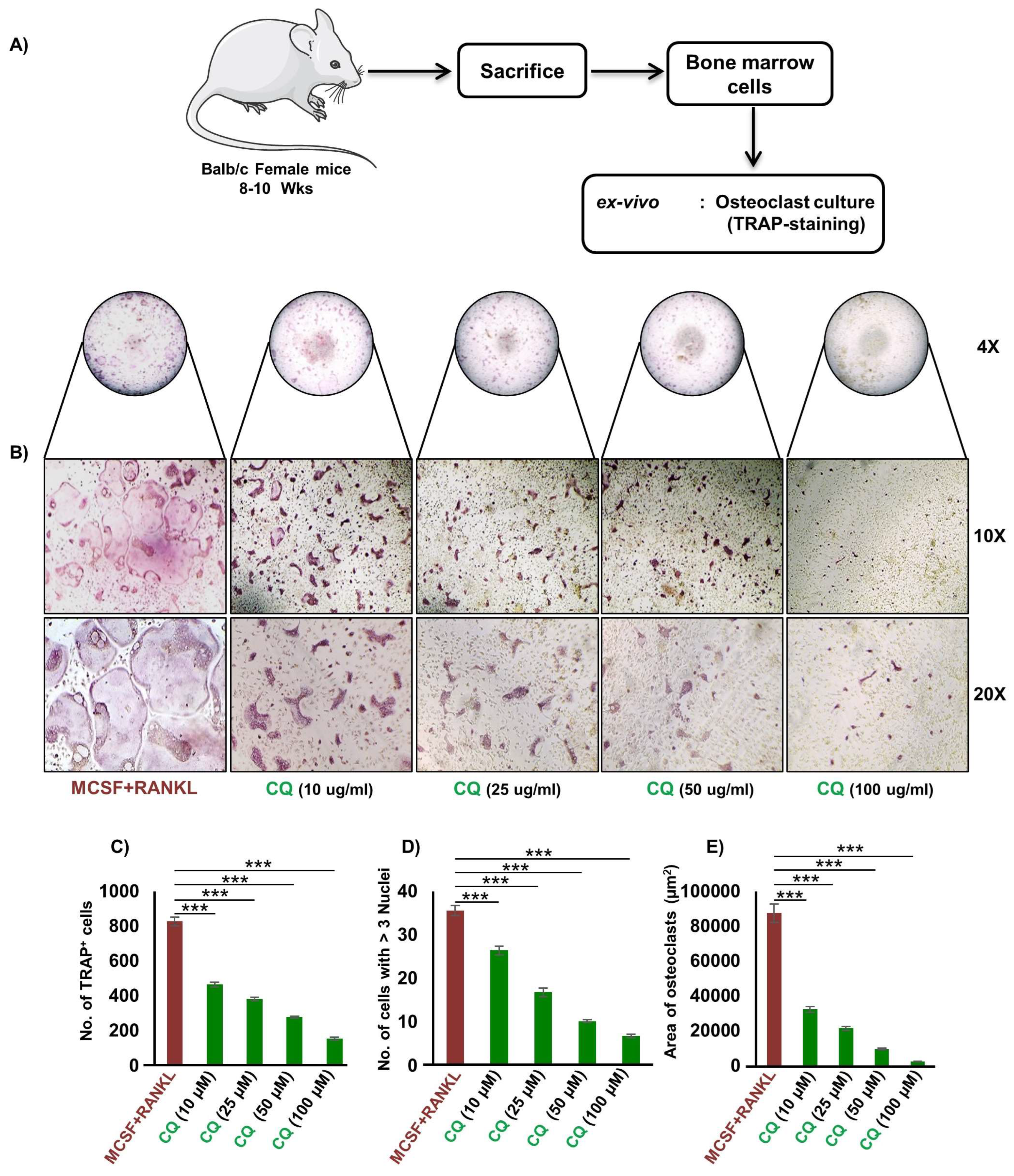
3.1. CQ Inhibits RANKL-Induced Osteoclastogenesis
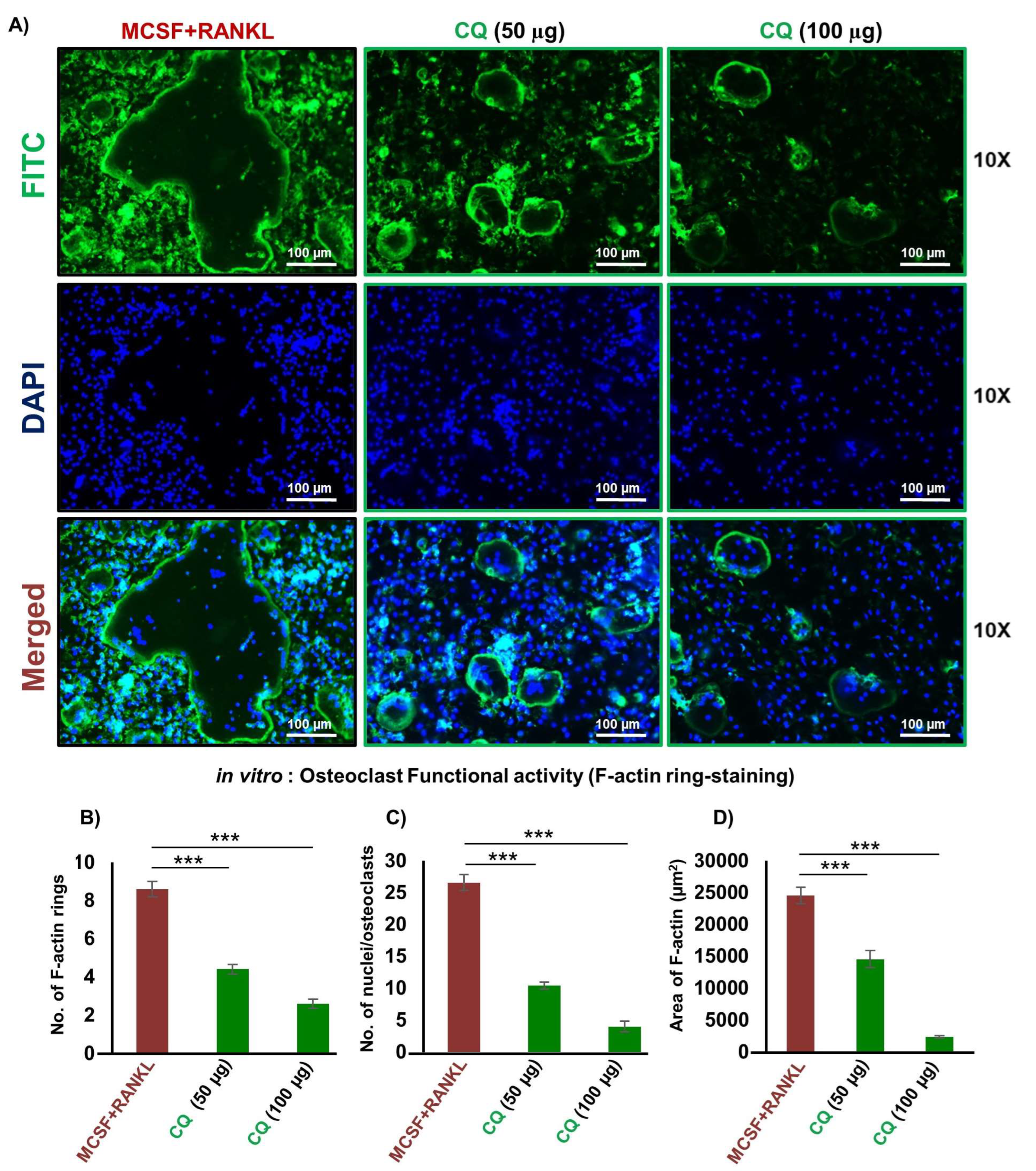
3.2. CQ Attenuates the Functional Activity of Osteoclasts
3.3. CQ Augments Bone Health under Postmenopausal Osteoporotic Conditions
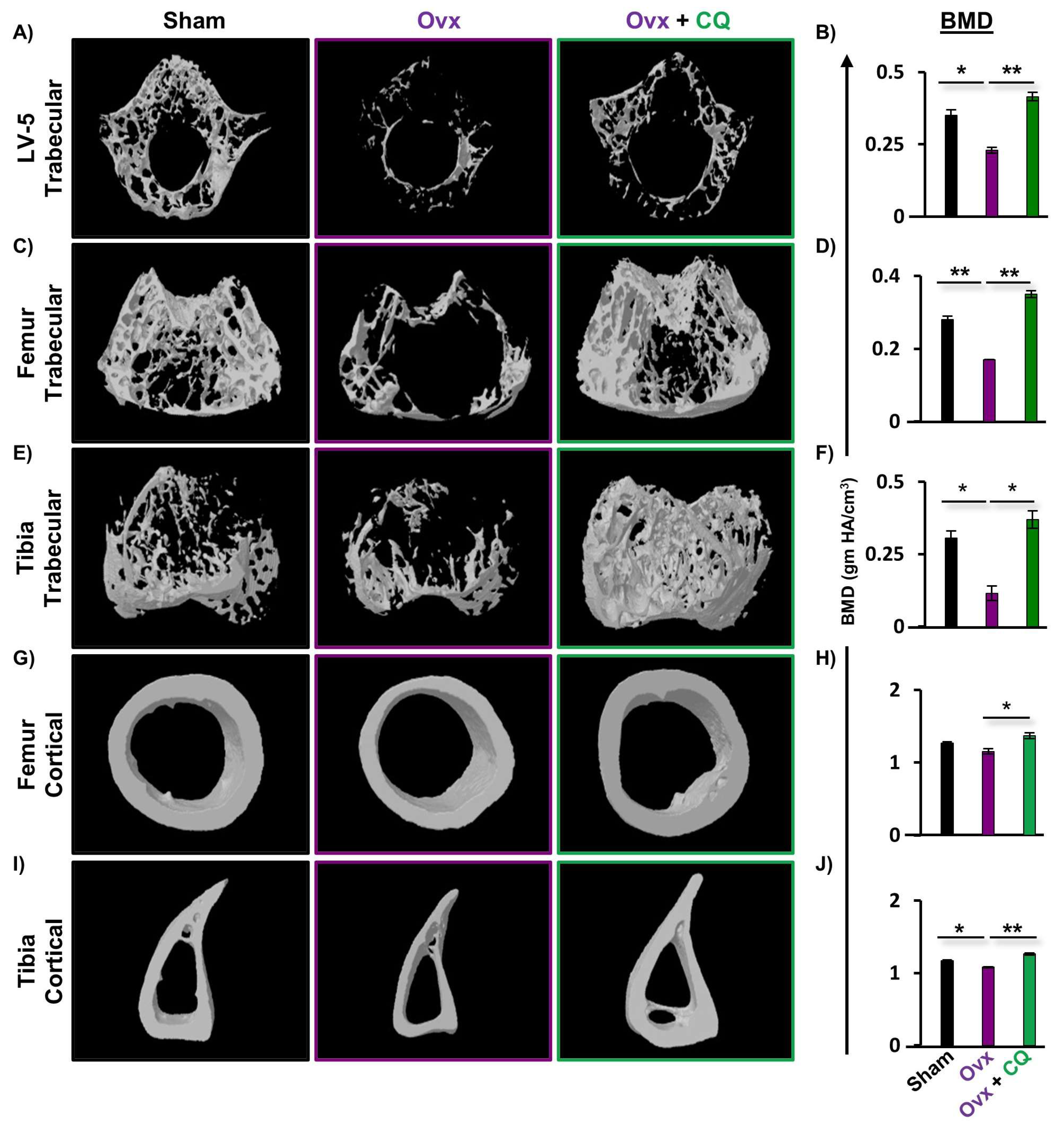
3.4. CQ Enhances Bone Micro-Architecture and Histomorphometric Indices
3.5. CQ Improves Bone Mineral Density (BMD)
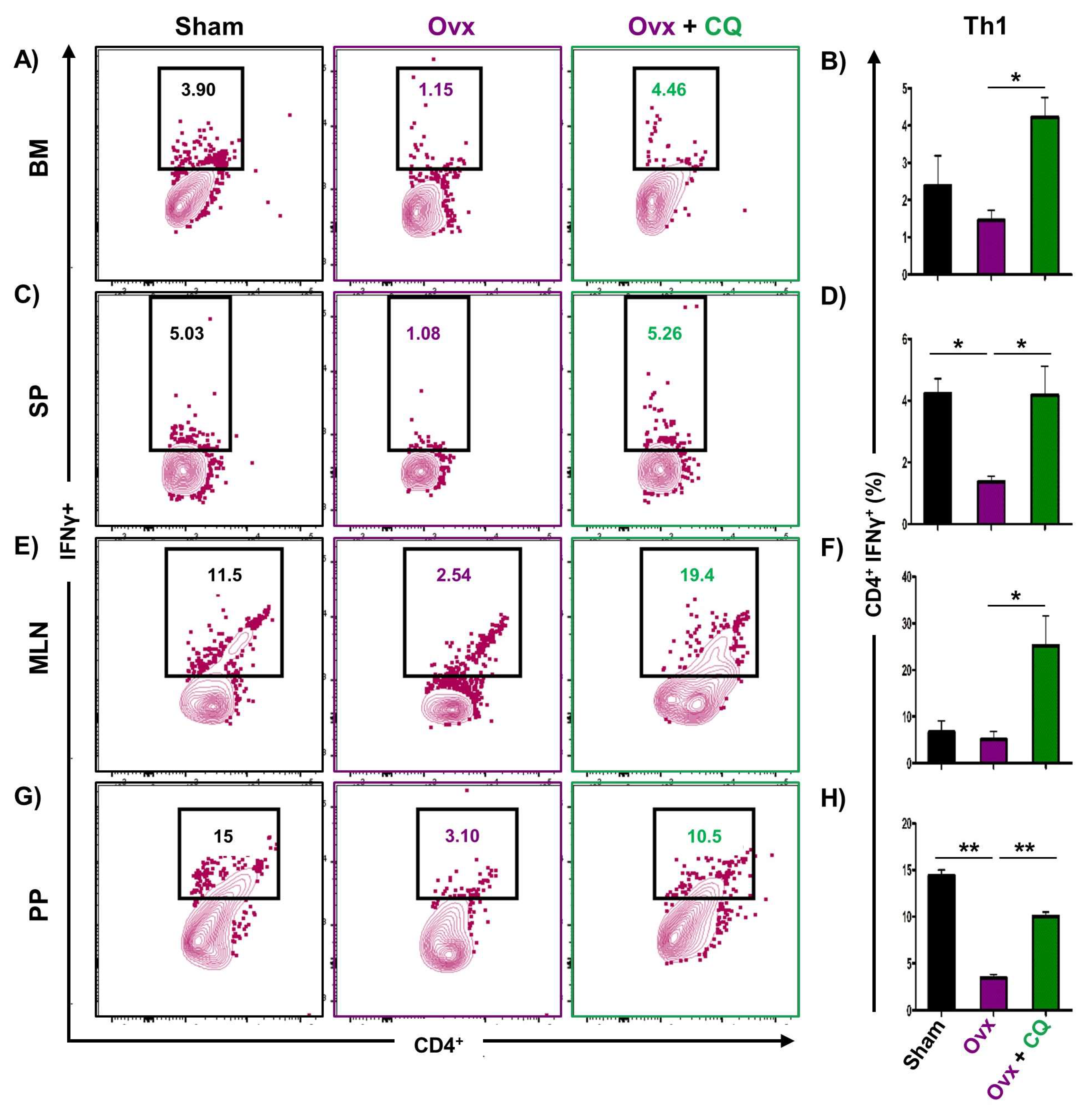
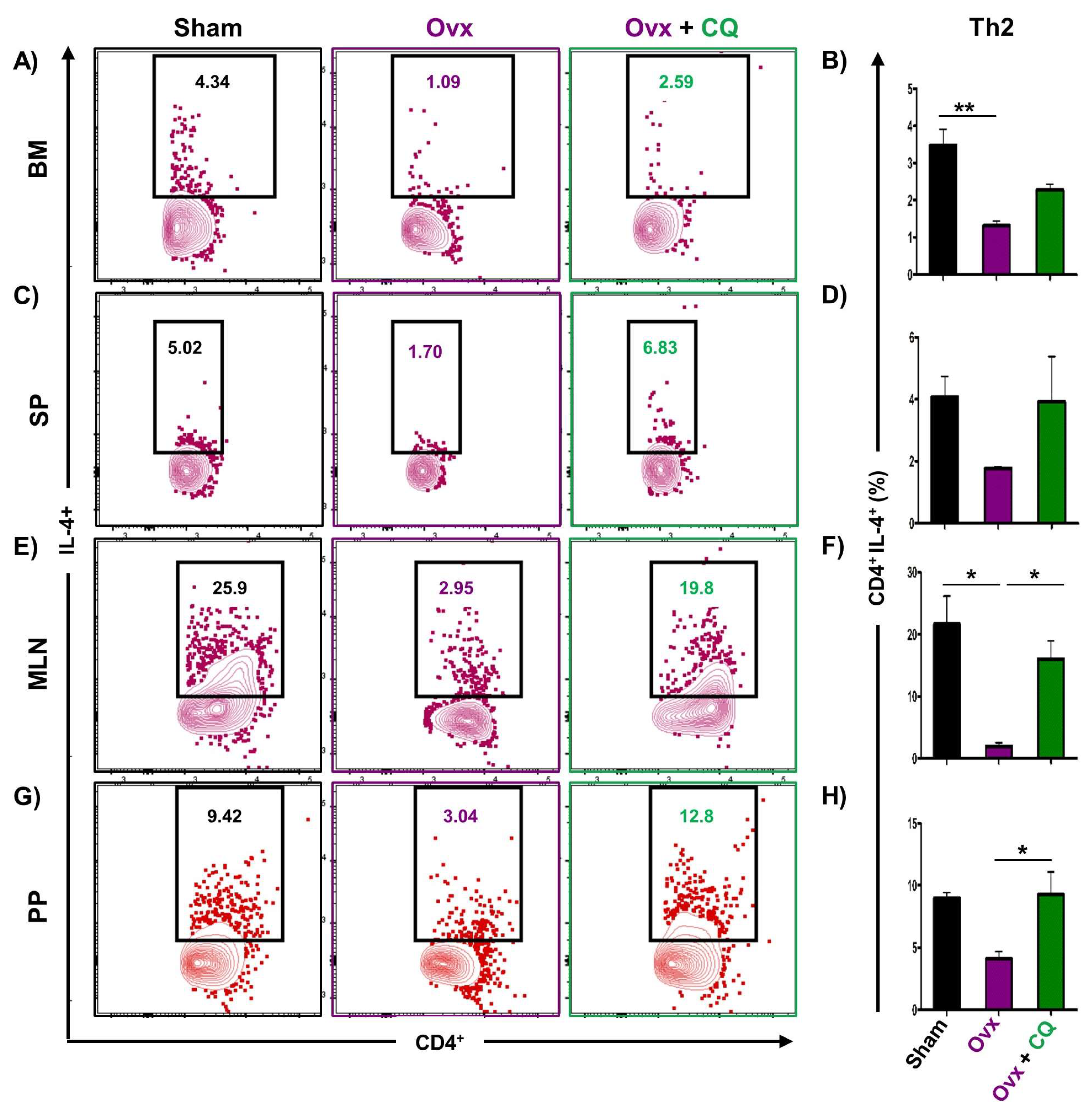
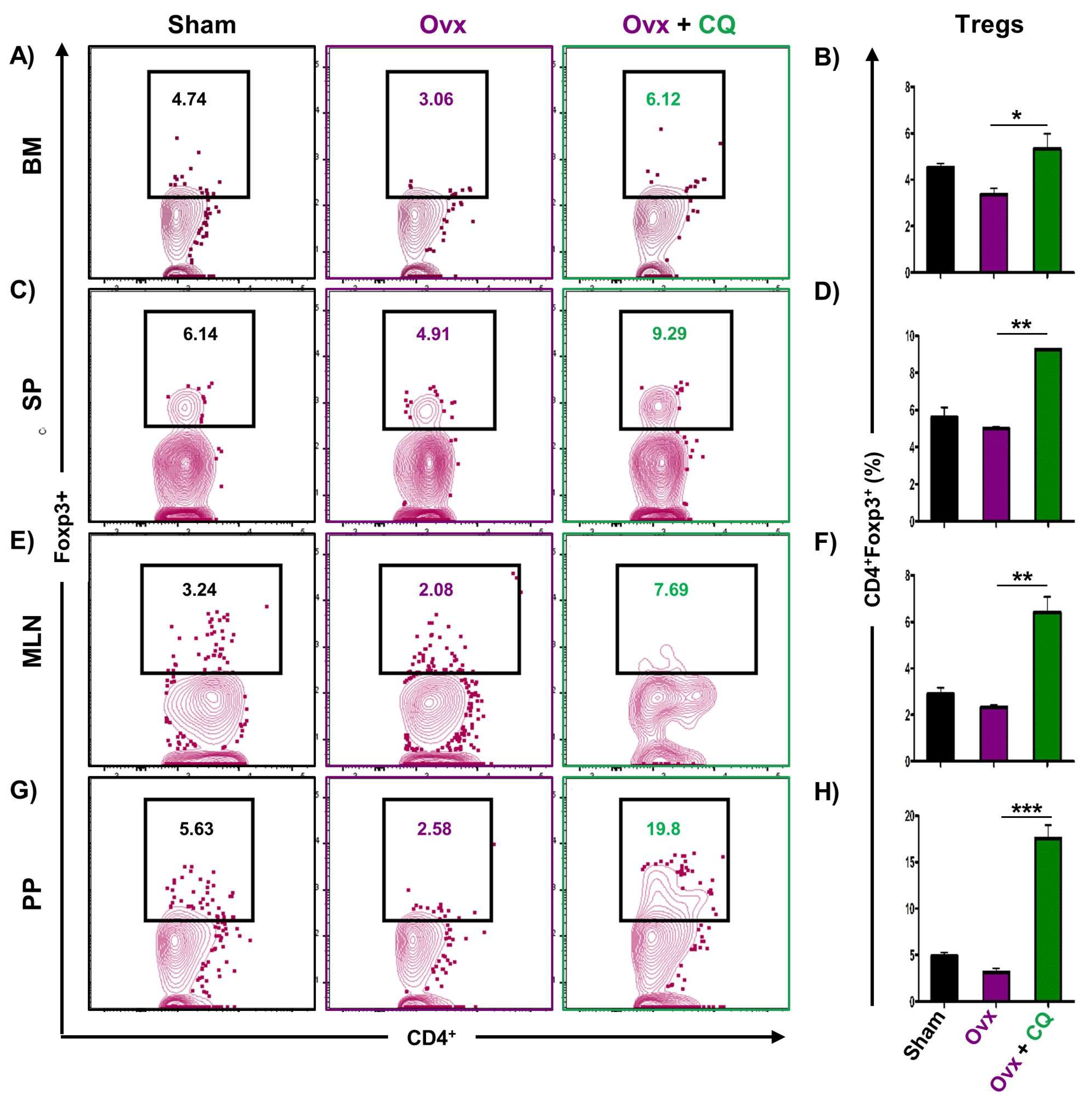
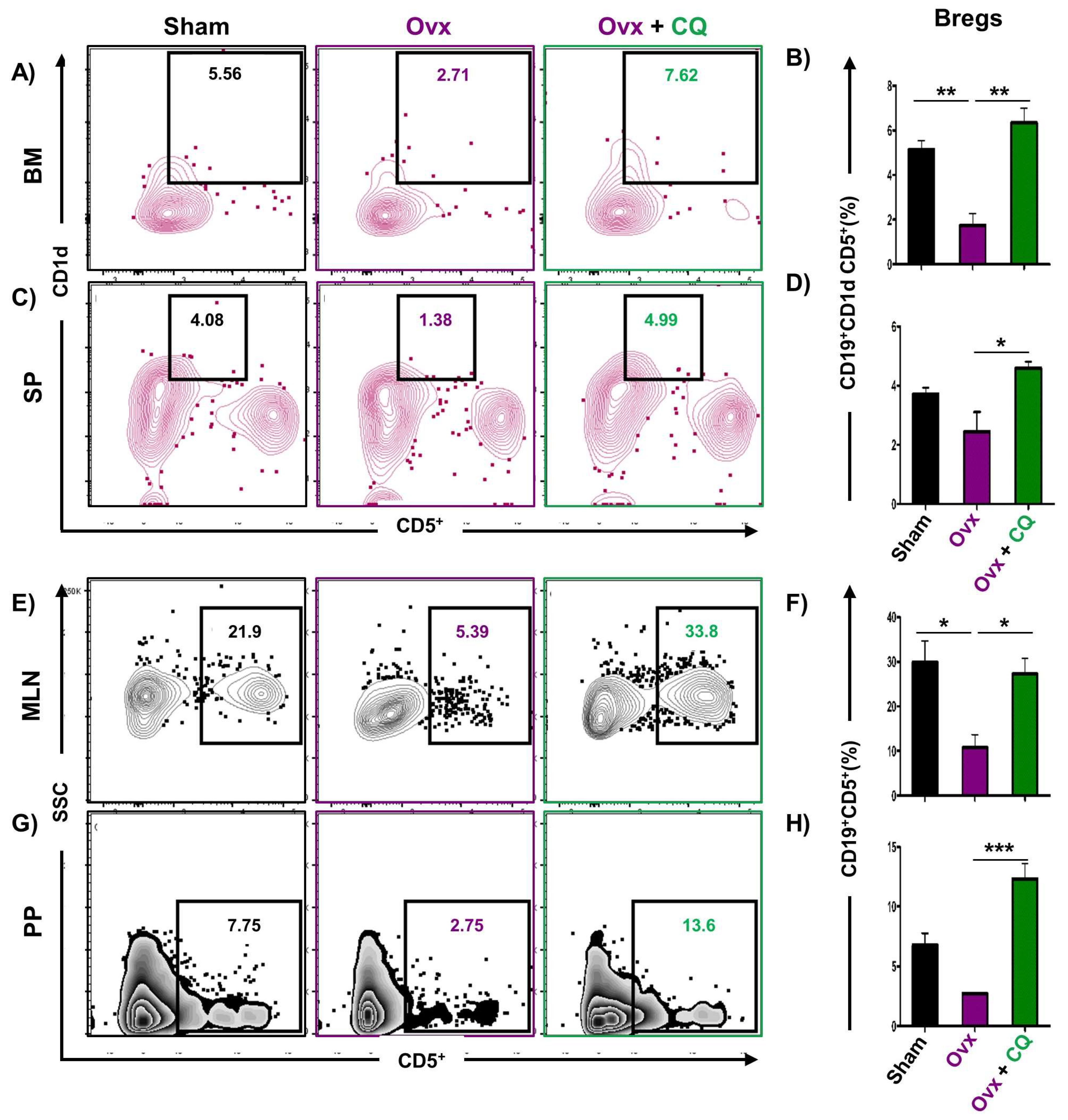
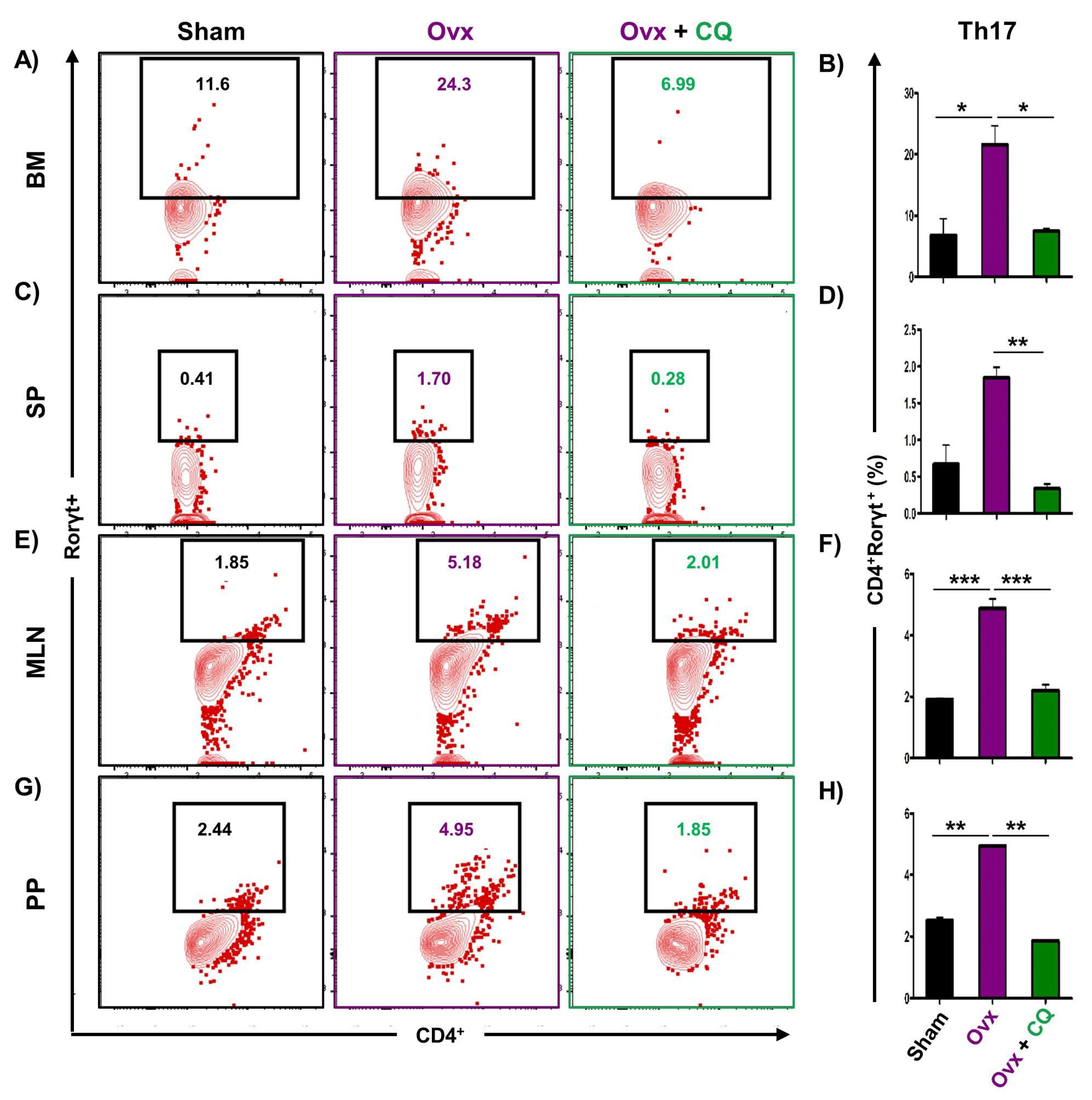
3.6. CQ Augments Bone Health Via Modulating Immunoporotic Cells
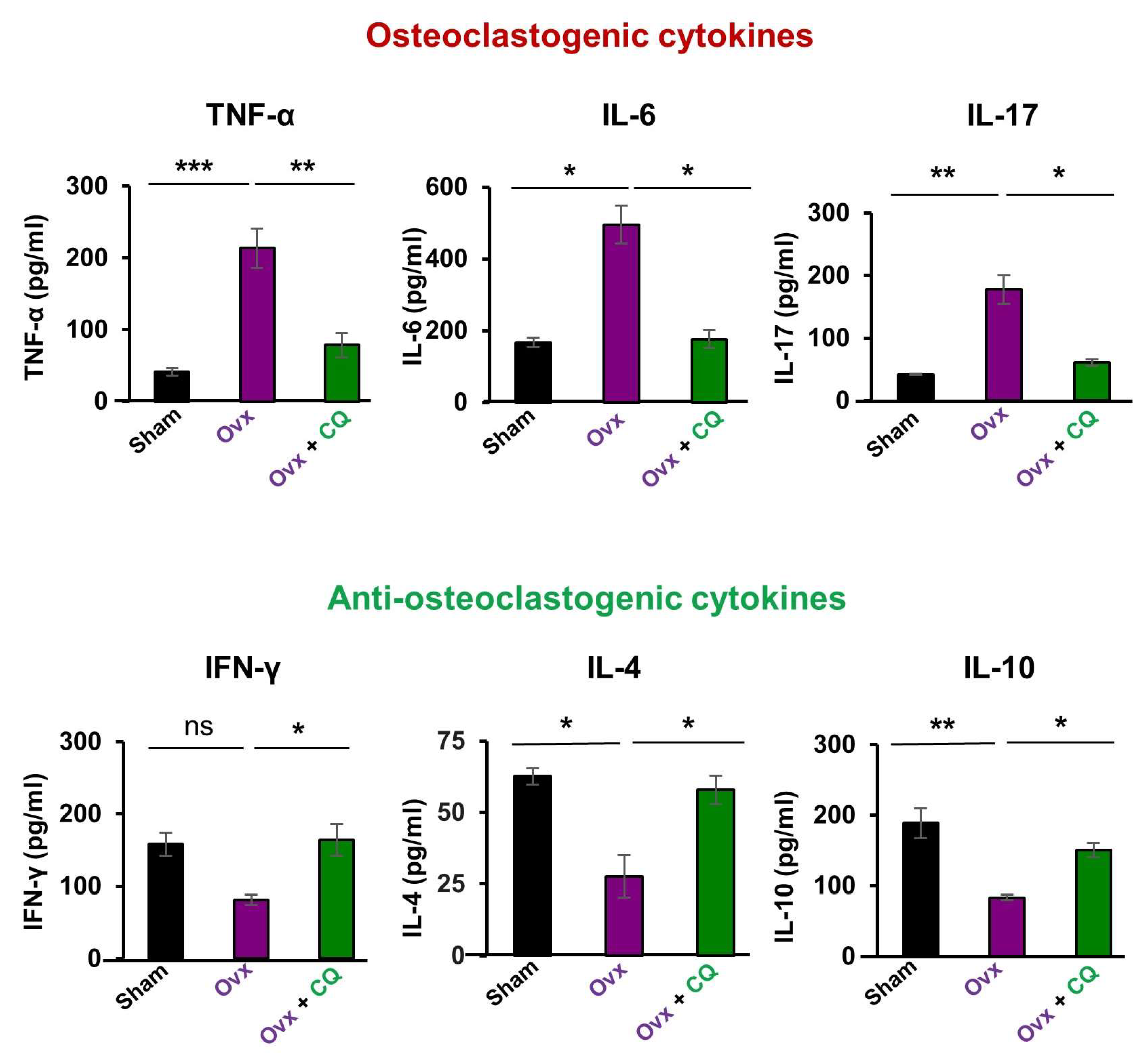
3.7. CQ Skews the Cytokine Balance under Estrogen-Deficient Conditions
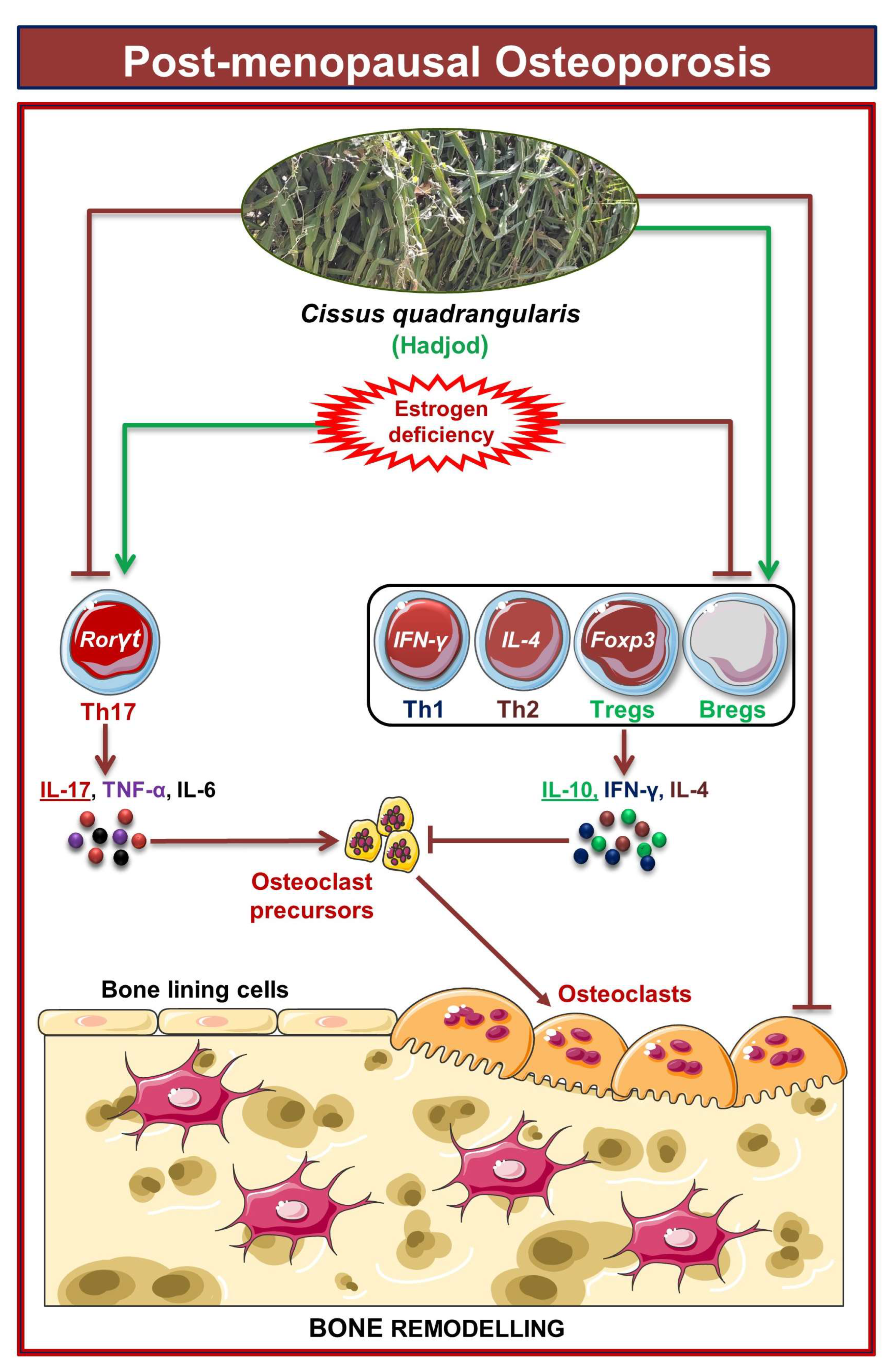
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sözen, T.; Özışık, L.; Başaran, N.Ç. An Overview and Management of Osteoporosis. Eur. J. Rheumatol. 2017, 4, 46. [Google Scholar] [CrossRef] [PubMed]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast Differentiation and Activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Redlich, K.; Smolen, J.S. Inflammatory Bone Loss: Pathogenesis and Therapeutic Intervention. Nat. Rev. Drug Discov. 2012, 11, 234–250. [Google Scholar] [CrossRef] [PubMed]
- McClung, M.; Harris, S.T.; Miller, P.D.; Bauer, D.C.; Davison, K.S.; Dian, L.; Hanley, D.A.; Kendler, D.L.; Yuen, C.K.; Lewiecki, E.M. Bisphosphonate Therapy for Osteoporosis: Benefits, Risks, and Drug Holiday. Am. J. Med. 2013, 126, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Tanvetyanon, T.; Stiff, P. Management of the Adverse Effects Associated with Intravenous Bisphosphonates. Ann. Oncol. 2006, 17, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Bowring, C.; Francis, R. National Osteoporosis Society’s Position Statement on Hormone Replacement Therapy in the Prevention and Treatment of Osteoporosis. Menopause Int. 2011, 17, 63–65. [Google Scholar] [CrossRef]
- Keshishi, D.; Makunts, T.; Abagyan, R. Common Osteoporosis Drug Associated with Increased Rates of Depression and Anxiety. Sci. Rep. 2021, 11, 23956. [Google Scholar] [CrossRef]
- Riancho, J.A.; Zarrabeitia, M.T.; Gonzalezmacias, J. Interleukin-4 Modulates Osteoclast Differentiation and Inhibits the Formation of Resorption Pits in Mouse Osteoclast Cultures. Biochem. Biophys. Res. Commun. 1993, 196, 678–685. [Google Scholar] [CrossRef]
- Miossec, P.; Chomarat, P.; Dechanet, J.; Moreau, J.-F.; Roux, J.-P.; Delmas, P.; Banchereau, J. Interleukin-4 Inhibits Bone Resorption through an Effect on Osteoclasts and Proinflammatory Cytokines in an Ex Vivo Model of Bone Resorption in Rheumatoid Arthritis. Arthritis Rheum. 1994, 37, 1715–1722. [Google Scholar] [CrossRef]
- McInnes, I.B.; Schett, G. Cytokines in the Pathogenesis of Rheumatoid Arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef]
- Pacifici, R. T Cells: Critical Bone Regulators in Health and Disease. Bone 2010, 47, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoreschi, K.; Laurence, A.; Yang, X.-P.; Hirahara, K.; O’Shea, J.J. T Helper 17 Cell Heterogeneity and Pathogenicity in Autoimmune Disease. Trends Immunol. 2011, 32, 395–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raphael, I.; Nalawade, S.; Eagar, T.N.; Forsthuber, T.G. T Cell Subsets and Their Signature Cytokines in Autoimmune and Inflammatory Diseases. Cytokine 2015, 74, 5–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dar, H.Y.; Pal, S.; Shukla, P.; Mishra, P.K.; Tomar, G.B.; Chattopadhyay, N.; Srivastava, R.K. Bacillus Clausii Inhibits Bone Loss by Skewing Treg-Th17 Cell Equilibrium in Postmenopausal Osteoporotic Mice Model. Nutrition 2018, 54, 118–128. [Google Scholar] [CrossRef]
- Dar, H.Y.; Shukla, P.; Mishra, P.K.; Anupam, R.; Mondal, R.K.; Tomar, G.B.; Sharma, V.; Srivastava, R.K. Lactobacillus Acidophilus Inhibits Bone Loss and Increases Bone Heterogeneity in Osteoporotic Mice via Modulating Treg-Th17 Cell Balance. Bone Rep. 2018, 8, 46–56. [Google Scholar] [CrossRef]
- Azam, Z.; Sapra, L.; Bhardwaj, A.; Yadav, S.; Mishra, P.K.; Shukla, P.; Sharma, V.; Srivastava, R.K. Crocin Attenuates Osteoclastogenesis and Enhances Bone Health by Skewing the Immunoporotic “Treg-Th17” Cell Axis in Post-Menopausal Osteoporotic Mice Model. Phytomed. Plus 2022, 2, 100302. [Google Scholar] [CrossRef]
- Dar, H.Y.; Azam, Z.; Anupam, R.; Mondal, R.K.; Srivastava, R.K. Osteoimmunology: The Nexus between Bone and Immune System. Front. Biosci.-Landmark 2018, 23, 464–492. [Google Scholar]
- Dar, H.Y.; Singh, A.; Shukla, P.; Anupam, R.; Mondal, R.K.; Mishra, P.K.; Srivastava, R.K. High Dietary Salt Intake Correlates with Modulated Th17-Treg Cell Balance Resulting in Enhanced Bone Loss and Impaired Bone-Microarchitecture in Male Mice. Sci. Rep. 2018, 8, 2503. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, R.K.; Dar, H.Y.; Mishra, P.K. Immunoporosis: Immunology of Osteoporosis—Role of T Cells. Front. Immunol. 2018, 9, 657. [Google Scholar] [CrossRef]
- Fischer, L.; Herkner, C.; Kitte, R.; Dohnke, S.; Riewaldt, J.; Kretschmer, K.; Garbe, A.I. Foxp3+ Regulatory T Cells in Bone and Hematopoietic Homeostasis. Front. Endocrinol. 2019, 10, 578. [Google Scholar] [CrossRef]
- Sapra, L.; Bhardwaj, A.; Mishra, P.K.; Garg, B.; Verma, B.; Mishra, G.C.; Srivastava, R.K. Regulatory B Cells (Bregs) Inhibit Osteoclastogenesis and Play a Potential Role in Ameliorating Ovariectomy-Induced Bone Loss. Front. Immunol. 2021, 12, 691081. [Google Scholar] [CrossRef] [PubMed]
- Sapra, L.; Shokeen, N.; Gupta, K.; Saini, C.; Bhardwaj, A.; Mathew, M.; Mishra, P.K.; Chattopadhyay, N.; Verma, B.; Srivastava, R.K. Bifidobacterium Longum Attenuates Ovariectomy-Induced Bone Loss Via Modulating the Immunoporotic Breg-Treg-Th17 Cell Axis. Front. Immunol. 2022, 13, 875788. [Google Scholar] [CrossRef] [PubMed]
- Sofowora, A.; Ogunbodede, E.; Onayade, A. The Role and Place of Medicinal Plants in the Strategies for Disease Prevention. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 210–229. [Google Scholar] [CrossRef] [PubMed]
- Udupa, K.; Chaturvedi, G.; Tripathi, S. Advances in Research in Indian Medicine; Banaras Hindu University: Varanasi, India, 1970. [Google Scholar]
- Ganguly, A.; Ganguly, D.; Banerjee, S. Topical Phytotherapeutic Treatment: Management of Normalization of Elevated Levels of Biochemical Parameter During Osteoarthritic Disorders: A Prospective Study. J. Orthop. Rheumatol. 2018, 5, 14. [Google Scholar]
- Sivarajan, V.; Balachandran, I. Ayurvedic Drugs and Their Plant Sources; Oxford and IBH Publishing: New Delhi, India, 1994. [Google Scholar]
- Udupa, K.N.; Prasad, G.C. Further Studies on the Effect of Cissus Quadrangularis in Accelerating Fracture Healing. Indian J. Med. Res. 1964, 52, 26–35. [Google Scholar]
- Shirwaikar, A.; Khan, S.; Malini, S. Antiosteoporotic Effect of Ethanol Extract of Cissus Quadrangularis Linn. On Ovariectomized Rat. J. Ethnopharmacol. 2003, 89, 245–250. [Google Scholar] [CrossRef]
- Stohs, S.J.; Ray, S.D. A Review and Evaluation of the Efficacy and Safety of Cissus Quadrangularis Extracts. Phytother. Res. 2013, 27, 1107–1114. [Google Scholar] [CrossRef]
- Udupa, K.N.; Prasad, G.; Sen, S.P. The Effect of Phytogenic Anabolic Steroid in the Acceleration of Fracture Repair. Life Sci. 1965, 4, 317–327. [Google Scholar] [CrossRef]
- Prasad, G.C.; Udupa, K.N. Pathways and Site of Action of a Phytogenic Steroid from Cissus Quadrangularis. J. Res. Indian Med. 1972, 4, 132. [Google Scholar]
- Padmalochana, K.; Rajan, M.S.D.; Lalitha, R.; Sivasankari, H. Evaluation of the Antioxidant and Hepatoprotective Activity of Cryptolepis Buchanani. J. Appl. Pharm. Sci. 1930, 3, 099–104. [Google Scholar] [CrossRef] [Green Version]
- Muthusami, S.; Ramachandran, I.; Krishnamoorthy, S.; Govindan, R.; Narasimhan, S. Cissus Quadrangularis Augments Igf System Components in Human Osteoblast Like Saos-2 Cells. Growth Horm. IGF Res. 2011, 21, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Sapra, L.; Dar, H.Y.; Bhardwaj, A.; Pandey, A.; Kumari, S.; Azam, Z.; Upmanyu, V.; Anwar, A.; Shukla, P.; Mishra, P.K.; et al. Lactobacillus Rhamnosus Attenuates Bone Loss and Maintains Bone Health by Skewing Treg-Th17 Cell Balance in Ovx Mice. Sci. Rep. 2021, 11, 1807. [Google Scholar] [CrossRef] [PubMed]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, A.M. Standardized Nomenclature, Symbols, and Units for Bone Histomorphometry: A 2012 Update of the Report of the Asbmr Histomorphometry Nomenclature Committee. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2013, 28, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.Y.; Chassaing, B.; Tyagi, A.M.; Vaccaro, C.; Luo, T.; Adams, J.; Darby, T.M.; Weitzmann, M.N.; Mulle, J.G.; Gewirtz, A.T.; et al. Sex Steroid Deficiency–Associated Bone Loss Is Microbiota Dependent and Prevented by Probiotics. J. Clin. Investig. 2016, 126, 2049–2063. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.K.; Tomar, G.B.; Barhanpurkar, A.P.; Gupta, N.; Pote, S.T.; Mishra, G.C.; Wani, M.R. Il-3 Attenuates Collagen-Induced Arthritis by Modulating the Development of Foxp3+ Regulatory T Cells. J. Immunol. 2011, 186, 2262–2272. [Google Scholar] [CrossRef] [Green Version]
- Azam, Z.; Pandey, V.; Gupta, N.; Sapra, L.; Dar, H.Y.; Shokeen, N.; Soni, V.; Srivastava, R.K. Phytoconstituents as Novel Osteo-Protective Agents: Implications in Bone Health. Front. Biosci.-Landmark 2020, 25, 1259–1296. [Google Scholar]
- Mahmoudi, Z.; Soleimani, M.; Khamisipour, G.; Azizsoltani, A. Effects of Foeniculum Vulgare Ethanol Extract on Osteogenesis in Human Mecenchymal Stem Cells. Avicenna J. Phytomed. 2013, 3, 135. [Google Scholar]
- Kanis, J.A.; McCloskey, E.V.; Johansson, H.; Oden, A.; Melton, L.J., III; Khaltaev, N. A Reference Standard for the Description of Osteoporosis. Bone 2008, 42, 467–475. [Google Scholar] [CrossRef]
- Park, J.A.; Ha, S.K.; Kang, T.H.; Oh, M.S.; Cho, M.H.; Lee, S.Y.; Park, J.-H.; Kim, S.Y. Protective Effect of Apigenin on Ovariectomy-Induced Bone Loss in Rats. Life Sci. 2008, 82, 1217–1223. [Google Scholar] [CrossRef]
- Kalalinia, F.; Ghasim, H.; Farzad, S.A.; Pishavar, E.; Ramezani, M.; Hashemi, M. Comparison of the Effect of Crocin and Crocetin, Two Major Compounds Extracted from Saffron, on Osteogenic Differentiation of Mesenchymal Stem Cells. Life Sci. 2018, 208, 262–267. [Google Scholar] [CrossRef]
- Mishra, G.; Srivastava, S.; Nagori, B. Pharmacological and Therapeutic Activity of Cissus Quadrangularis: An Overview. Int. J. Pharmtech Res. 2010, 2, 1298–1310. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bone Parameters | Sham | Ovx | Ovx + CQ |
|---|---|---|---|
| LV-5 | |||
| BV/TV (%) | 68.8 ± 0.1 | 56.4 ± 2.5 | 83.35 ± 1.35 ** |
| Tb. Th (mm) | 0.05 ± 0 | 0.045 ± 0.005 | 0.055 ± 0.005 * |
| Tb. Sp (mm−3) | 0.038 ± 0.002 | 0.058 ± 0.002 | 0.037 ± 0.0015 * |
| Conn. D (mm−3) | 198.38 ± 40.22 | 93.87 ± 27.52 | 223.153 ± 17.29 * |
| Tb. N (mm−1) | 9.8 ± 0.1 | 13.45 ± 0.45 | 16.95 ± 1.75 * |
| Tb. Pf (%) | 15.05 ± 2.65 | 21.25 ± 0.25 | 7.05 ± 0.65 * |
| Femur Trabecular | |||
| BV/TV (%) | 85.3 ± 2.6 | 63.35 ± 1.25 | 89.15 ± 0.75 ** |
| Tb. Th (mm) | 0.05 ± 0 | 0.041 ± 0.0015 | 0.07 ± 0 *** |
| Tb. Sp (mm−3) | 0.2 ± 0 | 0.4 ± 0 | 0.75 ± 0.0015 ** |
| Conn. D (mm−3) | 185.51 ± 14.85 | 83.39 ± 7.75 | 179.04 ± 9.13 *** |
| Tb. N (mm−1) | 7.36 ± 4.78 | 3.56 ± 2.21 | 11.4 ± 7.20 * |
| Tb. Pf (%) | 14.4 ± 0.62 | 19.03 ± 1.26 | 12.53 ± 1.65 ** |
| Tibia Trabecular | |||
| BV/TV (%) | 81.65 ± 2.75 | 62.15 ± 2.05 | 86.35 ± 2.85 * |
| Tb. Th (mm) | 0.07 ± 0 | 0.046 ± 0.005 | 0.00071 ± 0 *** |
| Tb. Sp (mm−3) | 0.037 ± 0.0035 | 0.046 ± 0.0015 | 0.025 ± 0.005 ** |
| Conn. D (mm−3) | 50.33 ± | 27.89 ± | 86.30 ± 53.60 * |
| Tb. N (mm−1) | 9.13 ± 1.73 | 6.7 ± 0.22 | 13.5 ± 1.20 ** |
| Tb. Pf (%) | 13.63 ± 2.35 | 20 ± 1.79 | 12.86 ± 0.96 * |
| Femur Cortical | |||
| Tt. Ar (mm2) | 1.35 ± 0.01 | 0.8 ± 0.1 | 2.03 ± 0.06 ** |
| T. Pm (mm) | 4.38 ± 0.025 | 3.22 ± 0.045 | 5.4 ±0.08 *** |
| Ct. Th (mm) | 0.16 ± 0.005 | 0.13 ± 0.005 | 0.17 ± 0.005 * |
| Ct. Ar (mm2) | 0.59 ± 0.57 | 0.54 ± 0.55 | 0.7 ± 0.08 * |
| B. Pm (mcm) | 7.60 ± 0.03 | 6.57 ± 0.56 | 9.39 ± 0.77 |
| J (mm4) | 0.21 ± 0.02 | 0.13 ± 0.04 | 0.26 ± 0.08 |
| Tibia Cortical | |||
| Tt. Ar (mm2) | 0.975 ± 0.015 | 0.65 ± 0.05 | 1.39 ± 0.05 ** |
| T. Pm (mm) | 4.51 ± 0 | 4.68 ± 0.02 | 5.57 ± 0.115 ** |
| Ct. Th (mm) | 0.14 ± 0.01 | 0.11 ± 0.01 | 0.16 ± 0.01 * |
| Ct. Ar (mm2) | 0.53 ± 0.05 | 0.46 ± 0.02 | 0.66 ± 0.02 ** |
| B. Pm (mcm) | 5.27 ± 3.22 | 4.72 ± 2.90 | 6.77 ± 4.15 * |
| J (mm4) | 0.14 ± 0.02 | 0.13 ± 0.01 | 0.31 ± 0.03 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azam, Z.; Sapra, L.; Baghel, K.; Sinha, N.; Gupta, R.K.; Soni, V.; Saini, C.; Mishra, P.K.; Srivastava, R.K. Cissus quadrangularis (Hadjod) Inhibits RANKL-Induced Osteoclastogenesis and Augments Bone Health in an Estrogen-Deficient Preclinical Model of Osteoporosis Via Modulating the Host Osteoimmune System. Cells 2023, 12, 216. https://doi.org/10.3390/cells12020216
Azam Z, Sapra L, Baghel K, Sinha N, Gupta RK, Soni V, Saini C, Mishra PK, Srivastava RK. Cissus quadrangularis (Hadjod) Inhibits RANKL-Induced Osteoclastogenesis and Augments Bone Health in an Estrogen-Deficient Preclinical Model of Osteoporosis Via Modulating the Host Osteoimmune System. Cells. 2023; 12(2):216. https://doi.org/10.3390/cells12020216
Chicago/Turabian StyleAzam, Zaffar, Leena Sapra, Kalpana Baghel, Niharika Sinha, Rajesh K. Gupta, Vandana Soni, Chaman Saini, Pradyumna K. Mishra, and Rupesh K. Srivastava. 2023. "Cissus quadrangularis (Hadjod) Inhibits RANKL-Induced Osteoclastogenesis and Augments Bone Health in an Estrogen-Deficient Preclinical Model of Osteoporosis Via Modulating the Host Osteoimmune System" Cells 12, no. 2: 216. https://doi.org/10.3390/cells12020216