

Purine Biosynthesis Pathways Are Required for Myogenesis in Xenopus laevis


 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Media
2.2. Yeast Strains and Plasmids
2.3. Yeast Growth Test
2.4. Embryo Culture
2.5. mRNA Synthesis and Morpholino Oligonucleotides
2.6. Microinjections
2.7. In Situ Hybridization
2.8. Immunostaining
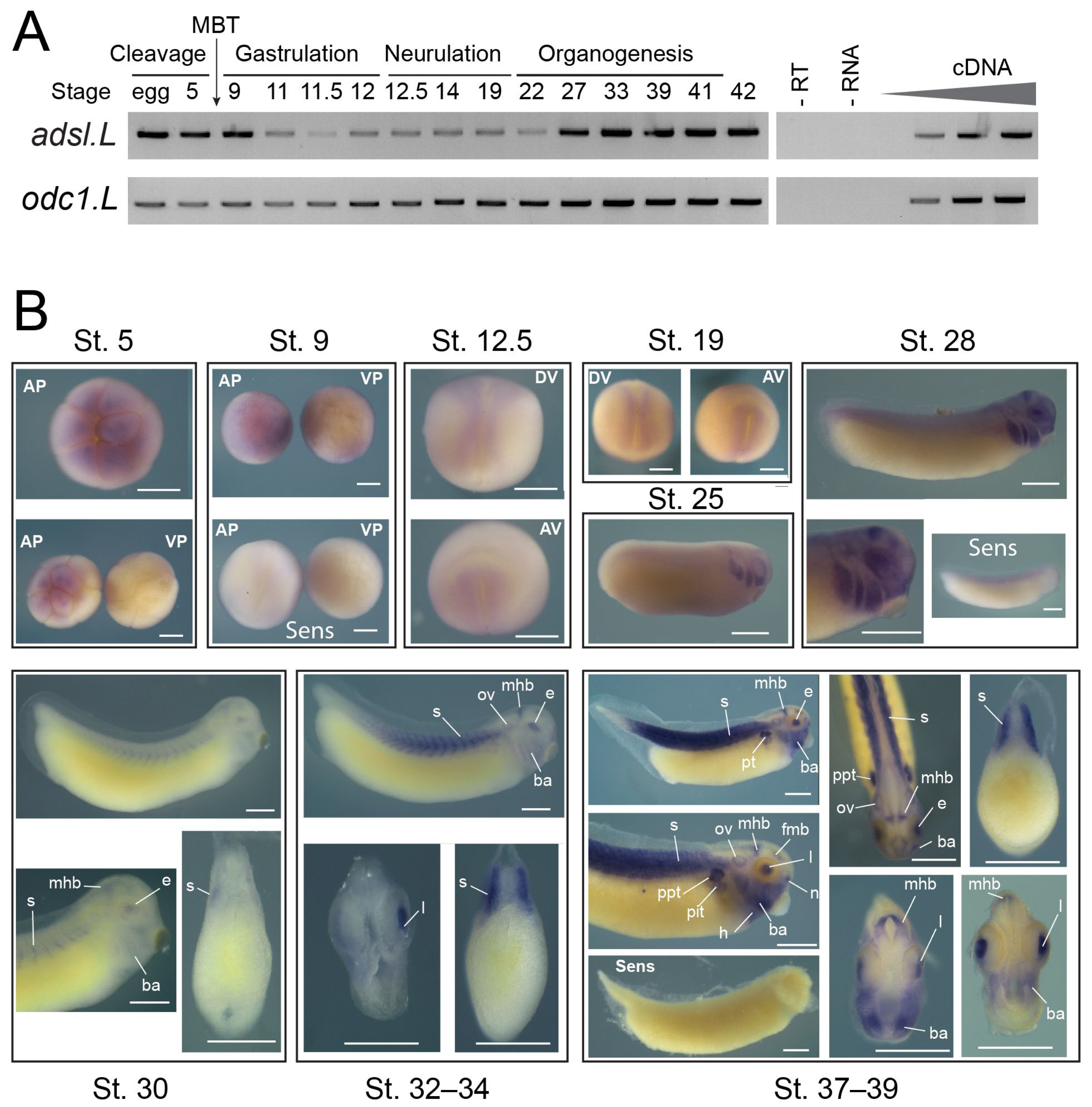
2.9. Temporal Expression of Genes Established Using RT-PCR
2.10. Embryo Scoring and Photography
2.11. Statistics and Reproducibility
2.12. Bioinformatics
3. Results
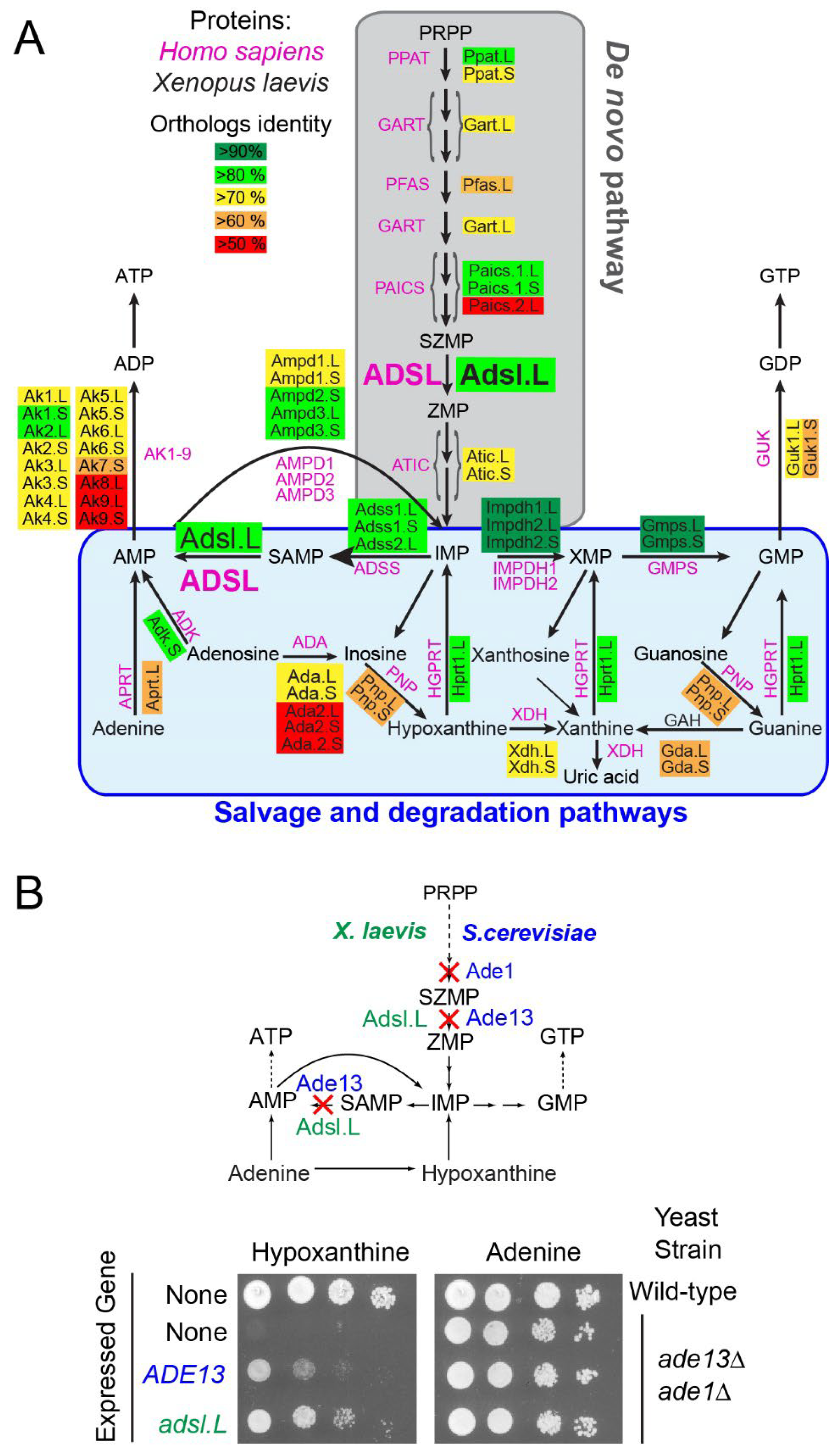
3.1. Identification of X. laevis Purine Pathway Genes
3.2. The adsl.L Gene Was Mostly Expressed in Muscular and Neuronal Tissues and Their Precursors during X. laevis Development
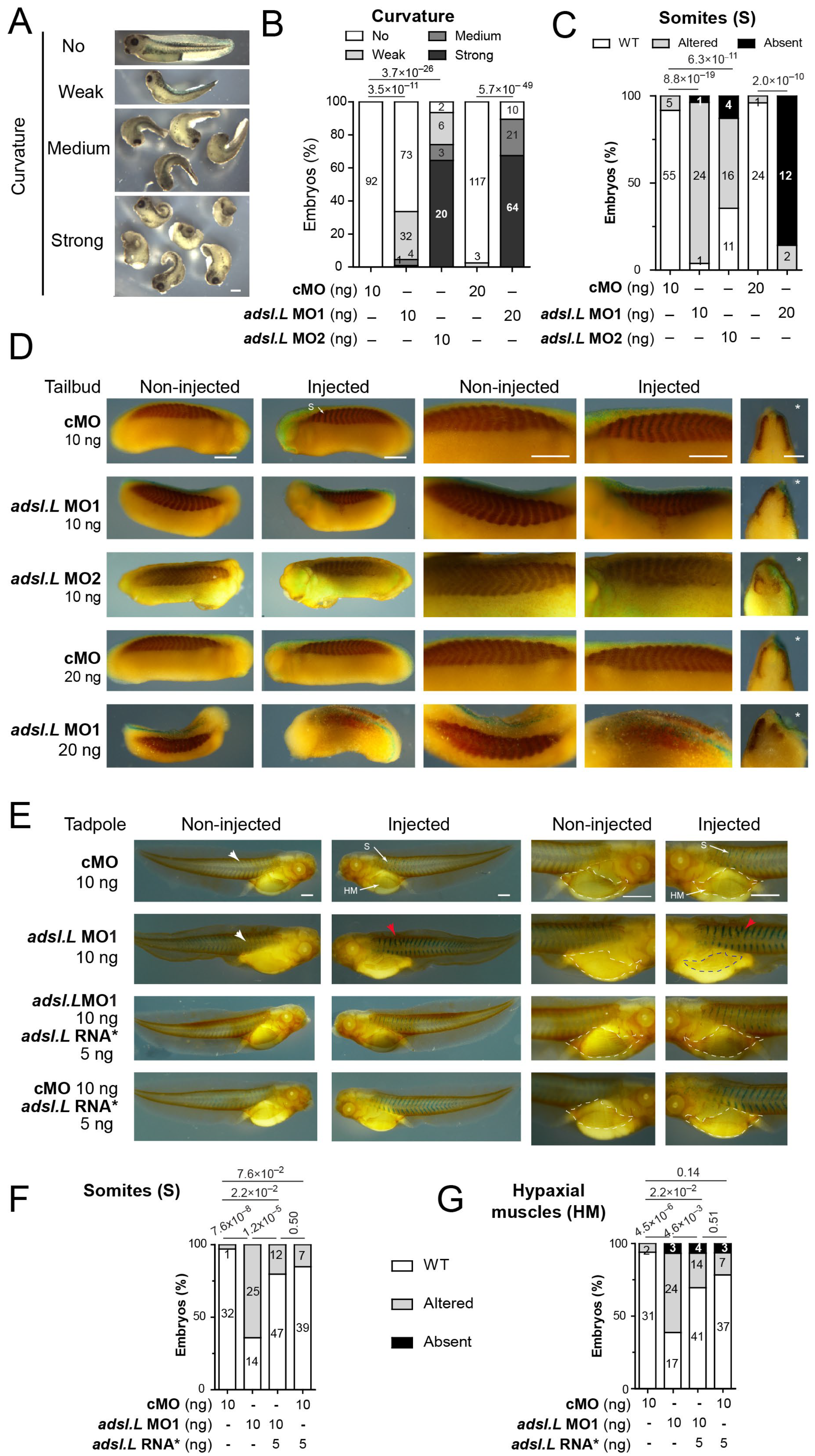
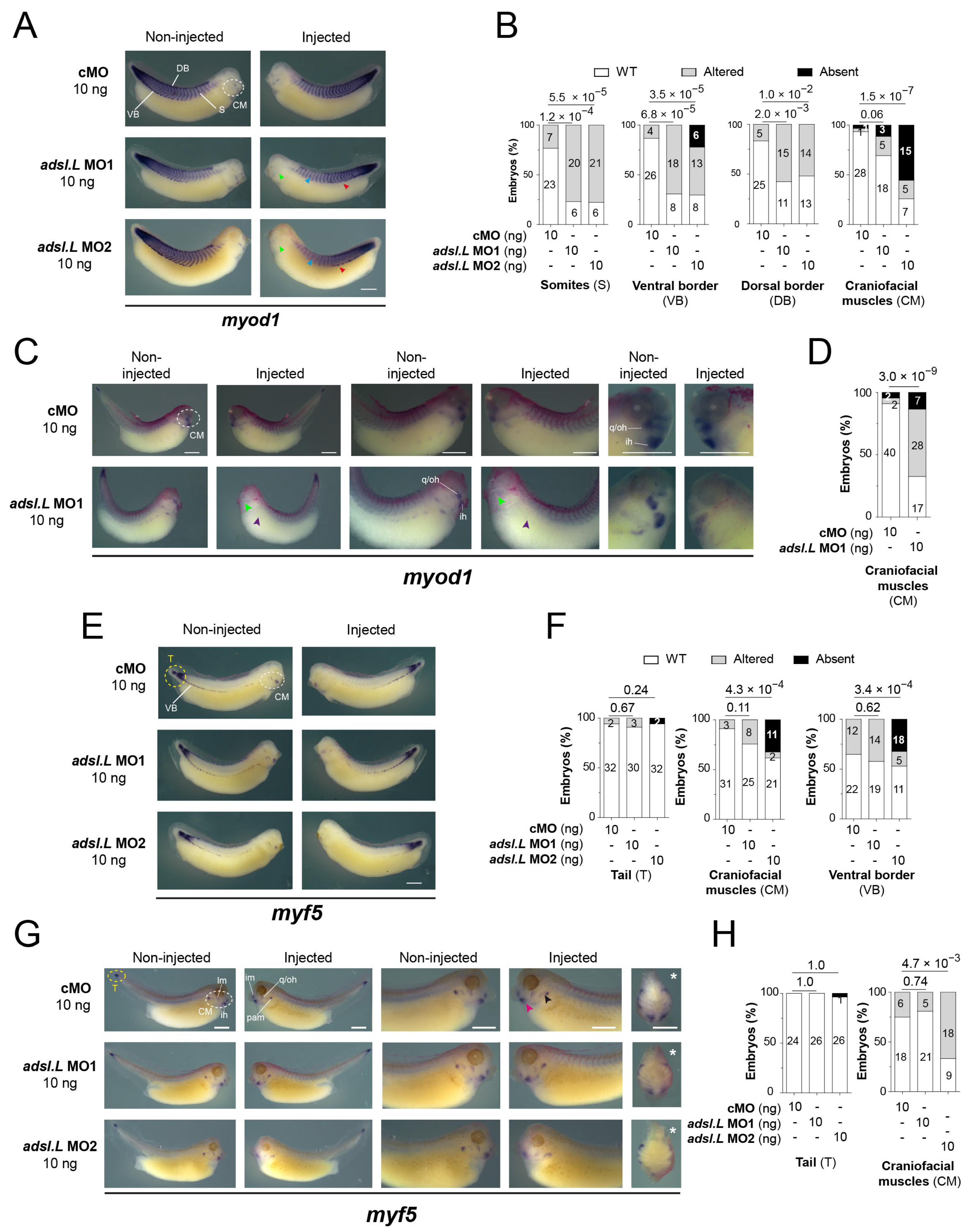
3.3. The adsl.L Gene Was Required for X. laevis Myogenesis
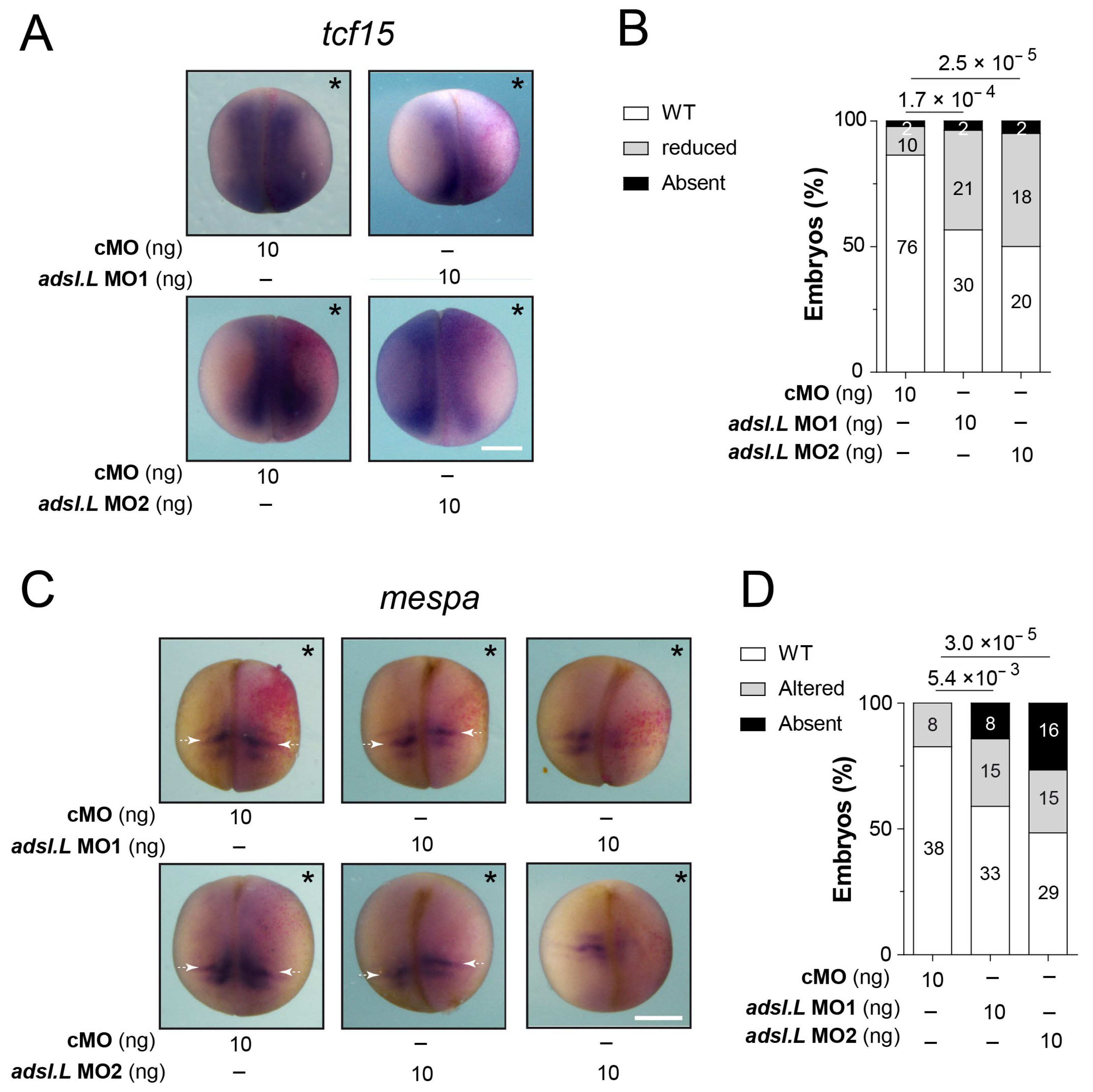
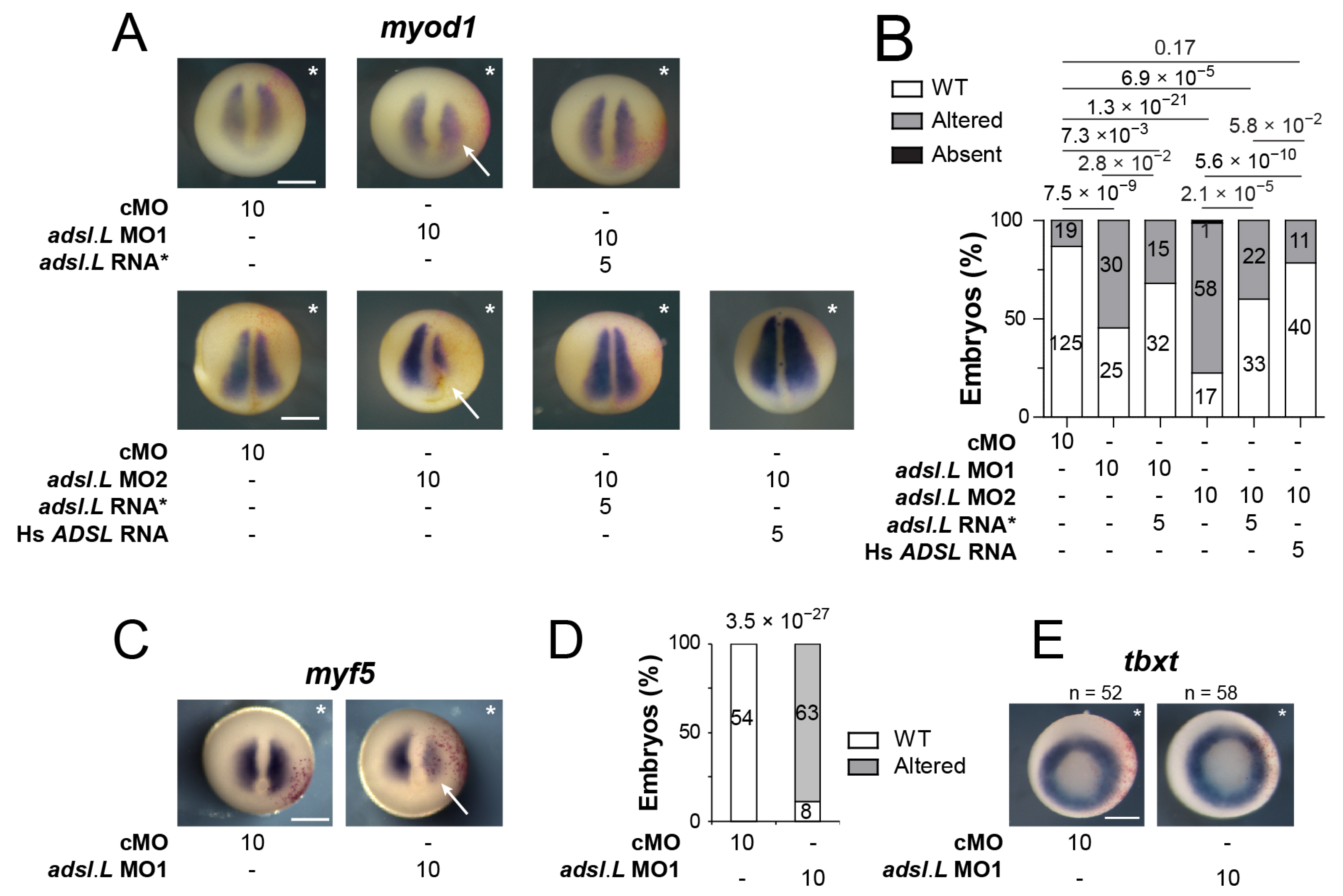
3.4. Expression of the myod1 and myf5 Genes Required Functional Purine Pathways
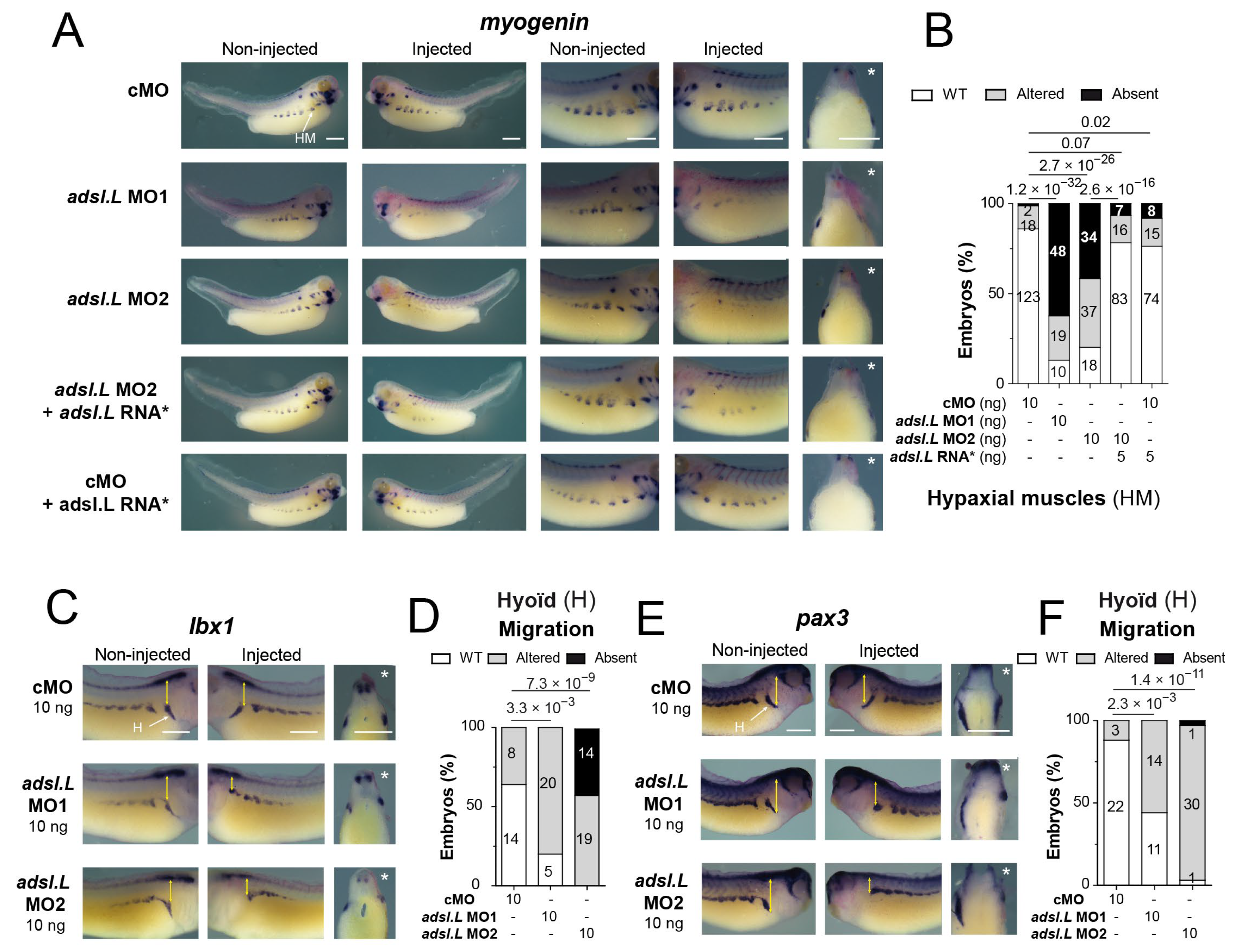
3.5. The adsl.L Gene Was Required for Hypaxial Muscle Formation and Migration
4. Discussion
4.1. X. laevis as a Vertebrate Model to Study Purine-Deficiency-Related Pathologies
4.2. Role of Purine Pathway Genes during X. laevis Development
4.3. Muscle Defects Associated with Purine Pathways Dysfunction Were Related to Altered Expressions of MRF Genes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daignan-Fornier, B.; Pinson, B. Yeast to Study Human Purine Metabolism Diseases. Cells 2019, 8, 67. [Google Scholar] [CrossRef]
- Massé, K.; Dale, N. Purines as Potential Morphogens during Embryonic Development. Purinergic Signal. 2012, 8, 503–521. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, H.A.; Duley, J.A.; Fairbanks, L.D.; McBride, M.B. When to Investigate for Purine and Pyrimidine Disorders. Introduction and Review of Clinical and Laboratory Indications. J. Inherit. Metab. Dis. 1997, 20, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, S.; Duley, J.A.; Christodoulou, J. Inborn Errors of Purine Metabolism: Clinical Update and Therapies. J. Inherit. Metab. Dis. 2014, 37, 669–686. [Google Scholar] [CrossRef] [PubMed]
- Jurecka, A. Inborn Errors of Purine and Pyrimidine Metabolism. J. Inherit. Metab. Dis. 2009, 32, 247–263. [Google Scholar] [CrossRef]
- Nyhan, W.L. Disorders of Purine and Pyrimidine Metabolism. Mol. Genet. Metab. 2005, 86, 25–33. [Google Scholar] [CrossRef]
- van den Berghe, G.; Vincent, M.-F.; Marie, S. Disorders of Purine and Pyrimidine Metabolism. In Inborn Metabolic Diseases: Diagnosis and Treatment; Fernandes, J., Saudubray, J.-M., van den Berghe, G., Walter, J.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 433–449. ISBN 978-3-540-28785-8. [Google Scholar]
- Pinson, B.; Vaur, S.; Sagot, I.; Coulpier, F.; Lemoine, S.; Daignan-Fornier, B. Metabolic Intermediates Selectively Stimulate Transcription Factor Interaction and Modulate Phosphate and Purine Pathways. Genes Dev. 2009, 23, 1399–1407. [Google Scholar] [CrossRef]
- Jaeken, J.; Van den Berghe, G. An Infantile Autistic Syndrome Characterised by the Presence of Succinylpurines in Body Fluids. Lancet 1984, 2, 1058–1061. [Google Scholar]
- Stone, R.L.; Aimi, J.; Barshop, B.A.; Jaeken, J.; Van den Berghe, G.; Zalkin, H.; Dixon, J.E. A Mutation in Adenylosuccinate Lyase Associated with Mental Retardation and Autistic Features. Nat. Genet. 1992, 1, 59–63. [Google Scholar] [CrossRef]
- Mao, X.; Li, K.; Tang, B.; Luo, Y.; Ding, D.; Zhao, Y.; Wang, C.; Zhou, X.; Liu, Z.; Zhang, Y.; et al. Novel Mutations in ADSL for Adenylosuccinate Lyase Deficiency Identified by the Combination of Trio-WES and Constantly Updated Guidelines. Sci. Rep. 2017, 7, 1625. [Google Scholar] [CrossRef]
- Van den Bergh, F.; Vincent, M.F.; Jaeken, J.; Van den Berghe, G. Residual Adenylosuccinase Activities in Fibroblasts of Adenylosuccinase-Deficient Children: Parallel Deficiency with Adenylosuccinate and Succinyl-AICAR in Profoundly Retarded Patients and Non-Parallel Deficiency in a Mildly Retarded Girl. J. Inherit. Metab. Dis. 1993, 16, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Kmoch, S.; Hartmannová, H.; Stibůrková, B.; Krijt, J.; Zikánová, M.; Sebesta, I. Human Adenylosuccinate Lyase (ADSL), Cloning and Characterization of Full-Length cDNA and Its Isoform, Gene Structure and Molecular Basis for ADSL Deficiency in Six Patients. Hum. Mol. Genet. 2000, 9, 1501–1513. [Google Scholar] [CrossRef] [PubMed]
- Mouchegh, K.; Zikánová, M.; Hoffmann, G.F.; Kretzschmar, B.; Kühn, T.; Mildenberger, E.; Stoltenburg-Didinger, G.; Krijt, J.; Dvoráková, L.; Honzík, T.; et al. Lethal Fetal and Early Neonatal Presentation of Adenylosuccinate Lyase Deficiency: Observation of 6 Patients in 4 Families. J. Pediatr. 2007, 150, 57–61.e2. [Google Scholar] [CrossRef] [PubMed]
- Jurecka, A.; Zikanova, M.; Tylki-Szymanska, A.; Krijt, J.; Bogdanska, A.; Gradowska, W.; Mullerova, K.; Sykut-Cegielska, J.; Kmoch, S.; Pronicka, E. Clinical, Biochemical and Molecular Findings in Seven Polish Patients with Adenylosuccinate Lyase Deficiency. Mol. Genet. Metab. 2008, 94, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Zikanova, M.; Skopova, V.; Hnizda, A.; Krijt, J.; Kmoch, S. Biochemical and Structural Analysis of 14 Mutant Adsl Enzyme Complexes and Correlation to Phenotypic Heterogeneity of Adenylosuccinate Lyase Deficiency. Hum. Mutat. 2010, 31, 445–455. [Google Scholar] [CrossRef]
- Spiegel, E.K.; Colman, R.F.; Patterson, D. Adenylosuccinate Lyase Deficiency. Mol. Genet. Metab. 2006, 89, 19–31. [Google Scholar] [CrossRef]
- Gitiaux, C.; Ceballos-Picot, I.; Marie, S.; Valayannopoulos, V.; Rio, M.; Verrieres, S.; Benoist, J.F.; Vincent, M.F.; Desguerre, I.; Bahi-Buisson, N. Misleading Behavioural Phenotype with Adenylosuccinate Lyase Deficiency. Eur. J. Hum. Genet. EJHG 2009, 17, 133–136. [Google Scholar] [CrossRef]
- Jurecka, A.; Zikanova, M.; Kmoch, S.; Tylki-Szymańska, A. Adenylosuccinate Lyase Deficiency. J. Inherit. Metab. Dis. 2015, 38, 231–242. [Google Scholar] [CrossRef]
- Marie, S.; Race, V.; Vincent, M.F.; Van den Berghe, G. Adenylosuccinate Lyase Deficiency: From the Clinics to Molecular Biology. Adv. Exp. Med. Biol. 2000, 486, 79–82. [Google Scholar] [CrossRef]
- Race, V.; Marie, S.; Vincent, M.F.; van den Berghe, G. Clinical, Biochemical and Molecular Genetic Correlations in Adenylosuccinate Lyase Deficiency. Hum. Mol. Genet. 2000, 9, 2159–2165. [Google Scholar] [CrossRef]
- Mastrogiorgio, G.; Macchiaiolo, M.; Buonuomo, P.S.; Bellacchio, E.; Bordi, M.; Vecchio, D.; Brown, K.P.; Watson, N.K.; Contardi, B.; Cecconi, F.; et al. Clinical and Molecular Characterization of Patients with Adenylosuccinate Lyase Deficiency. Orphanet J. Rare Dis. 2021, 16, 112. [Google Scholar] [CrossRef] [PubMed]
- Keller, K.E.; Tan, I.S.; Lee, Y.-S. SAICAR Stimulates Pyruvate Kinase Isoform M2 and Promotes Cancer Cell Survival in Glucose-Limited Conditions. Science 2012, 338, 1069–1072. [Google Scholar] [CrossRef] [PubMed]
- Gooding, J.R.; Jensen, M.V.; Dai, X.; Wenner, B.R.; Lu, D.; Arumugam, R.; Ferdaoussi, M.; MacDonald, P.E.; Newgard, C.B. Adenylosuccinate Is an Insulin Secretagogue Derived from Glucose-Induced Purine Metabolism. Cell Rep. 2015, 13, 157–167. [Google Scholar] [CrossRef]
- Mazzarino, R.C.; Baresova, V.; Zikánová, M.; Duval, N.; Wilkinson, T.G.; Patterson, D.; Vacano, G.N. The CRISPR-Cas9 crADSL HeLa Transcriptome: A First Step in Establishing a Model for ADSL Deficiency and SAICAR Accumulation. Mol. Genet. Metab. Rep. 2019, 21, 100512. [Google Scholar] [CrossRef]
- Dutto, I.; Gerhards, J.; Herrera, A.; Souckova, O.; Škopová, V.; Smak, J.A.; Junza, A.; Yanes, O.; Boeckx, C.; Burkhalter, M.D.; et al. Pathway-Specific Effects of ADSL Deficiency on Neurodevelopment. eLife 2022, 11, e70518. [Google Scholar] [CrossRef]
- Marsac, R.; Pinson, B.; Saint-Marc, C.; Olmedo, M.; Artal-Sanz, M.; Daignan-Fornier, B.; Gomes, J.-E. Purine Homeostasis Is Necessary for Developmental Timing, Germline Maintenance and Muscle Integrity in Caenorhabditis elegans. Genetics 2019, 211, 1297–1313. [Google Scholar] [CrossRef]
- Sater, A.K.; Moody, S.A. Using Xenopus to Understand Human Disease and Developmental Disorders. Genesis 2017, 55. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; Ott, T. Xenopus: An Undervalued Model Organism to Study and Model Human Genetic Disease. Cells Tissues Organs 2018, 205, 303–313. [Google Scholar] [CrossRef]
- Garí, E.; Piedrafita, L.; Aldea, M.; Herrero, E. A Set of Vectors with a Tetracycline-Regulatable Promoter System for Modulated Gene Expression in Saccharomyces cerevisiae. Yeast Chichester Engl. 1997, 13, 837–848. [Google Scholar] [CrossRef]
- Nieuwkoop, P.D.; Faber, J. Normal Table of Xenopus Laevis (Daudin): A Systematical and Chronological Survey of the Development from the Fertilized Egg Till the End of Metamorphosis; Garland Pub.: Spokane, WA, USA, 1994; ISBN 978-0-8153-1896-5. [Google Scholar]
- Massé, K.; Bhamra, S.; Eason, R.; Dale, N.; Jones, E.A. Purine-Mediated Signalling Triggers Eye Development. Nature 2007, 449, 1058–1062. [Google Scholar] [CrossRef]
- Blanchard, C.; Boué-Grabot, E.; Massé, K. Comparative Embryonic Spatio-Temporal Expression Profile Map of the Xenopus P2X Receptor Family. Front. Cell. Neurosci. 2019, 13, 340. [Google Scholar] [CrossRef]
- Massé, K.; Eason, R.; Bhamra, S.; Dale, N.; Jones, E.A. Comparative Genomic and Expression Analysis of the Conserved NTPDase Gene Family in Xenopus. Genomics 2006, 87, 366–381. [Google Scholar] [CrossRef]
- Della Gaspera, B.; Armand, A.-S.; Lecolle, S.; Charbonnier, F.; Chanoine, C. Mef2d Acts Upstream of Muscle Identity Genes and Couples Lateral Myogenesis to Dermomyotome Formation in Xenopus laevis. PLoS ONE 2012, 7, e52359. [Google Scholar] [CrossRef] [PubMed]
- Della Gaspera, B.; Armand, A.-S.; Sequeira, I.; Chesneau, A.; Mazabraud, A.; Lécolle, S.; Charbonnier, F.; Chanoine, C. Myogenic Waves and Myogenic Programs during Xenopus Embryonic Myogenesis. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2012, 241, 995–1007. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.L.; Harland, R.M. A Novel Role for Lbx1 in Xenopus Hypaxial Myogenesis. Dev. Camb. Engl. 2006, 133, 195–208. [Google Scholar] [CrossRef]
- Kintner, C.R.; Brockes, J.P. Monoclonal Antibodies Identify Blastemal Cells Derived from Dedifferentiating Limb Regeneration. Nature 1984, 308, 67–69. [Google Scholar] [CrossRef]
- Massé, K.; Bhamra, S.; Allsop, G.; Dale, N.; Jones, E.A. Ectophosphodiesterase/Nucleotide Phosphohydrolase (Enpp) Nucleotidases: Cloning, Conservation and Developmental Restriction. Int. J. Dev. Biol. 2010, 54, 181–193. [Google Scholar] [CrossRef]
- Bassez, T.; Paris, J.; Omilli, F.; Dorel, C.; Osborne, H.B. Post-Transcriptional Regulation of Ornithine Decarboxylase in Xenopus Laevis Oocytes. Dev. Camb. Engl. 1990, 110, 955–962. [Google Scholar] [CrossRef]
- Bowes, J.B.; Snyder, K.A.; Segerdell, E.; Gibb, R.; Jarabek, C.; Noumen, E.; Pollet, N.; Vize, P.D. Xenbase: A Xenopus Biology and Genomics Resource. Nucleic Acids Res. 2008, 36, D761–D767. [Google Scholar] [CrossRef]
- Karpinka, J.B.; Fortriede, J.D.; Burns, K.A.; James-Zorn, C.; Ponferrada, V.G.; Lee, J.; Karimi, K.; Zorn, A.M.; Vize, P.D. Xenbase, the Xenopus Model Organism Database; New Virtualized System, Data Types and Genomes. Nucleic Acids Res. 2015, 43, D756–D763. [Google Scholar] [CrossRef]
- Session, A.M.; Uno, Y.; Kwon, T.; Chapman, J.A.; Toyoda, A.; Takahashi, S.; Fukui, A.; Hikosaka, A.; Suzuki, A.; Kondo, M.; et al. Genome Evolution in the Allotetraploid Frog Xenopus laevis. Nature 2016, 538, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Ljungdahl, P.O.; Daignan-Fornier, B. Regulation of Amino Acid, Nucleotide, and Phosphate Metabolism in Saccharomyces cerevisiae. Genetics 2012, 190, 885–929. [Google Scholar] [CrossRef] [PubMed]
- Kragtorp, K.A.; Miller, J.R. Integrin Alpha5 Is Required for Somite Rotation and Boundary Formation in Xenopus. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2007, 236, 2713–2720. [Google Scholar] [CrossRef]
- Moreno, T.A.; Kintner, C. Regulation of Segmental Patterning by Retinoic Acid Signaling during Xenopus Somitogenesis. Dev. Cell 2004, 6, 205–218. [Google Scholar] [CrossRef]
- Sparrow, D.B.; Jen, W.C.; Kotecha, S.; Towers, N.; Kintner, C.; Mohun, T.J. Thylacine 1 Is Expressed Segmentally within the Paraxial Mesoderm of the Xenopus Embryo and Interacts with the Notch Pathway. Dev. Camb. Engl. 1998, 125, 2041–2051. [Google Scholar] [CrossRef]
- Tocco, A.; Pinson, B.; Thiébaud, P.; Thézé, N.; Massé, K. Comparative Genomic and Expression Analysis of the Adenosine Signaling Pathway Members in Xenopus. Purinergic Signal. 2015, 11, 59–77. [Google Scholar] [CrossRef]
- Della Gaspera, B.; Weill, L.; Chanoine, C. Evolution of Somite Compartmentalization: A View From Xenopus. Front. Cell Dev. Biol. 2021, 9, 790847. [Google Scholar] [CrossRef]
- Wilson-Rawls, J.; Hurt, C.R.; Parsons, S.M.; Rawls, A. Differential Regulation of Epaxial and Hypaxial Muscle Development by Paraxis. Dev. Camb. Engl. 1999, 126, 5217–5229. [Google Scholar] [CrossRef]
- Martin, B.L.; Harland, R.M. Hypaxial Muscle Migration during Primary Myogenesis in Xenopus laevis. Dev. Biol. 2001, 239, 270–280. [Google Scholar] [CrossRef]
- Kelley, R.E.; Andersson, H.C. Disorders of Purines and Pyrimidines. Handb. Clin. Neurol. 2014, 120, 827–838. [Google Scholar] [CrossRef]
- Iijima, R.; Kunieda, T.; Yamaguchi, S.; Kamigaki, H.; Fujii-Taira, I.; Sekimizu, K.; Kubo, T.; Natori, S.; Homma, K.J. The Extracellular Adenosine Deaminase Growth Factor, ADGF/CECR1, Plays a Role in Xenopus Embryogenesis via the Adenosine/P1 Receptor. J. Biol. Chem. 2008, 283, 2255–2264. [Google Scholar] [CrossRef] [PubMed]
- Maguire, R.J.; Isaacs, H.V.; Pownall, M.E. Early Transcriptional Targets of MyoD Link Myogenesis and Somitogenesis. Dev. Biol. 2012, 371, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.B.; Nelson, J.W.; Breaker, R.R. An Ancient Riboswitch Class in Bacteria Regulates Purine Biosynthesis and One-Carbon Metabolism. Mol. Cell 2015, 57, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Nakagawara, K.; Takeuchi, C.; Ishige, K. 5′-CMP and 5′-UMP Promote Myogenic Differentiation and Mitochondrial Biogenesis by Activating Myogenin and PGC-1α in a Mouse Myoblast C2C12 Cell Line. Biochem. Biophys. Rep. 2022, 31, 101309. [Google Scholar] [CrossRef] [PubMed]
- Sabillo, A.; Ramirez, J.; Domingo, C.R. Making Muscle: Morphogenetic Movements and Molecular Mechanisms of Myogenesis in Xenopus laevis. Semin. Cell Dev. Biol. 2016, 51, 80–91. [Google Scholar] [CrossRef]
- Knight, J.D.; Kothary, R. The Myogenic Kinome: Protein Kinases Critical to Mammalian Skeletal Myogenesis. Skelet. Muscle 2011, 1, 29. [Google Scholar] [CrossRef]
- Burnstock, G.; Arnett, T.R.; Orriss, I.R. Purinergic Signalling in the Musculoskeletal System. Purinergic Signal. 2013, 9, 541–572. [Google Scholar] [CrossRef]
- Pietrangelo, T.; Guarnieri, S.; Fulle, S.; Fanò, G.; Mariggiò, M.A. Signal Transduction Events Induced by Extracellular Guanosine 5′ Triphosphate in Excitable Cells. Purinergic Signal. 2006, 2, 633–636. [Google Scholar] [CrossRef]
- Meyer, M.P.; Gröschel-Stewart, U.; Robson, T.; Burnstock, G. Expression of Two ATP-Gated Ion Channels, P2X5 and P2X6, in Developing Chick Skeletal Muscle. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1999, 216, 442–449. [Google Scholar] [CrossRef]
- Ryten, M.; Dunn, P.M.; Neary, J.T.; Burnstock, G. ATP Regulates the Differentiation of Mammalian Skeletal Muscle by Activation of a P2X5 Receptor on Satellite Cells. J. Cell Biol. 2002, 158, 345–355. [Google Scholar] [CrossRef]
- Maire, P.; Dos Santos, M.; Madani, R.; Sakakibara, I.; Viaut, C.; Wurmser, M. Myogenesis Control by SIX Transcriptional Complexes. Semin. Cell Dev. Biol. 2020, 104, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Janesick, A.; Tang, W.; Nguyen, T.T.L.; Blumberg, B. RARβ2 Is Required for Vertebrate Somitogenesis. Dev. Camb. Engl. 2017, 144, 1997–2008. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Q. Implication of Retinoic Acid Receptor Selective Signaling in Myogenic Differentiation. Sci. Rep. 2016, 6, 18856. [Google Scholar] [CrossRef] [PubMed]
- Baló-Banga, J.M.; Leibinger, J.; Molnár, L.; Király, K. The Effect of Retinoic Acid on the Synthesis of Phosphoribosylpyrophosphate in Human Erythrocytes in Psoriasis. A Preliminary Note. Dermatologica 1978, 157 (Suppl. S1), 45–51. [Google Scholar] [CrossRef] [PubMed]
- Paschaki, M.; Schneider, C.; Rhinn, M.; Thibault-Carpentier, C.; Dembélé, D.; Niederreither, K.; Dollé, P. Transcriptomic Analysis of Murine Embryos Lacking Endogenous Retinoic Acid Signaling. PLoS ONE 2013, 8, e62274. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duperray, M.; Hardet, F.; Henriet, E.; Saint-Marc, C.; Boué-Grabot, E.; Daignan-Fornier, B.; Massé, K.; Pinson, B. Purine Biosynthesis Pathways Are Required for Myogenesis in Xenopus laevis. Cells 2023, 12, 2379. https://doi.org/10.3390/cells12192379
Duperray M, Hardet F, Henriet E, Saint-Marc C, Boué-Grabot E, Daignan-Fornier B, Massé K, Pinson B. Purine Biosynthesis Pathways Are Required for Myogenesis in Xenopus laevis. Cells. 2023; 12(19):2379. https://doi.org/10.3390/cells12192379
Chicago/Turabian StyleDuperray, Maëlle, Fanny Hardet, Elodie Henriet, Christelle Saint-Marc, Eric Boué-Grabot, Bertrand Daignan-Fornier, Karine Massé, and Benoît Pinson. 2023. "Purine Biosynthesis Pathways Are Required for Myogenesis in Xenopus laevis" Cells 12, no. 19: 2379. https://doi.org/10.3390/cells12192379